The global distribution of Paragonimus species
Yuan-Biao Lu, Kai Hu, Mei-Lin Mao, Lin-Bo Shi, Chun-Chao Zhu, Jie-Xin Zou

TL;DR
This paper maps the global distribution of lung fluke species that cause paragonimiasis, highlighting where they occur and where more research is needed.
Contribution
A comprehensive global map of Paragonimus species distribution, identifying species-specific geographic ranges and highlighting gaps in knowledge.
Findings
Paragonimus species are found in Asia, Africa, and the Americas, but none are distributed across multiple continents.
Eight species can infect humans, with distinct geographic ranges such as P. westermani in East Asia and P. kellicotti in North America.
The distribution of Paragonimus species and cases is uneven, with many suitable habitats lacking reported occurrences.
Abstract
Paragonimiasis, a foodborne zoonotic disease caused by Paragonimus (lung fluke) species, is prevalent mainly in tropical and subtropical regions. It is estimated that 23 million people are infected worldwide. Many reviews have been published in recent years, but very few reviews focused on distribution have been published. Here, we performed a review to map the global distribution of Paragonimus species. We systematically searched Google Scholar, Web of Science, PubMed, ScienceDirect, African Journals Online, the Chinese National Knowledge Infrastructure (CNKI), Wanfang Database, and Chongqing VIP Chinese Science and Technology Journal Database to identify studies and case reports documenting the occurrences of Paragonimus species (OPSs) and cases of paragonimiasis. Studies were included only if both the OPSs and the geographical locations of Paragonimus species occurrence had been…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
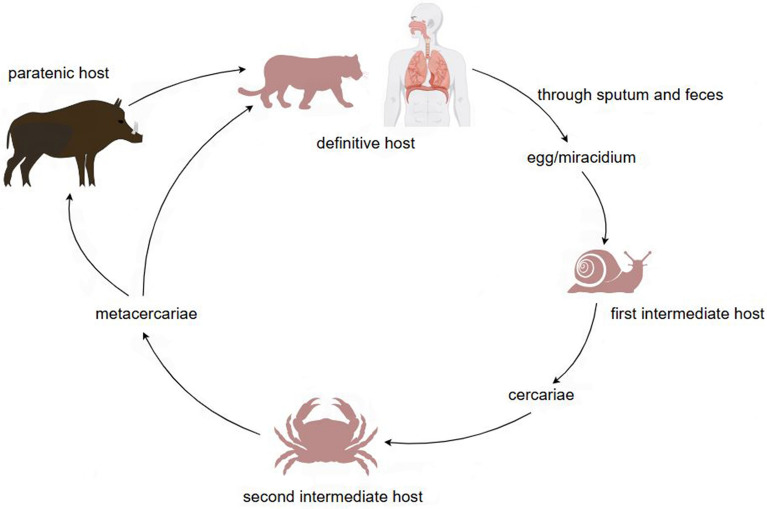 Figure 1
Figure 1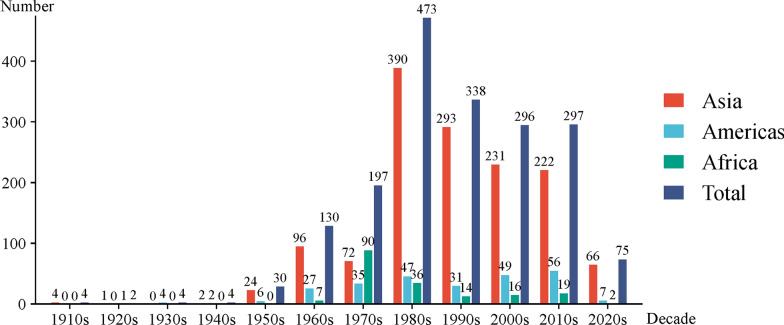 Figure 2
Figure 2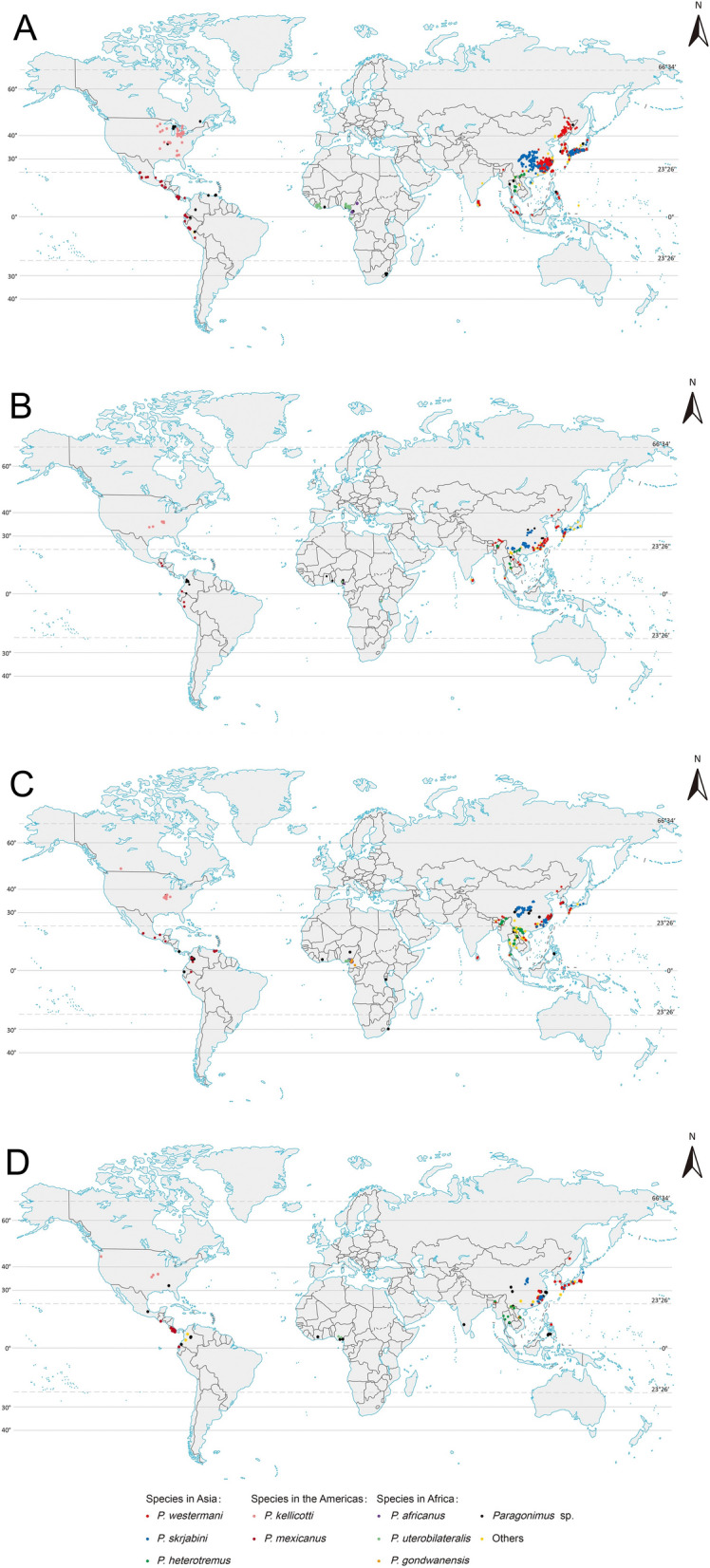 Figure 3
Figure 3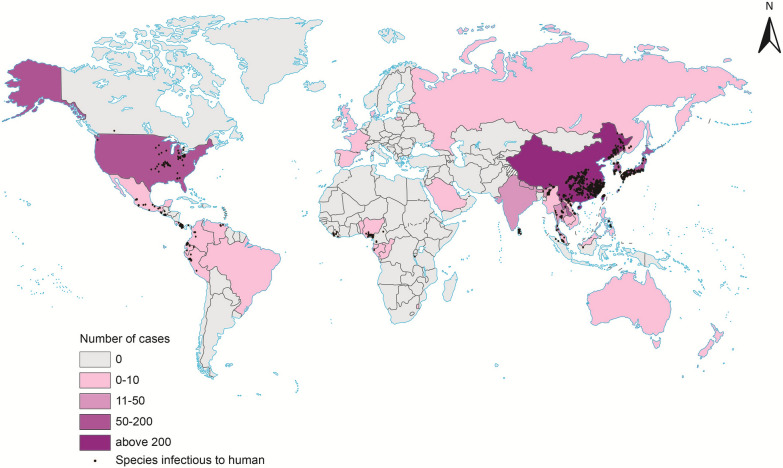 Figure 4
Figure 4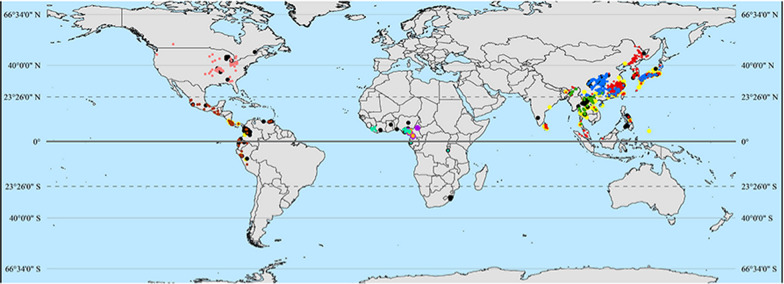 Figure 5
Figure 5- —the National Parasitic Resources Center
- —https://doi.org/10.13039/501100001809National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasites and Host Interactions · Helminth infection and control · Parasitic Diseases Research and Treatment
Background
Paragonimiasis, a neglected tropical disease [1], is a foodborne zoonotic disease caused by Paragonimus (lung fluke) species. It is broadly distributed in tropical and subtropical regions, including Asia, Africa and the Americas [2–5]. It was estimated that 23 million people are infected worldwide, mostly in Asia [6]. Paragonimus species are hermaphroditic trematodes with complex life cycles and at least three types of hosts: first intermediate hosts, second intermediate hosts and definitive hosts [2, 7]. Eggs of Paragonimus species produced by adult worms infecting definitive hosts can enter water through feces and sputum. Eggs develop into miracidia in water at suitable temperatures (Fig. 1). When miracidia meet the first intermediate hosts (usually freshwater snails), they enter the snails and give rise eventually to cercariae. Cercariae released by snails infect second intermediate hosts (usually freshwater crabs and crayfish) and develop into metacercariae in crustaceans (Fig. 1). Animals (definitive hosts) can be infected by ingesting crustaceans, and then metacercariae can develop into adults. Paratenic hosts are optionally involved in the life cycle, because sometimes metacercariae do not develop into adults but remain as juveniles in the tissues of the hosts [2, 3, 5, 7]. Among the more than 50 species worldwide, eight can infect humans [5]: Paragonimus westermani (Asia), P. skrjabini (Asia), P. heterotremus (Asia), P. africanus (Africa), P. uterobilateralis (Africa), P. gondwanensis (Africa), P. kellicotti (North America), and P. mexicanus (Central and South America). The human cases of infection with P. gondwanensis are based on parasite eggs in a couple of humans only, so that there is some uncertainty about this diagnosis. Humans generally get infected by consuming raw or undercooked crustaceans. Sometimes, infection in humans occurs through the consumption of paratenic hosts (such as case reports of the consumption of wild boars in Japan) [7–10]. In parts of Asia, paragonimiasis is more common due to pickling or eating raw or uncooked freshwater crabs, such as the “drunk crab” in China and the “kejiang” in the Republic of Korea [11–14]. In Africa, however, diseases caused by foodborne trematodes are rare because the thorough cooking of most foods, a tradition in Africa, will kill metacercariae [15, 16]. Among Paragonimus species that typically cause pulmonary disease, some worms may cause pleural disease, or stray beyond the thoracic cavity to cause ectopic paragonimiasis [17], which can affect almost any part of the body. In areas endemic for tuberculosis (TB), patients initially misdiagnosed with drug-resistant TB were found to be infected with paragonimiasis, confirming that the region is also endemic for paragonimiasis [18].Fig. 1. Life cycle typical of Paragonimus species including names of life-cycle stages in the corresponding hosts
Traditionally, the taxonomy of Paragonimus species was based on the morphology of adult worms and the form of metacercariae [5]. However, for many Paragonimus species, there are too few morphological features of the adult stage to distinguish between species, and the morphological characteristics of the metacercariae stage are considered unreliable [19, 20]. With the development of sequencing technology, molecular data have become popular in the areas of taxonomy and phylogeny. The addition of molecular data has helped solve some taxonomy problems. For example, the systematic position of P. caliensis Little, 1968, was determined via the use of molecular data [21]. However, the application of molecular data simultaneously adds to the complexity of taxonomy [5]. Whether the challenge of taxonomic classification becomes simpler or more complex hinges on the degree to which molecular phylogenetic information aligns with the morphological characteristics of Paragonimus species.
Species
We here present the world register of Paragonimus species based on studies by Blair et al. 1999, Yoshida et al. 2019, and Zhou et al. 2021 [2–4]. In the twenty-first century, five new species of Paragonimus were reported: P. vietnamensis from Vietnam (Asia), P. pseudoheterotremus from Thailand (Asia), P. sheni from China (Asia), and P. gondwanensis and P. kerberti from Cameroon (Africa) (Table 1). Two species of the genus Euparagonimus were included in Table 1 in this study. The reason was that multiple studies have shown that Euparagonimus is highly similar to species of the Paragonimus in terms of phylogenetic relationships and both should belong to the genus Paragonimus, despite some differences between them in morphology and life cycle [5, 22]. The two Euparagonimus species were distributed only in southeastern China. Although there are 57 nominal Paragonimus species worldwide, no species are distributed across continents (the Americas is regarded as a whole). More information about the taxonomy, synonyms and distribution of Paragonimus species is provided in Table 1. In the field of public health research, there are a total of 8 human-infecting species globally, with three in Asia, two in the Americas and three in Africa. A brief introduction to the eight species follows. Table 1. The world register of Paragonimus speciesNoSpeciesSynonymsSecond hosts (family)Distribution1P. bangkokensis (Miyazaki and Vajrasthira, 1967)No synonymPotamidaeThailand, China, Laos, Vietnam2P. cheni (Hu, 1963)No synonymPotamidaeChina3P. compactus (Cobbold, 1859)No synonymNot sureIndia, Sri Lanka4P. fukienensis (Tang and Tang, 1962)No synonymPotamidae, GecarcinucidaeChina5P. harinasutai (Miyazaki and Vajrasthira, 1968)No synonymPotamidae, GecarcinucidaeThailand, China, Laos, Vietnam6P. heterorchis (Zhou et al., 1982)No synonymPotamidaeChina7P. heterotremus (Chen and Hsia, 1964)P. tuanshanensis (Chung et al., 1964)Potamidae, GecarcinucidaeChina, Thailand, Laos, Vietnam, India8P. jiangsuensis (Cao et al., 1983)P. xiangshanensis (He et al., 1995)PotamidaeChina9P. macrorchis (Chen, 1962)No synonymGecarcinucidae, PotamidaeChina, Sri Lanka, Thailand10P. microchis (Hsia et al., 1978)No synonymPotamidaeChina11P. minqinensis (Li and Chen, 1983)No synonymPotamidaeChina12P. mungoi (Mishra et al., 1976)No synonymNot sureIndia13P. ohirai* (Miyazaki, 1939)P. sadoensis (Miyazaki et al., 1968)P. iloktsuenensis (Chen, 1940)Potamidae, Sesarmidae, VarunidaeJapan, China, the Republic of Korea14P. paishuihoensis (Tsao and Chung, 1965)P. divergens (Liu et al., 1980)PotamidaeChina, Laos, Thailand15P. pantheri (Mishra and Patal, 1976)No synonymNot sureIndia16P. proliferus* (Hsia and Chen, 1964)P. menglaensis (Chung et al., 1964)P. hokuoensis (Ho et al., 1965)PotamidaeChina, Vietnam17P. siamensis (Miyazaki and Wykoff, 1965)No synonymGecarcinucidae, PotamidaeThailand, Sri Lanka18P. skrjabini (Chen, 1959)P. hueitungensis (Chung et al., 1975)P. miyazakii (Kamo et al., 1961)P. szechuanensis (Chung and Tsao, 1962)P. veocularis (Chen and Li, 1979)PotamidaeChina, Japan, Thailand, India, Vietnam19P. sheni (Shan, Lin et al., 2009.)No synonymPotamidaeChina20P. taipingini (Kurochkin, 1987)No synonymNot sureChina21P. westermani (Kerbert, 1878)P. asymmetricus (Chen, 1977)P. edwardsi (Gulati, 1926)P. filipinus (Miyazaki, 1978)P. macacae (Sandosham, 1953)P. philippinensis (Ito et al., 1978)P. pulmonalis (Baelz, 1880)P. ringeri (Cobbold, 1880)Potamidae, Gecarcinucidae, Cambaroididae, VarunidaeIndia, Nepal, Thailand, the Philippines, Indonesia, Papua New Guinea, Myanmar, Vietnam, Cambodia, Laos, the Republic of Korea, China, Japan, Far East of Russia22P. yunnanensis (Ho et al., 1959)No synonymPotamidaeChina23P. vietnamensis (Doanh et al., 2007)No synonymPotamidaeVietnam24P. pseudoheterotremus (Waikagul, 2007)No synonymPotamidaeThailand25P. africanus Voelker and Vogel, 1965No synonymPotamonautidaeCameroon, Nigeria, Gabon, Equatorial Guinea, Ivory Coast26P. kerberti Bayssade-dufour et al., 2015No synonymPotamonautidaeCameroon27P. gondwanensis Bayssade-dufour et al., 2014No synonymPotamonautidaeCameroon, Ivory Coast28P. uterobilateralis Voelker and Vogel, 1965No synonymPotamonautidaeCameroon, Nigeria, Liberia, Guinea, Gabon29P. amazonicus Miyazaki, Grados and Uyema, 1973No synonymPseudothelphusidaePeru30P. caliensis Little, 1968No synonymPseudothelphusidaeColombia, Peru, Panama, Mexico, Costa Rica31P. inca Miyazaki, Mazabel, Grados and Uyema, 1975No synonymPseudothelphusidae, TrichodactylidaePeru32P. kellicotti Ward, 1908No synonymCambaroididaeUSA, Canada33P. mexicanus Miyazaki and Ishii, 1968P. ecuadoriensis Voelker and Arzube, 1979P. peruvianus Miyazaki et al., 1969Pseudothelphusidae, TrichodactylidaeMexico, Costa Rica, Panama, Guatemala, Ecuador, Peru34P. napensis Amunarriz, 1991No synonymNot sureEcuador35P. rudis (Diesing, 1850) Stiles and Hassall, 1900No synonymNot sureBrazil36Euparagonimus cenocopiosus (Chen, 1962)No synonymPotamidae, GecarcinucidaeChina37Euparagonimus hongzesiensis (Fu et al., 1990)No synonymNot sureChinaThe synonyms, second hosts and country-level distributions of Paragonimus species are also shown^*^: Nomen nudum^**^: Species inquirenda
Paragonimus westermani
The first reported Paragonimus species was described from the lungs of the otter Pteronura braziliensis (Gmelin, 1788) in Brazil [2]. In 1878, Kerbert described a new species, Distomum westermani, from a Bengal tiger at the Amsterdam zoological gardens in the Netherlands [23]. The following year, Dr. Ringer detected a species of parasite, Distomum ringeri (Cobbold, 1880), in the body of a Portuguese sailor from Taiwan, China, which was the first time that human paragonimiasis was reported [24]. Shortly thereafter, Baelz reported a species, D. pulmonalis (Baelz, 1883), in Japan [24]. In 1899, Braun split the genus Distomum to many genera*,* one of which is Paragonimus, and the name “Paragonimus” is still used [24]. The referred two species above (D. ringeri and D. pulmonalis) were regarded as synonyms of Paragonimus westermani. In later studies, many new species, such as P. asymmetricus (Chen, 1977), P. edwardsi (Gulati, 1926) and P. filipinus (Miyazaki, 1978), were reported and regarded as synonyms of P. westermani [2, 25–30].
Paragonimus skrjabini
Chen first briefly described P. skrjabini from the lungs of the viverrid Paguma larvata, which was purchased from markets in Guangzhou (a city in southern China) in 1959, and more morphological descriptions were provided for the following year [27]. In 1962, Chung and Tsao described P. szechuanensis from cats in Sichuan Province [31, 32]. However, Chen compared this species with P. skrjabini in terms of the morphological aspects of the cuticular spines, the shape and size of the eggs, the body proportions of the adults and the species of the hosts and reported that P. szechuanensis is the same as P. skrjabini [33]. In 2003, Cui et al. collected five types of geographical strains of P. skrjabini from different provinces in China and verified that P. szechuanensis is a synonym of P. skrjabini on the basis of the morphology and phylogeny of different geographical strains [34, 35]. Paragonimus hueitungensis, P. veocularis and P. miyazakii are also regarded as synonyms of P. skrjabini [2, 27, 28, 36, 37] (Table 1).
Paragonimus heterotremus
Paragonimus heterotremus was first reported in Guangxi Zhuang Autonomous Region, China by Chen and Hsia in 1964, but specific collection locations, collection times and collectors were not provided in this study [38]. In the same year, Chung et al. described P. tuanshanensis from Xishuangbanna Dai Autonomous Prefecture, Yunnan Province [39]. According to Chen in 1965, P. tuanshanensis is regarded as a synonym of P. heterotremus [40]. Li et al. performed a detailed comparison of the morphology of the two species and considered the two to be the same species [27].
Paragonimus kellicotti
The discovery of P. kellicotti was almost simultaneously reported by Ward and Kellicott in 1894 [41]. After the description of infection of this species in a cat that was found in Michigan, United States by Ward, Kellicott described the infection in a dog in Columbus, Ohio, the United States later that year [42]. However, Ward identified the species as Distoma westermani at that time [43]. Subsequently, Ward and Hirsch determined that the North American species was a distinct Paragonimus species [44], which they named P. kellicotti. P. kellicotti is the only Paragonimus species endemic to North America [24]. Paragonimiasis caused by P. kellicotti is rare in humans but is usually observed in wild and domestic animals [24, 45].
Paragonimus mexicanus
The species was described from Colima, Mexico, by Miyazaki and Ishii in 1968 [46]. Paragonimus ecuadoriensis and P. peruvianus were described from coastal Ecuador in 1979 and from Peru in 1969, respectively. These two species were regarded as synonyms of P. mexicanus [47] by Miyazaki et al. However, López-Caballero considered that P. mexicanus is a species complex [48]. Tongu suggested that the other six species in Latin America are P. mexicanus [49]. Usually, only P. ecuadoriensis and P. peruvianus are regarded as synonyms of P. mexicanus [2]. Studies on the taxonomy of Paragonimus species in Latin America are ongoing [50, 51]. P. mexicanus is distributed in all countries in Latin America where Paragonimus species are endemic [2, 5, 51].
Paragonimus africanus
Paragonimus africanus was described from Lower Bakossi, West Cameroon, by Voelker and Vogel in 1965 [52]. The species is distributed mainly in West Africa [52, 53]. The species richness of Paragonimus species in Africa is low (only four), and none of the species has synonyms. Paragonimiasis caused by this species is usually pulmonary [5].
Paragonimus uterobilateralis
This species was reported together with P. africanus from Cameroon in the same study [52]. The distributions of P. uterobilateralis and P. africanus strongly overlapped. Paragonimiasis caused by the species P. uterobilateralis is usually pulmonary [5].
Paragonimus gondwanensis
In 2014, Paragonimus gondwanensis was described from the lungs of naturally infected cats and civets in Cameroon by Bayssade-Dufour et al. [54]. This species is the most recently reported human-infecting Paragonimus species. The following year, Bayssade-Dufour et al. described another species, P. kerberti, but there are no reports of paragonimiasis in humans caused by this species [5]. There are few studies on this species, so more studies on its distribution and disease characterization are needed.
Human cases caused by P. westermani have been reported in China, Japan, the Republic of Korea, the Far East of Russia and the Philippines, whereas in other countries with this species’ range (e.g., Thailand, Malaysia), human cases caused by this species have not been reported. Given the taxonomy complexity and controversy of Paragonimus species, we will not discuss much about it in this study. The taxonomy we used in this article follows that of Blair et al. 1999, Yoshida et al. 2019, and Zhou et al. 2021 [2–4].
In recent years, many general reviews have focused on the biology, evolution and medical significance of Paragonimus or paragonimiasis [3, 5, 16, 55–59]. These studies cover many aspects and are highly valuable. However, very few studies have focused on mapping the occurences of Paragonimus species (OPSs) or paragonimiasis (only two studies in recent years): one focused on Africa [16], and the other focused on China [4]. However, these two studies are regional and not global. Mapping is essential for identifying endemic foci; e.g., a study revealed that infections of Clonorchis sinensis in animals are primarily concentrated in regions with low altitudes and high precipitation [60]. Thus, we aimed to (1) update the World Register of Paragonimus species, (2) map the distributions of OPSs and paragonimiasis cases globally, and (3) present a list of the second hosts of Paragonimus species. The list of the second hosts is included in this study because of its importance. This is not only because humans are usually infected with Paragonimus by ingesting crustaceans [2, 4, 56], but more importantly, isolating Paragonimus metacercariae carried by crustacean hosts is the most commonly used way in which epidemiologists and parasitologists obtain Paragonimus samples.
Methods
Search strategy
The search for target literature was conducted via four global and four regional databases: Google Scholar, Web of Science, PubMed, ScienceDirect, African Journals Online (Africa), Chinese National Knowledge Infrastructure (CNKI, https://www.cnki.net/, China), Wanfang Database (https://www.wanfangdata.com.cn/, China), and Chongqing VIP Chinese Science and Technology Journal Database (http://qikan.cqvip.com/, China). The search terms used were as follows: Paragonimus, lung fluke and paragonimiasis (see Additional file 1). The search terms in Chinese were used for the three Chinese databases, and the search terms in English were used for the other databases from 1 January, 1900 to 31 December, 2024.
Inclusion and exclusion criteria
Studies of OPSs and paragonimiasis were included if they met the following criteria: (1) the OPSs were verified according to the study, (2) the site where the Paragonimus species occurred was clear, or (3) the case reports of patients with a confirmed diagnosis of paragonimiasis were included. Studies of OPSs that met the following criteria were excluded: (1) the OPSs were not confirmed (e.g., a diagnosis of paragonimiasis using ELISA only), (2) studies that reported the OPSs in a large area (e.g., a country or a large-area state or province), and (3) case reports that did not show where the patients ate crustaceans and got infected (Given inherent human mobility, we cannot reliably equate the location where paragonimiasis is diagnosed with the area where Paragonimus species are endemic). Publications of case reports were extracted for mapping the distribution of paragonimiasis cases.
Data extraction and quality assessment
Eligible studies of OPSs were divided into three groups on the basis of the study region: Asia, the Americas, and Africa. The following data were extracted from the included studies: Paragonimus species, second hosts, site, longitude, latitude, publication year, decade, study language, and corresponding literature (see Additional file 2). Most studies do not provide information on the longitude and latitude of the OPSs, so we obtained this information via Google Maps (https://www.google.com/maps) based on the corresponding sites. “Decade” means the decade in which the study was published. Part of the data from China and Africa were obtained directly from the two studies mentioned above and another study focusing on China in 2000 [4, 5, 61]. The data of eligible studies of paragonimiasis cases can be seen in Additional file 3. The first author and the second author independently assessed the risk of bias for the included studies via the Joanna Briggs Institute (JBI) Critical Appraisal Tools [62]. “Low risk of bias” represents a summary score of 0–3, “moderate risk of bias” represents a summary score of 4–6, and “high risk of bias” represents a summary score of 7–10. Each study was assigned a low, moderate or high risk of bias.
The names of different second hosts recorded in the file are the scientific names used currently. Updated names of crustacean hosts were obtained via the WoRMS database (https://www.marinespecies.org/index.php). In some publications, the species names were wrongly used. Some could be easily identified (such as Eriocheir japonica wrongly used as Eriocheir japonicus and Eriocher japonieum), so that we can rectify these species names in Additional file 4. However, some species names cannot be related to any of the scientific names used now. These names are marked with asterisks.
Data analysis and mapping
The data were organized and analyzed in Microsoft Excel (version 2016; Microsoft Corporation, Redmond, USA). Figures except for the maps and the life cycle figures were produced on the online platform “https://www.bioinformatics.com.cn”. The map data set is provided by Geospatial Data Cloud site, Computer Network Information Center, Chinese Academy of Sciences. (http://www.gscloud.cn) [63]. The maps of different time periods of Paragonimus species were produced via ArcGIS 10.2 (Environment System Research Institute, Redlands, USA), by importing the coordinate data to the software. The period was divided according to the estimation of paragonimiasis in 1995 by the WHO [64] and in 2005 and 2015 by the Global Burden of Disease Study [65]. The different periods of OPSs in the five maps were as follows: “total” (mapping all the OPSs), “before 1995” (mapping the OPSs before 1995), 1996–2005 (mapping the OPSs from 1996 to 2005), 2006–2015 (mapping the OPSs from 2006 to 2015), and 2016–2024 (mapping the OPSs from 2016 to 31-December, 2024). The coordinate information for each map is provided in Additional file 5.
Results
Study selection and data characteristics
For OPSs studies, a total of 19,854 articles were identified in eight databases. After title and abstract screening and duplicate removal, 17,611 articles were excluded. After full-text assessment, 931 of 2243 articles meeting the inclusion criteria were included. In all the included studies, those in English were most frequently used (approximately 48.7%). Chinese studies were the second most common, followed by Japanese, Spanish, Korean, French, and German studies (see Additional file 6). In the Americas, English studies were also the most frequently included, followed by Spanish studies (see Additional file 6). In Africa, English studies were most frequently included, followed by French and German studies (see Additional file 6). In contrast to studies in the Americas and Africa, studies in Chinese have the maximum quantity in Asia, followed by studies in English. Notably, 39 and 21 Japanese and Korean studies, respectively, were included (see Additional file 6).
Few studies recorded the OPSs before the 1950s. Since the 1950s, the number of OPSs has increased rapidly and peaked in the 1980s. Since the 1980s, the number of OPSs has declined gradually and remained stable in the 2000s and 2010s. In contrast to other regions, Asia experienced a decline in the 1970s. In nearly all decades, the number of OPSs in Asia reached a maximum, and that in Africa reached a minimum. Notably, in the 1970s, the number of OPSs in Africa (90) surpassed that in Asia (72) and reached a maximum (Fig. 2; see Additional file 7).Fig. 2. Bar chart showing OPSs (Occurrences of Paragonimus species) by decade (e.g. 1990s include the years of “1990 to 1999” in this bar chart) and region. Different colors indicate different regions. “Total” represents the sum of the three regions (Asia, the Americas and Africa)
Mapping of Paragonimus species
We constructed five maps to visualize the OPSs, especially the eight species that infect humans (Fig. 3). Among the 1915 OPSs extracted from the included studies, the majority were reported in Asia (1466, 76.6%), followed by the Americas (264, 13.8%) and Africa (185, 9.6%). (see Additional file 5). Three human-infecting species are distributed in Asia (P. westermani, P. skrjabini and P. heterotremus), two human-infecting species (P. kellicotti and P. mexicanus) from the Americas and three from Africa (P. africanus, P. uterobilateralis and P. gondwanensis). This study suggests that Paragonimus species are distributed from 12° S to 50° N globally, except four “outliers” in South Africa (approximately 30° S). Notably, Paragonimus species in the Northern Hemisphere can extend to high-latitude regions, whereas those in the Southern Hemisphere can only live in tropical regions. The number of second hosts of Paragonimus species globally is highest in Asia, medium in the Americas and lowest in Africa (see Additional file 4). Additional file 4 was extracted from Additional file 2 to clearly display the second hosts from different regions.Fig. 3. Maps of the OPSs (Occurrences of Paragonimus species) globally during different periods. A. (1900–1995), B. (1996–2005), C. (2006–2015), D. (2016–31-December, 2024). The dots of different colors indicate different Paragonimus species. Undetermined Paragonimus species were categorized as "Paragonimus sp.", and those not infecting humans as "Others". The map data set is provided by Geospatial Data Cloud site, Computer Network Information Center, Chinese Academy of Sciences. (http://www.gscloud.cn). Map approval number: GS(2026)0525
Asia
There are three human-infecting species distributed in Asia. P. westermani has the broadest distribution (East Asia, the Far East, Southeast Asia, India, Sri Lanka, probably Nepal, New Guinea) not only in Asia but also in the world. P. skrjabini is distributed mainly in China and Japan. P. heterotremus is limited to the Indochina Peninsula and southwestern border regions of China and northeastern India [66] (Table 1; Fig. 3).
China, where three Asian human-infecting species are present, has the greatest species richness of Paragonimus [2, 4]. P. westermani and P. skrjabini are the dominant species. P. westermani is distributed mainly in southeastern China and northeastern China, whereas P. skrjabini is distributed mainly in central China. In contrast to the two widely distributed species, P. heterotremus is limited to the southwestern border regions of China (Guangxi Zhuang autonomous region and Yunnan Province) [4] (Fig. 3). In Japan, there are two human-infecting species: P. westermani and P. skrjabini miyazakii. The distributions of the two species overlapped across nearly all of Japan except Hokkaido (Fig. 3). The occurrence of P. ohirai is also frequently reported in Japan [7, 67]. There is only one human-infecting species in the Republic of Korea: P. westermani. This species is distributed throughout the country. Articles on the Far East of Russia were so rare that only a few occurrences were mapped (Fig. 3). In this area, there is only one human-infecting species: P. westermani ichunensis. In India, there are three human-infecting species that are distributed mainly in northeastern regions: P. westermani, P. skrjabini and P. heterotremus. In Sri Lanka, there is only one human-infecting species: P. westermani. Furthermore, the situation is slightly different in Southeast Asia compared with that in other regions. In Malaysia, the Philippines and Indonesia, P. westermani is the dominant species without a distribution of P. heterotremus, whereas in other countries on the Indochina Peninsula, P. heterotremus is the dominant species (Table 1; Fig. 3).
The endemic foci of Paragonimus species in Asia have been stable since 1995 (Fig. 3). OPSs in northeastern China were reported mainly before 1995. After 1995, few occurrences were reported in this region (Fig. 3). After 2015, there was no occurrence in this region (Fig. 3D). Occurrences in the Indochina Peninsula from 2006 to 2015 are very intensive, even more than those in the period before 1995 (Fig. 3A, C).
In Asia, China has the greatest species richness of the second hosts of Paragonimus species (62), which is far greater than that in other regions of the world (see Additional file 4). The species richness of the second hosts of Paragonimus species in all other Asian countries is lower than 10. In northeastern China, the Republic of Korea and the Far East in Russia, freshwater crayfish are the second hosts of Paragonimus species, whereas freshwater crabs are the second hosts in other regions of Asia. More information on the second intermediate hosts of Paragonimus species in Asia can be found in Additional file 4.
The Americas
There are two human-infecting Paragonimus species distributed in the Americas: P. kellicotti and P. mexicanus. The two species are entirely geographically separated: P. kellicotti is distributed in North America, whereas P. mexicanus is distributed in Latin America (Fig. 3). There is a “gap region” between the USA and Mexico, in which there are no reports of OPSs (Fig. 3).
In the USA and Canada, only one species has been reported: P. kellicotti. It is distributed mostly in the eastern part of the USA, and Ontario and Quebec in Canada (Fig. 3). A study published in 2020 reported the OPSs in canine feces at three off-leash dog parks in Portland, Oregon, USA, which is located on the Pacific coast and is far from the main endemic foci in the USA [68] (Fig. 3). Another study published in 2011 detected P. kellicotti from coyote feces in Calgary, Alberta, Canada, which is also far from the main endemic foci in Canada [69] (Fig. 3). In countries in Latin America, there is only one human-infecting species: P. mexicanus. In Mexico, the species is distributed in the southern part of the country. In Colombia, the species is distributed in the northwest of the country. In Venezuela, P. mexicanus is distributed in the northern coastal region. In Ecuador, the species is distributed in the western coastal region and northern part of the country. In Peru, the species is distributed in the western part (Fig. 3).
In the Americas, Mexico has the highest species richness of the second hosts (9), which is one more than that of Colombia. In the USA and Canada, freshwater crayfish are the second hosts of Paragonimus species as freshwater crabs do not occur there, whereas freshwater crabs are the second hosts of Paragonimus species in other regions of the Americas (Additional file 2; Additional file 4). More information on the second intermediate hosts of Paragonimus species in the Americas can be found in Additional file 4.
Africa
Three human-infecting species have been reported in Africa: P. africanus, P. uterobilateralis and P. gondwanensis. The three species are mostly distributed in West and Central Africa. In Cameroon, all three human-infecting species are present and distributed mainly in the western coastal regions. In Gabon, only one species, P. uterobilateralis, which is distributed in the northwestern coastal regions, has been reported. In Liberia, only one species has been reported: P. uterobilateralis. In Nigeria, three human-infecting Paragonimus species have been reported, and they are distributed mainly in the southern coastal regions. Occurrences of P. uterobilateralis and Paragonimus species undetermined were reported in Tanzania and South Africa, respectively (Fig. 3).
In Africa, Cameroon has the greatest species richness (5) of second hosts, with one more than that of Nigeria (see Additional file 4). There are no freshwater crayfish species that serve as second hosts of Paragonimus species in Africa, unlike in Asia and the Americas (see Additional file 4). More information on the second intermediate hosts of Paragonimus species in Africa can be found in Additional file 4.
Mapping of paragonimiasis case distribution
To further analyze the relationship between the distribution of Paragonimus species and the prevalence of paragonimiasis from a public health perspective, we searched relevant literature and created a global map of paragonimiasis case distribution. In total, 666 papers were included in the data set, numbering 6561 cases. To date, according to our study, cases of paragonimiasis have been reported in 35 countries (Additional file 3). Generally, the ranking of paragonimiasis case reports by number is Asia, the Americas, Africa, Europe, and Oceania (from high to low). China reported the highest number of paragonimiasis cases (n = 6047). With the exception of China, no other country reported over 200 cases, indicating a global imbalance in the reporting of paragonimiasis cases. One potential explanation is the difference in research attention devoted to Paragonimus studies across nations. In some countries including Malaysia (number of paragonimiasis cases = 1), Vietnam (1) and Colombia (3), OPSs were reported frequently, whereas only a small number of paragonimiasis cases were documented. In contrast, in countries such as Saudi Arabia (1), Denmark (1), Switzerland (1), Australia (1), New Zealand (1), Spain (1), England (3), and France (4), no OPSs were detected, yet paragonimiasis cases were reported. It may be attributed to imported cases from other paragonimiasis-endemic foci, highlighting the need for vigilance against biological invasion by these parasites.
Discussion
This study comprehensively investigated the global distribution of Paragonimus on the basis of literature reports. For some species with disputed names, we did not conduct further species identification, including the cryptic species within the complex species [5, 70]. We hope to provide public health workers with more reliable evidence for the prevention and control of paragonimiasis from a global perspective.
A comparison of different periods revealed that there was an “outbreak period” of reports of new Paragonimus species (32 species) in the 1960s and 1970s (Table 1). In contrast, since the start of the twenty-first century, only five new species were reported. The potential reasons may include the following: fewer as-yet-unknown cryptic Paragonimus species than previously; and diminished research interest among investigators.
Due to the complexity of Paragonimus life cycle, from the perspective of public health researchers, strategies for preventing and controlling paragonimiasis should not only focus on the human population but also pay attention to the hosts of Paragonimus, especially the crustacean hosts. Different epidemic areas should develop targeted prevention strategies due to variations in intermediate hosts and dietary habits. Notably, the invasive species “Procambarus clarkii”, which is native to North America, has been reported to harbor Paragonimus species in Asia and South America [71–73]. In particular, “Procambarus clarkii” is cooked as food and is very popular in Asia. The occurrence of biological invasions and species extinction may alter the burden of paragonimiasis. Attention should be paid to such invasions in endemic foci because it may increase the burden of paragonimiasis.
Based on the global distribution of Paragonimus species, Paragonimus has a dominant distribution in the Northern Hemisphere between 0°N and 50°N, in tropical and subtropical regions. In Africa, Paragonimus species are distributed mainly in West Africa and Central Africa, where the tropical rainforest climate is common, with a hot climate and sufficient precipitation. Other regions in Africa that lack precipitation are mainly deserts and grasslands, which may not be suitable habitats for Paragonimus species and their first and second hosts. Notably, some regions in Australia in which freshwater snails and crabs are distributed [74, 75] are suitable habitats for Paragonimus species; however, no occurrences of the species have been reported in these regions. Specifically, some countries with warm or hot climates and sufficient precipitation, such as India, Cambodia, Myanmar and Brazil, are suitable habitats for Paragonimus species. However, there are very few or even no reports of the OPSs in these countries. In contrast, a certain number of cases were reported in these countries or regions. Therefore, there is a speculation here that the occurrences of the Paragonimus species identified via mapping may have been underestimated. Studies in many countries have not been performed enough. The most direct evidence is that there have been no reports of the OPSs in Cambodia, around which countries all have endemic foci of paragonimiasis. In Myanmar, there is one report of the occurrence of P. heterotremus [76]. The situations of many other countries are similar to those of Cambodia and Myanmar, especially countries with humid climates in tropical and subtropical regions. More attention should be paid to these countries, as there may be new species that have never been found in these countries.
Figures 2 and 3 show that the number of reports of Paragonimus species has decreased over the past 40 years. This may have resulted from control measures implemented by governments, economic growth, improvements in public health awareness, decreased research interest, and so on. In the 1970s, the number of OPSs in Africa surpassed that in Asia (Fig. 2), which is abnormal. This may be attributed to food shortages and simultaneous consumption of freshwater crabs resulting from the Nigerian Civil War of 1967–1970 [77–79]. Crabs are more likely to be consumed by people living in rural regions due to poverty and food shortages. While this practice has become rare today, it is crucial to remain aware that it may resurface. Such a resurgence could be triggered by factors like infectious diseases, wars, or other crises that lead to regional food shortages.
Based on paragonimiasis case reports mapping (Fig. 4), in countries, such as Australia, some North Africa countries and European countries (Fig. 4), no Paragonimus species were reported, while paragonimiasis cases were documented. This phenomenon may be attributed to the global population mobility driven by enhanced transportation accessibility. In this era of advanced development, we should remain vigilant against the potential establishment of Paragonimus and other endemic parasitic organisms in previously non-endemic regions via population mobility.Fig. 4. Global map showing the number of paragonimiasis per country, from reported cases. The purpler colors indicate a higher number of paragonimiasis case reports, while pinker shades indicate lower number. White indicates no case reports in the country. The black dots indicate the exact location of human-infecting Paragonimus species. The map data set is provided by Geospatial Data Cloud site, Computer Network Information Center, Chinese Academy of Sciences. (http://www.gscloud.cn). Map approval number: GS(2026)0525
Importantly, we are trying to collect more data to present the authentic “Paragonimus world”. However, our study only reflects the distribution of Paragonimus species and cases caused by them based on these literatures we compiled, and is by no means an authentic distribution. How much data we can compile depends on the interest of researchers, surveillance and reporting biases, literature accessibility, and so on.
Conclusions
Our study updated the world register of Paragonimus species and mapped their global distribution. On the basis of these findings, Paragonimus species and paragonimiasis cases exhibit an imbalanced distribution across hemispheres, continents, and countries. The prevalence of Paragonimus species in many regions and countries may be underestimated. It is recommended that more epidemiological surveys or other surveys on Paragonimus species be conducted in countries or regions suitable for the survival of Paragonimus species but with relatively few or no reported occurrences. This study enhances the understanding of the spatiotemporal distribution of Paragonimus species worldwide and can provide support for the intervention and control of paragonimiasis (lung fluke disease), thereby promoting the improvement of public health.
Supplementary Information
Supplementary Material 1. Table S1. Search terms for literature in eight databasesSupplementary Material 2. Data recording OPSs extracted from eligible studiesSupplementary Material 3. Data recording paragonimiasis case reportsSupplementary Material 4. The list of second intermediate hosts of ParagonimusSupplementary Material 5. Coordinates of OPSs for the mapsSupplementary Material 6. The number of studies by different languagesSupplementary Material 7. The number of OPSs by decade
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ward HB. On the presence of Distoma westermanni in the United States. Vet Mag. 1894:355–357.
- 2Tongu Y. The species of Paragonimus in Latin America. Summary of Health Care Department of School of Medicine at Okayama University. 2001;12(1):1–5.
- 3Geospatial Data Cloud site. http://www.gscloud.cn. Accessed 21 Apr 2025.