A Glimpse in the Metal Ion Selectivity Rules: Zn(II), Cd(II) and Co(II) Interplay With Different Protein Coordination Spheres in Determining Variable Thermal Stability and Folding Scenarios
Martina Dragone, Gaetano Caputo, Gianluca D’Abrosca, Getasew Shitaye, Ilaria Baglivo, Paolo Vincenzo Pedone, Luigi Russo, Roberto Fattorusso, Gaetano Malgieri, Carla Isernia

TL;DR
This study explores how different metal ions and amino acid arrangements in proteins affect thermal stability and folding behavior.
Contribution
The study reveals how specific coordination spheres influence folding mechanisms and metal ion selectivity in Ros87 protein variants.
Findings
CysAspHis2 coordination induces a two-state folding mechanism with Zn(II), Co(II), and Cd(II).
Wild-type Ros87 with Cys2His2 coordination forms a stable folding intermediate with Zn(II) or Co(II).
Metal ion and coordination sphere interplay determines dissociation constants and folding pathways.
Abstract
Models designed to study protein/metal ion interaction are helpful to provide insights into the rules of metal ion selectivity. In this context, the prokaryotic zinc finger family Ros/MucR offers an example of several naturally occurring homologues binding a structural zinc ion with coordination spheres characterized by different amino acid arrays. In particular, Ros87, the zinc binding domain of the protein Ros from A. tumefaciens, binds Zn(II) with a classical Cys2His2 coordination sphere, but most of its homologues show a substitution of the second cysteine by an aspartate. In this study, the binding properties to Zn(II), Co(II) and Cd(II) of the protein Ros87‐C27D, Ros87 mutant with a CysAspHis2 coordination sphere, are investigated by means of UV–vis, CD and NMR spectroscopies. Dissociation constants, structural effects and the resulting mechanisms of folding are compared with the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
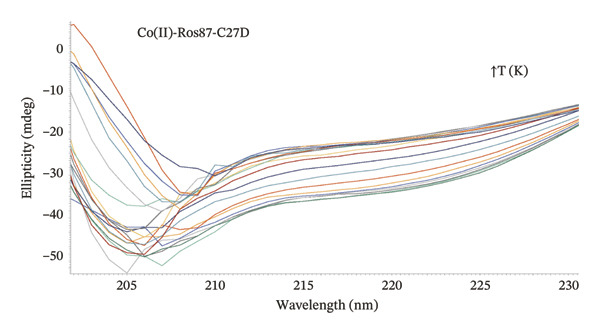 Figure 1
Figure 1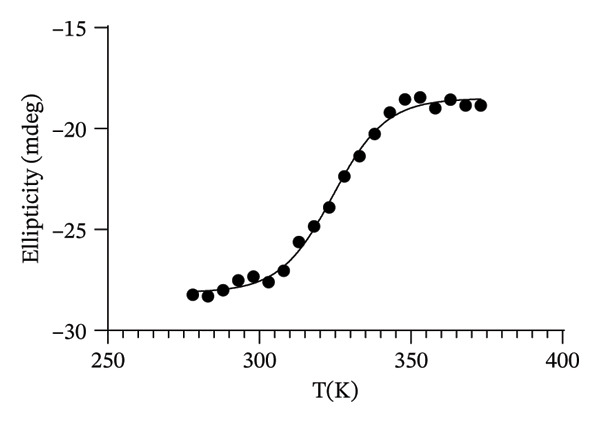 Figure 2
Figure 2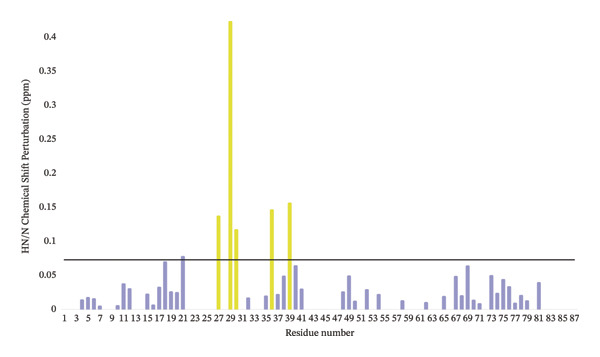 Figure 3
Figure 3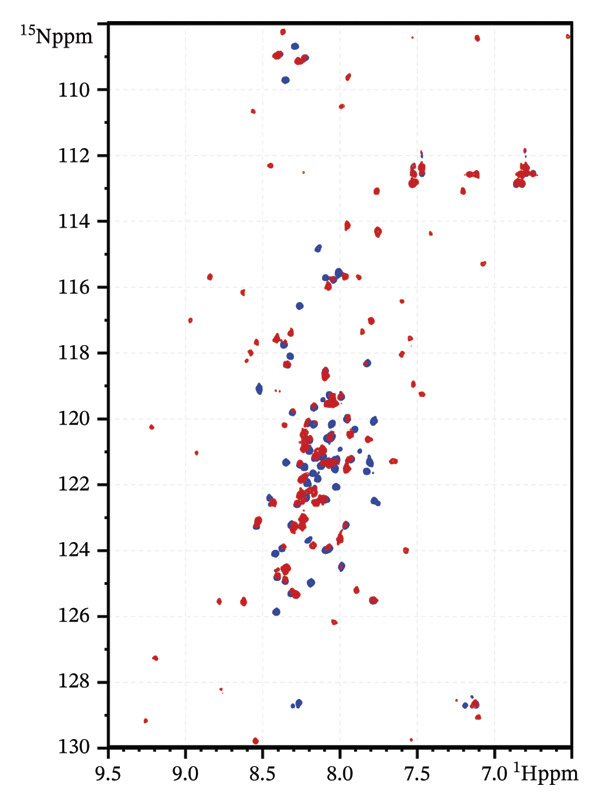 Figure 4
Figure 4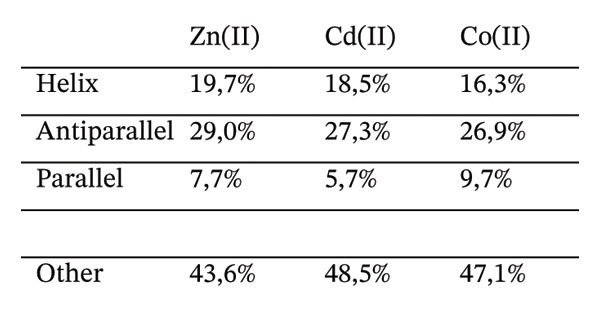 Figure 5
Figure 5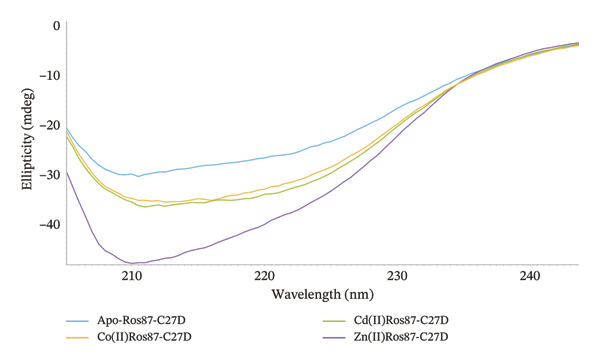 Figure 6
Figure 6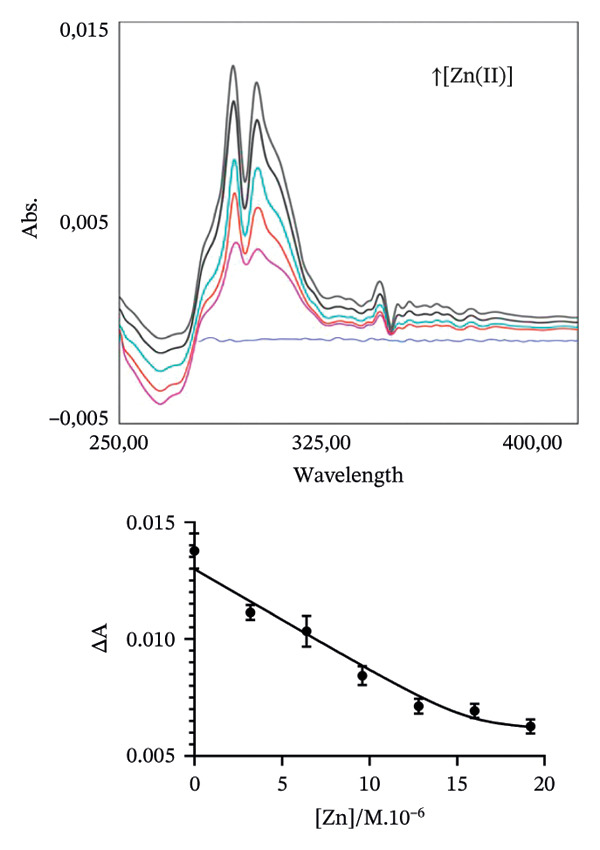 Figure 7
Figure 7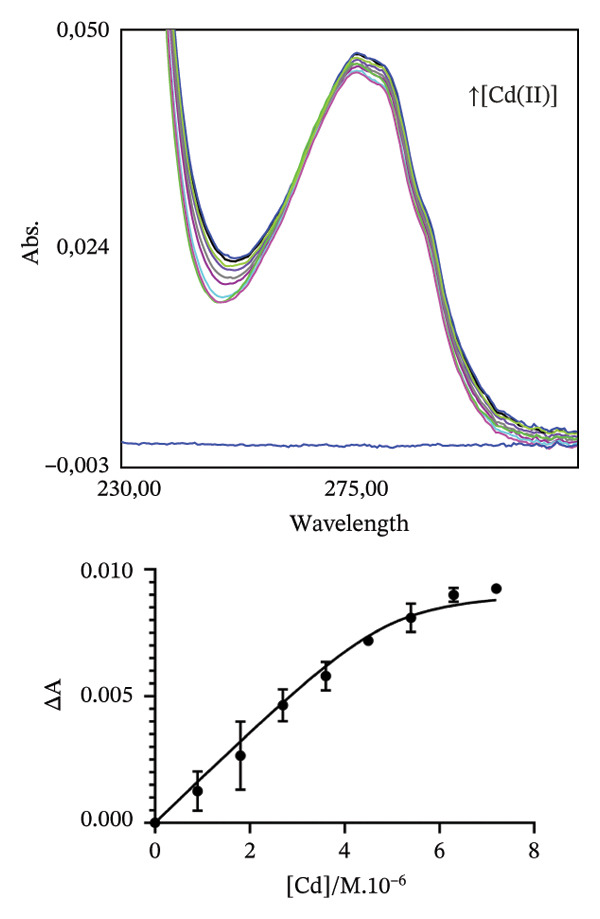 Figure 8
Figure 8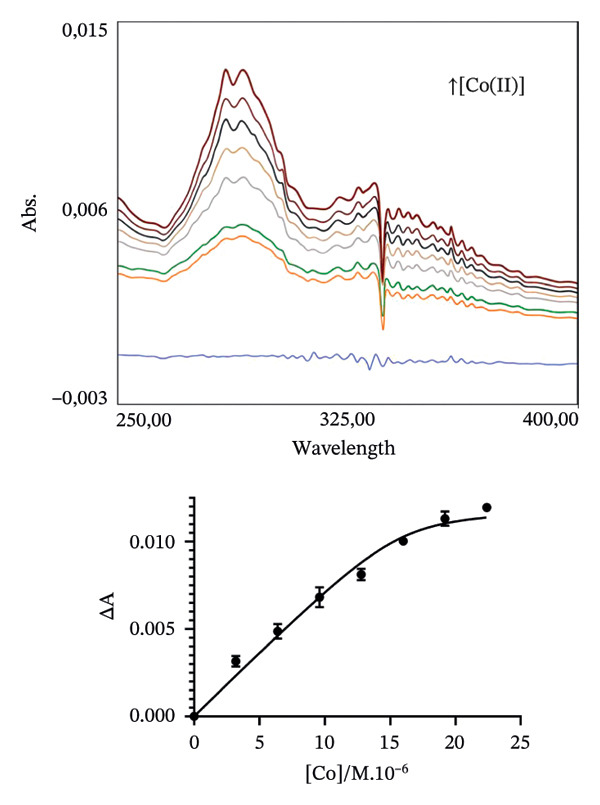 Figure 9
Figure 9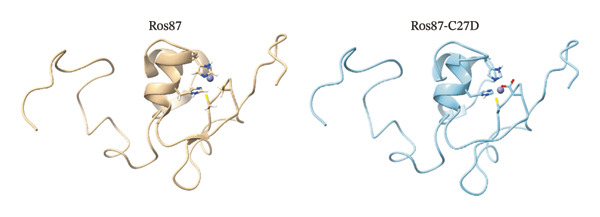 Figure 10
Figure 10 Figure 11
Figure 11 Figure 12
Figure 12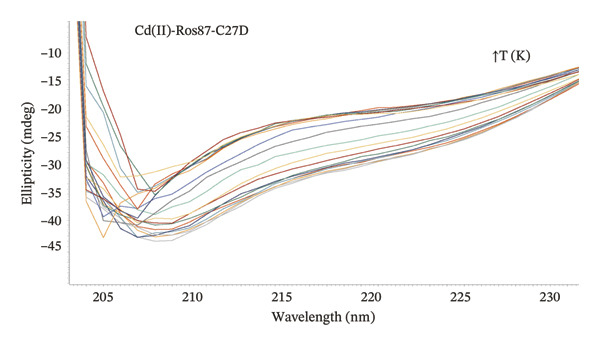 Figure 13
Figure 13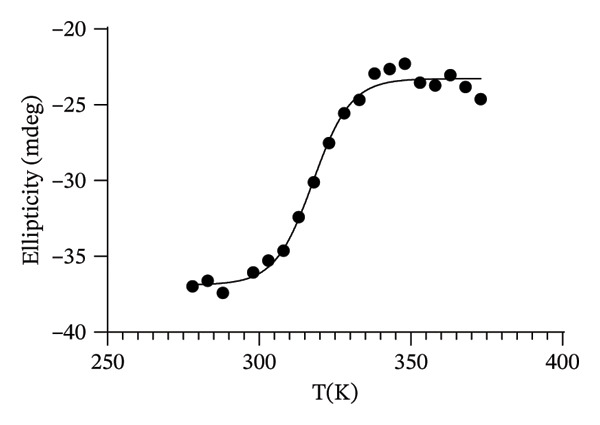 Figure 14
Figure 14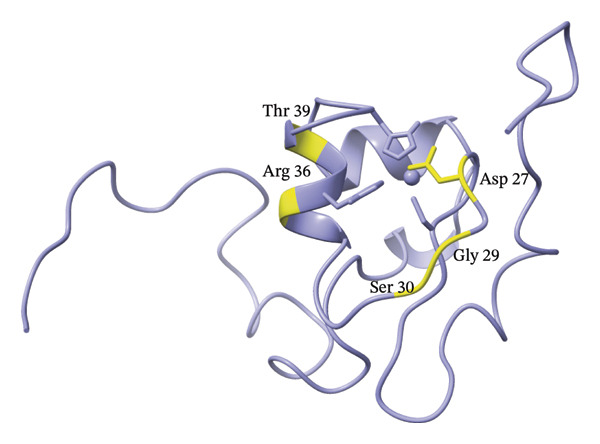 Figure 15
Figure 15| Kd∗ Co(II) (M) | Kd∗ Cd(II) (M) | Kd∗ Zn(II) (M) | |
|---|---|---|---|
| Ros87‐C27D | 4.3 (±1.4) x 10 −7 | 1.6 (±0.4) x 10−7 | 2.7 (±1.1) x 10 −9 |
| Ros87 | 5.6 (±2.0) x 10−8 | 1.98 (±0.8) x 10−8 | 5.8 (±2.0) x 10−10 |
- —Ministero dell'Università e della Ricerca10.13039/501100021856
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTrace Elements in Health · Metalloenzymes and iron-sulfur proteins · Protein Structure and Dynamics
1. Introduction
The Irving–Williams series allows the recognition of the ion properties that govern metal binding [1]. However, when it comes to proteins and their specific metalation, the rules appear more complex [2, 3]. Metal availability, type of binding side chains, binding affinity and geometrical requirements of the specific site render more obscure the establishment of rules [4–8]. The challenge faced by a metallo‐protein is to favour the binding of the “desired” metal and to elude the binding of “wrong” ions [9–12]. In fact, the binding of an “unwanted” metal can disrupt the specific structural and functional requirements of a metal binding protein [13, 14]. As a metal cofactor is required for the function of a huge amount of proteins [15], how they acquire the correct metal avoiding the toxicity of wrong metals or even the improper handling of the proper one is a fascinating field that still needs exploration [16–18].
In this regard, the accurate description of the metal binding abilities of model proteins carefully designed can give a strong contribution [19–21]. Within this context, the Ros\MucR family of zinc finger containing proteins [22, 23] offers a set of natural mutants that shows an array of metal coordinating residues perfectly suited to deepen the exploration of the rules for metal selection in proteins.
The structure of the Cys^2^His^2^ zinc binding domain (named Ros87 and obtained by deletion of the first 55 amino acids) of the Ros protein from A. tumefaciens (Uniprot Code:Q04152) [24, 25] has shown an architecture in which a β‐sheet and one α‐helix are held together by the four coordinating residues binding a structural Zn(II) ion and by a large hydrophobic core that is also formed by residues belonging to a second α‐helix that bends on this zinc binding core with an axis almost orthogonal to that of the first helix.
A substantial number of Ros homologues have been identified across various bacterial species, predominantly pathogenic to plants and mammals, and largely belonging to the proteobacteria phylum. Despite high sequence identity to the Ros protein, the zinc‐coordinating residues in these proteins are generally poorly conserved (Figure 1) [26, 27].
FIGURE 1(a) Sequence alignment of 5 Ros homologues coordination spheres identified in M. loti, residues coordinating Zn(II) [40] and substituting the coordinating ones in the zinc‐free protein Ml4 and Ml5 are reported in red. (b) Primary sequence of Ros87‐C27D; the zinc‐coordinating residues are reported in red. (c) Ribbon drawing of Ros87 (left) (PDB: 2JSP) and the mutant Ros87‐C27D (right) [27] bound to the zinc ion. The zinc ion is in blue and the metal ion coordinating residues are shown.(a)(b)(c)
In the case of Mesorhizobium loti proteins, three out of four zinc‐coordinating residues in the Ros protein exhibit poor conservation displaying various residue combinations. Specifically, the first cysteine can be substituted by a serine while all zinc‐bound proteins contain aspartic acid at the second coordinating position, leading to a CysAspHis^2^ coordination sphere. The histidine residue that could serve as the fourth zinc‐coordinating site is not always conserved. Nonetheless, the high sequence identity of these proteins to Ros implies a similar three‐dimensional structure. Notably, while the residues forming the extensive hydrophobic core of Ros are highly conserved, the lack of three of the canonical four zinc‐coordinating residues in M. loti proteins suggests a secondary role for the zinc ion in protein folding and the emergence of different zinc coordination ligands [27].
Given the rarity of structural zinc coordination by aspartate in eukaryotic zinc fingers, whether and how Ros homologues can bind zinc or other metals such as cobalt and cadmium when an aspartate occupies the second coordination position have been investigated. To this aim, a functional point mutant of Ros87 (Ros87‐C27D), which contains an aspartate replacing the second coordinating cysteine, was studied [28] by UV–vis, circular dichroism (CD) and NMR. Our findings aim at contributing to the general discussion regarding the interplay between metal ion selection in proteins, stability of the formed metal complexes, protein folding and protein functions.
2. Materials and Methods
The protein used in this article is a mutant of the full‐length Ros protein from A. tumefaciens, obtained by deletion of the first 55 amino acids. The entire Ros protein is constituted by two independent domains: N‐terminal oligomerization domain [29] and zinc‐binding DNA‐binding domain (Ros87). The amino acid numbering of the protein under study will start giving to the alanine 56 of the wild‐type protein the number 1. Thus, the mutation C27D would be C82D in the full‐length protein.
2.1. Protein Expression, Purification and Preparation of the Metal Ion–Ros87‐C27D Complexes
^15^N labelled for NMR experiments or un‐labelled proteins employed for the UV–vis and CD experiments were over‐expressed and purified as reported in D’Abrosca [28].
Concisely, transformed E. coli host strain BL21(DE3) bacteria were plated onto an LB‐agar plate, containing 100 μg/mL of ampicillin. Protein expressions that lasted 90 min were obtained by adding to the bacterial culture 1.0 mM IPTG (isopropyl‐b‐D‐1‐thiogalactopyranoside) when the absorbance of the culture measured at 600 nm was 0.6 OD. The expression of ^15^N labelled proteins was obtained simply by growing the cells at 37 °C in a minimal medium with only 0.5 g/L of ^15^NH_4_Cl as the nitrogen source. At the end of each expression, cells were harvested by centrifugation at 3750 rpm for 40 min. The obtained pellet was suspended in 20 mM Na_2_HPO_4_ buffer at pH 6.8, lysed by sonication and centrifuged (16.500 rpm to 40 min). The obtained supernatant, after a filtration with a 0.22 mm filter membrane, was then applied to a Mono S HR 5/5 cation exchange chromatography column (Amersham Bioscences, Amersham, UK). The column was previously equilibrated with phosphate buffer. The fractions containing the protein were then loaded into a HiLoad 26/60 Superdex 75 gel filtration chromatography column (Amersham Bioscences, Amersham, UK) equilibrated with 20 mM Na_2_HPO_4_ at pH 6.8, 0.2 M NaCl.
At the end of the purification procedure, the purity of the obtained samples has been assessed by means of SDS‐PAGE. The zinc ion removal from the native Zn(II)‐Ros87‐C27D was obtained by acidifying sample at pH 3 adding aliquots of HCl 0.1M; the sample was then dialysed against 10 mM Tris buffer and 150 μM TCEP at pH 3. Finally, the sample was dialysed a second time against 10 mM Tris buffer and 150 μM TCEP at pH 6.5.
As a last step, proteins were concentrated to the desired final concentration by means of an Amicon ultra‐15 centrifugal filter (Merck, Burlington, MA, USA).
2.2. UV–Vis Spectroscopy
Ros87‐C27D was complexed to the different metals (Zn^2+^, Co^2+^ or Cd^2+^), while Apo‐Ros87‐C27D indicates the protein obtained after the removal of the metal ion.
In all the titration experiments, fresh solutions of 5.0 mM metal chloride salts (ZnCl_2_, CoCl_2_ or CdCl_2_) were used and aliquots were slowly added to Apo‐Ros87‐C27D up to a final metal/protein ratio of 1.4:1.
The experiments were conducted at the lowest protein concentration possible to obtain binding isotherms and avoid metal binding at full saturation. The pH was strictly controlled and re‐measured multiple times during each step of the titration procedure. UV–vis spectra in the range 200–800 nm were recorded at 298 K by means of a Shimadzu UV‐1800 spectrophotometer. UV–vis absorption at 280 nm was used to determine the Apo‐Ros87‐C27D concentration. The molar absorption coefficient used was 9970 M^−1^ cm^−1^. In order to estimate Co(II) binding affinity, a binding curve was built upon a direct titration of Apo‐Ros87‐C27D (∼10 μM) with 5 mM CoCl_2_ solution. We followed the absorption LMCT band at 340 nm to estimate the dissociation constant of the Co(II)‐Ros87‐C27D complex [30]. For the fitting of the data, the following binding isotherm [31] was used:
where f _ p M _ indicates the fractional saturation; A 0 and A max are, respectively, the absorbance values in the absence and presence of cobalt; [M]tot is the total concentration of the Co(II) added; K_d_ ^M^ is the apparent dissociation constant of the Co(II)‐Ros87‐C27D complex; and [P]tot is the protein concentration. The same direct titration was performed also in the case of the Cd(II)‐Ros87‐C27D complex evaluating the increase in absorption at 240 nm as reported [32, 33]. Moreover, zinc affinity was estimated by reverse titrations in which the decrease of the characteristic absorption band of the complexed cobalt followed the addition of aliquots of 5 mM ZnCl_2_ to Co(II)‐Ros87‐C27D up to a Zn(II)/Co(II)‐Ros87‐C27D final ratio of 2.4:1.
Fractional saturation values were used to estimate the dissociation constant. For the fitting of the data, the following equation was used [30]:
f _ p Z n _ stands for the fractional saturation; A 0 is the absorbance value in the absence of Zn(II) and A max stands for the absorbance value in the presence of Zn(II); [Zn]tot is the final concentration of the zinc added. K_d_ ^Zn^ is the apparent dissociation constant of the Zn(II)–Ros87‐C27D complex.
Each experiment has been run in triplicate. A good R‐square for data fitting (≥ 0.98) was obtained in all the cases. The program GraphPad Prism 8.0 was used to fit the data.
2.3. CD Spectroscopy
CD spectra of the Co(II)‐Ros87‐C27D, Cd(II)‐Ros87‐C27D and Zn(II)‐Ros87‐C27D in 10 mM Tris, 150 μM TCEP at pH 6.5 were acquired by means of a JASCO J‐815 CD spectropolarimeter furnished of a Peltier temperature control. Quartz cuvettes with a 1 cm path‐length were used. Spectra were acquired on ∼8 μM samples in a wavelength range of 200–260 nm; a 1 nm data pitch, a 50 nm/min scanning speed and a 1 nm bandwidth were used. Thermal unfolding spectra were acquired at 5 K intervals in the 278–373 K temperature range. After the final spectrum, samples were cooled back to 298 K and the spectrum obtained at this temperature was compared to the spectrum recorded at the same temperature before the unfolding procedure. Data were fitted to a two‐state model, reporting the molar ellipticity at 222 nm by GraphPad Prism 8.0 software [34]. Each experiment has been run in triplicate. A good R‐square for data fitting (≥ 0.99) was obtained in all the cases. The server BestSel [35] was used for the spectra deconvolution.
2.4. NMR Spectroscopy
The NMR spectra were recorded in 10 mM Tris buffer containing 150 μM TCEP. Experiments were run at pH 6.5 and at 298 K on a Bruker Advance III HD 600 MHz equipped with a triple‐resonance Prodigy N2 cryo‐probe with a z‐axis pulse field gradient.
The NMR samples (90% H_2_O/10% ^2^H_2_O) in 3 mm tubes contained 250 μM of ^15^N‐labelled proteins into 250 μL of 10 mM Tris, 150 μM TCEP solution at pH 6.5; zinc, cobalt and cadmium fully loaded Ros87‐C27D samples were obtained after the addition of 1.5:1 ratio of metal chloride salts (ZnCl_2_, CoCl_2_ or CdCl_2_) to Apo‐Ros87‐C27D.
The experimental matrix yielding 1024 × 90 complex data points was transformed to a final matrix yielding 2048 × 1024 data points.
The following equation [36] was used to estimate HN and ^15^N chemical shift perturbations:
in which ΔHN and ΔN indicate, respectively, the differences between ^1^H and ^15^N chemical shifts of Zn(II)‐Ros87‐C27D and Cd(II)‐Ros87‐C27D. TopSpin 3.5 (Bruker) and NMRPIPE [37] were utilized to process NMR data, while CARA [38] and SPARKY [39] were employed to analyse them.
3. Results and Discussion
3.1. Metal Ion Binding Affinity
The prokaryotic zinc finger domain Ros87 exploits the typical Cys_2_His_2_ coordination sphere to bind the native structural zinc ion. The mutant protein object of this study, namely, Ros87‐C27D, uses a CysAspHis2 coordination sphere (Figure 1).
We have previously showed that this mutant protein shows the same length and three‐dimensional organization of the structural elements characterizing Ros87 [28], thus preserving all the important structural interactions [25].
To estimate, in the chosen conditions, the metal binding affinity of apoRos87‐C27D (Figure 2), which is largely unstructured as indicated by the CD data (Figure 3a), we have used a UV–vis direct titration method for Co(II) and Cd(II) complexes while a reverse titration was carried out for Zn(II)‐Ros87‐C27D. The direct titrations with Co(II) and Cd(II) show that the complexes definitively form when the metal/protein molar ratio is equal to 1.4. Monitoring the LMCT band at 340 nm and using a 1:1 model to fit the data, we obtain a lower limit for the Kd^∗^ constant of 4.3(±1.4) ^∗^10^−7^M for Co(II) while, using the band at 245 nm, the same procedure gives 1.6(±0.4) ^∗^10^−7^M for Cd(II). In the case of Zn(II), the titration of the Co(II)‐loaded protein with Zn(II) induces a progressive reduction of the Co(II) bands [41–43] that disappear after the addition of twofold excess of Zn(II) ion, indicating the substitution of the Co(II) ion with the spectroscopically inert Zn(II) ion. In this case, the estimated Kd∗ constant has a magnitude of 5.8 (±2.0) x 10^−10^M.
FIGURE 2Portion of the UV–vis spectra and fitting curves of (a) direct titration with Co(II) of Ros87‐C27D; (b) direct titration with Cd(II) of Ros87‐C27D; (c) reverse titration of Co(II) fully loaded Ros87‐C27D with Zn(II).(a)(b)(c)
FIGURE 3(a) CD spectra of Apo‐Ros87‐C27D in light blue, Zn(II)‐Ros87‐C27D in purple, Co(II)‐Ros87‐C27D in green and Cd(II)‐Ros87‐C27D in yellow. The spectra are recorded at 1.4:1 metal/apo‐protein molar ratio. (b) Comparison of the Ros87‐C27D secondary structure content in the presence of the different metal ions estimated from the CD data by the server BeStSel.(a)(b)
The obtained UV–vis spectra and the related fitting curves are shown and compared in Figure 2. The three metal ions show different Kd^∗^ constant magnitudes (Table 1), zinc being the metal ion that shows the highest affinity to the protein binding site. The data are compared in Table 1 to those already reported for the wild‐type domain [30–32], demonstrating that the introduction of the mutation in the second position of the coordination sphere affects the affinity of the protein for each metal ion, resulting in all the cases in about one order of magnitude lower.
3.2. The Secondary Structure of Co(II)‐Ros87‐C27D and Cd(II)‐Ros87‐C27D
The far‐UV CD spectrum of apo‐Ros87‐C27D in the chosen buffer indicates that Ros87‐C27D after the removal of the structural metal ion is a largely unfolded polypeptide, with a drastic decrease in the secondary structure content when compared with that of the zinc‐loaded protein. Nonetheless, some amount of the residual secondary structure is suggested by the overall shape and the intensity of the spectrum around the wavelength of 222 nm and 208 nm. This could be due to the presence of transient secondary structures and/or of the existence of a small core region that resists to the destabilizing structural effects induced by the absence of the metal ion. Otherwise, the far‐UV CD spectra of Co(II)‐Ros87‐C27D and Cd(II)‐Ros87‐C27D (Figure 3) are largely similar in shape to that of the zinc‐loaded protein, but both exhibit a similar and reproducibly decreased signal amplitude. Such a difference in signal amplitude, also in consideration of the different magnitude of the metal binding constants, is likely the result either of a minor structural perturbation of the two different metal‐bound proteins or of an increased population of unfolded protein in equilibrium with the well‐folded metal‐bound form that leads to a global destabilization of the metal complexes if compared to the zinc‐loaded protein.
Accordingly, we have used the CD data to estimate the content of the secondary structure for the three metal‐loaded proteins using the server BeStSel [35] (Figure 3(b)). The data indicate that Co(II)‐Ros87‐C27D and Cd(II)‐Ros87‐C27D structures have a content of secondary structure similar to that of Zn(II)‐Ros87‐C27D.
3.3. Cd(II)‐Ros87‐C27D
To further evaluate the structural effects of the zinc ion replacement with cadmium, 2D ^1^H‐^15^N NMR HSQC spectrum was acquired for Cd(II)‐Ros87‐C27D and compared with that of the apo form and of Zn(II)‐Ros87‐C27D (Figure 4). Cd(II)‐Ros87‐C27D spectrum clearly shows a higher degree of resonance dispersion if compared to the spectrum of apo‐Ros87‐C27D, indicating that upon Cd(II) addition, like the Zn(II)‐loaded protein, Ros87‐C27D populates a folded conformation with well‐formed elements of α‐helix and β‐sheet structure equivalent to that of the wild‐type protein. Moreover, many of the resonances in the spectra of each of Cd(II)‐Ros87‐C27D and Zn(II)‐Ros87‐C27D are superposable (Figure 4), indicating that large regions of the polypeptides adopt a highly similar structure in both proteins. Nonetheless, to identify the residues experiencing the greatest metal substitution‐induced perturbations, we estimated the weighted average of the 1 H and 15 N chemical shift change for each resonance in the spectra and produced a graph reporting the data as a function of the residue number (Figure 4(c)). Residues with chemical shift variations higher than average plus one standard deviation were then colour‐mapped onto the structure of Zn(II)‐Ros87‐C27D [28]. Clearly, the inspection of both figures shows that significant chemical shift changes are evident for those resonances arising from the region surrounding the metal ion coordination sphere: the largest changes are obtained for the amino acids Asp27, Gly29, Ser30, Arg36 and Thr39, which are located near the zinc coordination sphere, thus reflecting a different chemical environment. Together, these results confirm that the substitution of the Zn(II) ion with Cd(II) allows the protein to achieve the same fold but at the same time induces a substantial structural reorganization of the coordination sphere of the protein accompanied by only a minor structural rearrangement of the protein hydrophobic core.
FIGURE 4(a) Overlaid ^1^H‐^15^N HSQC spectra for Apo‐Ros87‐C27D in blue and Cd(II)‐Ros87‐C27D in red. (b) Weighted average of the chemical shift perturbations between the ^1^H^15^N HSQC of Cd(II)‐Ros87‐C27D and the ^1^H^15^N HSQC of Zn(II)‐Ros87‐C27D; the black line represents the mean value plus the standard deviation. (c) Chemical shift perturbations estimated mapped onto the Zn(II)‐Ros87‐C27D 3D structure.(a)(b)(c)
3.4. Thermal Unfolding
In order to investigate the effect of the structural metal ion changes on the protein stability, a thermal unfolding characterization of Co(II)‐Ros87‐C27D and Cd(II)‐Ros87‐C27D has been carried out using CD spectroscopy. Figures 5(a), 5(b) report the CD spectra recorded for both proteins as a function of temperature: data show how the raise of temperature results in all the cases in a progressive loss of secondary structure content. The Ɵ value at 222nm was followed at regular intervals of 5K in a range from 278 to 372 K. The data have been fitted using a two‐state model obtaining a melting temperature of 317.8 (±1.9) K and 324.3 (±1.7) K, respectively (Figures 5(c), 5(d)).
FIGURE 5(a) CD spectra of Co(II)‐Ros87‐C27D recorded in the 278–372 K temperature range. (b) CD spectra of Cd(II)‐Ros87‐C27D recorded in the 278–372 K temperature range. (c) Unfolding curves of Co(II)‐Ros87‐C27D monitored at 222 nm and fitted with a two‐state model. Error bars are included in the symbols plotted. (d) Unfolding curves of Cd(II)‐Ros87‐C27D monitored at 222 nm and fitted with a two‐state model. Error bars are included in the symbols plotted.(a)(b)(c)(d)
As already found for the Zn(II)‐Ros87‐C27D protein [28, 45], Zn(II)‐Ros87‐C27D remains the most stable structure with an obtained melting temperature of 338K (±0.2) [45], while Cd(II)‐Ros87‐C27D appears more stable (324 K) than Co(II)‐Ros87‐C27D for which a temperature of 318K was obtained.
This trend reflects the magnitude of the metal ion binding affinity reported above.
Interestingly, the same trend in the metal‐loaded Ros87‐C27D stabilities was obtained for Ros87: Zn(II)‐Ros87 ca. 365K; Cd(II)‐Ros87 ca. 348K; Co(II)‐Ros87 ca. 338K.
However, Zn(II)‐Ros87 folding/unfolding mechanism has been shown to be a two‐transition process [46]: in the first transition, the metal ion recruitment leads to the formation of a metal binding intermediate that, in the second transition, converts to the native state through a downhill barrierless mechanism [47–50]. In particular, the downhill reversible transition occurs in the temperature range between 300 and 350 K, while the irreversible transition due to the loss of the metal ion happens in the range of 370–390 K [46]. In this protein, the substitution with both metals implied a change in this thermodynamic scenario, with Cd(II)‐Ros87 showing the most dramatic difference: this protein folds/unfolds through a classical two‐state model, and no stable intermediate states are detected during the structural transition between the folded and the unfolded forms. Also, Co(II)‐Ros87 shows differences in terms of unfolding mechanism when compared to Zn(II)‐Ros87 but remains characterized by two transitions [51]. Interestingly, the mutation of the second zinc‐coordinating cysteine to aspartate (thus transforming Zn(II)‐Ros87 in Zn(II)‐Ros87‐C27D) has been previously evaluated in terms of influence on the mechanism of folding, showing that this simple mutation allows the structure and the metal ion centre of the protein to collapse in a single event, i.e., the mutant protein shows a two‐state folding mechanism.
The same two‐state mechanism of folding is here demonstrated to characterize Co(II)‐Ros87‐C27D and Cd(II)‐Ros87‐C27D. Altogether, this behaviour suggests that the interplay between the metal ion characteristics and the organization of metal ion coordination sphere is determinant in the folding process.
4. Conclusions
Protein structures, enzymes and also many therapeutic and diagnostic tools all rely on metal ion selectivity to properly function. Selectivity requires the optimized affinity for one metal ion while the affinity for all the others is reduced [52]. The search for ligands capable of coordinating selectively specific metal ions is quite difficult [53–55]. Electronic and steric effects, entropy and solvent dependencies are all terms that contribute to the complexity in the prediction of metal ion discrimination [56–58]. When it comes to proteins, metal concentration within the cellular compartments where the protein performs its function and the presence of metallo‐chaperones are also important factors [59–61]. On top of this, a considered metallo‐protein can often display an affinity for multiple metal ions, further complicating the determination of the relationship between protein structure and metal binding selectivity [62, 63]. In this framework, the study of simplified models of metal binding proteins can offer hints to guide researchers through the complexity of the relationship between metal ions, protein structures along with their folding pathway and protein functions [20, 64, 65]. Our group has been long involved in the study of the structure–function relationship of zinc finger proteins. Within these special metal binding domains, the prokaryotic zinc finger family offered us a paradigmatic example of metal binding sites in which the same structural architecture can functionally fold exploiting different sets of metal coordinating amino acids or even substitute the stabilizing role of the metal cofactor with a network of hydrogen bonds and hydrophobic interactions.
The same proteins have been used to study the effects on the structure and folding mechanism of the native metal ion replacement. In particular, we have earlier shown that, although with different affinities, the prokaryotic zinc finger domain Ros87 binds its native zinc or other metal ions, such as cadmium (with a larger ionic radius) and cobalt (with a smaller radius), still remaining capable of folding in a functional structure [30, 32]. We have also demonstrated that the substitution of the native Zn(II) with Cd(II) or Co(II) has a profound effect on the unfolding mechanism of Ros87 [30, 51].
In the present work, we have investigated whether the differences of binding affinities for the same metals due to a different coordination sphere influence the structural tolerance and the folding mechanism of Ros87 iso‐structural mutant, Ros87‐C27D. Altogether, our study reports evidences on how the minor structural reorganization of the metal binding core around Co(II) and Cd(II) also allows the mutant protein to retain the same globular architecture of the zinc‐loaded protein, demonstrating also the mutant protein to be able to structurally tolerate different ionic radii.
However, differently from the wild‐type protein (the Cys_2_His_2_ Ros87), Ros87‐C27D (CysAspHis_2_) shows a two‐state folding/unfolding mechanism in the presence of each of the studied metal ions. Interestingly, both proteins, when cadmium‐loaded, show a reversible two‐state mechanism of folding. In the case of Zn(II) or Co(II), the protein bearing the cysteine in the second position shows a two‐step folding process with the formation of a metal binding intermediate while the folding mechanism of the mutant protein is a reversible two state. These findings underline that the interplay between the different metal ions, with their different electronic structures and different ionic radii, and the coordinating amino acid patterns is determinant in defining the different dissociation constants and the folding pathway of a given protein.
The thiol group confers to cysteines the ability to strongly interact with the different metal ions but cysteines are quite delicate residue to handle for a given protein. Indeed, free cysteines frequently form aberrant disulphide bonds triggering misfolding and aggregation [66, 67]. The fact that the organization of the metal ion centre has such a dramatic effect in determining the folding scenario allows us to infer that, for a given coordination sphere, the evolutionary advantage in recruiting and retaining a specific metal ion has been driven by the most diverse chemical conditions that might have required the formation or not of stable metal binding intermediates. Our data indicate that, at least in the systems reported in the present study, the presence of the two cysteines requires the formation of a metal containing structural intermediate in which zinc or cobalt protects these residues from the formation of disulphide bonds. The intermediate is no longer required when the metal cofactor is cadmium with this known preferential binding to thiols. The mutation of the second coordinating cysteine to aspartic acid cancels the necessity of a structural intermediate resulting in all the cases in a two‐state scenario.
Concluding, it is important to study the same protein structure coordinating different metal ions or to study the same metallo‐protein varying its coordination sphere in order to offer to the scientific community a body of data helpful to fine‐tune, in both computational and experimental fields, the rules of metal ion selectivity.
Funding
This work was supported by the Ministero dell’Università e della Ricerca, 2022MBK24T P2022AW2H9 2022JXSA9C.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Irving H. and Williams R. J. P. , Order of stability of metal complexes, Nature. (1948) 162, no. 4123, 746–747.
- 2Haas K. L. and Franz K. J. , Application of metal coordination chemistry to explore and manipulate cell biology, Chem Rev. (2009) 109, no. 10, 4921–4960.19715312 10.1021/cr 900134 a PMC 2761982 · doi ↗ · pubmed ↗
- 3Choi T. S. and Tezcan F. A. , Overcoming universal restrictions on metal selectivity by protein design, Nature. (2022) 603, no. 7901, 522–527.35236987 10.1038/s 41586-022-04469-8PMC 9157509 · doi ↗ · pubmed ↗
- 4Young T. R. and Xiao Z. , Principles and practice of determining metal-protein affinities, Biochem J. (2021) 478, no. 5, 1085–1116.33710331 10.1042/BCJ 20200838 PMC 7959690 · doi ↗ · pubmed ↗
- 5Holm R. H. , Kennepohl P. , and Solomon E. I. , Structural and functional aspects of metal sites in biology, Chem Rev. (1996) 96, no. 7, 2239–2314.11848828 10.1021/cr 9500390 · doi ↗ · pubmed ↗
- 6Dokmanić I. , Sikić M. , and Tomić S. , Metals in proteins: Correlation between the metal-ion type, coordination number and the amino-acid residues involved in the coordination, Acta Crystallogr D Biol Crystallogr. (2008) 64, no. Pt 3, 257–263.18323620 10.1107/S 090744490706595 X · doi ↗ · pubmed ↗
- 7Noroozi Tiyoula F. , Vafaee Sharbaf F. , Rahimian K. , Gholami M. , Zabihi M. R. , Salehi Z. et al., Artificial intelligence in metalloprotein binding site prediction: A systematic review bridging bioinformatics and biotechnology, Int J Biol Macromol. (2025) 323, no. 2.10.1016/j.ijbiomac.2025.14666640885350 · doi ↗ · pubmed ↗
- 8Essien C. , Wang N. , Yu Y. , Alqarghuli S. , Qin Y. , Manshour N. et al., Predicting the location of coordinated metal ion-ligand binding sites using geometry-aware graph neural networks, Computational and Structural Biotechnology Journal. (2025) 27, 137–148.39840139 10.1016/j.csbj.2024.12.016PMC 11750443 · doi ↗ · pubmed ↗