Maize ZmbZIP92 transcription factor positively regulates drought tolerance in Arabidopsis
Zukuan Liu, Jian Shang, Min Liu, Jietian Kong, Aiya Huang, Yang Zhao, Qing Ma, Wei Dai

TL;DR
This study identifies a maize gene, ZmbZIP92, that helps plants better tolerate drought by influencing stress response pathways.
Contribution
ZmbZIP92 is a novel maize bZIP transcription factor that positively regulates drought tolerance in Arabidopsis.
Findings
ZmbZIP92 is a nuclear protein that binds to the G-box element but lacks self-activation activity in yeast.
Overexpression of ZmbZIP92 in Arabidopsis improves drought tolerance and ABA sensitivity.
ZmbZIP92 influences ABA signaling and stress response pathways in plants.
Abstract
Plants are susceptible to various environmental stresses, but basic leucine zipper (bZIP) transcription factors play a key role in regulating stress responses. In this study, a drought response-related candidate gene (ZmbZIP92) was cloned from maize (Zea mays L.) following a comparative genomic analysis. This gene is highly homologous to the rice (Oryza sativa L.) gene OsbZIP62, exhibits tissue-specific expression patterns, and is significantly induced by drought, high salinity, and abscisic acid (ABA) treatments. Subcellular localization revealed that ZmbZIP92 is a nuclear protein. Additionally, yeast-based assays of ZmbZIP92 detected a lack of transcriptional self-activation. Dual-luciferase reporter assays demonstrated that ZmbZIP92 binds specifically to the G-box (CACGTG) cis-element. Overexpressing ZmbZIP92 in Arabidopsis thaliana significantly promoted root elongation, enhanced…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
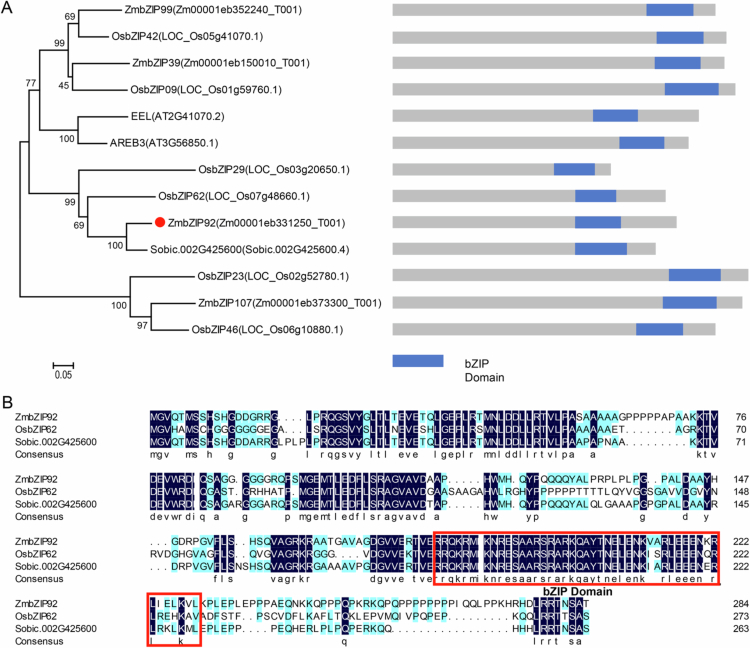 Figure 1
Figure 1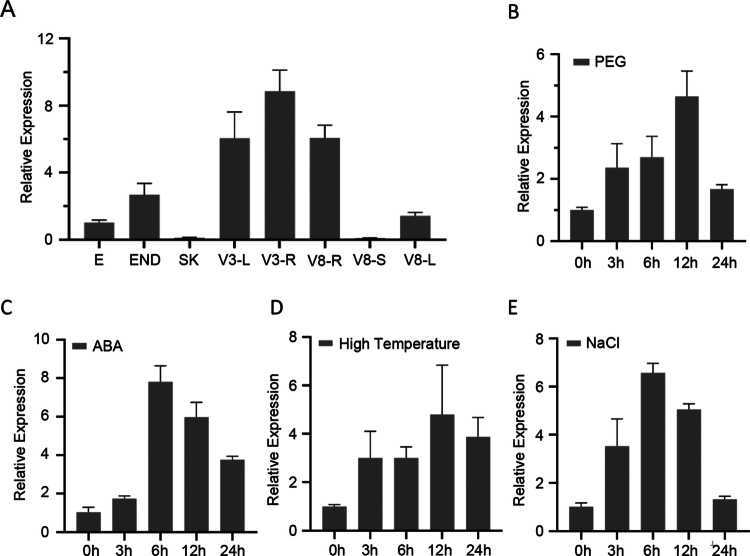 Figure 2
Figure 2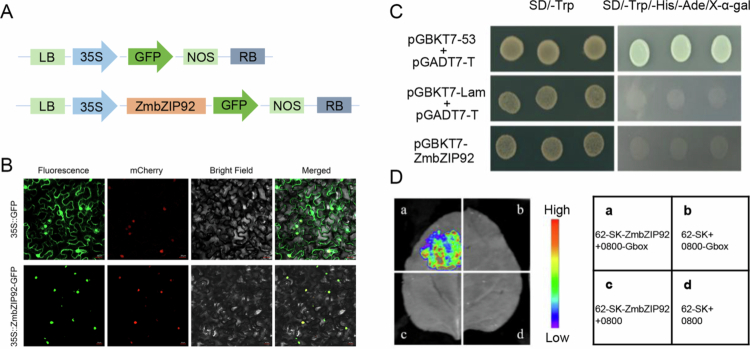 Figure 3
Figure 3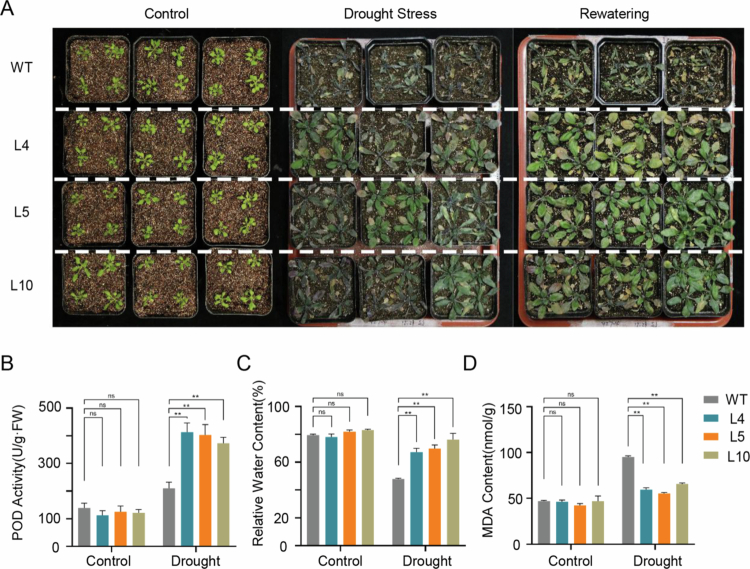 Figure 4
Figure 4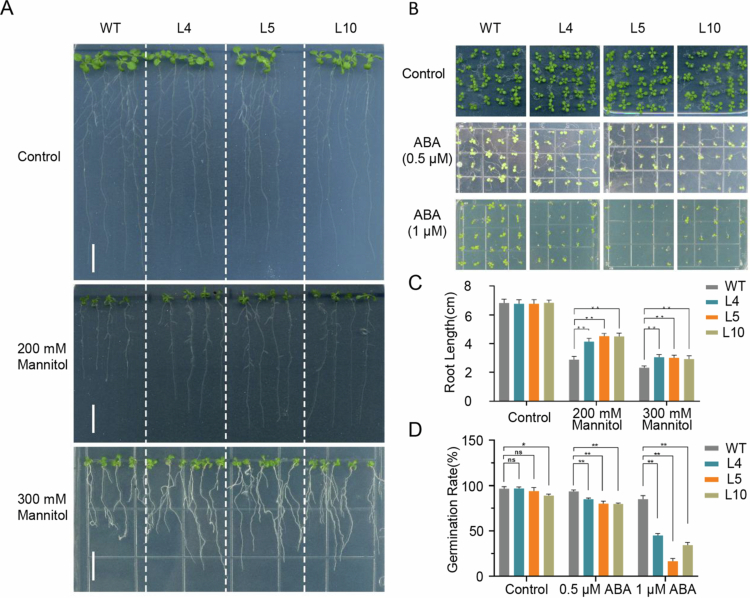 Figure 5
Figure 5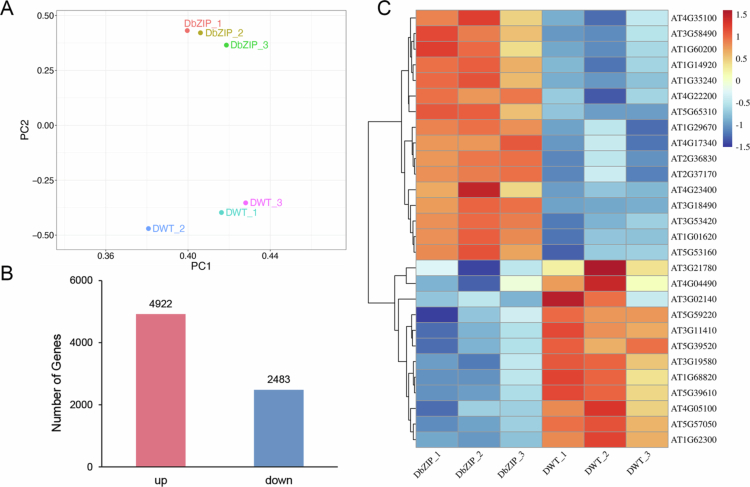 Figure 6
Figure 6- —The Technology Innovation Guidance Program of Gansu Province
- —University Natural Science Research Project of Anhui Province10.13039/501100009558
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Stress Responses and Tolerance · Plant Gene Expression Analysis · Plant Molecular Biology Research
Introduction
Water, which constitutes approximately 80%–90% of the plant body, is essential for all physiological processes.1 Drought decreases the soil moisture content in agricultural systems, with the resulting impaired crop growth and development decreasing yield and quality.2^,^3 To adapt to the uneven spatiotemporal distribution and varying intensity of natural precipitation, plants have evolved diverse regulatory mechanisms involving morphological, physiological, and molecular processes.4 In recent years, substantial progress has been made in elucidating the molecular basis of plant drought responses and in identifying genes that regulate stress resistance.5^,^6 Consequently, improving plant drought tolerance has become a key focus in abiotic stress research and breeding for crop stress resilience.7
Transcription factors regulate target gene expression by binding to specific DNA motifs in promoter regions. Numerous transcription factor families, including MYB, WRKY, NAC, AP2/ERF, and basic leucine zipper (bZIP), mediate plant responses to environmental stress.8 In eukaryotes, bZIP family members are one of the most widely distributed and evolutionarily conserved transcription factors.9 Notably, they are activated by various abiotic stresses, including salinity, drought, cold, and heavy metals.9^,^10 In rice (Oryza sativa L.), OsbZIP62 and OsbZIP66 enhance drought tolerance through the abscisic acid (ABA) signaling pathway,11–13 whereas OsbZIP71 binds to the promoters of abiotic stress response-related genes and improves salt tolerance.14 In Arabidopsis thaliana (Arabidopsis), silencing HY5, which encodes a key component of the light signaling pathway, leads to increased sensitivity to salinity.15 Additionally, AtbZIP63 mediates glucose–ABA crosstalk under stress conditions.16 In the complex network of epigenetic and hormonal interactions, transcription factors serve as critical hubs that directly decode signals, such as ABA, to activate downstream stress-responsive genes, highlighting the importance of elucidating their functions.17
ABA is the central phytohormone regulating plant adaptations to abiotic stress.18 The canonical ABA-dependent signaling pathway has been thoroughly characterized. Specifically, stress-induced ABA accumulation promotes the binding of PYR/PYL/RCAR receptors to clade A PP2C phosphatases, thereby relieving their repressive effects on SnRK2 kinases.19^,^20 The resulting activated SnRK2s phosphorylate downstream transcription factors, including extensively studied bZIP family members. Comparative genomic analysis of maize identified 125 bZIP genes that encode 170 distinct proteins; they were phylogenetically classified into 11 subfamilies on the basis of their homology to Arabidopsis and rice sequences.21 A previous study showed that overexpressing subfamily D member ZmbZIP4 increases ABA levels to enhance stress tolerance.22 Additionally, ZmbZIP76 in the F subclass of the bZIP family can bind to ACGT elements to enhance reactive oxygen species (ROS) scavenging and improve plant tolerance to salt and osmotic stresses.23 Subfamily S member ZmbZIP72 modulates abiotic stress responses through the ABA-dependent pathway,24 whereas subfamily I member ZmbZIP113 influences seed germination at low temperatures.25 Although bZIP transcription factors have been broadly characterized, the functions of subfamily I members in maize remain largely unexplored, leaving their potential role in abiotic stress responses an open question.
In this study, ZmbZIP92 was characterized as a drought- and ABA-inducible bZIP transcription factor that binds specifically to G-box elements. To dissect the potential molecular function of ZmbZIP92 within a controlled genetic background, the model plant Arabidopsis was used for heterologous expression analysis. The advantages of Arabidopsis include the availability of a fully sequenced genome, a mature genetic manipulation system, and a high degree of standardization in physiological assays, making it ideal for preliminary gene functional analysis and in-depth mechanistic investigations in plants.26^,^27 This strategy has been widely employed to elucidate the functions of stress-responsive genes in crops and reveal conserved regulatory mechanisms.28 In the current study, ZmbZIP92 overexpression in Arabidopsis enhanced drought tolerance by improving ROS scavenging. This study provides valuable insights into a key molecular mechanism underlying drought stress adaptation as well as a promising genetic resource for breeding drought-resilient maize varieties.
Materials and methods
Plant materials and growth conditions
The plant materials used in this study were as follows: B73 homozygous maize inbred line, wild-type (WT) Arabidopsis Columbia (Col-0) ecotype, and Nicotiana benthamiana. All plant materials were provided by the National Engineering Laboratory for Crop Stress Resistance Breeding. Maize plants were cultivated in a greenhouse set at 28 ± 2 °C with a 14-h light/10-h dark photoperiod. Arabidopsis plants were grown under a 24 °C (16-h light)/22 °C (8 h dark) cycle. Tobacco plants were maintained at 24 °C with a 16-h light/8-h dark cycle. In the Arabidopsis root elongation assay, 200 and 300 mM mannitol were used to simulate drought conditions.
Phylogenetic analysis of ZmbZIP92
On the basis of homology with the rice OsbZIP62 protein sequence in the NCBI database (https://www.ncbi.nlm.nih.gov/), ZmbZIP92 was identified as a homologous gene in maize. Its genomic and coding sequences were retrieved from MaizeGDB (https://maizegdb.org/) and Phytozome V13 (https://phytozome-next.jgi.doe.gov/). Homologous protein sequences from related species were downloaded from NCBI. Multiple sequences were aligned using MEGA 6.0 software, after which a phylogenetic tree was constructed according to the maximum likelihood method. Topological robustness was assessed using 1,000 bootstrap replicates to quantify branch support. The multiple sequence alignment was visualized using the default parameters of DNAMAN 6.0 to highlight conserved functional domains.
Plant transformation
The full-length ZmbZIP92 coding region was cloned into the p1301a vector to construct the 35S::ZmbZIP92 overexpression vector. WT Arabidopsis (Col-0) was transformed using an Agrobacterium-mediated floral dip method. Harvested T_0_ seeds were screened for positive transgenic plants on half-strength solid Murashige and Skoog (MS) medium containing 20 mg/L hygromycin B. ZmbZIP92 expression in transgenic lines was detected by quantitative real-time PCR (qRT-PCR) using primers qzmbzip92-F/R (Table S1) and AtActin as the internal reference gene.
RNA extraction and qRT-PCR analysis
Total RNA was extracted from maize or Arabidopsis tissues using TRIzol reagent (Huamaike Bio, Beijing, China). Purified RNA and an Evo M-MLV RT Mix Kit (Accurate Biology, Changsha) were used to synthesize cDNA via reverse transcription. The subsequent qRT-PCR analysis was performed using a LightCycler 480 system. Relative gene expression levels were calculated according to the 2^−ΔΔCt^ method. Statistical significance was determined by one-way ANOVA and Tukey’s post-hoc test, which were completed using GraphPad Prism version 9.1.1 (Graph-Pad, San Diego, CA, USA). All experiments were conducted using three biological replicates.
Subcellular localization of ZmbZIP92
The ZmbZIP92 coding region without a stop codon was fused in-frame with the green fluorescent protein (GFP)-encoding sequence to construct the 35S::ZmbZIP92-GFP/pCAMBIA1305 vector. The recombinant vector was inserted into Agrobacterium tumefaciens strain GV3101 (p19) cells for the subsequent infiltration of N. benthamiana cells on the abaxial side of leaves. After a 36-h incubation in darkness, GFP fluorescence in the transiently transformed leaves was observed using an LSM800 laser scanning confocal microscope.
Transcriptional self-activation assay of ZmbZIP92
The full-length ZmbZIP92 coding region was cloned into the yeast expression vector pGBKT7. Y2HGold yeast competent cells were transformed with the recombinant vector (pGBKT7-ZmbZIP92). Transformants were sequentially plated on solid SD/−Trp and SD/−Trp/−His/−Ade (containing X-α-gal) media, which were incubated at 30 °C for 3–5 days to monitor growth and color development. pGBKT7-53 and pGADT7-T were used as positive controls, whereas pGBKT7-Lam and pGADT7-T were used as negative controls.
Dual-luciferase reporter assay
The ZmbZIP92 coding region was cloned into the pGreenII 62-SK vector to serve as the effector. A fragment containing four tandem repeats of the G-box element (CACGTG) was inserted into the pGreenII 0800-LUC vector as the reporter. Effector and reporter vectors were used for the co-transformation of A. tumefaciens strain GV3101 (pSoup-p19) cells for the subsequent co-infiltration of N. benthamiana leaves. With the empty pGreenII 0800-LUC vector serving as a control, luciferase activity was measured at 36 h post-infiltration using a dual-luciferase reporter assay kit and a Tanon-5200 in vivo imaging system.
ABA sensitivity assay
Surface-sterilized seeds of transgenic and WT Arabidopsis were sown evenly on half-strength solid MS medium supplemented with 0, 0.5, or 1 μM ABA in plates. Each treatment was performed in triplicate. Plates were placed horizontally and cultured for 7 days, after which the seed germination rate was recorded. Data are presented herein as the mean ± standard deviation of at least three biological replicates (sample size ≥ 50).
Physiological analysis
Peroxidase (POD) activities and malondialdehyde (MDA) contents were determined using commercial assay kits (Solarbio, Beijing, China). The relative water content (RWC) of leaves was measured using a gravimetric method and the following formula: RWC (%) = [(fresh weight − dry weight)/(turgid weight − dry weight)] × 100. To determine the turgid weight, leaves were immersed in water for 8–10 h and then carefully blotted to remove surface moisture before weighing. Three biological replicates were used in each group.
RNA sequencing and analysis
Seeds of WT and ZmbZIP92-overexpressing line L5 Arabidopsis plants were surface-sterilized and sown on half-strength solid MS medium. After a 10-day incubation, seedlings were transplanted in nutrient soil. Following a 14-day cultivation, seedlings were exposed to drought stress. Leaves at the same developmental stage were collected (three biological replicates per group), flash-frozen in liquid nitrogen, and stored at −80 °C. Sequencing was performed using an Illumina NovaSeq 6000 system (LC Bio Technology Co., Ltd. Hangzhou, China). Differentially expressed genes (DEGs) were identified on the basis of FPKM values and the following thresholds: |log_2_(fold-change)| > 1 and p < 0.05.
Statistical analysis
Physiological and biochemical data were processed using GraphPad Prism 9.0 software (Graph-Pad). Data are presented herein as the mean ± standard deviation of at least three biological replicates (n ≥ 3). Statistical significance (^∗^p < 0.05; ^∗∗^p < 0.01) was determined by Student’s t-test and one-way ANOVA.
Results
ZmbZIP92 was identified as a novel bZIP gene in maize
Considering the reported contribution of OsbZIP62 to the rice response to drought stress,12 its homolog in maize was identified (i.e., ZmbZIP92) on the basis of comparative genomic analysis (Figure 1A). The encoded protein, which was classified in subgroup I among the 11 bZIP subclasses in maize, was revealed to contain a basic region for binding DNA and a leucine zipper domain that mediates protein dimerization. According to the constructed phylogenetic tree, maize ZmbZIP92 is closely related to rice OsbZIP62 and sorghum Sobic-002G425600 (Figure 1B), suggesting that ZmbZIP92 may encode a regulator of maize drought tolerance.
Phylogenetic tree of ZmbZIP92 and related bZIP proteins. (A) Phylogenetic tree of ZmbZIP92 and homologous bZIP proteins from various plant species. The tree was constructed by the neighbor-joining method with 1,000 bootstrap replicates using MEGA 7.0. All protein sequences were obtained from Phytozome v14.0. The blue regions indicates the conserved bZIP domain. (B) Multiple sequence alignment of amino acid sequences. The conserved bZIP and DNA-binding domains are indicated by solid lines beneath the sequences.
ZmbZIP92 expression pattern analysis
ZmbZIP92 expression profiles in various maize tissues and in response to ABA, NaCl, PEG-6000 (simulated drought stress), and 42 °C (heat stress) were determined via qRT-PCR. The generated data indicated that all tested abiotic stresses induced ZmbZIP92 expression, with a rapid initial increase followed by a gradual decrease (Figure 2B–E). In terms of tissue-specific expression, ZmbZIP92 transcript abundance was relatively high in roots, leaves, embryos, and endosperm, which was in contrast to the low transcript abundance in the stem at the eight-leaf stage and in silk at the flowering stage (Figure 2A).
Expression patterns of ZmbZIP92. (A) Tissue-specific expression of ZmbZIP92 at vegetative (V3: roots and leaves; V8: roots, stems, and leaves) and reproductive (SK: silks; E: embryo; END: endosperm) stages. (B-E) Expression of ZmbZIP92 under various abiotic stress treatments: simulated drought (20% PEG-6000), 100 μM ABA, heat (42 °C), and high salinity (200 mM NaCl). Samples were collected at the indicated time points after treatment.
Nuclear localization and transcriptional self-activation analysis of ZmbZIP92
To determine the subcellular localization and molecular properties of ZmbZIP92, we fused its coding sequence without the stop codon to the GFP-encoding sequence in an expression vector (Figure 3A) for the subsequent transient expression in N. benthamiana leaves. Green fluorescence due to the ZmbZIP92-GFP fusion protein was localized to the nucleus of tobacco leaf cells and its fluorescence signal highly overlapped with the spatial distribution of the Nuclear Localization Signal-mCherry-labeled nuclear region (Figure 3B), whereas the fluorescence of GFP alone was detected in the nucleus and cell membrane. Hence, ZmbZIP92 is a nuclear protein.
Identification and functional analysis of the transcription factor ZmbZIP92. (A) Schematic diagram of the construction of recombinant fluorescence vector 35S::GFP. (B) Subcellular localization of 35S::ZmbZIP92-GFP fusion protein in transiently transformed tobacco leaves. (Images acquired by laser scanning confocal microscopy). (C) Transcriptional activation assays. Yeast cells containing different plasmids were screened for growth on SD/-Trp and SD/-Trp/-His/-Ade/X-α-gal media. (D) The dual-LUC assay for ZmbZIP92 with G-box cis-acting elements. G-box-0800, mG-box-0800, pGreenII 62-SK empty vector, and ZmbZIP92-62-SK were transiently expressed in N. benthamiana. The LUC signal was captured at 72 h post-transfection. The LUC/REN ratio indicating the relative luciferase activity in the dual-LUC assay.
To analyze the transcriptional self-activation of ZmbZIP92, its full-length coding sequence was cloned into the pGBKT7 vector, which was then inserted into yeast cells. Colonies that grew on solid SD/−Trp medium were spotted onto solid SD/−Trp/−His/−Ade selection medium containing X-α-gal in plates. Yeast cells carrying pGBKT7-ZmbZIP92, like the negative control, failed to grow on the selection medium (Figure 3C), reflecting a lack of transcriptional self-activation under experimental conditions.
ZmbZIP92 bound specifically to the G-box element
The G-box (core sequence: CACGTG) is a conserved cis-regulatory element that serves as a binding site for bZIP transcription factors.29 To determine whether ZmbZIP92 binds specifically to this cis-regulatory element, recombinant vectors pGreenII 62-SK-ZmbZIP92 and pGreenII 0800-G-box were constructed for a dual-luciferase reporter assay. Significant luminescence was detected for the experimental group (62-SK-ZmbZIP92 and 0800-G-box) (Figure 3D), but not for any of the control groups (62-SK and 0800-G-box, 62-SK*-*ZmbZIP92 and 0800, and 62-SK and 0800), indicating that ZmbZIP92 binds specifically to the G-box element.
ZmbZIP92 overexpression enhanced drought resistance in Arabidopsis
Transgenic Arabidopsis plants overexpressing ZmbZIP92 under the control of the CaMV 35S promoter were generated. Three independent transgenic lines were identified by PCR and qRT-PCR, with the highest ZmbZIP92 expression level detected in line L5 (Figure S1A, B). Lines L4, L5, and L10 were selected for subsequent analyze on the basis of GUS staining results confirming that they were transgenic plants (Figure S1C).
Drought stress adversely affected both WT and ZmbZIP92-overexpressing plants (inhibited growth, wilting, and yellowing leaves) (Figure 4A). After rewatering, transgenic plants recovered nearly completely, which was in contrast to the highly impaired recovery of WT plants. These results reflect the enhanced drought tolerance of the transgenic plants.
Enhanced drought tolerance in ZmbZIP92-overexpressing Arabidopsis lines. (A) Representative phenotypes of wild-type (WT) and three independent ZmbZIP92-overexpressing lines (L4, L5, L10) after 14 days of drought stress followed by 3 days of re-watering. (B-D) Physiological indices of WT and transgenic lines under drought conditions: Peroxidase (POD) activity, relative water content (RWC), and malondialdehyde (MDA) content in leaves. Data are presented as means ± SD of three biological replicates. Statistical significance was determined by Student’s t-test (, P < 0.05, *, P < 0.01).
Physiological analysis revealed that ZmbZIP92 overexpression provided significant advantages under drought conditions. Compared with WT plants, transgenic plants had higher POD activities and RWC. Moreover, they accumulated less ROS and had lower MDA levels (Figure 4B–D; Figure S2). Under mannitol-simulated drought conditions, root growth was suppressed in both WT and transgenic plants (relative to that of untreated controls). However, in response to 200 and 300 mM mannitol treatments, ZmbZIP92-overexpressing plants developed significantly longer roots than WT plants, further supporting their improved tolerance to drought-like conditions (Figure 5A, C).
*Phenotypic analysis of ZmbZIP92-overexpressing Arabidopsis under simulated drought and ABA treatment. (A) Root lengths of wild-type (WT) and transgenic lines grown on 1/2 MS medium supplemented with 0, 200, and 300 mM mannitol. (B) Germination of WT and transgenic lines on 1/2 MS medium containing 0, 0.5, or 1 μM ABA. (C) Statistical comparison of root lengths under the indicated mannitol concentrations. Roots were measured after 10 days of growth in the greenhouse. (D) Statistical analysis of ZmbZIP92 overexpression lines and wild-type seed germination rates under ABA (0, 0.5, and 1 μM), respectively. Arabidopsis plants with two leaves after 7 days of growth were defined as successfully germinated. Each data point represents the mean of three independent biological replicates (mean ± SD, *, P < 0.05, *, P < 0.01).
Enhanced ABA sensitivity of ZmbZIP92-overexpressing Arabidopsis
Exogenous ABA applications rapidly induced ZmbZIP92 expression. To assess whether this induction altered ABA responsiveness, we compared seed germination rates between WT and transgenic plants treated with 0.5 or 1 μM ABA. At both ABA concentrations, germination was inhibited significantly more for the ZmbZIP92-overexpressing lines (L4, L5, and L10) than for the WT control (Figure 5B, D). Thus, ZmbZIP92 overexpression appeared to increase the sensitivity of Arabidopsis to ABA.
ZmbZIP92 mediated the drought response through the ABA signaling pathway
WT and L5 transgenic Arabidopsis plants exposed to drought stress underwent a transcriptome analysis. After confirming high sample consistency and sequencing data quality via PCA (Figure 6A), 7,405 DEGs were identified (Table S2). These DEGs consisted of 4,922 and 2,483 genes with upregulated and downregulated expression levels, respectively, in transgenic plants (Figure 6B). A set of 10 DEGs (five upregulated and five downregulated) was selected for further validation (Figure S3A, B). A total of 440 significantly enriched Gene Ontology (GO) terms from the molecular function, cellular component, and biological process categories were associated with the identified DEGs (Figure S4). These GO terms included key drought response-related terms, such as response to water deprivation (GO:0009414), ABA response (GO:0009737), defense response (GO:0006952), oxidative stress response (GO:0006979), salt stress response (GO:0009651), plant signal transduction (GO:0007165), and oxidation-reduction processes (GO:0055114). The subsequent Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis identified seven significantly enriched pathways, with plant hormone signal transduction revealed as the most enriched pathway (Figure S5).
Comparative RNA-seq analysis of Arabidopsis thaliana wild-type and ZmbZIP92-overexpressing line. (A) Principal component analysis (PCA) of global gene expression profiles across all sequenced samples. The plot visualizes the overall variance and grouping pattern of samples based on their transcriptional profiles. (B) Number of differentially expressed genes (DEGs) between wild-type (WT) and ZmbZIP92-overexpressing (L5) Arabidopsis plants under drought stress (|log₂FC| > 1, FDR < 0.05). (C) Heatmap of expression patterns for 28 selected genes associated with ABA signaling and stress adaptation under drought conditions, illustrating coordinated up- or down-regulation in the transgenic line.
By combining GO and KEGG analyses with protein functional annotations, 28 genes associated with ABA signaling and stress adaptations were identified. Cluster analysis indicated that 16 of these genes were upregulated and 12 were downregulated under drought conditions (Figure 6C). Functional annotations revealed that the upregulated genes included ASPG1 (AT3G18490) and RCAR3 (AT5G53160),30^,^31 which are implicated in ABA-related pathways, as well as PIP2A (AT3G53420), PIP1;5 (AT4G23400), PIP1C (AT1G01620), PIP2B (AT2G37170), PIP3 (AT4G35100), and TIP2;2 (AT4G17340),32–37 which encode regulators of transmembrane water transport. By contrast, the downregulated genes (AT3G02140, AT3G11410, AT3G19580, AT1G68820, and AT4G04490) are reportedly associated with ABA-inhibitory responses.38–42 Decreases in the expression of these genes may contribute to ABA accumulation and enhanced drought tolerance (Table S3). Considered together, these findings suggest that ZmbZIP92 may enhance drought tolerance by modulating ABA-related signaling pathways and coordinating the expression of key regulatory genes involved in stress adaptations and water homeostasis.
Discussion
Maize, the most important grain, feed, and energy crop globally, is susceptible to drought stress, which can lead to severe yield and quality losses. There is increasing evidence of the involvement of multiple gene types in plant drought stress responses that alleviate the adverse effects of drought on growth and development.43^,^44 According to earlier research, bZIP transcription factors are key regulators of plant responses to abiotic stresses, including salinity, drought, and temperature extremes.45^,^46 In plants, bZIP transcription factors are encoded by a relatively large gene family. A total of 170 bZIP transcription factors (encoded by 125 genes) have been identified and grouped into 11 subfamilies according to their phylogenetic relationships with orthologs in Arabidopsis (75 bZIP family members) and rice (89 bZIP family members).21 Numerous studies have clarified bZIP transcription factor functions and the associated mechanisms underlying abiotic stress responses. For example, in tomato, SlbZIP38 overexpression markedly decreases drought and salt tolerance, which may be attributed to decreased chlorophyll and free proline contents and increased MDA contents, suggesting that SlbZIP38 negatively regulates abiotic stress responses by modulating ABA signaling.47
In this study, the maize drought response‑related gene ZmbZIP92 was identified according to its homology to OsbZIP62, which positively regulates drought tolerance in rice.12 The Arabidopsis homolog of ZmbZIP92, EEL, reportedly enhances drought tolerance.48 Our analysis of gene expression profiles indicated that ZmbZIP92 is expressed in roots, leaves, and selected tissues, with transcript abundance peaking in roots and leaves at the three-leaf stage. As mentioned above, the expression of plant bZIP transcription factor genes can be induced by diverse abiotic stresses, with ZmbZIP92 overexpression significantly improving phytohormone treatments.10^,^45 Earlier research demonstrated that the rice bZIP genes OsbZIP62 and OsbZIP16, which are functional orthologs of ZmbZIP92, are transcriptionally activated by several abiotic stresses, such as drought and high salinity, as well as exogenous ABA.12^,^49 We also examined the effects of multiple stress treatments (200 mM NaCl, 20% PEG-6000, 42 °C, and 100 μM ABA) on ZmbZIP92 expression. Our findings showed that ZmbZIP92 expression was significantly induced by various abiotic stresses and exogenously applied ABA, but especially by drought and ABA. Hence, this gene may encode a protein with crucial functions during abiotic stress responses. The nuclear localization of ZmbZIP92 further supports its identification as a canonical bZIP transcription factor. Transcriptional self-activation is a common feature of transcription factors. However, transcriptional self-activation of ZmbZIP92 was undetectable in yeast. Interestingly, similar findings were reported for OsbZIP62 and OsbZIP46.12^,^50 Although full-length OsbZIP62 cannot activate transcription, its N-terminal fragment can if the C-terminal domain is absent.12 Similarly, full-length OsbZIP46 does not activate transcription, but removing part of its C-terminal leads to detectable transcriptional activation.50 Notably, most bZIP transcription factors are transcriptional activators, including BhbZIP58, CsbZIP55, and CsbZIP65.51^,^52 Specifically, ZmbZIP92 has characteristics analogous to those of OsbZIP62 and OsbZIP46, implying that they may have a common conserved regulatory mechanism.
ZmbZIP92-overexpressing transgenic Arabidopsis plants were obtained to clarify the ZmbZIP92 function under drought conditions. Mannitol treatments promoted root elongation in transgenic plants, with the subsequent water deficit stress assays revealing a significant increase in drought tolerance. Thus, ZmbZIP92 expression appears to be important for plant adaptive responses to drought stress. The enhanced stress tolerance of transgenic plants was reflected by increased POD activity and RWC along with a decrease in MDA contents. Collectively, these findings indicate that ZmbZIP92 overexpression confers improved drought tolerance because of the associated positive regulation of drought stress responses. Therefore, ZmbZIP92 functions as a transcription factor that positively regulates drought stress responses; the biological functions of ZmbZIP92 and its rice ortholog OsbZIP62 are conserved.12 Transcription factors mainly modulate the expression of downstream target genes by binding to their promoter regions, thereby enabling plants to adapt to various external stresses.53^,^54 We determined that ZmbZIP92 can bind to the G-box element of downstream target genes and mediate expression.29 Genes potentially regulated by ZmbZIP92 were identified on the basis of RNA sequencing data. Moreover, according to significantly enriched GO terms, the 7,405 identified DEGs were associated with responses to ABA, salinity stress, water deprivation, and oxidative stress. KEGG pathway analysis indicated that most DEGs were involved in the MAPK signaling pathway and plant hormone signal transduction. Hence, these DEGs may contribute to the adaptive response of Arabidopsis to drought stress. Among the identified upregulated DEGs, AT3G18490 encodes an aspartic protease (ASPG1) that promotes drought tolerance in Arabidopsis through the ABA signaling pathway,30 while AT5G53160 encodes an ABA receptor (PYL8) that positively regulates ABA signaling.31^,^55 The downregulated genes included AT3G02140, which encodes tandem MAC motif-containing protein 2; this protein contains two or more ABRE elements and is a key negative regulator of ABA and salt stress responses.38 Another downregulated gene, AT3G11410, encodes protein phosphatase 2CA, which negatively regulates ABA signaling.42^,^56 In addition, genes encoding proteins that regulate water transport, such as PIP1D (AT4G23400) and PIP1C (AT1G01620), had upregulated expression levels.33^,^57ZmbZIP92 expression is significantly induced by the application of ABA, with the corresponding transgenic plants exhibiting increased sensitivity to exogenous ABA. This suggests that ZmbZIP92 may not uniformly amplify all downstream effects of ABA signaling, thereby optimizing plant resource allocation under stress conditions.
Increased ABA sensitivity enables plants to initiate basic defense responses, such as stomatal closure, relatively early and rapidly.18^,^58 More importantly, after integrating transcriptomic data, we speculated that ZmbZIP92 preferentially activates ABA-responsive genes directly related to stress adaptations, including genes related to water retention and ROS scavenging. Simultaneously, ZmbZIP92 may have limited effects on ABA signaling that strongly inhibits cell growth and expansion. This differential regulation of downstream ABA signaling pathways enables the plant to prioritize survival under drought conditions, thereby enhancing overall stress resistance without incurring excessive growth costs.59 However, the absence of direct binding evidence (e.g., ChIP‑qPCR and EMSA) makes it unclear whether the identified DEGs are directly targeted by ZmbZIP92. Alternatively, the observed changes in the expression of these genes may be due to indirect responses within the regulatory network. This possibility decreases the direct mechanistic evidence of ZmbZIP92 binding to the G‑box promoter element and activating downstream gene expression.
Although our analysis of heterologous expression in Arabidopsis suggests that ZmbZIP92 participates in ABA‑dependent drought responses, whether this transcription factor functions similarly in maize remains to be determined. The distinct regulatory networks of monocot species may alter the molecular mechanism or target gene specificity of ZmbZIP92. Definitive functional validation is required in future loss‑ and gain‑of‑function analysis of maize.
Conclusion
This study functionally characterized ZmbZIP92 as a maize bZIP transcription factor. Phylogenetic analysis classified it as a Group I member in the bZIP family, while also revealing its homology to bZIP proteins from Arabidopsis and rice. Subcellular localization confirmed that ZmbZIP92 is a nuclear protein. Moreover, ZmbZIP92 expression was induced by abiotic stresses, including salinity and drought. Although the full-length protein lacked transcriptional self-activation, it was observed to bind specifically to the G-box cis-element. Under drought conditions, Arabidopsis plants overexpressing ZmbZIP92 exhibited enhanced drought tolerance, characterized by significant increases in RWC and POD activity as well as decreased MDA accumulation. Furthermore, ZmbZIP92-overexpressing plants were more sensitive to ABA than WT plants. Molecular analysis revealed that in response to drought stress, ZmbZIP92 overexpression modulated the expression of key ABA signaling pathway genes, concurrently upregulating positive regulators and downregulating negative regulators. In summary, the study data imply that ZmbZIP92 positively regulates the maize drought stress response, while also providing important molecular insights relevant to deciphering the mechanisms mediating maize drought tolerance.
Supplementary Material
Figure S1.tifFigure S1.tif
Figure S2.tifFigure S2.tif
Figure S3.tifFigure S3.tif
Figure S4.tifFigure S4.tif
Figure S5.tifFigure S5.tif
Table S1.xlsxTable S1.xlsx
Table S2.xlsxTable S2.xlsx
Table S3.xlsxTable S3.xlsx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brodersen CR, Roddy AB, Wason JW, Mc Elrone AJ. Functional status of xylem through time. Annu Rev Plant Biol. 2019;70:407–433. doi: 10.1146/annurev-arplant-050718-100455.30822114 · doi ↗ · pubmed ↗
- 2Blum A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017;40(1):4–10. doi: 10.1111/pce.12800.27417527 · doi ↗ · pubmed ↗
- 3Hussain HA, Hussain S, Khaliq A, Ashraf U, Anjum SA, Men SN, Wang LC. Chilling and drought stresses in crop plants: implications, cross talk, and potential management opportunities. Front Plant Sci. 2018;9:393. doi: 10.3389/fpls.2018.00393.29692787 PMC 5902779 · doi ↗ · pubmed ↗
- 4Meng S, Cao Y, Li HG, Bian Z, Wang DL, Lian CL, Yin WL, Xia XL. Pe SHN 1 regulates water-use efficiency and drought tolerance by modulating wax biosynthesis in poplar. Tree Physiol. 2019;39(8):1371–1386. doi: 10.1093/treephys/tpz 033.30938421 · doi ↗ · pubmed ↗
- 5Chen F, Chen L, Yan Z, Xu JY, Feng LL, He N, Guo ML, Zhao JX, Chen ZJ, Chen HQ, et al. Recent advances of CRISPR-based genome editing for enhancing staple crops. Front Plant Sci. 2024;15:1478398. doi: 10.3389/fpls.2024.1478398.39376239 PMC 11456538 · doi ↗ · pubmed ↗
- 6Oyebamiji YO, Adigun BA, Shamsudin NA, Ikmal AM, Salisu MA, Malike FA, Lateef AA. Recent advancements in mitigating abiotic stresses in crops. Horticulturae. 2024;10(2):156. doi: 10.3390/horticulturae 10020156. · doi ↗
- 7Sato H, Mizoi J, Shinozaki K, Yamaguchi-Shinozaki K. Complex plant responses to drought and heat stress under climate change. Plant J. 2024;117(6):1873–1892. doi: 10.1111/tpj.16612.38168757 · doi ↗ · pubmed ↗
- 8Thilakarathne AS, Liu F, Zou ZW. Plant signaling hormones and transcription factors: key regulators of plant responses to growth, development, and stress. Plants-Basel. 2025;14(7):1070. doi: 10.3390/plants 14071070.40219138 PMC 11990802 · doi ↗ · pubmed ↗