Callus-mediated plant regeneration and CRISPR/Cas9-targeted mutagenesis in Oenanthe javanica
Kai Feng, Cheng Yao, Hui Lv, Zhiyuan Yang, Jialu Liu, Ziqi Zhou, Nan Sun, Shuping Zhao, Peng Wu, Aisheng Xiong, Liangjun Li

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
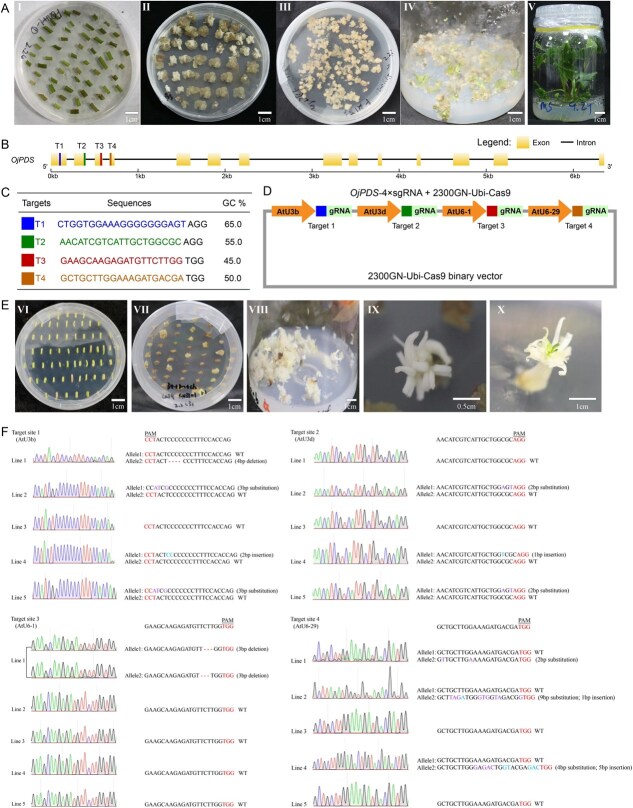 Figure 1
Figure 1- —National Natural Science Foundation of China10.13039/501100001809
- —China Agriculture Research System
- —Jiangsu seed industry revitalization project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCRISPR and Genetic Engineering · Plant tissue culture and regeneration · Plant Molecular Biology Research
Dear Editor,
Oenanthe javanica (Blume) DC., a perennial aquatic herb in the Apiaceae family, is widely cultivated in East Asian countries [1]. Oenanthe javanica is a medicinal and edible plant, which has high economic value and is popular for its distinctive aroma and crisp texture. Rich nutrients and pharmacological substances confer on O. javanica the therapeutic potentials, such as calming the liver, reducing blood pressure, preventing thrombosis, and anticancer effects [2]. Due to germination disorder and seed dormancy, O. javanica is commonly propagated through stem-cuttings in cultivation. Traditional asexual propagation has many disadvantages, such as high propagation costs, low reproductive coefficient, season dependence, and viral disease occurrence [3]. The long-term asexual propagation of O. javanica restricted the genetic diversity and germplasm innovation through traditional crossbreeding. The emergence of plant tissue culture and genetic transformation provides the efficient strategy to address these challenges [4].
Plant tissue culture technology has been developed on the basis of the totipotency of plant cells, including micropropagation, adventitious shoot regeneration, and somatic embryogenesis. Varieties, types of explants, disinfection processes, and the composition of the medium are the main factors influencing the plant tissue culture. The composition of the medium mainly includes macro elements, trace elements, iron salts, sucrose, vitamin, and plant growth hormone. Efficient tissue culture technology is an important basis for constructing a plant genetic transformation system. Genetic transformation can be used for the overexpression and silencing expression of functional genes in plants. The CRISPR/Cas (clustered regularly interspaced short palindromic repeat/CRISPR-associated protein) system for genome editing has been widely used in precise engineering of genomes and crop breeding [5]. PDS gene is a key gene in carotenoid biosynthesis, responsible for converting the colorless compound phytoene into the colored compound ζ-carotene. It plays an important role in photosynthesis and pigment biosynthesis in plants, and disruption of its function leads to albinism. Recently, the T2T genome of O. javanica has been published, which provides important genetic information for the gene editing in O. javanica [6].
Here, the callus-mediated regeneration and CRISPR/Cas9-targeted mutagenesis system in O. javanica were established. The vigorous axillary buds of O. javanica were sterilized and cultivated to obtain the aseptic seedlings. The petioles, leaf blades, and roots of O. javanica were selected as different explant types to evaluate the callus induction efficiency. The different kinds of explants were cut into ~5-mm-sized fragments and cultivated under dark conditions. Based on the quantitative comparisons of callus induction efficiency of different explants, petioles were investigated to be the most suitable explants for callus induction in O. javanica (Fig. S1). Similarly, petioles were also used as the suitable explant in other Apiaceae species, including celery [7] and carrot [8]. The callus induction efficiency is usually affected by many factors, such as plant growth regulators (PGRs) and basic culture medium. Murashige & Skoog (MS) medium and Gamborg B5 medium were selected as the basic mediums for callus induction of O. javanica. The petioles of aseptic seedling were used as explants and inoculated on MS and B5 medium supplemented with different concentrations of 2,4-D, NAA and 6-BA. Based on the comprehensive analysis of all treatments, B5 culture medium supplemented with 0.5 mg/l of 2,4-D and 1.0 mg/l of 6-BA showed the highest induction rates and better callus growth status than other treatments (Tables S1–S4). Notably, B5 culture medium supplemented with 1.0 mg/l of 2,4-D and 0.5 mg/l of 6-BA was the most suitable treatment for callus proliferation (Tables S5 and S6). MS basic medium supplemented with 0.5 mg/l of NAA and 2.0 mg/l of 6-BA was the optimum medium for callus differentiation of O. javanica (Table S7). Hence, the efficient callus-mediated regeneration system of O. javanica was established (Fig. 1A and Fig. S2A).
Loss-function of phytoene desaturase (PDS) gene usually lead to the albino phenotype, which is widely used as visual target gene for evaluating the efficiency of gene editing in plants. To obtain the PDS homologous alleles, the sequence of DcPDS (GenBank accession XM_017385289.1) was used to conduct the BLAST alignment in the genome of O. javanica [6]. One single-copy PDS gene, Oj17G000040.1, was identified from O. javanica and designated as OjPDS (Figs S3 and S4). The OjPDS gene was cloned from O. javanica cv. ‘Fuqin No.1’ using the specific primers OjPDS-F and OjPDS-R (Table S8). OjPDS protein contains the phytoene–desat domain, which belongs to the phytoene dehydrogenase family and was involved in the carotene biosynthesis (Fig. S5). To ensure the precise editing of OjPDS gene, four target sites (T1–T4) individually located in the first four exons were designed using CRISPR-GE tool (Fig. 1B and C) [9]. Four Arabidopsis promoters (AtU3b, AtU3d, AtU6-1, and AtU6-29) were selected to individually drive the expression of four sgRNA cassette containing T1–T4 targets to improve the gene editing efficiency (Fig. S6). Then, the four sgRNA expression cassettes were constructed into the modified pYLCRISPR/Cas9Pubi-H vector, and the recombinant OjPDS-2300GN-Ubi-Cas9 binary vector was transformed into the Agrobacterium tumefaciens GV3101 for genetic transformation assay (Fig. 1D) [7].
Based on our optimized callus-mediated regeneration system, 603 explants were infected with A. tumefaciens GV3101 containing OjPDS-2300GN-Ubi-Cas9. To screen the positive transformed callus, the transformed explants were cultivated on B5 culture medium supplemented with 0.5 mg/l of 2,4-D, 1.0 mg/l of 6-BA, 50 mg/l of kanamycin, 100 mg/l of l-serine and 300 mg/l of carbenicillin. Finally, a total of 14 lines of O. javanica plants were regenerated from the callus via somatic embryogenesis pathway, including albino, green, and chimeric plants (Fig. 1E and Fig. S2B). To determine the efficiency of CRISPR/Cas9-mediated targeted mutagenesis, the genomic DNA was extracted from the positive plants using CTAB method. The genomic sequences harboring four target sites were amplified from the extracted DNA using PDS-detection-F and PDS-detection-R primers (Table S8). The PCR products were direct sequenced or cloned into the pCE2 TA/Blunt-Zero vector followed by Sanger sequencing. The decoded sequences indicated that ten lines of O. javanica were edited, indicating the CRISPR/Cas9-mediated targeted mutagenesis was constructed with the efficiency of 1.7%. Multiple mutation types were detected from the gene edited O. javanica plants, including base deletion, base insertion, base substitution and combination mutagenesis. The mutation efficiency at T1 (targeted by AtU3b-driven sgRNA) was 90%, including two homozygous mutations and four biallelic mutations, while the remaining were heterozygous mutations. At T2 (targeted by AtU3d-driven sgRNA), the mutation efficiency was 70%, consisting of three homozygous mutations and four heterozygous mutations. The mutation efficiency at T3 (targeted by AtU6-1-driven sgRNA) was 20%, including one heterozygous mutation and one biallelic mutation. The mutation efficiency at T4 (targeted by AtU6–29-driven sgRNA) was the same as that at T2, including six heterozygous mutations and one biallelic mutation. The mutation efficiency at T1, T2, and T4 was significantly higher than at T3 (Fig. 1F and Fig. S7). Homozygous mutations were observed at the T1 and T2 targets, while no such mutations were detected at T3 and T4. This indicates that the observed differences in mutation efficiency among T1, T2, T3, and T4 are related to promoter type. Furthermore, the mutation efficiency at T1 was higher than that at T2. The mutation efficiency at T1 was driven by AtU3b promoter, which has been widely used in various crops for gene editing [10]. Among the gene-edited plants, four lines were exhibited as heterozygotes, with a frequency of 40%. The frequency of chimeras and homozygous phenotypes were calculated as 20% and 40%, respectively.
In conclusion, this study constructed the efficient callus-mediated plant regeneration by optimizing the explant type, basic culture medium, varieties and proportion of PGRs. This study also provided the first report of CRISPR/Cas9-mediated targeted mutagenesis in O. javanica. These results will have tremendous application prospects in functional genes verification and innovation of excellent germplasm through molecular breeding in O. javanica.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wang XJ, Luo Q, Li T. et al. Origin, evolution, breeding, and omics of Apiaceae: a family of vegetables and medicinal plants. Hortic Res. 2022;9:uhac 07638239769 10.1093/hr/uhac 076PMC 10795576 · doi ↗ · pubmed ↗
- 2Lu CL, Li XF. A review of Oenanthe javanica (Blume) DC. as traditional medicinal plant and its therapeutic potential. Evid Based Complement Alternat Med. 2019;2019:1–1710.1155/2019/6495819 PMC 646358831057651 · doi ↗ · pubmed ↗
- 3He Z, Sheng SY, Wang LQ. et al. Cucumber mosaic virus-induced gene and micro RNA silencing in water dropwort (Oenanthe javanica (Blume) DC). Plant Methods. 2024;20:638212839 10.1186/s 13007-023-01129-4PMC 10782793 · doi ↗ · pubmed ↗
- 4Zhang X, Xu G, Cheng C. et al. Establishment of an Agrobacterium-mediated genetic transformation and CRISPR/Cas 9-mediated targeted mutagenesis in Hemp (Cannabis sativa L.). Plant Biotechnol J. 2021;19:1979–8733960612 10.1111/pbi.13611 PMC 8486249 · doi ↗ · pubmed ↗
- 5Wolabu TW, Mahmood K, Jerez IT. et al. Multiplex CRISPR/Cas 9-mediated mutagenesis of alfalfa FLOWERING LOCUS Ta 1 (Ms F Ta 1) leads to delayed flowering time with improved forage biomass yield and quality. Plant Biotechnol J. 2023;21:1383–9236964962 10.1111/pbi.14042 PMC 10281603 · doi ↗ · pubmed ↗
- 6Feng K, Liu JL, Sun N. et al. Telomere-to-telomere genome assembly reveals insights into the adaptive evolution of herbivore-defense mediated by volatile terpenoids in Oenanthe javanica. Plant Biotechnol J. 2025;23:2346–5740112135 10.1111/pbi.70062 PMC 12120883 · doi ↗ · pubmed ↗
- 7Liu JX, Li T, Wang H. et al. CRISPR/Cas 9-mediated precise targeted mutagenesis of phytoene desaturase in celery. Hortic Res. 2022;9:uhac 16236204201 10.1093/hr/uhac 162PMC 9531335 · doi ↗ · pubmed ↗
- 8Xu ZS, Feng K, Xiong AS. CRISPR/Cas 9-mediated multiply targeted mutagenesis in orange and purple carrot plants. Mol Biotechnol. 2019;61:191–930644027 10.1007/s 12033-018-00150-6 · doi ↗ · pubmed ↗