Pressure‐dependent adaptation strategies implied by the dissimilatory iron reducer Orenia metallireducens Z6
Shuyi Li, Jiahao Pei, Jiasong Fang, Rulong Liu, Yuli Wei, Xianyu Huang, Guang Yang, Min Liu, Qin Lin, Robert R. Sanford, Hongbo Shao, Yongguang Jiang, Yidan Hu, Zhou Jiang, Qi Feng, Yu He, Chenxi Zhang, Yizhou Fan, Yiran Dong, Liang Shi

TL;DR
This paper studies how a deep subsurface bacterium adapts to high pressure by using iron minerals to generate energy and survive.
Contribution
The study reveals novel pressure adaptation strategies involving carbon flux redirection and iron mineral utilization in a piezotolerant bacterium.
Findings
At 40 MPa, Orenia metallireducens redirects carbon flux toward energy generation and biosynthesis.
Iron minerals act as an electron sink to support survival under high hydrostatic pressure.
Both universal and pressure-dependent strategies help the organism withstand varying pressures.
Abstract
Tolerance of high hydrostatic pressure (HHP) is the hallmark of deep subsurface microorganisms, while its mechanisms remain under‐investigated. This study explores HHP adaptation in the piezotolerant bacterium Orenia metallireducens across its near‐full pressure range (0.1–40 MPa). At inhibitory pressure (40 MPa), the organism redirected carbon flux toward more favorable energy generation and biosynthesis using ferric mineral as the “electron sink.” Furthermore, both universal and pressure‐dependent strategies enabled the organism to withstand varying pressures. These findings highlight the role of iron minerals in microbial HHP adaptation and reveal novel survival strategies, advancing our understanding of deep‐life evolution and biogeochemical impacts.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
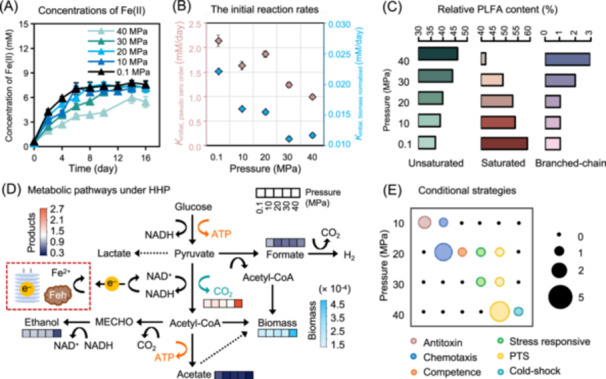 Figure 1
Figure 1- —National Natural Science Foundation of China10.13039/501100001809
- —Hadal Science and Technology Research Center at Shanghai Ocean University
- —Fundamental Research Funds for the Chinese Central Government via China University of Geosciences (Wuhan)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Fuel Cells and Bioremediation · Microbial Inactivation Methods · Bacterial biofilms and quorum sensing
The deep subsurface, one of the Earth's largest microbial habitat, is characterized by high hydrostatic pressure (HHP) as an essential environmental stressor1, 2. HHP profoundly influences microbial life, affecting community structure, metabolic flexibility, and evolutionary adaptation. Research has revealed that HHP affects virtually all aspects of cellular physiology, including transcription, translation, membrane composition, multimeric protein assemblages, protein structure, and cellular motility3. While microbial adaptation to HHP is crucial for global biogeochemical cycles4, and microorganisms use strategies such as synthesizing osmolytes (compatible solutes) and activating antioxidant defenses5, 6, 7, research has historically focused on piezophiles, leaving common and ecologically important piezotolerant organisms underinvestigated8, 9. Among different microbial reactions, thermodynamic models indicated that microbial iron reduction was energetically more favorable under HHP compared to other electron‐accepting processes10. However, their specific adaptations to a pressure gradient and the role of ubiquitous iron minerals in this process remain poorly understood5. To address these gaps, we investigated the piezotolerant iron reducer Orenia metallireducens strain Z6, isolated from a 2.01 km‐deep terrestrial reservoir11, 12, across a hydrostatic pressure gradient from 0.1 to 40 MPa (Figure 1A). Our findings reveal a multi‐level, pressure‐dependent adaptation strategy that integrates physiological, metabolic, membrane, and genomic responses.
Physiologically, strain Z6 showed stable iron‐reducing activity at 0.1–30 MPa but was significantly inhibited at 40 MPa (Figure 1A). In contrast, no biological iron reduction was observed in any of the abiotic controls across all pressure conditions, suggesting that iron reduction was mediated by enzymatic reactions (Figure S1). While the final Fe(II) production was reduced by approximately 25% at 40 MPa, the initial Fe(III) reduction rates showed a more pronounced decline, decreasing from 2.14 ± 0.12 mM/day at 0.1 MPa to 0.99 ± 0.04 mM/day at 40 MPa (Figure 1B). Protein analysis showed that biomass production peaked at 20 MPa (Figure S2), a pressure that closely aligned with its native habitat (~17 MPa)12. The optimal growth at its native pressure suggests a nuanced adaptation. While the rate of iron reduction per cell (biomass‐normalized rate) decreased at 20 MPa compared to 0.1 MPa, the organism compensated by accumulating significantly more biomass to maintain overall metabolic function (Figures 1B and S2).
A key physiological adaptation to HHP is the maintenance of cell membrane fluidity. Strain Z6 actively remodeled its membrane lipids in response to pressure (Figure 1C), a process known as homeoviscous and homeophasic adaptation3. Membrane analyses revealed phospholipid fatty acids (PLFAs) with 13–18 carbons in strain Z6, among which 15‐ and 17‐carbon species predominated (Table S1). As pressure increased, the proportions of unsaturated and branched‐chain fatty acids significantly increased, while the saturated fatty acid content decreased (Figure 1C and Table S2). Unsaturated and branched‐chain lipids have lower melting points and increase membrane fluidity, counteracting the pressure‐induced gelling or rigidity of the lipid bilayer, thereby maintaining essential cellular functions13, 14. Compared to their straight‐chain counterparts, branched‐chain PLFAs reduce melting temperatures and prevent the formation of crystalline structures between acyl chains, thereby further enhancing membrane fluidity under pressure3. This aligns with a previous study showing that the monounsaturated‐to‐polyunsaturated PLFA ratios for marine microorganisms correlate positively with their isolation depth13. Similarly, Photobacterium profundum SS9 requires monounsaturated fatty acids for growth at high pressure and low temperature14.
Critically, a metabolic shift under the inhibitory pressure (40 MPa) was observed (Figure 1D). Specifically, glucose consumption slowed, and the final concentrations of formate and acetate decreased significantly. While ethanol yield decreased, CO_2_ production and the final Fe(II)/glucose ratio increased significantly (Figures 1D and S3; Table S2). This indicates a pressure‐induced redirection of carbon and electron flux. Metabolic reconstruction revealed that strain Z6 decomposed glucose via glycolysis and pyruvate metabolism11. These pathways include two steps in glycolysis and one step in the downstream pyruvate metabolism (acetyl‐CoA to acetate) that generate ATP. Beyond energy production, pyruvate metabolism also regulates the NADH/NAD^+^ balance. Pyruvate‐to‐ethanol conversion consumes NADH to form NAD^+^, while pyruvate decarboxylation generates NADH along with the formation of C1 compounds (CO_2_ or formate) and acetyl‐CoA15. Our observed carbon flux redirection might be due to pressure‐induced inhibition of key enzymes such as alcohol dehydrogenase16. This hypothesis aligns with the earlier findings on Clostridium paradoxum 16. For this organism, elevated pressure attenuates ethanol production kinetics by reducing enzymatic activity, which is attributed to a pressure‐induced conformational state change characterized by lower substrate affinity for NAD⁺ and restricted hydride transfer16. Thus, a metabolic shift occurs, moving away from ethanol formation and toward pyruvate decarboxylation. This shift provides key advantages. It generates more acetyl‐CoA, which is a key biosynthetic precursor15. It also produces more reduced electron carriers (e.g., NADH), which then support enhanced ferrihydrite reduction by strain Z6. This mechanism highlights the central role of Fe(III) minerals as an “electron sink.” It reframes the conventional theory, which often regards ferric minerals in fermentation as merely incidental sinks. In contrast, our work suggests that they are active facilitators that modify electron and carbon fluxes, ultimately enhancing microbial growth under inhibitory HHP. While thermodynamic modeling predicts increased favorability of microbial iron reduction under HHP10, our calculations show that the changes in Gibbs free energy (ΔG) across our experimental pressure range are not significant (Table S2). Instead, the observed thermodynamic difference between the inhibitory pressure (40 MPa) and the relatively lower pressures primarily relates to the altered stoichiometry between glucose decomposition and iron reduction. By accepting excess electrons, the amended ferrihydrite actively facilitates this metabolic rerouting, allowing strain Z6 to optimize energy production and growth yield even under inhibitory pressure. This adaptation makes the overall metabolism thermodynamically more favorable at 40 MPa (ΔG = −361.12 kJ) than that under lower pressures (ΔG = −343.51 to −303.15 kJ) (Table S2), explaining how comparable biomass yields were maintained despite physiological stress.
Sequencing of the cDNA libraries for the pressure‐adapted cultures yielded 2.00–2.43 Gb of high‐quality reads per sample (Table S3). Compared to the gene expression under 0.1 MPa controls, 463, 1794, 2082, and 2059 genes were identified as differentially expressed genes (DEGs) (Figure S4). Transcriptomic analysis revealed the genetic basis for these integrated adaptations (Figures 1E and S5). We identified general pressure adaptation strategies based on the genes upregulated across all high‐pressure conditions (10–40 MPa) (Figure S5 and Table S4). These included universal stress response genes for: (i) antioxidant defense (e.g., the grx gene encoding glutaredoxin). Upregulation of antioxidant systems represents a universal bacterial defense strategy. HHP is known to induce oxidative stress; thus, activating these pathways is a critical strategy to maintain cellular integrity, similar to the upregulation of gpx and katE by Shewanella piezotolerans WP3 under HHP and cold stress17. (ii) DNA/protein repair. The gene encoding the LexA repressor, which represses SOS response genes, was upregulated. In contrast, recA, which is essential for homologous DNA repair, was downregulated. This may result from a negative feedback loop in the bacterial SOS response system, ensuring an appropriate repair response while preventing potential harm from an overactive one. For example, the clpX and clpP genes encoding ATP‐dependent Clp proteases critical for protein quality control were downregulated, which likely plays a key role in protein degradation, stress response, and cellular physiology regulation, perhaps as a mechanism to conserve cellular energy under stress. (iii) Ion transport. Upregulation of afuB and fbpB encoding putative ferric transporters highlights the critical challenge of iron homeostasis under pressure. The organism must acquire essential iron while managing the risk of oxidative stress from free iron. Thus, upregulation of these specific transporters suggests a fine‐tuned mechanism to balance metabolic need against potential toxicity. (iv) Compatible solute biosynthesis. Genes such as gltB/D, proW/X, and proX, encoding the proteins for glutamate, glycine, and betaine transport, respectively, were upregulated. These allow uptake or production of the organic piezolytes to stabilize proteins and membranes against pressure stress6. In addition, upregulation of the glutamate synthase genes (gltB/D) is particularly significant, as glutamate is a key piezolyte observed in many piezophiles (e.g., Desulfovibrio spp.)7.
More significantly, conditional pressure adaptation strategies on the specific pressure regime were identified (Figure 1E and Table S5). At 10–30 MPa, where active microbial growth occurred, strain Z6 upregulated the genes related to chemotaxis (e.g., histidine kinase) and natural competence18. The flagellar system is usually important for piezophilic growth, serving a critical function for nutrient acquisition and predator avoidance3. Nearly all the genes associated with flagellar synthesis and regulation in strain Z6, however, were downregulated at elevated pressures (Table S6). Thus, the upregulation of chemotaxis proteins under elevated pressures by strain Z6 suggests an alternative strategy for fine‐tuning environmental sensing. Interestingly, the genes related to the TonB system, which powers outer membrane transport at the cost of the proton motive force, were downregulated at 10–40 MPa compared to that at standard pressure (Table S4). This downregulation implies an active strategy to reallocate energy toward detecting favorable microniches for activating natural competence, potentially enabling nutrient recycling from lysed cells as an energetically efficient strategy. In contrast, at the inhibitory 40 MPa, a different suite of genes (e.g., those encoding the phosphotransferase system (PTS) and cold‐shock proteins) was overexpressed, indicating a shift to a “survival” posture (Figure 1E and Table S5). PTS is a central regulator for carbon metabolism and stress responses. Studies suggest that PTS may modulate oxidative stress tolerance or exert genetic regulation in non‐ or slow‐growing cells exposed to lethal stress19. In this study, it may function as a high‐affinity scavenging system or as a key sensor in a broader stress‐response cascade, helping the cells withstand inhibitory pressure while in a slow‐growing state. Meanwhile, the cold‐shock protein response confirms the known mechanistic linkage between adaptation to HHP and cold stress19.
The ecological relevance of these findings is underscored by the global distribution of the genus Orenia. A survey of isolated strains and metagenomic data showed their presence in diverse habitats ranging from surface hypersaline lakes to deep subsurface (Figure S6 and Table S7). These environments span the pressure range, aligning with that investigated in this study. This strongly suggests that the multi‐level, Fe‐dependent adaptive strategies observed in strain Z6 are not unique, but rather represent fundamental and ecologically significant survival mechanisms for this widespread genus across the shallow to deep biosphere.
In conclusion, O. metallireducens strain Z6 uses multi‐level and pressure‐dependent adaptation strategies under HHP conditions. The strategies are conditional: it maintains active metabolism (0.1–30 MPa) via chemotaxis and membrane remodeling, but shifts to a conservative “survival mode” at inhibitory pressure (40 MPa). This shift involves actively rerouting carbon flux away from ethanol and toward CO_2_/biomass to optimize energy generation. Crucially, we identify ubiquitous Fe(III) minerals not as passive acceptors, but as active “electron sinks” essential for this metabolic flexibility and enhanced biomass production under stress. Given the global distribution of the Orenia genus (Figure S6), this Fe‐dependent adaptation likely represents a fundamentally and ecologically significant survival strategy for the deep biosphere20.
ETHICS STATEMENT
This article does not contain any study with human participants or animals.
Supporting information
Supporting information.
Supporting information.
Supporting information.
Supporting information.
Supporting information.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Flemming HC , Wuertz S . Bacteria and archaea on Earth and their abundance in biofilms. Nat Rev Microbiol. 2019;17:247–260.30760902 10.1038/s 41579-019-0158-9 · doi ↗ · pubmed ↗
- 2Magnabosco C , Lin LH , Dong H , Bomberg M , Ghiorse W , Stan‐Lotter H , et al. The biomass and biodiversity of the continental subsurface. Nat Geosci. 2018;11:707–717.
- 3Oger PM , Jebbar M . The many ways of coping with pressure. Res Microbiol. 2010;161:799–809.21035541 10.1016/j.resmic.2010.09.017 · doi ↗ · pubmed ↗
- 4Yang N , Lv Y , Ji M , Wu S , Zhang Y . High hydrostatic pressure stimulates microbial nitrate reduction in hadal trench sediments under oxic conditions. Nat Commun. 2024;15:2473.38503798 10.1038/s 41467-024-46897-2PMC 10951307 · doi ↗ · pubmed ↗
- 5Zhong M , Li Y , Deng L , Fang J , Yu X . Insight into the adaptation mechanisms of high hydrostatic pressure in physiology and metabolism of hadal fungi from the deepest ocean sediment. m Systems. 2024;9:0108523.10.1128/msystems.01085-23PMC 1080494138117068 · doi ↗ · pubmed ↗
- 6Yancey PH . Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. J Exp Biol. 2005;208:2819–2830.16043587 10.1242/jeb.01730 · doi ↗ · pubmed ↗
- 7Amrani A , van Helden J , Bergon A , Aouane A , Ben Hania W , Tamburini C , et al. Deciphering the adaptation strategies of Desulfovibrio piezophilus to hydrostatic pressure through metabolic and transcriptional analyses. Environ Microbiol Rep. 2016;8:520–526.27264199 10.1111/1758-2229.12427 · doi ↗ · pubmed ↗
- 8Dutta A , Peoples LM , Gupta A , Bartlett DH , Sar P . Exploring the piezotolerant/piezophilic microbial community and genomic basis of piezotolerance within the deep subsurface Deccan traps. Extremophiles. 2019;23:421–433.31049708 10.1007/s 00792-019-01094-8 · doi ↗ · pubmed ↗