Molecular Mechanisms of Algicidal Bacteria in Controlling Harmful Algal Blooms: Advances in Bacteria‐Algae Interactions
Jiaxin Wang, Binfu Xu, Lixing Huang

TL;DR
This review explains how certain bacteria kill harmful algae by disrupting their structure and function, offering a potential solution for controlling algal blooms.
Contribution
The paper systematically summarizes the molecular mechanisms and bacterial behaviors involved in algicidal activity, emphasizing extracellular vesicles and calcium signaling.
Findings
Bacterial behaviors like chemotaxis, adhesion, and quorum sensing are critical in the algicidal process.
Extracellular vesicles released by bacteria play a significant role in killing algal cells.
Calcium signaling regulation is increasingly recognized as important in algal cell death.
Abstract
The frequent occurrence of harmful algal blooms (HAB) poses severe threats to aquatic ecosystems, aquaculture industries and human health. Recently, algicidal bacteria have emerged as a promising biocontrol strategy. However, the precise mechanisms underlying their algicidal effects remain poorly understood, limiting their practical application in environmental management. This review systematically summarises the interactions between bacteria and algae, as well as the various algicidal modes employed by bacteria, with a particular focus on the mechanisms driving bacterial algicidal activity. Key bacterial behaviours such as chemotaxis, adhesion, quorum sensing and the release of extracellular vesicles have been identified as critical factors in the algicidal process, among which the role of bacterial extracellular vesicles warrants special attention. Furthermore, we elaborate on the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
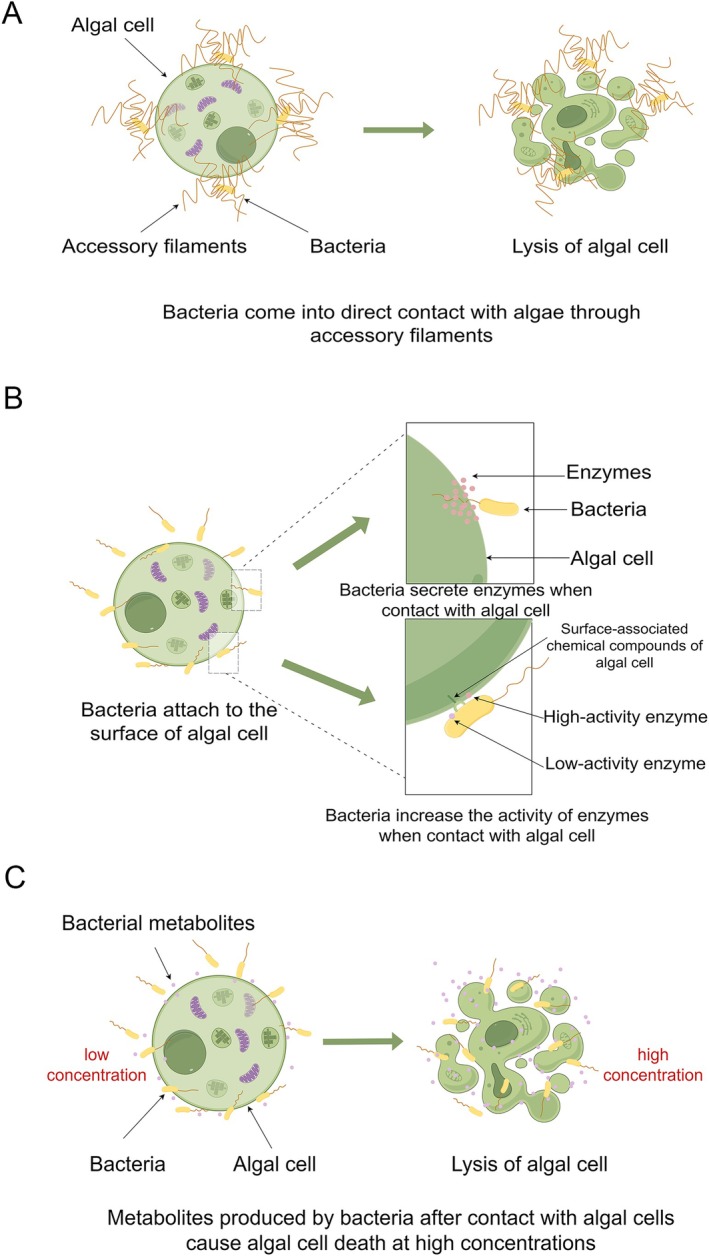 FIGURE 1
FIGURE 1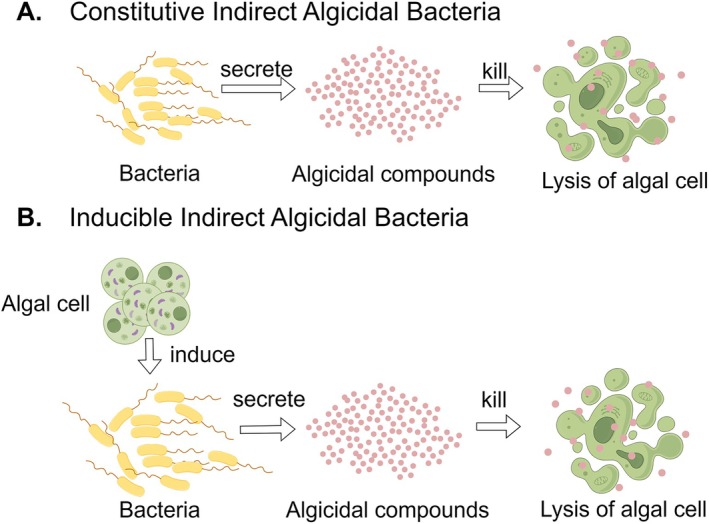 FIGURE 2
FIGURE 2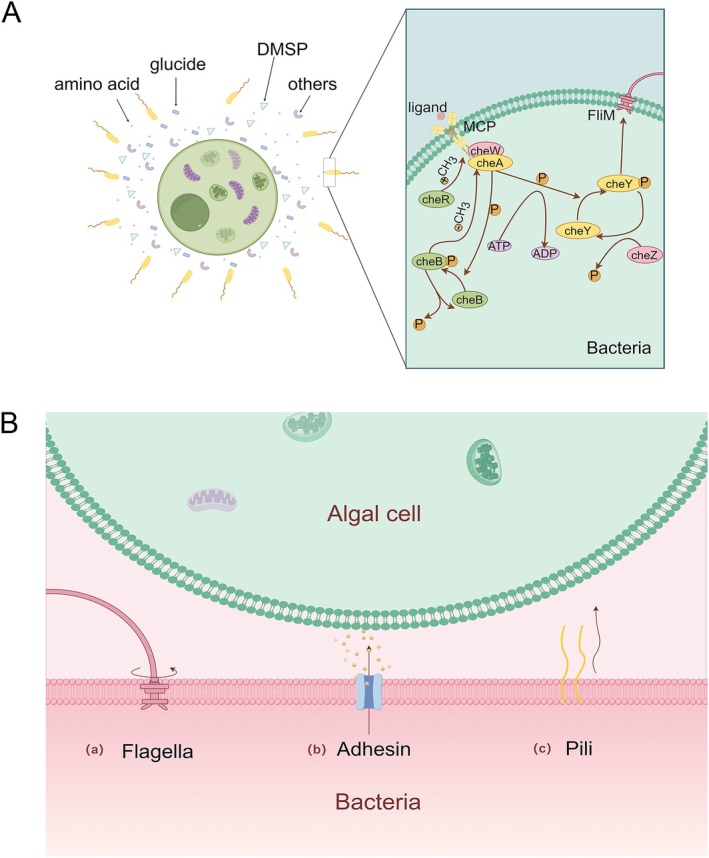 FIGURE 3
FIGURE 3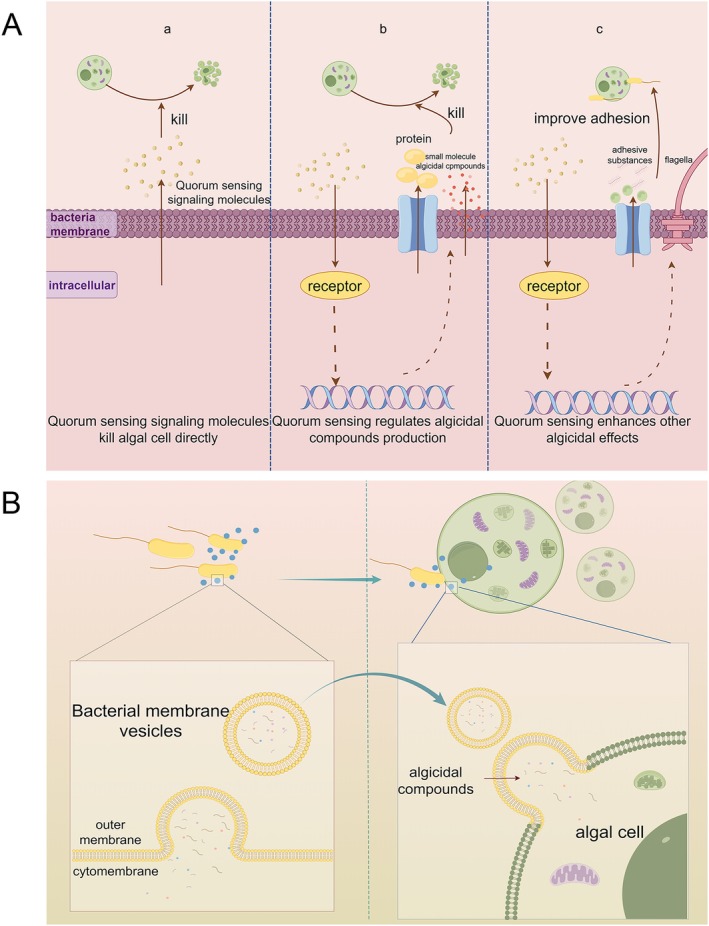 FIGURE 4
FIGURE 4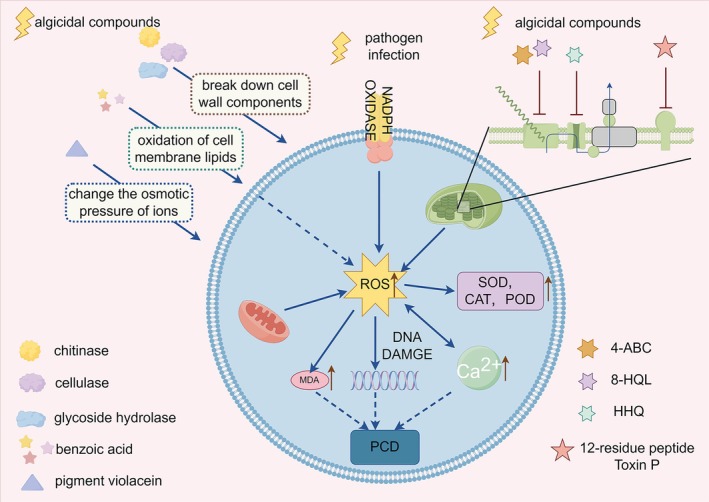 FIGURE 5
FIGURE 5- —Natural Science Foundation of Fujian Province10.13039/501100003392
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Community Ecology and Physiology · Vibrio bacteria research studies · Protist diversity and phylogeny
Introduction
1
Harmful algal blooms (HABs) refer to phytoplankton outbreaks in aquatic environments, such as marine and freshwater ecosystems, that occur under specific conditions, posing significant threats to the ecological balance of the water. The primary species involved in these blooms include various dinoflagellates (Yu et al. 2023), cyanobacteria (Paerl et al. 2001), diatoms (Smayda 1997), etc. These harmful algae are widely distributed and have been reported across global marine regions, including China, Japan, South Korea, eastern Russia (Sakamoto et al. 2021), the coasts of Northern Europe (Karlson et al. 2021), the United States (Anderson et al. 2021) and the Mediterranean Sea (Zingone et al. 2021). HABs pose significant threats to global ecosystems, aquaculture industries and human health. The extensive proliferation of harmful algae can lead to the depletion of oxygen and other essential nutrients in aquatic environments, jeopardising the survival of other species and disrupting ecological balance (Li et al. 2017). These blooms also cause irreparable economic damage. For instance, in 2012, a bloom of Karenia mikimotoi along the Chinese coast resulted in mass mortality of marine organisms, particularly cultured abalone, leading to economic losses of up to $330 million (Li et al. 2017). Similar events in Japan, South Korea and other countries have incurred losses amounting to billions of dollars (Sakamoto et al. 2021). Beyond the impacts on aquaculture, the toxins produced by these harmful algae accumulate in filter‐feeding shellfish, and consumption of these contaminated organisms by humans can lead to poisoning. In addition to impacting fisheries, toxins produced by these harmful algae can accumulate in marine organisms such as shellfish through feeding. When humans consume contaminated seafood, poisoning may occur, leading to public health concerns. These toxins are commonly categorised into five groups based on their clinical symptoms: ciguatera fish poisoning (CFP), paralytic shellfish poisoning (PSP), neurotoxic shellfish poisoning (NSP), amnesic shellfish poisoning (ASP) and diarrhetic shellfish poisoning (DSP) (Grattan et al. 2016). This is sorted out in a review published by Grattan et al. (2016). For instance, ingestion of fish containing CFP toxins can lead to acute gastrointestinal symptoms within 24 h, potentially accompanied by cardiovascular or neurological complications. The other four toxin types primarily accumulate in shellfish. PSP typically causes oral numbness or tingling shortly after consumption, which may progress to facial numbness, along with headache, abdominal pain, nausea, vomiting, dizziness and paresthesia. NSP induces acute gastroenteritis and neurological symptoms such as paresthesia and vertigo, although most patients recover within a short period. ASP presents with acute symptoms including vomiting, abdominal cramps, diarrhoea and headache, and may also involve memory loss. DSP is mainly characterised by severe diarrhoea accompanied by nausea, vomiting and abdominal cramps (Grattan et al. 2016). Beyond direct consumption of contaminated seafood, toxins from HABs can also become airborne and affect humans via aerosol transmission (Lim et al. 2023). Furthermore, the accumulation of toxins in water bodies compromises the quality of water used for human consumption (Brooks et al. 2016; Qin et al. 2010). Despite increased awareness of environmental protection in recent years, these risks remain poorly controlled. Survey data indicate that the frequency of coastal red tide events in the Northern South China Sea (NSCS) increased from 76 occurrences in 1998–2007 to 90 occurrences in 2008–2018 (Liu et al. 2025). Globally, HABs have increased in frequency since the 1980s (Feng et al. 2024), and projections indicate that the number of days with cyanobacterial blooms in large reservoirs and lakes in the United States could rise from 7 days in 2017 to between 18 and 39 days by 2090 (Chapra et al. 2017). Although improvements in the monitoring and reporting systems for HABs may introduce a positive bias in trend estimates, climate warming (Gobler et al. 2017) and nutrient inputs (Wang et al. 2023; Lan et al. 2024) are still likely to increase the frequency of HAB events. Rising global atmospheric CO_2_ levels and elevated temperatures lead to ocean warming, acidification and deoxygenation (Gobler 2020). Under these changing conditions, certain harmful algal species thrive: cyanobacteria, due to their adaptation to warmer waters, often dominate in freshwater systems (Paerl and Huisman 2008), while CO_2_‐limited harmful dinoflagellates may benefit from elevated CO_2_ concentrations and thus gain a competitive advantage (Reinfelder 2011). Furthermore, there is an observed trend of HAB expansion towards polar regions as sea temperatures rise (Griffith et al. 2019). Nutrient enrichment from agricultural, industrial and urban wastewater discharges promotes eutrophication, enabling rapid algal growth driven by nitrogen and phosphorus availability. This is especially evident in freshwater systems close to urban areas, where cyanobacterial blooms frequently occur (Paerl et al. 2001). When these nutrient‐laden freshwaters flow into coastal zones, they can induce sustained eutrophication in marine environments, potentially leading to more severe ecological and economic impacts (Wurtsbaugh et al. 2019). Beyond these primary factors, hydrodynamic conditions such as poor water circulation, along with extreme weather events, including coastal typhoons, heavy rainfall, droughts or prolonged sunny periods, can also trigger algal bloom formation (Feng et al. 2024; Aoki et al. 2019). In summary, the occurrence of algal blooms results from a complex interplay of environmental and anthropogenic factors.
Current strategies to control HABs are largely physical, including ultrasound treatment (Kong et al. 2022), photocatalysts (Wei et al. 2024), and UV‐C irradiation (Gallardo‐Rodríguez et al. 2019). Chemical approaches rely on materials such as clays to promote flocculation and sedimentation of blooms (Jiang et al. 2021), while other chemical methods employ plant extracts (Ni et al. 2023) and aquatic environmental remediation agents (Chai et al. 2025; Zhou et al. 2014) to treat affected waters. Biocontrol options are also being explored, including the use of algicidal bacteria (Yu et al. 2024), algal parasites (Li, Song, et al. 2014), algicidal viruses (Takano et al. 2018) and protists (Li, Gu, et al. 2024). However, physical and chemical methods exhibit several drawbacks: slower action, variable efficacy, higher costs, limited scope, greater energy demands, potential environmental pollution and limited specificity (Balaji‐Prasath 2022; Anabtawi et al. 2024). In contrast, biocontrol approaches, particularly those employing algicidal bacteria, offer environmental compatibility and favourable cost‐effectiveness and are regarded as a promising avenue for HAB mitigation (Anabtawi et al. 2024). Currently, algicidal bacteria can be immobilised for the removal of algal blooms in natural environments. For example, one study immobilised the algicidal bacterium Shewanella sp. IRI‐160 on a porous substrate, which preserved its algicidal activity while allowing for recovery after use, thereby minimising environmental impact (Wang and Coyne 2020). Although progress has been made in using algicidal bacteria for algal bloom control, the complexity of natural ecosystems often leads to discrepancies between laboratory results and field applications (Noh et al. 2017). Addressing these challenges will be crucial for advancing real‐world implementation in future applied research.
The relationship between bacteria and microalgae has long been a focal point in environmental research (Azam and Malfatti 2007; Buchan et al. 2014; Hu et al. 2025). As primary producers in marine ecosystems, microalgae utilise photosynthesis to fix inorganic carbon (Field et al. 1998) and release substantial amounts of organic matter into their surroundings. These extracellular metabolites not only attract bacteria to colonise the phycosphere, the zone immediately surrounding algal cells, but also shape bacterial community composition (Patidar 2025). Bacteria and microalgae engage in intricate interactions that significantly influence algal metabolism and growth dynamics. Certain bacteria establish interdependent relationships with algal cells, some of which can reside within the cell envelope or periplasmic space (Coale et al. 2024). Others communicate via chemical signalling or exchange of metabolites (Amin et al. 2015). Additionally, bacteria can release bioactive compounds that inhibit algal growth (Li, Zhu, et al. 2014) or even induce algal cell lysis (Zeng et al. 2021). Following algal bloom senescence, heterotrophic bacteria play a crucial role in decomposing algal‐derived organic matter (detritus), thereby profoundly shaping pelagic energy fluxes and biogeochemical nutrient cycling (Azam and Malfatti 2007).
To address HABs more effectively, research attention has increasingly turned to algicidal bacteria, microorganisms capable of selectively killing certain algal species. In recent years, numerous algicidal strains have been isolated from diverse aquatic environments, and a range of algicidal compounds have been identified, demonstrating promising potential for bloom mitigation (Zhang et al. 2024; Ding et al. 2021). However, the practical application of these bacteria in natural settings remains constrained by an incomplete understanding of their molecular mechanisms of action. This review therefore focuses on elucidating the molecular basis of bacterial algicidal activity. We systematically examine bacterial‐algal interactions, categorise modes of algicidal action, summarise known molecular mechanisms and describe algal cell death pathways. By synthesising current knowledge in these areas, this work aims to deepen the understanding of bacterial‐mediated algal lysis, clarify the ecological roles of bacteria in algal population dynamics, and provide a foundation for developing more effective and targeted bacterial strategies for HAB control.
Bacterial‐Algal Interactions
2
In aquatic environments, particularly in marine ecosystems, the relationship between bacteria and microalgae is both complex and intricate. The concept of the phycosphere is similar to the rhizosphere of plants. The environment surrounding algal cells undergoes modifications due to their metabolic activities. For example, algal respiration can influence the surrounding oxygen levels and pH, while the exudation of organic substances from algae contributes to an organic‐rich environment surrounding the algal cells. This creates an environment with specific spatial dimensions and stability (Seymour et al. 2017). Similar to the rhizosphere of plants, the phycosphere also harbours a diverse assemblage of bacterial taxa. Broadly, the bacterial‐algal relationship can be classified into three categories.
Mutualistic Symbiosis
2.1
Soluble organic compounds released into the phycosphere, such as amino acids, carbohydrates, sugar alcohols and organic acids (Seymour et al. 2017), are generally available for bacterial utilisation. Upon entering this microenvironment, bacteria benefit by acquiring nutrients while also positively influencing algal growth, thus fostering a mutualistic relationship. For instance, when nutrient levels are low in the environment, phytoplankton growth heavily depends on nitrogen and phosphorus supplied by bacteria (Cole 1982). Additionally, bacteria can act as providers of nutrients through remineralization, which enhances the bioavailability of trace elements to algae (Amin et al. 2009). Furthermore, the synthesis of essential vitamins, such as vitamin B_12_, required by algae, often relies on the contribution of symbiotic bacteria (Croft et al. 2005). Recent studies indicate that bacteria also influence the dormancy and proliferation of algae. In particular, recent work reports that bacteria, via chemical signalling, can induce vesicle formation in ageing diatoms Coscinodiscus radiatus , facilitating the extrusion of reactive oxygen species (ROS), oxidised fatty acids and other deleterious metabolic byproducts from aged cells and thereby enabling renewed proliferation (Deng et al. 2024). Symbiotic bacteria can also play a role in interspecies competition among algae. For example, the addition of the symbiotic bacterium Bacillus sp. increases the abundance of Cyclotella atomus , a species of the centriae diatom, thereby displacing the previously dominant Fistulifera pelliculosa and becoming the new dominant species in the environment (Zhou et al. 2025).
Competition
2.2
The nutrients in the algal habitat not only attract symbiotic partners desired by algal cells but also lure opportunistic enemies. The availability of nutrients in the environment is a significant factor influencing the structure of microbial communities and can regulate algal populations (Jia, Lu, et al. 2023). Certain bacteria can suppress algal growth by modulating nutrient availability. For example, in interactions between Stenotrophomonas maltophilia , a freshwater oligotrophic bacterium and algal cells, the bacteria produce a hydroxamate‐type siderophore, an amino acid derivative, which limits the uptake of iron by cyanobacteria, thereby negatively impacting their growth (Z. Z. Liu 2014). Of course, algal cells are not easily eliminated; HABs and bloom‐forming algae, in particular, are adept at producing allelopathic compounds to inhibit competitors or outcompete other species for essential nutrients (Coyne et al. 2022). Nitrogen, for instance, is a crucial macronutrient in diatom‐bacteria competition. Benthic diatoms, with their high affinity for ammonia, limit the growth and metabolic activity of ammonia‐oxidising bacteria through competitive exclusion (Martens‐Habbena et al. 2009; Risgaard‐Petersen et al. 2004).
Antagonistic Interactions
2.3
Competition between algae and bacteria is not always subtle; in many cases, it escalates into active antagonism. Of particular threat to algal cells are algicidal bacteria, which, upon chemotactic attraction to the phycosphere, can eliminate algal cells to access released nutrients for their own proliferation. These attacks are mediated either through the secretion of algicidal compounds or via direct physical interactions. However, algae are not passive victims; they have evolved multiple strategies to counteract bacterial aggression. Studies have shown that, upon bacterial induction, algae can release a suite of antimicrobial compounds. For instance, diatoms secrete proteins (Paul and Pohnert 2011) and oxidised fatty acids (Ianora et al. 2011) with potent bactericidal properties. Additionally, algae may evade contact with lytic bacteria through dormancy strategies, such as the formation of cysts (Roth et al. 2008). Recent evidence also suggests a novel defence mechanism whereby diatoms reduce their cell size during reproduction to decrease surface area for bacterial adhesion (Cai et al. 2023). Algal cells are also capable of disrupting bacterial quorum sensing (QS) systems. Chlamydomonas reinhardtii , for example, produces chromophores that mimic bacterial acyl‐homoserine lactones (AHLs) (Teplitski et al. 2004), and further secretes bromocyanin, a secondary metabolite that catalyses the cleavage of AHL peptide bonds (Vanelslander et al. 2012), thereby interfering with bacterial QS signalling and mitigating virulence. Likewise, certain microalgae in aquatic systems secrete auxin analogues with indole structures that inhibit QS in Vibrio harveyi , impairing bacterial motility and biofilm formation (Yang et al. 2017).
With the variations in physicochemical conditions and the physiological state of algal cells, the interaction between bacteria and algae becomes increasingly complex (Zhang et al. 2021). For instance, members of the Rhodobacteraceae family, such as Phaeobacter gallaeciensis , can establish a potentially mutualistic relationship with the marine coccolithophore Emiliania huxleyi through the production of growth‐promoting hormones like phenylacetic acid and antibiotics such as protocatechuic acid. However, when E. huxleyi cells undergo senescence, P. gallaeciensis switches its role to become an opportunistic pathogen. This transition occurs upon detection of byproducts released during algal cell ageing, such as coumaric acid, prompting the bacterium to release lytic molecules, namely roseobactin A and B, which contribute to algal cell lysis (Seyedsayamdost et al. 2011). Furthermore, rising temperatures can lead to the transformation of symbiotic bacteria into antagonistic ones (Lin et al. 2024). For instance, the bacterium Tamlana sp. MS1 enhances the growth of the diatom Skeletonema costatum at 20°C, but as the water temperature increases, its algicidal activity intensifies, with a lysis rate reaching up to 92.4% at 25°C. This increased activity is attributed to the enhanced motility and adhesion of the bacteria at higher temperatures, facilitating direct contact with the algal cells and thereby enhancing its algicidal effect. Additionally, literature indicates that the production of algicides by bacteria is influenced by environmental nutrient conditions. Under oligotrophic conditions, such as nitrogen and phosphorus limitation, the production of algicides is promoted, whereas under optimal bacterial growth conditions, the production of such compounds is not required (Ray and Bagchi 2001).
Modes of Algicidal Activity
3
To better elucidate the specific mechanisms underlying bacterial algicidal activity, the modes of action can be categorised into direct contact and indirect algicidal strategies. In the direct contact mode, bacteria must physically interact with algal cells to exert their lethal effects. Experimentally, this is typically demonstrated when both washed bacterial cells and their corresponding culture supernatants exhibit algicidal activity, whereas the filtered supernatant, lacking bacterial cells, fails to inhibit algal growth (Shi et al. 2023). Several studies have provided further validation by co‐culturing algae and bacteria in compartmentalised systems in which dialysis membranes or similar barriers physically separate the two organisms, thereby preventing direct cell–cell contact while allowing exchange of metabolites between the compartments (Yu et al. 2024; Zeng et al. 2021). This approach is considered more rigorous, as it excludes bacteria that do not require physical contact but may produce algicidal compounds only in response to chemical cues released by algal cells. In contrast, the indirect mode does not require physical association between bacteria and algal cells. Instead, bacteria secrete diffusible algicidal compounds into the environment. In laboratory, the culture filtrate of bacteria or the bacterial secretions induced by algal metabolites exhibits algicidal activity, resulting in the death of algal cells (Zhang et al. 2024).
These two mechanisms are not entirely exclusive; on the contrary, they may co‐occur (Dai et al. 2024). For instance, Paucibacter sp. DH15 exhibits a strong removal effect on Microcystis aeruginosa through both sterile supernatants and bacterial suspensions after washing (Le et al. 2022). Additionally, Streptomyces sp. HY can simultaneously attack Anabaena through direct interaction, indirect interaction and biosorption mechanisms (Xie et al. 2024). The Enterobacterium sp. H6 has been shown to primarily exert indirect algicidal effects, with direct action being secondary, particularly against M. aeruginosa (Dai et al. 2024). These mechanisms may also vary depending on the host; for example, Bacillus cereus CZBC1 removes Oscillatoria chlorina and Oscillatoria tenuis through direct contact, while it employs indirect methods against Oscillatoria planctonica (Hu et al. 2019).
Direct‐Contact Algicidal Mechanisms
3.1
The known mechanisms of direct contact algicidal activity can generally be classified into three types. The first mechanism involves bacteria with well‐developed appendage‐like filaments, which directly contact and kill algae via these filaments. A classic example is the interaction between the algicidal bacterium * Streptomyces globisporus G9* and the cyanobacterium M. aeruginosa , where the bacterium wraps its hyphal filaments around the cyanobacterial cells to capture and kill them. Notably, cyanobacterial cells that are not in direct contact with the hyphae remain morphologically intact (Zeng et al. 2021). Similarly, the filamentous structures of Saprospira sp. SS98‐5, a saprophytic spirochete bacterium, can capture diatom cells, dissolve the diatom frustule at the point of contact, and subsequently invade and lyse the diatom cells (Furusawa et al. 2003). Literature suggests that this direct invasion mechanism shares similarities with bacterial predation behaviour (Bauer and Forchhammer 2021) (Figure 1A).
Bacterial mechanisms of algicidal activity through direct contact. (A) Streptomyces utilises its hyphal network to target and lyse algal cells. (B) Following contact with algal cells, bacteria secrete proteases or enhance protease activity. (C) After bacterial contact with algal cells, the concentration of algicidal compounds within algal remnants increases.
The second mechanism occurs when bacteria only generate active algicidal compounds upon contact with molecular structures on the surface of algal cells. For example, Chitinimonas prasina LY03 uses its flagella to anchor itself to the surface of algal cells. Upon attachment, it produces chitinase that degrades the algal cell wall (Li, Lei, et al. 2016), Similarly, the free‐living Alteromonas sp. L15 does not exhibit high protease activity unless it adheres to algal cells, at which point it maintains high protease activity capable of hydrolyzing the glycoproteins in the cell wall. This suggests that the chemical cues on the surface of Thalassiosira pseudonana cells may play a crucial role in triggering the protease activity of L15 (Cai et al. 2023) (Figure 1B).
The third mechanism involves bacterial metabolites accumulate around algal cells after contact, increasing local concentrations and potentially reaching algicidal doses. For example, Roseobacter sp. can adhere to the green alga E. huxleyi , initially utilising tryptophan secreted by the algae to enhance the production of the plant hormone indole‐3‐acetic acid (IAA). At this stage, the concentration of IAA does not harm the algal cells. However, after co‐culturing for 3 weeks, when the concentration of IAA reaches a threshold, it becomes lethal to the algal cells (Segev et al. 2016) (Figure 1C).
Indirect Bacterial Algicidal Mechanisms
3.2
Indirect algicidal activity refers to bacterial inhibition or killing of algal cells without physical contact, instead relying on the secretion of algicides. These secreted compounds may be constitutive (Whalen et al. 2018), metabolites inherently toxic to algal cells, or inducible, in which case the production of algicidal substances is triggered by specific algal‐derived molecules. For example, the algal metabolite dimethylsulfoniopropionate (DMSP) can induce bacteria to produce algicides, thereby enabling bacteria to acquire additional sulphur resources (Wang and Seyedsayamdost 2017; José 2018) (Figure 2).
Constitutive and inducible indirect algal toxic bacteria.
To date, a wide diversity of indirect algicides has been identified. Broadly, these include alkaloids, amino acid derivatives, peptides and proteins, enzymes, polyketides, terpenoids, fatty acids and their derivatives, etc. (Li, Xing, et al. 2024). Yang et al. (2025) compiled a comprehensive survey of approximately 250 natural algicidal compounds isolated from bacteria between the 1960s and 2024, categorising them into algal‐associated and incidental agents, and further distinguishing their relative algicidal potency.
These algicides act on diverse cellular targets. For instance, 79 derivatives were synthesised from the parent compound N ^1^‐benzyl‐N ^3^, N ^3^‐diethylpropane‐1,3‐diamine to probe structure–activity relationships and infer specific sites of action (Park et al. 2023). Another example is the γ‐proteobacterium Microbulbifer sp. RZ01, which produces 3,3′,5,5′‐tetrabromo‐2,2′‐dihydroxydiphenyl (4‐BP). This compound exerts algicidal effects by inhibiting plastoquinone biosynthesis, thereby disrupting multiple essential metabolic pathways in phytoplankton (Zhang et al. 2023). Genes responsible for 4‐BP biosynthesis are widespread among diverse bacterial taxa in marine environments, suggesting that it may represent a globally relevant bacterial tool for mediating antagonistic bacteria–algae interactions. Mechanistic studies have revealed that 4‐BP competes with homogentisate for the active site of homogentisate solanesyltransferase (HST), a key enzyme in the plastoquinone biosynthetic pathway, ultimately suppressing photosynthesis and impairing algal viability.
Molecular Mechanisms Underlying Bacterial‐Mediated Algal Lysis
4
In aquatic environments, bacteria associated with algal cells do not kill algae by a simple secretion of algicidal compounds. Instead, they need to sense and locate planktonic algae, mobilise towards them and even enter the phycosphere; upon contacting algal cells, they actively or passively regulate their own behaviour to release algicidal substances or to enact other algicidal actions. This process constitutes an integrated, system‐level programme of resource acquisition and interaction with the algae (Meyer et al. 2017). In this section, we summarise the molecular mechanisms that underlie bacterial behaviour during algicidal activity, including how bacteria sense cues from algae, approach and adhere to algal cells, how collective (group) behaviours contribute to algicidal function, and the emerging mechanism of vesicle‐mediated delivery of algicidal substances.
Bacterial Chemotaxis Towards Algal Cells
4.1
Chemotaxis refers to the ability of bacteria to sense environmental chemoattractants or chemorepellents and modulate flagellar rotation to direct their movement accordingly, thereby adapting to environmental cues. In the context of bacterial algicidal activity, chemotaxis primarily serves to reduce the physical distance between bacteria and algal cells, enhancing the efficiency of attack (Xu, Ali, et al. 2024). Similar to the rhizosphere bacteria that chemotactically respond to plant‐derived signals, algal cells and their extracellular products act as attractants for algicidal bacteria (Seymour et al. 2017, 2009). These extracellular products are mainly low‐molecular‐weight compounds such as amino acids, monosaccharides, organic acids and secondary metabolites (Bell and Mitchell 1972; Sjoblad and Mitchell 1979). Recent studies have identified oxygen as an additional chemoattractant for bacteria targeting algae (Prakash et al. 2025). Among these chemoattractants, DMSP has been reported as a potent attractant for multiple marine algicidal bacteria (Seymour et al. 2010). The DSYE gene responsible for DMSP synthesis is present in a wide range of algal taxa (Wang et al. 2024), suggesting that DMSP is a broadly conserved chemical cue mediating bacteria–algae interactions in marine environments. Furthermore, the addition of DMSP in bacteria–algae co‐culture systems can enhance bacterial algicidal potency (José 2018), implying a potential role in bloom regulation (Xu, Ali, et al. 2024). Beyond such common attractants, different bacterial species display distinct preferences when interacting with dinoflagellates (Yang et al. 2024). For instance, Polaribacter marinivivus and Lentibacter algarum preferentially assimilate amino acids and dipeptides, whereas Litoricola marina favours nucleotides (Han et al. 2021). Algal cells can also release macromolecules such as glycoproteins, although the secretion of these larger compounds generally lags behind that of small molecules and is more prominent during bloom decline (Seymour et al. 2017). Although less studied, chemotaxis towards algal macromolecules has also been documented. Marine bacteria exhibit strong chemotactic responses to kelp‐derived polysaccharides, providing direct evidence that bacteria can actively target abundant polymeric substrates. Highly chemotactic strains often belong to Pseudomonadaceae, Alteromonadaceae and Pseudoalteromonadaceae, taxa with notable algicidal capabilities. It has been hypothesised that kelp polysaccharides may strongly attract algicidal bacteria during bloom senescence. Intriguingly, pre‐exposure to DMSP can enhance bacterial chemotaxis towards kelp polysaccharides, likely by providing sufficient methyl donors to increase the sensitivity of methyl‐accepting chemotaxis proteins (MCPs), thereby amplifying chemotactic responsiveness (Clerc et al. 2023). In addition to secreted products, algal cells themselves possess inherent attractant properties. For example, in the chemotaxis of Vibrio coralliilyticus towards K. mikimotoi, the intact algal cells were found to strongly attract bacteria, with membrane components identified as key mediators of this effect (Yu et al. 2024).
Unlike stationary plant cells, planktonic algae in aquatic environments are not merely fixed in place. They are passively displaced by turbulence (Cencini et al. 2019), and some planktonic algae possess flagella and exhibit motility (Basterretxea et al. 2020). This dynamic behaviour places higher demands on the motility capabilities of associated bacteria, which must relocate and navigate to encounter and interact with their algal hosts. Marine bacteria often exhibit chemotactic responses more than an order of magnitude faster than the model organism Escherichia coli (Stocker et al. 2008), with greater swimming speeds and directional persistence (Stocker et al. 2008; Stocker and Seymour 2012; Garren et al. 2014; Son et al. 2016). This enables them to closely track motile algal cells (Barbara and Mitchell 2003). Certain species, such as Vibrio alginolyticus , employ specialised swimming strategies, propelled by a single polar flagellum; they alternate between rapid runs and sharp 180° reversals or 90° ‘flicks’ (Son et al. 2016), conferring advantages in nutrient foraging within oligotrophic seawater. Despite their high motility, marine bacteria adopt slower swimming speeds when tracking small phytoplankton, which prolongs the sensory integration time and improves search efficiency (Foffi et al. 2024).
In the classical E. coli chemotaxis paradigm, extracellular ligands are detected by MCPs (Parkinson 2015), which activate the CheA/CheY two‐component system to modulate flagellar rotation and thus swimming direction. In nutrient‐poor marine environments, bacteria have evolved an expanded repertoire of MCPs, enhancing their adaptive capacity (Lacal et al. 2010). Notably, the genomes of algicidal Roseobacter clade (MRC) bacteria typically encode c‐di‐GMP signalling systems, with over half of MRC genomes containing genes for motility, chemotaxis and diverse chemoreceptors, likely facilitating algal localization and stable interspecies interactions (Dang and Lovell 2016) (Figure 3A).
(A) Algicidal bacteria migrate towards algal cells via the canonical chemotaxis pathway. (B) Bacterial attachment to algal cells is mediated through flagella, adhesins, and pili.
Chemotaxis is also implicated in the initial stages of bacterial surface colonisation. For instance, deletion of the CheA/CheB two‐component system in Marinobacter adhaerens HP15 significantly reduced its attachment to diatoms (Sonnenschein et al. 2012). Similarly, non‐motile or morphologically altered mutants of Silicibacter sp. TM1040 exhibited impaired attachment to dinoflagellates (Miller and Belas 2006). Once chemotactic bacteria enter the phycosphere and attach to phytoplankton cells (Mayali et al. 2011) or extracellular matrices, they can maintain proximity to their targets and maximise algicidal activity (Seymour et al. 2009, 2008).
Bacterial Attachment to Algal Cells and the Formation of Bacterial–Algal Biofilm Consortia
4.2
Attachment represents a critical step in the direct algicidal activity of bacteria, as it prolongs the period of interaction between bacterial and algal cells. Live‐cell imaging combined with scanning and transmission electron microscopy has directly visualised the attachment of algicidal bacterium GD3 and V. coralliilyticus to the cell membrane of K. mikimotoi cells (Shi et al. 2023; Yu et al. 2024). Depending on their lifestyle, bacteria can be classified as particle‐attached or free‐living (Shi et al. 2023), and field surveys have shown that the abundance of particle‐attached bacteria with algicidal activity in diatom blooms is approximately fivefold higher than that of free‐living counterparts (Park et al. 2010). In co‐culture experiments, the number of attached bacteria per diatom cell is strongly and positively correlated with the rate of algal lysis (Cai et al. 2023). Furthermore, the addition of organic amendments that enhance bacterial adhesion significantly increases algal lysis rates. Collectively, these observations underscore the pivotal role of bacterial adhesion in mediating algicidal effects. The algicidal bacteria, through their adhesion to algal cells, not only prolong the duration of their interaction (Slightom and Buchan 2009), but may also serve as a prerequisite for the algicidal effect. In some cases, adhesion is not merely advantageous but may be essential. For instance, Alteromonas sp. L15 requires adhesion to the diatom surface to sense algal surface‐derived chemical cues that activate extracellular proteases capable of degrading frustulin, thereby enabling algal cell lysis (Cai et al. 2023).
The molecular mechanisms underlying bacterial adhesion to algal cells remain incompletely understood, but both flagella and pili appear to be important mediators. In the case of the chitinase‐producing strain LY03 interacting directly with the diatom T. pseudonana , bacteria were observed to attach to algal surfaces via flagella before secreting chitinases to degrade the algal cell wall, ultimately leading to cell rupture (Li, Lei, et al. 2016). This process may be regulated by a two‐component system, as Pseudoalteromonas piscicida has been shown to modulate chitinase gene expression via the CdsS/CdsR pathway following surface colonisation (Miyamoto et al. 2007). Metagenomic surveys of bacteria during bloom events dominated by diatoms have identified tad gene clusters associated with tight adhesion, suggesting that these clusters encode pili enabling firm attachment to diatoms (Isaac et al. 2021). Beyond adhesion, pili have also been implicated in virulence regulation, as seen in Pseudomonas aeruginosa (Persat et al. 2015). Some members of the phylum Bacteroidota, which lack flagella, may attach to algal surfaces through stochastic encounters and subsequently move across the cell surface via gliding motility (Buchan et al. 2014). Although the precise mechanisms of gliding remain elusive, type IV pili, extracellular polysaccharides (EPS) and adhesins are likely involved (McBride 2001). Genomic analyses of the gliding bacterium Flavobacterium have identified adhesins with conserved peptide motifs, cadherins and bundled filament proteins, all of which may contribute to adhesion (Woyke et al. 2009) (Figure 3B).
Typically, particle‐associated bacteria produce extracellular polymeric substances (EPS), adhesins and other matrix components upon attachment to organic matter, linking individual cells into structured microbial aggregates known as biofilms (O'Toole et al. 2000). These biofilms possess interconnected channels that facilitate the exchange of nutrients, water and gases, thereby optimising resource use and conferring resistance to shear forces in aquatic environments (Bartual et al. 2017) (Steele et al. 2014). In bacterial–algal interactions, initial bacterial attachment to algal surfaces can be reinforced through the reciprocal stimulation of extracellular product secretion, resulting in bacterial–algal consortia with enhanced adhesion and algicidal efficiency. It has been demonstrated in the model system involving the marine coccolithophore E. huxleyi and the algicidal bacterium Phaeobacter inhibens (Lipsman et al. 2024) that succinate, secreted by algal cells, may act as a chemical signal that promotes bacterial secretion of EPS. This signalling enhances bacterial adhesion and facilitates EPS release, which in turn stimulates the production of transparent exopolymer particles (TEP) by the algal cells. The formation of TEP contributes to the establishment of an extracellular matrix (ECM), thereby promoting the formation of a distinct algal‐bacterial consortium. In such consortia, bacteria are better equipped to exert their algicidal effects. Moreover, other studies suggest that IAA secreted by Vibrio species (Gutierrez et al. 2009) may play a role in stimulating the mucus exudation of marine algae and in the formation of Vibrio‐algal consortia (Mazur and Homme 1993). Once attached to algal cell surfaces, bacteria can enrich IAA concentrations, and at high levels, IAA exhibits potent algicidal activity (Segev et al. 2016).
Role of QS in Algicidal Processes
4.3
QS is a specialised intra‐ and interspecific communication mechanism among bacteria, enabling population density‐dependent perception, signal transduction and adaptive responses. Canonical QS pathways are characterised by the production, release and detection of low‐molecular‐weight signalling molecules, collectively termed autoinducers, which, upon reaching threshold concentrations indicative of sufficient population size, trigger coordinated behaviours (Dang and Lovell 2016).
In recent years, the role of QS in bacterial algicidal activity has attracted increasing attention. Certain marine bacteria produce small‐molecule compounds that act as QS signals. Within the phylum Proteobacteria, cell–cell communication is commonly mediated by AHLs, encompassing both short‐chain and long‐chain variants. Short‐chain AHLs include N‐hexanoyl‐l‐homoserine lactone (C6‐HSL), N‐3‐hydroxyhexanoyl‐l‐homoserine lactone (OH‐C6‐HSL) and N‐3‐oxohexanoyl‐L‐homoserine lactone (oxo‐C6‐HSL), while long‐chain AHLs include N‐tetradecanoyl‐L‐homoserine lactone (C14‐HSL) and N‐3‐hydroxytetradecanoyl‐l‐homoserine lactone (Stock et al. 2020). These QS molecules contribute to algicidal activity through three primary mechanisms. First, some act directly as algicidal agents. For example, Pseudomonas piscicida releases 2‐heptyl‐4‐quinolone (HHQ), which induces mortality in E. huxleyi (Harvey and Whalen 2016), potentially by disrupting electron transport in the photosynthetic system (Dow et al. 2020). In diatom‐focused studies, short‐chain AHLs generally did not affect growth, whereas certain long‐chain AHLs exhibited inhibitory or lethal effects. Transcriptomic analyses suggest that these long‐chain AHLs may impede growth by modulating lipid metabolism and cell cycle progression (Stock et al. 2020). Evidence exists that bacterial quorum‐sensing molecules alter the composition and abundance of lipids in algal cells (Parveen and Patidar 2023). Second, QS enables coordinated regulation of diverse algicidal compounds (Dow 2021). In Aeromonas sp. GLY‐2107, the production of two algicidal metabolites is regulated by short‐chain AHLs, with C4‐HSL serving as a key signal. Disruption of the agyI AHL synthase gene abolished production of these compounds (Guo et al. 2016). Similarly, the AI‐2 signal of Deinococcus sp. Y35 markedly enhances the synthesis of algicidal pigments, increasing its algicidal potency (Han et al. 2023). Third, QS molecules can enhance algicidal efficiency by modulating bacterial motility and adhesion, thereby improving colonisation of the phycosphere. QS signalling has been shown to suppress motility while promoting adhesion via flagellar regulation (Fei et al. 2020). During their investigation of the phycosphere microecosystem associated with the diatom Asterionellopsis glacialis , Fei et al. (2020) demonstrated that bacterial attachment to algal cells is restricted to taxa equipped with a fully functional quorum‐sensing system. The QS molecule 3‐oxo‐C16:1‐HSL strongly inhibited bacterial motility and promoted attachment, and metagenomic analyses have detected increased AHL concentrations during bacterial colonisation of algal cells (Figure 4A) (Isaac et al. 2021).
(A) The role of bacterial quorum sensing in algal killing (a). Quorum‐sensing signals in bacteria directly induce cytotoxicity in algal cells. (b) These signals orchestrate the biosynthesis of algicidal metabolites. (c) Quorum‐sensing pathways modulate extracellular polysaccharide secretion and flagellar function, strengthening bacterial adhesion and promoting algal cell demise. (B) Bacterial delivery of algicidal substances via vesicle‐mediated transport.
The genetic basis of QS‐mediated algicidal activity varies across taxa. In Bacillus, the NprR/NprX system mediates QS‐dependent regulation of high‐molecular‐weight algicidal compounds against M. aeruginosa , with gene disruption leading to reduced activity (Wu et al. 2017). In Gram‐negative bacteria, loss of luxS results in decreased algicidal compound production and diminished activity (Liu et al. 2022).
Numerous studies have reported a positive correlation between algicidal efficacy and bacterial density (F. Liu 2023; Jia, Cheng, et al. 2023; Shi et al. 2013). Among the most extensively investigated algicidal bacteria are genera such as Pseudomonas sp., Pseudoalteromonas sp., Vibrio sp. and Alteromonas sp., whose algicidal activity is often regulated by QS systems (Kahla et al. 2021). The activation of QS is closely linked to bacterial density: as cell density increases, the concentration of signalling molecules rises until a threshold is reached, triggering receptor activation and downstream gene expression (Williams et al. 2007). Therefore, in studies of QS‐mediated algicidal bacteria, bacterial concentration is likely a key factor influencing their algicidal capacity (Shi et al. 2013). However, it remains to be conclusively demonstrated whether QS‐dependent algicidal activity is strictly density‐dependent. Thus, further research to elucidate the interplay among bacterial density, QS activation and algicidal function will be crucial for advancing our mechanistic understanding and practical application of bacterial agents in algal bloom control.
Bacterial Delivery of Algicidal Compounds via Extracellular Vesicles
4.4
Extracellular vesicles (EVs) are membrane‐bound structures secreted into the extracellular milieu, enriched with proteins, nucleic acids, signalling molecules and toxins. They have been identified in bacteria, archaea and eukaryotes (Di Naro et al. 2025), functioning as mediators of intercellular communication. In recent years, EVs have attracted considerable attention, with most studies focusing on mammalian cells and model bacterial species; however, their ecological roles in aquatic systems are increasingly recognised (Schatz 2018). Algal cells can secrete vesicles as an early response to viral infection. Upon infection, algae release large quantities of vesicles that encapsulate viral particles, thereby prolonging viral half‐life in the environment and increasing the risk of infection. When these infection‐derived vesicles are internalised by healthy algal cells, they enhance the cells' susceptibility to subsequent viral attack, resulting in accelerated lysis and higher viral yields. This process sustains effective viral infection and transmission within the population and can contribute to the termination of algal blooms (Schatz et al. 2017).
Bacterially derived EVs also play a pivotal role in algicidal interactions. For instance, outer membrane vesicles of the coral pathogen Vibrio shilonii have been found to contain quorum‐sensing signal molecules such as AHL (Li, Azam, et al. 2016), Given the close association between coral disease and its symbiotic dinoflagellates (Symbiodinium) (Banin et al. 2001), it is plausible that bacteria may influence coral health by delivering vesicle‐contained factors that adversely affect these symbionts. Moreover, bacterial EVs can directly transport algicidal compounds. Recent work on Chitinimonas sp. LY03 revealed that its membrane vesicles exhibit algicidal activity comparable to that of the producing bacterial strain. Purification and structural elucidation via nuclear magnetic resonance spectroscopy and high‐resolution mass spectrometry identified the active agent as a pyrrole‐core macrolactam belonging to the tambjamine antibiotic family, designated Tambjamine LY2. Microscopic imaging confirmed that LY2 enters Heterosigma akashiwo and T. pseudonana cells via membrane fusion. These findings demonstrate that bacterial vesicles can encapsulate and efficiently deliver hydrophobic algicidal metabolites to algal targets, opening a new chapter in our understanding of bacteria–algae interactions (Figure 4B) (Li, Wang, et al. 2024).
Mechanisms of Algal Cell Death
5
Algal cells collapse and lyse under the stress imposed by algicidal substances secreted by bacteria, releasing their intracellular contents and thereby supplying nutrients to the bacteria. The dominant mechanisms by which different algicidal compounds induce algal cell death may differ (Yang et al. 2025). Current evidence indicates that bacterial induction of algal cell death predominantly occurs through disruption of cell membrane integrity, impairment of photosynthesis, induction of oxidative stress and perturbation of calcium homeostasis.
Loss of Cell Integrity
5.1
Algicidal compounds produced by bacteria can act on the constituents of algal cell walls and plasma membranes, leading to cell lysis. For example, chitinase secreted by Chitinophaga sp. can degrade chitin, which is a major constituent of diatom cell walls (Li, Lei, et al. 2016). In addition, some bacteria secrete beta‐glucosidase and other glycosidases as algicidal agents. Their targets may lie in the cell wall. It has been reported that, in coculture with Prorocentrum micans , Alteromonas sp. exhibit increased beta‐glucosidase gene expression, while the alga's cell‐wall polysaccharide content declines, providing a potential piece of evidence for this mechanism (Shi et al. 2018). Marine bacterium Flammeovirga yaeyamensis shows activities of amylase, cellulase and xylanase, which are associated with the degradation of algal cell walls (Chen et al. 2013). Some small‐molecule algicidal compounds can cross the cell membrane into the cytoplasm, causing cytosolic acidification over time (Lu et al. 2016), potentially triggering ROS production and lipid peroxidation, ultimately leading to intracellular dissolution of algal cells. Beyond targeting cell‐wall and membrane components, certain algicidal substances can alter ion permeability, resulting in cell rupture (Jeong and Son 2021). For instance, violacein produced by Chromobacterium violaceum can rapidly disrupt algal cell permeability, provoking prolonged oxidative stress and causing algal cell rupture and death (Cai et al. 2024).
Photosynthetic Uncoupling and Inhibition of Photosystem Activity
5.2
Photosynthetically fixed solar energy is the primary energy source for unicellular phytoplankton, and damage to the photosynthetic apparatus can directly lead to cell death (Li, Chen, et al. 2024). Chlorophyll A, a key pigment of the photosynthetic system, not only reflects the photosynthetic state of algal cells but is also frequently used as an indicator of algal mortality in many bacterium‐derived algicidal studies (Zhang et al. 2024; Li, Chen, et al. 2024; Liu et al. 2024). When the photosynthetic system is inhibited or disrupted, chlorophyll A content, photosynthetic efficiency and electron transport rate all decline markedly (Jia, Lu, et al. 2023), indicating that the cells are near collapse.
Certain algicidal compounds act by perturbing the electron transport chain within the photosynthetic apparatus to suppress photosynthetic activity. Sphingomonas sp. M‐17 produces argimycin A, which can selectively inhibit cyanobacterial growth; studies suggest this may disrupt the function of the phycobilisome within PSII, preventing energy transfer from the phycobilisome to chlorophyll A and thereby affecting light‐energy conversion (Hibayashi and Imamura 2003). In addition, algicidal substances secreted by Bacillus subtilis , such as 4‐acetamido‐butanoic acid (4‐ABC) and 8‐hydroxyquinoline (8‐HQL), have been shown, via measurements of chlorophyll fluorescence, to suppress photosystem activity in M. aeruginosa . They reduce photosynthetic parameters Fv/Fm, ѱ_0_ and φ_E0_ (the PSII photochemical efficiency and the electron transport efficiency beyond QA), while increasing the energy‐dissipation quantum yield φD0, indicating these compounds hinder PSII electron transfer and compromise photosynthetic performance (Liu et al. 2024). The 2‐MECHD compound produced by Pseudomonas fragi YB2, which resembles an acetylacetonate structure capable of chelating metals, suggests that its target within the photosynthetic electron transport chain may be non‐heme iron in PSII, plastoquinone (PQ) and the iron–sulphur cluster in ferredoxin (Fd) (Zhang et al. 2024). Further, the cytochrome b6f complex has been identified as a major binding site for 2‐alkyl‐4‐quinolones (HHQ) isolated from algicidal marine bacteria (Dow et al. 2020).
Beyond action on the electron transport chain, some algicidal compounds can uncouple the algal photosynthetic system, altering the outcome of photosynthesis. For example, a marine Vibrio sp. (Vibrio shiloi) (Banin et al. 2001) produces a linear proline‐rich 12‐mer peptide toxin P that, in the presence of ammonium ions, inhibits photosynthesis in a dinoflagellate, likely by promoting ammonium uptake and perturbing the cellular proton gradient, thereby causing photosynthetic uncoupling. The photosynthetic system of algal cells operates as a tightly integrated, sensitive machine; disruption of any link can derail energy conversion, and given its central role in algal survival, the photosynthetic apparatus represents a favourable target for algicidal strategies.
Oxidative Stress and ROS Generation
5.3
ROS refer to hydrogen peroxide, superoxide anion, singlet oxygen, hydroxyl radicals and other species generated during electron transfer in photosynthesis and respiration as well as in other enzymatic reactions. These molecules are highly oxidising, and under normal conditions cellular antioxidant enzymes and antioxidants promptly detoxify them to maintain redox balance. When cells face environmental stress, ROS production increases. If not promptly scavenged, accumulated ROS can attack cellular structures, including DNA and proteins, promote lipid peroxidation and compromise membrane integrity, ultimately leading to cell death (Fulda et al. 2010; Mallick and Mohn 2000).
ROS generation is a common phenomenon when algal cells are subjected to algicidal bacteria or algicidal substances (Li et al. 2015; Lin et al. 2023; Zhang et al. 2014). This may reflect multi‐factorial causes: ROS production can arise as part of algal defence responses (Low and Merida 1996), or result from damage to cellular structures caused by algicidal compounds, such as impairment of photosynthetic systems that elevates ROS (Zhang et al. 2014). The algal antioxidant system comprises enzymatic antioxidants (SOD, CAT, APX and POD) and non‐enzymatic antioxidants (ascorbate, glutathione and carotenoids), which are commonly used to assess the oxidative stress status of algal cells (Mallick and Mohn 2000). Malondialdehyde (MDA) content is another important parameter for judging oxidative stress, as MDA is one of the terminal products of lipid peroxidation and reflects the extent of membrane lipid oxidative damage (Tsikas 2017). For example, in the indirect algicidal action of Bacillus subtilis on Spirogyra gracilis, the contents of MDA, POD, CAT and SOD increase, indicating that the algicidal compounds induce oxidative damage in the algal cells (Gu et al. 2024). Prolonged exposure to stress can lead to algal apoptosis. In diatom cells subjected to stress from the algicidal bacterium Bacillus mycoides , cysteine proteases involved in programmed cell death were activated, which can scavenge ROS and attenuate the ROS‐induced upregulation of glutathione synthetase expression; concurrently, the death‐specific protein (DSP), implicated in transcriptional regulation of photosynthesis, also increased. These findings suggest that diatoms employ mechanisms to neutralise excess ROS under oxidative stress while concurrently initiating programmed cell death (Bayramova et al. 2024). Other work suggests that excessive ROS produced by K. mikimotoi under stress can promote apoptosis by stimulating caspase‐3 (Han et al. 2018).
Disruption of Calcium Homeostasis
5.4
Calcium ions (Ca^2+^) act as pivotal signalling molecules in a wide range of cellular processes, including maintaining the stability of the cell wall and plasma membrane, regulating cell proliferation and modulating metabolic activity (Bagur and Hajnóczky 2017). In studies on algicidal bacteria targeting Chlamydomonas reinhardi, it was found that both pyrrolnitrin and pyoluteorin secreted by Pseudomonas protegens perturb intracellular Ca^2+^ dynamics. Exposure of algal cells to 20 μM pyrrolnitrin induced a rapid and pronounced intracellular Ca^2+^ surge, while treatment with 100 μM pyoluteorin similarly elevated Ca^2+^ levels, triggering explosive cell death. Permeability assays suggested that this interference with algal Ca^2+^ homeostasis may occur via a mechanism independent of membrane rupture (Rose et al. 2021). Further evidence was obtained in the interaction between P. fragi YB2 and Chlorella, where non‐invasive micro‐test technology (NMT) was used to directly quantify cytosolic Ca^2+^ fluxes without damaging algal cells. Application of YB2 supernatant significantly increased Ca^2+^ influx in Chlorella, disrupting intracellular calcium homeostasis (Zhang et al. 2024).
Other Mechanisms of Algal Cell Death
5.5
Beyond the commonly described algal cell death mechanisms, other death pathways have been explored. For example, algicidal substances can affect the cell cycle to induce programmed cell death in algae (Wang and Coyne 2023), and death can also result from inhibition of algal cell division (Li, Lin, et al. 2014; Van Tol et al. 2017). In the transcriptome of H. akashiwo exposed to Pseudalteromonas sp. LD‐B1, upregulation of autophagy‐related genes such as the ubiquitin‐like modifier‐activating enzyme (ATG7), Beclin (BECN) and the 5′‐AMP‐activated protein kinase (PRKAA) was observed, indicating an autophagic response in the algal cells (Xu, Chen, et al. 2024). Additionally, during Microbulbifer sp.‐mediated killing of Phaeocystis globosa , the algal cells may undergo autophagic cell death (Zhu et al. 2022).
Overall, algal cell death is a complex and multifaceted process. It can arise not only from direct lysis inflicted by algicidal substances but also from disruption of normal cellular physiology that triggers programmed cell death. ROS play a central role as key signalling mediators in death regulation. Mechanical damage, impairment of photosynthesis (Pospíšil 2012), and upregulation of intracellular Ca^2+^ caused by algicidal bacteria can all provoke ROS production; likewise, under stress, excessive ROS accumulation damages organelles and biomolecules, forcing the cell to choose between survival and death (Zhang et al. 2020). ROS can regulate downstream responses via eight distinct pathways to either rescue the cell or induce death (Tang et al. 2025). ROS not only act as signals of programmed cell death alongside elevated Ca^2+^, but ROS‐induced lipid peroxidation products also participate in death induction (Aguilera et al. 2022), and ROS is linked to autophagic death as well (Pérez‐Pérez et al. 2012). The modes of death are intricately interconnected; cumulative interactions lead to cellular collapse, and the dominant mode may be determined by the specific algicidal substance and the algal species involved. Therefore, when investigating algal death mechanisms, identifying the principal sites of action may be the most critical aspect to emphasise (Figure 5).
Mechanisms by which bacteria and their algicidal compounds induce algal cell death: Bacteria can disrupt the integrity of extracellular structures and impair algal photosynthesis. Algal defence responses trigger a substantial increase in reactive oxygen species (ROS), which elevate intracellular Ca2+ concentrations and activate programmed cell death in the algal cells.
Discussion
6
As researchers screen for bacteria with high algicidal activity from the phycosphere, notable progress has been achieved using algicidal bacteria to mitigate blooms. Bacillus subtilis S4, for example, can lyse M. aeruginosa with an algicidal kill rate of up to 98%, and about 89% of the released microcystins are degraded by the bacteria, suggesting potential for remediation of nutrient‐rich waters experiencing blooms (Chen et al. 2024). Similarly, Paenibacillus sp. can reduce toxin concentrations by approximately 90% when targeting M. aeruginosa (Jia, Cheng, et al. 2023). In addition, Pseudomonas sp. Ps3 demonstrates substantial bloom‐dissolving activity against the dinoflagellate Gymnodinium catenatum and K. mikimotoi, achieving algicidal rates of 83.0% and 78.3%, respectively (Zheng et al. 2023). Nevertheless, practical deployment of algicidal bacteria faces several challenges. In laboratory co‐culture systems, confined conditions allow bacteria to exert strong algicidal effects, but environmental variability in natural settings may compromise efficacy. Pseudomonas fluorescens SK09 is the most frequently studied algicidal bacterium in both laboratory studies and field surveys and can effectively control harmful blooms, and it exhibits robust, species‐specific algicidal activity against Stephanodiscus hantzschii in vitro (Jung et al. 2010). However, field trials show a significant reduction in algicidal activity (Noh et al. 2017). Moreover, the life state of algal cells can differ under varying growth conditions; for instance, M. aeruginosa tends to occur as colonies in natural waters but as single cells under laboratory culture, a difference driven by the production of extracellular polymers by associated bacteria that promotes aggregation in situ (Wang et al. 2016). Beyond inconsistent efficacy, safety and contamination concerns must also be addressed: do algicidal preparations affect non‐target organisms, and do the bacterial cultures leave residual nutrients or other contaminants (F. Liu 2023)? Additionally, can the toxins released from lysed cells be reliably removed or purified in practice? These considerations underscore the need for careful ecological risk assessment and the development of field‐relevant strategies before any widespread application.
As these challenges drive deeper investigation, we recognise that bacteria–algae interactions in natural settings encompass three main relationships: symbiosis, competition and antagonism. These interactions are largely governed by nutrient acquisition and may shift with environmental conditions. Algicidal bacteria can suppress algal cells through direct contact or via secreted active compounds. Direct‐contact killing may require bacterial appendages, such as flagella or pili, to deliver attack or may be triggered only when algal surface–released cues induce the production of active algicidal agents. Direct contact also shortens the effective distance and elevates local algicidal concentrations, thereby enhancing efficacy. Indirect algicidal action relies on diffusible algicidal substances; constitutive compounds are produced irrespective of algal exudates, whereas inducible compounds are generated only after sensing algal‐derived signals. In aquatic environments, bacteria detect algal attractants through MCPs to modulate flagellar activity and execute chemotaxis into the phycosphere, a key prerequisite for in situ algicidal activity. Adhesion following chemotaxis represents further engagement with the algal cell and is necessary for some direct‐contact algicidal strains; bacterial flagella, pili and secreted adhesins likely facilitate this adhesion, potentially under quorum‐sensing control. Moreover, quorum‐sensing signals themselves can act as algicidal molecules or serve as switches for algicidal gene expression. Emerging work indicates that bacterial vesicles can function as cargo vectors for algicidal substances, broadening the modes of bacterial algicidal action and revealing important facets of alga–bacteria communication. Under exposure to algicidal bacteria or substances, algal cell walls or membranes may be degraded, leading to cell lysis; damage to the photosynthetic apparatus triggers oxidative stress, and accumulated ROS oxidise membrane lipids and DNA, causing cellular injury. Calcium ions and ROS often act synergistically as signals that coordinate the death process.
Despite presenting a refined description of algicidal bacteria, this field lacks the depth typical of mechanistic studies in bacteria–host interactions elsewhere, highlighting the necessity for rigorous, high‐quality investigations to uncover the underlying mechanisms. In recent years, the maturation and cost reduction of omics technologies have led to an increasing number of studies employing multi‐omics approaches to investigate the molecular mechanisms of bacteria‐algae interactions, a development that could significantly aid future research. For instance, in the interaction between Brevibacillus laterosporus and M. aeruginosa , bacterial transcriptome sequencing following co‐culture indicated that bacteria may degrade amino acids and fatty acids as sources of ATP and energy, while membrane transport proteins secrete hydrolases and proteases into the external environment to disrupt algal cells, leading to cell death and achieving algicidal effects (Zhang et al. 2022). Jiaying Yu et al. through transcriptomic analysis of bacterial‐algal interactions not only explored the algicidal mechanisms of the direct killing bacterium V. coralliirubri against K. mikimotoi but also provided clues regarding the underlying mechanisms of algal cell death (Yu et al. 2024). Similarly, Lee et al. (2024) utilised tandem mass spectrometry‐based proteomics to investigate the interaction between the algicidal bacterium Maribacter dokdonesis P4 and the red tide algae K. mikimotoi. They found that bacterial stress led to the downregulation of key mitochondrial and chloroplast proteins in the algae, which inactivated their functions, ultimately resulting in cellular oxidative stress and cell death. Taken together, the rapid development of multi‐omics approaches provides a powerful toolkit for dissecting the molecular mechanisms of bacterium–algae interactions. However, omics‐derived insights must be validated by targeted molecular biology experiments, and current limitations in algal genome annotation, particularly the relatively low annotation rates among dinoflagellates (Yang et al. 2010; Lin 2024), remain a key bottleneck. Looking ahead, improvements in genome annotation methods, the development of more precise omics analyses, and rigorous cross‐disciplinary validation will help to delineate the mechanisms of bacterium–algae interactions more comprehensively and provide a more robust theoretical framework and technical support for the biocontrol of red tides.
Author Contributions
Jiaxin Wang: conceptualization, writing – original draft. Binfu Xu: writing – review and editing. Lixing Huang: conceptualization, funding acquisition, writing – review and editing.
Funding
This work was supported by the Natural Science Foundation of Fujian Province (No. 2022J02044).
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aguilera, A. , A. Distéfano , C. Jauzein , et al. 2022. “Do Photosynthetic Cells Communicate With Each Other During Cell Death? From Cyanobacteria to Vascular Plants.” Journal of Experimental Botany 73: 7219–7242.36179088 10.1093/jxb/erac 363 · doi ↗ · pubmed ↗
- 2Amin, S. A. , D. H. Green , and M. C. Hart . 2009. “Photolysis of Iron–Siderophore Chelates Promotes Bacterial–Algal Mutualism.” Proceedings of the National Academy of Sciences 106: 17071–17076. 10.1073/pnas.0905512106.PMC 276130819805106 · doi ↗ · pubmed ↗
- 3Amin, S. A. , L. R. Hmelo , H. M. Van Tol , et al. 2015. “Interaction and Signalling Between a Cosmopolitan Phytoplankton and Associated Bacteria.” Nature 522: 98–101.26017307 10.1038/nature 14488 · doi ↗ · pubmed ↗
- 4Anabtawi, H. M. , W. H. Lee , A. Al‐Anazi , M. M. Mohamed , and A. Aly Hassan . 2024. “Advancements in Biological Strategies for Controlling Harmful Algal Blooms (HA Bs).” Water 16: 224.
- 5Anderson, D. M. , E. Fensin , C. J. Gobler , et al. 2021. “Marine Harmful Algal Blooms (HA Bs) in the United States: History, Current Status and Future Trends.” Harmful Algae 102: 101975.33875183 10.1016/j.hal.2021.101975 PMC 8058451 · doi ↗ · pubmed ↗
- 6Aoki, K. , H. Kuroda , T. Setou , et al. 2019. “Exceptional Red‐Tide of Fish‐Killing Dinoflagellate Karenia Mikimotoi Promoted by Typhoon‐Induced Upwelling.” Estuarine, Coastal and Shelf Science 219: 14–23.
- 7Azam, F. , and F. Malfatti . 2007. “Microbial Structuring of Marine Ecosystems.” Nature Reviews Microbiology 5: 782–791.17853906 10.1038/nrmicro 1747 · doi ↗ · pubmed ↗
- 8Bagur, R. , and G. Hajnóczky . 2017. “Intracellular Ca 2+ Sensing: Its Role in Calcium Homeostasis and Signaling.” Molecular Cell 66: 780–788.28622523 10.1016/j.molcel.2017.05.028PMC 5657234 · doi ↗ · pubmed ↗