The Complete Chloroplast Genome of a Vietnam Endemic Aroid, Homalomena perplexa K.Z.Hein, Vuong, Bao & V.S.Dang, and Updated Comparative Genomics in Araceae
Nhat Nam Nguyen, Ngoc Trai Nguyen, Hoang Dang Khoa Do

TL;DR
This study sequenced the complete chloroplast genome of a rare Vietnamese plant and compared it to related species to better understand their evolutionary relationships.
Contribution
The study provides the first complete chloroplast genome of Homalomena perplexa and updates comparative genomics in Araceae.
Findings
The chloroplast genome of H. perplexa is 169,893 bp long with 79 protein-coding genes.
Phylogenetic analysis confirmed the close relationship between Homalomena and Furtadoa genera.
Comparative analysis revealed 13 junction variations in Aroideae chloroplast genomes.
Abstract
Homalomena perplexa K.Z.Hein, Vuong, Bao & V.S.Dang has been recently recognized and identified as a Vietnamese endemic species. Its chemical contents and bioactivities have been investigated, but its genomic information is still scarce. In the current study, we sequenced and characterized the complete chloroplast genome of H. perplexa using the next‐generation sequencing method. The circular genome was 169,893 bp in length and contained 79 protein‐coding genes, 30 transfer RNA genes, and four ribosomal RNA genes. Nucleotide diversity analysis revealed only two hypervariable regions between the two Homalomena chloroplast genomes. However, repeat analysis indicated that there were shared and unique repeats between H. perplexa and H. occulta chloroplast genomes. In Araceae chloroplast genomes, there was a high variation of the junctions among the large‐single copy, small‐single copy,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 FIGURE 1
FIGURE 1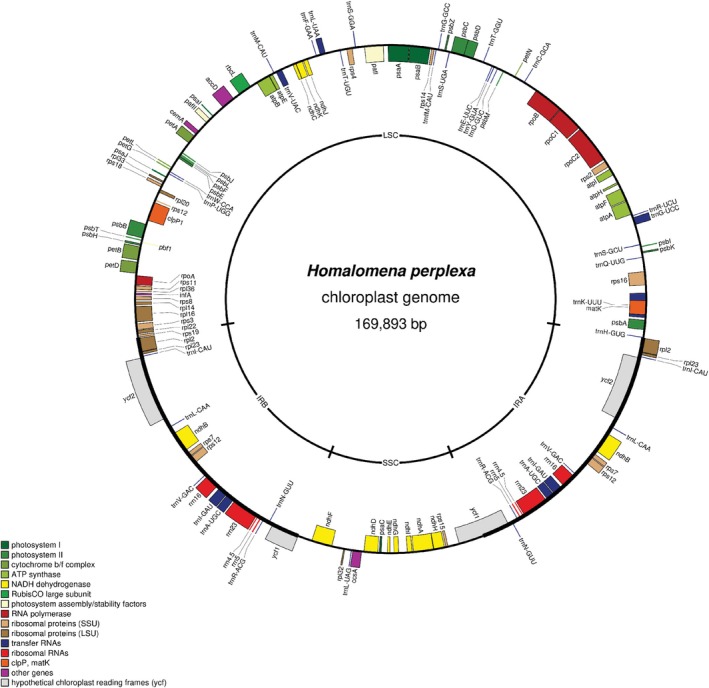 FIGURE 2
FIGURE 2 FIGURE 3
FIGURE 3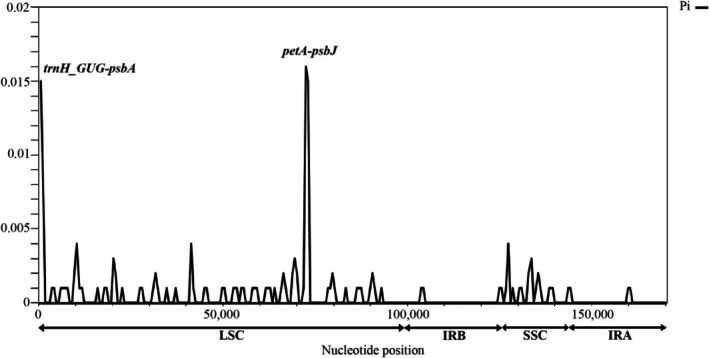 FIGURE 4
FIGURE 4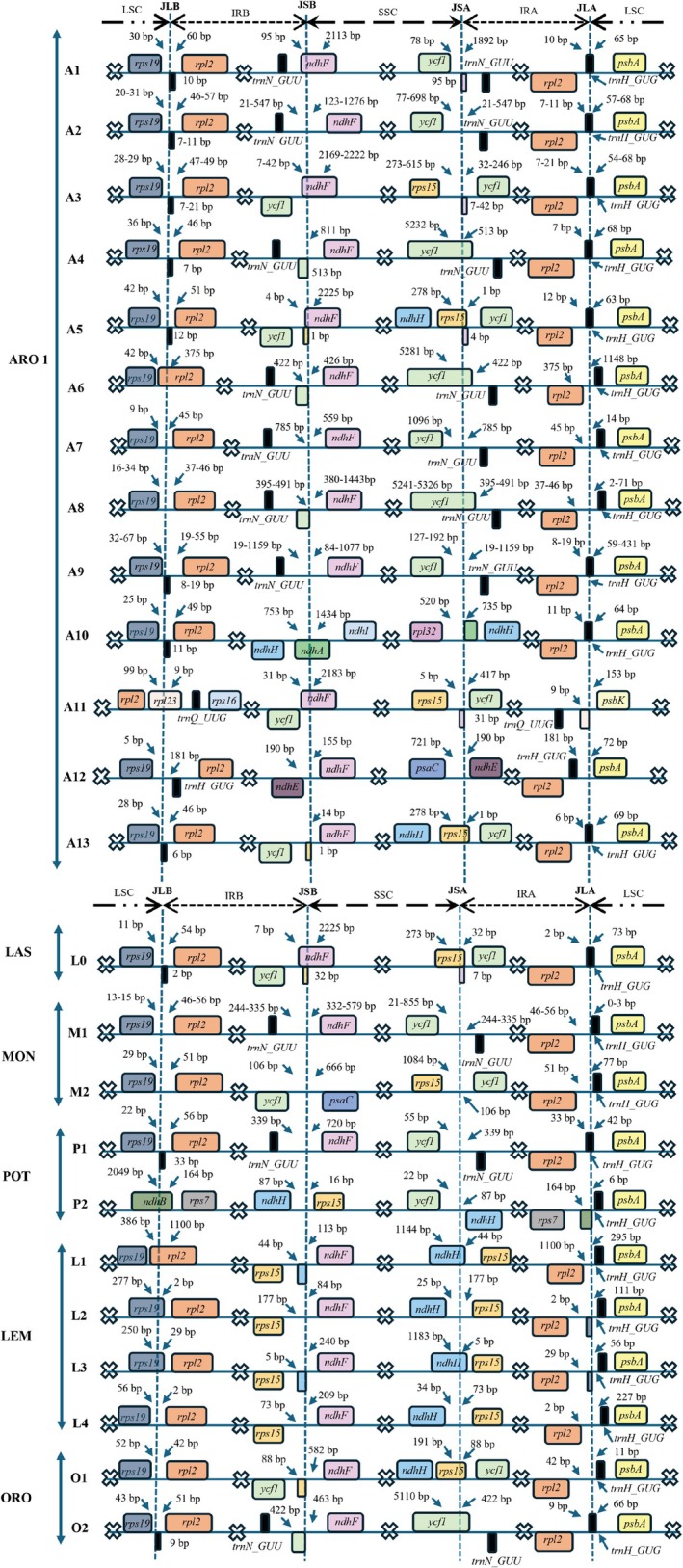 FIGURE 5
FIGURE 5 FIGURE 6
FIGURE 6| Family | Subfamily | Species | Accession number |
|---|---|---|---|
| Araceae | Aroideae |
| |
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Aroideae |
| ||
| Lasioideae |
| ||
| Lemnoideae |
| ||
| Lemnoideae |
| ||
| Lemnoideae |
| ||
| Lemnoideae |
| ||
| Monsteroideae |
| ||
| Monsteroideae |
| ||
| Monsteroideae |
| ||
| Monsteroideae |
| ||
| Orontioideae |
| ||
| Orontioideae |
| ||
| Pothoideae |
| ||
| Pothoideae |
| ||
| Tofieldiaceae |
| ||
|
| |||
|
| |||
| Acoraceae |
| ||
| Groups of genes | Name of genes |
|---|---|
| Ribosomal RNAs |
|
| Transfer RNAs |
|
| Large units of ribosome |
|
| Small units of ribosome |
|
| RNA polymerase |
|
| Translational initiation factor |
|
| Subunit of photosystem I |
|
| Subunit of photosystem II |
|
| Subunit of cytochrome |
|
| Subunit of ATP synthases |
|
| Large unit of Rubisco |
|
| Subunit of NADH dehydrogenase |
|
| Maturase |
|
| Envelope membrane protein |
|
| Subunit of acetyl‐CoA |
|
| C‐type cytochrome synthesis gene |
|
| ATP‐dependent protease subunit P |
|
| Hypothetical proteins and conserved reading frames |
|
|
|
| ||||||
|---|---|---|---|---|---|---|---|
| Type | Repeat Unit/length (bp) | Location | Type | Repeat Unit/length (bp) | Location | ||
| Start | End | Start | End | ||||
| Dinucleotide | TA/12 | 231 | 243 | Dinucleotide | TA/12 | 231 | 243 |
| Mononucleotide | A/10 | 2093 | 2102 | Mononucleotide | A/11 | 2132 | 2142 |
| Mononucleotide | T/10 | 5920 | 5929 | Mononucleotide | T/11 | 5959 | 5969 |
| Mononucleotide | A/10 | 6078 | 6087 | Mononucleotide | A/10 | 6110 | 6119 |
| Mononucleotide | A/14 | 7407 | 7420 | Mononucleotide | A/14 | 7413 | 7426 |
| Mononucleotide | A/10 | 7430 | 7439 | Mononucleotide | A/10 | 7436 | 7445 |
| Mononucleotide | T/10 | 10,980 | 10,989 | Mononucleotide | T/10 | 10,890 | 10,899 |
| Mononucleotide | A/10 | 11,021 | 11,030 | Mononucleotide | A/11 | 10,931 | 10,941 |
| Mononucleotide | T/11 | 11,093 | 11,103 | Mononucleotide | T/10 | 11,004 | 11,013 |
| Mononucleotide | A/11 | 11,482 | 11,492 | Mononucleotide | A/11 | 11,343 | 11,353 |
| Mononucleotide | A/11 | 13,083 | 13,093 | Mononucleotide | A/14 | 12,910 | 12,923 |
| Mononucleotide | T/11 | 13,201 | 13,211 | Mononucleotide | T/11 | 13,031 | 13,041 |
| Dinucleotide | TA/12 | 16,073 | 16,084 | Dinucleotide | TA/12 | 15,901 | 15,912 |
| Mononucleotide | A/10 | 16,101 | 16,110 | Mononucleotide | A/12 | 15,940 | 15,951 |
| Mononucleotide | A/14 | 17,091 | 17,104 | Mononucleotide | A/15 | 16,931 | 16,945 |
| Mononucleotide | T/11 | 19,668 | 19,678 | Mononucleotide | T/11 | 19,507 | 19,517 |
| Mononucleotide | T/11 | 21,861 | 21,871 | Mononucleotide | T/11 | 21,705 | 21,715 |
| Mononucleotide | T/12 | 25,883 | 25,894 | Mononucleotide | T/14 | 25,722 | 25,735 |
| Mononucleotide | T/10 | 29,568 | 29,577 | Mononucleotide | T/10 | 29,409 | 29,418 |
| Mononucleotide | T/11 | 30,718 | 30,728 | Mononucleotide | T/13 | 30,554 | 30,566 |
| Mononucleotide | A/10 | 34,782 | 34,791 | Mononucleotide | A/12 | 34,605 | 34,616 |
| Dinucleotide | TA/12 | 41,070 | 41,082 | Dinucleotide | TA/12 | 40,915 | 40,927 |
| Mononucleotide | T/12 | 41,094 | 41,105 | Mononucleotide | T/14 | 40,937 | 40,950 |
| Mononucleotide | A/10 | 41,684 | 41,693 | Mononucleotide | A/10 | 41,533 | 41,542 |
| Mononucleotide | A/10 | 41,702 | 41,711 | Mononucleotide | A/11 | 41,551 | 41,561 |
| Mononucleotide | T/10 | 47,293 | 47,302 | Mononucleotide | T/10 | 47,143 | 47,152 |
| Mononucleotide | A/12 | 49,236 | 49,247 | Mononucleotide | A/12 | 49,099 | 49,110 |
| Mononucleotide | A/10 | 49,498 | 49,507 | Mononucleotide | A/11 | 49,361 | 49,371 |
| Mononucleotide | A/10 | 50,504 | 50,513 | Mononucleotide | A/10 | 50,373 | 50,382 |
| Dinucleotide | TA/12 | 52,853 | 52,864 | Dinucleotide | TA/18 | 52,465 | 52,482 |
| Dinucleotide | TA/12 | 55,009 | 55,020 | Dinucleotide | TA/12 | 54,633 | 54,644 |
| Mononucleotide | T/11 | 59,495 | 59,505 | Mononucleotide | T/12 | 59,128 | 59,139 |
| Mononucleotide | A/12 | 61,701 | 61,712 | Mononucleotide | A/16 | 61,335 | 61,350 |
| Mononucleotide | T/10 | 61,993 | 62,002 | Mononucleotide | T/10 | 61,629 | 61,638 |
| Mononucleotide | T/10 | 69,091 | 69,100 | Mononucleotide | T/11 | 68,736 | 68,746 |
| Mononucleotide | A/10 | 69,297 | 69,306 | Mononucleotide | A/10 | 68,943 | 68,952 |
| Mononucleotide | A/13 | 69,315 | 69,327 | Mononucleotide | A/11 | 68,961 | 68,971 |
| Dinucleotide | AT/12 | 71,962 | 71,974 | Dinucleotide | AT/12 | 71,612 | 71,624 |
| Mononucleotide | T/11 | 75,308 | 75,318 | Mononucleotide | T/12 | 74,957 | 74,968 |
| Mononucleotide | T/11 | 79,334 | 79,344 | Mononucleotide | T/13 | 79,006 | 79,018 |
| Mononucleotide | T/11 | 82,567 | 82,576 | Mononucleotide | T/10 | 82,242 | 82,251 |
| Mononucleotide | A/13 | 83,381 | 83,393 | Mononucleotide | A/14 | 83,056 | 83,069 |
| Mononucleotide | T/10 | 86,460 | 86,469 | Mononucleotide | T/11 | 86,136 | 86,146 |
| Mononucleotide | A/11 | 86,506 | 86,516 | Mononucleotide | A/11 | 86,183 | 86,193 |
| Mononucleotide | A/11 | 88,327 | 88,337 | Mononucleotide | A/10 | 88,004 | 88,013 |
| Mononucleotide | T/11 | 90,857 | 90,867 | Mononucleotide | T/13 | 90,520 | 90,532 |
| Mononucleotide | T/12 | 91,338 | 91,349 | Mononucleotide | T/11 | 91,003 | 91,013 |
| Dinucleotide | TA/12 | 103,544 | 103,556 | Dinucleotide | TA/12 | 103,208 | 103,220 |
| Mononucleotide | T/12 | 106,409 | 106,420 | Mononucleotide | T/12 | 106,073 | 106,084 |
| Mononucleotide | T/10 | 112,187 | 112,196 | Mononucleotide | T/10 | 111,848 | 111,857 |
| Mononucleotide | A/16 | 117,223 | 117,238 | Mononucleotide | A/18 | 116,891 | 116,908 |
| Trinucleotide | TAA/15 | 118,347 | 118,361 | Trinucleotide | TAA/15 | 118,012 | 118,026 |
| Mononucleotide | T/10 | 126,208 | 126,217 | Mononucleotide | T/11 | 122,104 | 122,114 |
| Mononucleotide | A/11 | 128,528 | 128,538 | Mononucleotide | A/11 | 124,413 | 124,423 |
| Mononucleotide | T/11 | 131,797 | 131,807 | Mononucleotide | T/11 | 127,702 | 127,712 |
| Mononucleotide | T/10 | 132,734 | 132,743 | Mononucleotide | T/10 | 128,629 | 128,638 |
| Mononucleotide | T/11 | 137,709 | 137,719 | Mononucleotide | T/11 | 133,410 | 133,420 |
| Trinucleotide | TTA/15 | 144,730 | 144,744 | Trinucleotide | TTA/15 | 140,234 | 140,248 |
| Mononucleotide | T/16 | 145,853 | 145,868 | Mononucleotide | T/18 | 141,352 | 141,369 |
| Mononucleotide | A/10 | 150,895 | 150,904 | Mononucleotide | A/10 | 146,403 | 146,412 |
| Mononucleotide | A/12 | 156,671 | 156,682 | Mononucleotide | A/12 | 152,176 | 152,187 |
| Dinucleotide | AT/12 | 159,535 | 159,547 | Dinucleotide | AT/12 | 155,040 | 155,052 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| ||||
|
|
|
|
| ||||
| Type/Length (bp) | Sequence | Location in | Location in | ||
|---|---|---|---|---|---|
| Start | End | Start | End | ||
| Forward/34 | TTTGTCTAAGTCACTTCGTTTCTTTTTGTCCAAG | 97,625 | 97,658 | 97,961 | 97,994 |
| 97,670 | 97,703 | 98,006 | 98,039 | ||
| Forward/34 | GATATCGATATTGATGATAGTGACGATATTGATA | 100,086 | 100,119 | 100,422 | 100,455 |
| 100,134 | 100,167 | 100,470 | 100,503 | ||
| Forward/32 | AGACTACACACTACTAATTATATTATATAATA | 133,753 | 133,784 | 138,052 | 138,083 |
| 133,791 | 133,822 | 138,090 | 138,121 | ||
| Forward/29 | ATTTATTTATTATATATTTATTTATTTTT | 177 | 205 | 177 | 205 |
| 31,019 | 31,047 | 31,164 | 31,192 | ||
| Palindromic/29 | AAAAGTAAGAACTCAGCAGGACCGTACCC | 71,917 | 71,945 | 72,267 | 72,295 |
| 71,967 | 71,995 | 72,317 | 72,345 | ||
| Palindromic/26 | TATTTTAATTTTATTTATTATTATAT | 210 | 235 | 210 | 235 |
| 51,599 | 51,624 | 51,734 | 51,759 | ||
| Forward/26 | TTTATTTAATTTATTTATTTAATATT | 30,976 | 31,001 | 31,121 | 31,146 |
| 52,549 | 52,574 | 52,915 | 52,940 | ||
| Forward/25 | GGAGAGAGAGGGATTCGAACCCTCG | 9926 | 9950 | 9936 | 9960 |
| 40,130 | 40,154 | 40,284 | 40,308 | ||
| Forward/25 | TATGTATTTGAAATACAATTGTTAT | 10,755 | 10,779 | 10,845 | 10,869 |
| 10,789 | 10,813 | 10,879 | 10,903 | ||
| Palindromic/25 | AAGTTTTTTTGAGAACCATTTGACT | 124,452 | 124,476 | 128,567 | 128,591 |
| 124,482 | 124,506 | 128,597 | 128,621 | ||
| Palindromic/24 | AGAGAGGGATTCGAACCCTCGGTA | 9930 | 9953 | 9940 | 9963 |
| 50,214 | 50,237 | 50,345 | 50,368 | ||
| Palindromic/23 | GAAGTAATGAACCTCCCAATATG | 82,522 | 82,544 | 82,847 | 82,869 |
| 82,546 | 82,568 | 82,871 | 82,893 | ||
| Palindromic/24 | ATTTTTCTATTTTTTCTTTTAAT | 122,796 | 122,818 | 126,903 | 126,926 |
| 122,828 | 122,850 | 126,934 | 126,957 | ||
| Forward/22 | GATGCGGGTTCGATTCCCGCTA | 12,752 | 12,773 | 12,925 | 12,946 |
| 41,368 | 41,389 | 41,519 | 41,540 | ||
| Palindromic/21 | AATAATATAATAATAAATAAA | 51,595 | 51,615 | 51,730 | 51,750 |
| 126,788 | 126,808 | 130,905 | 130,925 | ||
| Forward/21 | TAATATTAATATAATGTAATA | 66,234 | 66,254 | 66,600 | 66,620 |
| 66,250 | 66,270 | 66,616 | 66,636 | ||
| Palindromic/21 | ATTATATATTATATAATATAA | 75,959 | 75,979 | 76,309 | 76,329 |
| 133,808 | 133,828 | 138,107 | 138,127 | ||
| Forward/21 | TATTATAATTTAAATTTAATT | 118,915 | 118,935 | 123,023 | 123,043 |
| 128,759 | 128,779 | 133,067 | 133,087 | ||
| Forward/20 | ATATTTTATTATATTTCATT | 31,002 | 31,021 | 31,147 | 31,166 |
| 126,765 | 126,784 | 130,882 | 130,901 | ||
| Palindromic/20 | AACCACTCGGCCATCTCTCC | 40,202 | 40,221 | 40,356 | 40,375 |
| 50,156 | 50,175 | 50,287 | 50,306 | ||
| Palindromic/20 | TATATATATTATATAAAATAA | 10,250 | 10,270 | 10,283 | 10,302 |
| 86,145 | 86,165 | 86,468 | 86,487 | ||
| Palindromic/20 | TTTTTTTTTTATTTTATATA | 86,137 | 86,156 | n/a | n/a |
| 128,807 | 128,826 | n/a | n/a | ||
| Forward/20 | AATATCAATATCCAAAATAA | 103,230 | 103,249 | n/a | n/a |
| 103,248 | 103,267 | n/a | n/a | ||
| Forward/20 | TATATATATTTTTATATATT | 31,059 | 31,078 | n/a | n/a |
| 40,919 | 40,938 | n/a | n/a | ||
| Forward/20 | TATATATATATTTAATTATT | 233 | 252 | n/a | n/a |
| 255 | 274 | n/a | n/a | ||
| Forward/21 | ATAATATAACATATATAAATA | 75,971 | 75,991 | n/a | n/a |
| 75,992 | 76,012 | n/a | n/a | ||
| Forward/21 | TAAAAAAAAAAAAGAAATAAA | 34,604 | 34,624 | n/a | n/a |
| 49,098 | 49,118 | n/a | n/a | ||
| Forward/22 | ATATATATATAATATATATATA | 15,902 | 15,923 | n/a | n/a |
| 71,601 | 71,622 | n/a | n/a | ||
| Palindromic/22 | TATTATATTTATTATATTTATA | 41,048 | 41,069 | n/a | n/a |
| 90,276 | 90,297 | n/a | n/a | ||
| Forward/21 | ATTATTATATATATATATTTA | 226 | 246 | n/a | n/a |
| 269 | 289 | n/a | n/a | ||
| Forward/21 | TATATTTAAATTATATATATT | 10,205 | 10,225 | n/a | n/a |
| 10,220 | 10,240 | n/a | n/a | ||
| Forward/23 | ATTATATTATTATATCATACTTA | 126,798 | 126,820 | n/a | n/a |
| 126,831 | 126,853 | n/a | n/a | ||
| Forward/24 | TTATTTATATATTATTTATATATT | 128,536 | 128,559 | n/a | n/a |
| 128,547 | 128,570 | n/a | n/a | ||
| Palindromic/23 | TATTATATTATAATATAAATATA | 10,353 | 10,375 | n/a | n/a |
| 66,857 | 66,879 | n/a | n/a | ||
| Forward/26 | TATATATTTATAATTATATTTTATAT | 35,243 | 35,268 | n/a | n/a |
| 35,266 | 35,291 | n/a | n/a | ||
| Forward/25 | ATTTAAATAATATTTAATTTAAATA | 6025 | 6049 | n/a | n/a |
| 6041 | 6065 | n/a | n/a | ||
| Forward/20 | TAAATATATATTATAATATA | n/a | n/a | 10,438 | 10,457 |
| n/a | n/a | 10,489 | 10,508 | ||
| Palindromic/20 | TATATATATATTTTTTTTTT | n/a | n/a | 41,084 | 41,103 |
| n/a | n/a | 49,498 | 49,517 | ||
| Palindromic/20 | TTTTTCAAAAAAAAAAAAGA | n/a | n/a | 61,695 | 61,714 |
| n/a | n/a | 91,336 | 91,355 | ||
| Forward/20 | AATTATAAATATAATAAATA | n/a | n/a | 90,601 | 90,620 |
| n/a | n/a | 90,664 | 90,683 | ||
| Palindromic/20 | CCTTTTTACGTCCCCATGTC | n/a | n/a | 111,600 | 111,619 |
| n/a | n/a | 111,633 | 111,652 | ||
| Palindromic/20 | TATTAATATATAAAATAAAA | n/a | n/a | 133,109 | 133,128 |
| n/a | n/a | 138,653 | 138,672 | ||
| Forward/20 | ATATTTCATTTATTTATTAT | n/a | n/a | 31,157 | 31,176 |
| n/a | n/a | 132,594 | 132,613 | ||
| Palindromic/20 | ATAAAATAAAAAAAAAAGAA | n/a | n/a | 34,775 | 34,794 |
| n/a | n/a | 86,457 | 86,476 | ||
| Palindromic/20 | ATAATATATATTTATATTAT | n/a | n/a | 262 | 281 |
| n/a | n/a | 122,291 | 122,310 | ||
| Forward/21 | TATAACAATATATAATTTATA | n/a | n/a | 122,069 | 122,089 |
| n/a | n/a | 122,086 | 122,106 | ||
| Forward/21 | TATATATATATATAATATAAA | n/a | n/a | 122,659 | 122,679 |
| n/a | n/a | 122,694 | 122,714 | ||
| Forward/21 | ATTTATTTATTATTTATATTT | n/a | n/a | 66,471 | 66,491 |
| n/a | n/a | 132,601 | 132,621 | ||
| Palindromic/22 | TATATATATATAATATATATAT | n/a | n/a | 55,009 | 55,030 |
| n/a | n/a | 122,649 | 122,670 | ||
| Forward/22 | TATTTATAATTTTATAATATAT | n/a | n/a | 138,499 | 138,520 |
| n/a | n/a | 138,520 | 138,541 | ||
| Palindromic/21 | ATATATTATATAATATATATT | n/a | n/a | 10,271 | 10,291 |
| n/a | n/a | 122,299 | 122,319 | ||
| Palindromic/21 | AAATATAAATATAATTATAAA | n/a | n/a | 11,576 | 11,596 |
| n/a | n/a | 41,033 | 41,053 | ||
| Forward/22 | AACTATGACTATAAATAATAAA | n/a | n/a | 6110 | 6131 |
| n/a | n/a | 6130 | 6151 | ||
| Forward/22 | ACCTTTTTTAATAATAATATTA | n/a | n/a | 10,094 | 10,115 |
| n/a | n/a | 10,131 | 10,152 | ||
| Forward/22 | ATATAAAATAAATATTAAAATA | n/a | n/a | 11,506 | 11,527 |
| n/a | n/a | 66,544 | 66,565 | ||
| Palindromic/23 | ATAAATATAATTAAATATAATAT | n/a | n/a | 11,432 | 11,454 |
| n/a | n/a | 52,600 | 52,622 | ||
| Palindromic/23 | ATATATATATAATATATATATAT | n/a | n/a | 71,951 | 71,973 |
| n/a | n/a | 122,647 | 122,669 | ||
| Forward/25 | ATATTATAATATATAAAATATAAAT | n/a | n/a | 10,458 | 10,482 |
| n/a | n/a | 10,496 | 10,520 | ||
| Forward/27 | ATTATTATAATAATTATAATAAAATAT | n/a | n/a | 122,584 | 122,610 |
| n/a | n/a | 122,609 | 122,635 | ||
| Forward/26 | ATATAAATATAATTATAAATATAATA | n/a | n/a | 11,578 | 11,603 |
| n/a | n/a | 90,591 | 90,616 | ||
| Forward/38 | ATATATTAAATATTATATTATTATAATTATATATATTA | n/a | n/a | 138,549 | 138,586 |
| n/a | n/a | 138,579 | 138,616 | ||
| Forward/21 | ATTTATATATATATATTTATA | n/a | n/a | 41,067 | 41,087 |
| n/a | n/a | 138,466 | 138,486 | ||
- —Trường Đại học Trà Vinh10.13039/100018444
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Plant Diversity and Evolution · Plant and Fungal Species Descriptions
Introduction
1
Homalomena Schott, a member of the Araceae family, contains 163 accepted species that are distributed in South Asia, East Asia, Southeast Asia, and Melanesia (Plants of the World Online 2025). Recently, various new Homalomena species have been identified, including Homalomena renda, Homalomena belitungensis A.S.D.Irsyam & M.R.Hariri from Indonesia, Homalomena punctifolia K.Z.Hein and Naive and Homalomena sigmoidea K.Z.Hein and Naive from Myanmar, and Homalomena perplexa K.Z.Hein, Vuong, Bao & V.S.Dang from Vietnam (Dang et al. 2023; Hein and Naive 2024; Hariri and Irsyam 2025; Irsyam et al. 2025). These new species indicate a potentially high biodiversity of the Homalomena genus and need further investigations. In addition, different compounds such as monoterpenes and linalool have been identified in Homalomena species extraction, which exhibited antimicrobial, antifungal, and anti‐inflammatory activities (Rozman et al. 2018; Dam and Van 2022; Nguyen, Doan, et al. 2023; Nguyen, Nguyen, et al. 2023; Gupta et al. 2025; Tamang et al. 2025). In the case of H. perplexa , the essential oil extraction from leaves and rhizomes contained various compounds (e.g., sabinene, limonene, and linalool) which displayed antibacterial activities (Truong, Nguyen, et al. 2025; Truong, Van, et al. 2025). Although high diversity and pharmacological features of Homalomena have been intensively studied, the genomic data (e.g., complete organelle and nuclear genomes) of Homalomena species are still scarce. Therefore, more genomic studies examining Homalomena species should be conducted.
Chloroplast is an essential organelle of land plants, which contains a circular genome (consisting of a large‐single copy (LSC), a small‐single copy (SSC), and two identical inverted repeat (IR) regions) encoding genes responsible for photosynthesis (Dobrogojski et al. 2020). Within the Araceae family, several complete chloroplast genomes have been reported (Abdullah, Henriquez, Mehmood, Shahzadi, et al. 2020; Henriquez et al. 2020; Abdullah et al. 2021; Li et al. 2025). These chloroplast genomes revealed polymorphic loci, repeat content, and genomic features across aroid species. In addition, the chloroplast genome provided useful data for mining molecular markers, exploring selective pressure, and phylogenetic relationships among Araceae species (Li et al. 2022). However, there has not been a comprehensive study examining whole chloroplast genomes of all Araceae subfamilies so far. Although there are more than 100 accepted Homalomena species, only one complete chloroplast genome of Homalomena occulta has been reported (Zhang et al. 2021). The lack of genomic data limits the exploration of evolutionary history among Homalomena species. Therefore, in the current study, we sequenced and characterized the complete chloroplast genome of H. perplexa , an aroid species endemic to Vietnam. This study aims to (1) compare the chloroplast genome features between H. perplexa and H. occulta, focusing on genone structure, gene content, and repeat composition, (2) locate hypervariable regions in Homalomena chloroplast genomes, (3) explore variations of junctions between LSC, SSC, and IR regions in Araceae chloroplast genomes, and (4) reconstruct phylogenetic relationships among Araceae subfamilies based on 79 protein‐coding regions. The results of this study provide new insights into chloroplast genome features of Homalomena species and an update on comparative chloroplast genome analysis of Araceae.
Materials and Methods
2
Plant Material, DNA Extraction, and Sequencing
2.1
The whole plant of H. perplexa was transplanted from Con Dao, Ho Chi Minh City to Tra Vinh University, Vinh Long province (Figure 1). The living sample of H. perplexa was available at Tra Vinh University, Vinh Long province (contact person: Nguyen Nhat Nam, [email protected]). No permission is required to collect the sample of H. perplexa . A healthy leaf was collected and dried using silica gel beads. The dried sample was stored in a deep freezer at −81°C before being used to extract total genomic DNA using Dneasy Plant Pro Kit (Qiagen, USA). The quality of DNA sample was checked using a NanoDrop One Microvolume UV–Vis Spectrophotometer (Thermo Fisher Scientific, USA) and 1% agarose gel electrophoresis. The DNA sample which had a clear band on agarose gel and a concentration over 100 ng/μl was used to prepare a library for sequencing using a Native Barcoding Kit 24 V14 (SQK‐NBD114.24, Oxford Nanopore Technologies, UK) following the manufacturer's protocol. The library was then sequenced using a PromethION flow cell (R10.4.1, FLO‐PRO114M) and a PromethION 2 Solo device, monitored by MINKNOW v24.11.10 with a High Accuracy option for the basecalling process. The quality of output reads in fastq format was checked using nanoplot v1.41.6 and then filtered using Filtlong v0.3.1 (https://github.com/rrwick/Filtlong) to remove reads that are shorter than 1000 bp and have a Qscore lower than 8 (De Coster and Rademakers 2023). Consequently, among 424,520 raw reads (accounting for 2.08 GB), 313,902 reads (composing 1.87 GB) were kept for further analyzes.
Photos of Homalomena perplexa. (A) Whole plant collected at Con Dao, Ho Chi Minh City. (B) Whole plant transplanted at Tra Vinh University, Vinh Long province. (C) Blooms with slender peduncles and green spathes. (D) Bloom at pistillate anthesis. The photos were taken by Nhat Nam Nguyen.
Genome Assembly and Annotation
2.2
The filtered reads were assembled to complete the chloroplast genome of Homalomena occulta (MW145396) to isolate chloroplast‐related reads using minimap2 with default settings (Li 2018). Then, the isolated reads were assembled using Flye v2.9.6 with the settings of ‐‐nano‐corr, ‐i 10, ‐g 0.17 m, and ‐t 60 to complete the chloroplast genome of H. perplexa (Kolmogorov et al. 2019). The complete chloroplast genome of H. perplexa was annotated using GeSeq (https://chlorobox.mpimp‐golm.mpg.de/geseq.html) with the default settings for the chloroplast genome (Tillich et al. 2017). The annotation of protein‐coding genes and transfer RNAs was confirmed using Geneious Prime v2025.0 for start and stop codons and tRNAscan‐SE, respectively (Chan and Lowe 2019). The map of the chloroplast genome was illustrated using OrganellarGenomeDRAW (OGDRAW) version 1.3.1 with default settings (Greiner et al. 2019). The complete chloroplast genome of H. perplexa was submitted to the GenBank database under accession number PX569150.
Comparative Chloroplast Genome Analyzes
2.3
For finding small sequence repeats (SSRs) in two chloroplast genomes of Homalomena species, Phobos embedded in Geneious Prime v2025.0 with a minimum threshold of 10, 12, 15, 20, and 24 bp for mononucleotide repeats, dinucleotide repeats, trinucleotide repeats, pentanucleotide repeats, and hexanucleotide repeats, respectively. Similarly, long repeats (e.g., forward, complement, reverse, and palindromic types) were located using the “Find Repeat” function in Genenious Prime v2025.0 with a minimum length of 20 bp. Nucleotide diversity analysis among two chloroplast genomes of H. occulta and H. perplexa was conducted using DnaSP 6 with a window length of 1000 and a step size of 200 (Rozas et al. 2017). The junctions among the large‐single copy, small‐single copy, and inverted repeat regions were located using IRplus and manually illustrated for 46 available complete chloroplast genomes, representing 45 genera of Araceae (Table 1) (Díez Menéndez et al. 2023).
Phylogenetic Analysis Among Araceae Species
2.4
The available complete chloroplast genome of 46 aroid species representing 45 genera of Araceae and four Tofieldiaceae and Acoraceae species serving as outgroups were retrived from the GenBank database (Table 1). Then, 79 protein‐coding regions were extracted from the chloroplast genomes using Geneious Prime v2025.0.2 before being aligned using MUSCLE v5.3 with defautl settings (Edgar 2022). TrimAl v1.5.1 was used to remove gaps in the aligned sequences (Capella‐Gutiérrez et al. 2009). The trimmed dataset was subjected to jModeltest 2 to find the best substitution model which was identified as GTR + I + G (Darriba et al. 2012). For reconstructing phylogenetic relationships among aroid species, maximum likelihood (ML) and Bayesian inference (BI) methods were employed. For ML method, the IQ‐TREE 2 was used with settings of 1000 bootstraps alignments and substitution model of GTR + I + G (Minh et al. 2020). Similarly, for BI analysis, MrBayes v 3.2.7a was used with settingd of GTR + I + G substitution model, 3,000,000 generations, and 25% discarded samples (Ronquist et al. 2012). The phylogenetic trees were illustrated using Figtree v1.4.5 (https://github.com/rambaut/figtree/releases).
Results
3
Features of
H. perplexa Chloroplast Genome
3.1
The complete chloroplast genome of H. perplexa (with average coverage depth of 433×) was 169,893 bp in length and had 35.4% GC content (Figures 2 and 3A). This circular genome consisted of a large‐single copy (LSC, 93,197 bp in length and 33.4% GC content), a small‐single copy (SSC, 20,014 bp in length and 27.3% GC content), and two identical inverted repeat regions (IR, 28,341 bp in length and 41.4% GC content each) (Figure 2). In Addition, there were 113 unique coding regions in this quadripartite genome, including 79 protein‐coding genes, 30 transfer RNA genes, and four ribosomal RNA genes (Figure 2, Table 2). Among coding regions, pafI and clpP1 genes had two introns whereas rps16, atpF, rpoC1, petB, petD, rpl16, rpl2, ndhB, ndhA, trnK_UUU, trnI_GAU, trnA_UGC, trnG_UCC, trnL_UAA, and trnV_UAC each had one intron (Figure 3B). The rps12 gene was transpliced, of which the exon 1 was in the LSC region whereas the exon 2 and exon 3 were in the IR regions (Figure 3C). Due to the presence of IR regions, rpl2, rpl23, trnI_CAU, ycf2, trnL_CAA, ndhB, rps7, rps12, trnV_GAC, rrn16, trnI_GAU, trnA_UGC, rrn23, rrn4.5, rrn5, trnR_ACG, trnN_GUU, and ycf1 were duplicated in the IR region, but ycf1 was present as an incomplete duplication (Figure 2, Table 2).
Chloroplast genome map of Homalomena perplexa. The inner and outer genes indicate the clockwise and counterclockwise translation directions, respectively. The inner circle indicates four regions of the chloroplast genome. IRA and IRB, Inverted repeat regions A and B, respectively; LSC, Large single‐copy region; SSC, Small single‐copy region.
Coverage depth and trans‐splicing and cis‐splicing gene content in chloroplast genome of Homalomena perplexa. (A) Coverage depth of H. perplexa chloroplast genome. (B) Cis‐splicing genes of H. perplexa chloroplast genome. C. Trans‐splicing gene of H. perplexa chloroplast genome.
In the H. perplexa chloroplast genome, there were 81 SSRs composed of 63 mononucleotide repeats, 16 dinucleotide repeats, and two trinucleotide repeats (Table 3). All these repeats were made up of A and T nucleotides. Similarly, H. occulta chloroplast genome possessed 79 SSRs, including 63 mononucleotide repeats, 12 dinucleotide repeats, and four trinucleotide repeats. Most of the SSRs in the H. occulta chloroplast genome contained A and T nucleotides, except one trinucleotide repeat made up of the TCC unit. Among identified SSRs, there were 62 shared repeats between two Homalomena chloroplast genomes, which included 52 mononucleotide repeats, eight dinucleotide repeats, and two trinucleotide repeats. However, H. occulata had four and two unique dinucleotide and trinucleotide repeats, respectively. Meanwhile, H. perplexa had eight unique dinucleotide repeats. There were no records of tetranucleotide, pentanucleotide, and hexanucleotide repeats in the two Homalomena chloroplast genomes examined. In addition, the length of mononucleotide repeats ranged from 10 to 16 bp in H. perplexa and from 10 to 18 bp in H. occulta. Similarly, H. occulta had 12–18 bp dinucleotide repeats, whereas H. perplexa contained 12–14 bp dinucleotide repeats. The length of trinucleotide repeats (15 bp) was identical in both Homalomena species (Table 3).
Analysis of long repeats revealed the presence of forward and palindromic repeats in the chloroplast genomes of H. occulta and H. perplexa (Table 4). There were 23 forward repeats (ranging from 20 to 34 bp) and 13 palindromic repeats (ranging from 20 to 29 bp) in H. occulta. Meanwhile, H. perplexa had 26 forward repeats (ranging from 20 to 38 bp) and 21 palindromic repeats (ranging from 20 to 23 bp). Between H. occulta and H. perplexa there were 11 shared forward repeats and 10 shared palindromic repeats, which ranged from 20 bp to 34 bp. In addition, there were 26 and 15 unique long repeats in H. perplexa and H. occulta chloroplast genomes, respectively (Table 4). Notably, there were no reverse and complement repeats in the two examined chloroplast genomes of Homalomena. Comparative analysis between H. occulta and H. perplexa chloroplast genomes revealed a high similarity (Figure 4). Only two regions (including trnH_GUG‐psbA and petA‐psbJ) had Pi values over 0.015. In contrast, most of the IRA and IRB regions had Pi values equal to 0. Other regions possessed Pi values under 0.005.
Nucleotide diversity among two Homalomena chloroplast genomes. Nucleotide positions are shown on the x‐axis and Pi values are shown on the y‐axis. IRA and IRB, Inverted repeat regions A and B, respectively; LSC, Large‐single copy region; SSC, Small‐single copy region.
Variations of Junctions Among the LSC, SSC and IR Regions in Araceae Chloroplast Genomes
3.2
Comparative genome structure analysis among 46 examined Araceae species revealed different junctions among LSC, SSC, and IR regions (Figure 5). In Aroideae, there were 13 junction variations inferred from 32 examined genera, whereas two variations were identified in four genera of Monsteroideae. In Lemnoideae, Orontioideae, Pothoideae, and Lasioideae, the number of junction variations was identical to the number of examined genera, which were 4,2,2, and 1, respectively. Although various junction variations were found, the gene content at the boundaries was quite similar. Specifically, the junction between the LSC and IRB regions (JLB) located in the intergenic spacer (IGS) between the rps19 and rpl22 genes. In some cases, the JLB moved to coding regions of rpl2 (i.e., L1 of Lemnoideae), rps19 (i.e., L2 and L3 of Lemnoideae), rpl23 (i.e., A11 of Aroideae), and ndhB (i.e., P2 of Pothoideae). In the case of the junction between the LSC and IRA regions (JLA), there was a shift of trnH_GUG from the LSC region to the IRA region until the whole trnH_GUG was located in the IRA region (i.e., A11 of Aroideae). Notably, due to the expansion of the IR region to the LSC region in Anchomanes hookeri (A11 of Aroideae), the JSA contained partial ndhF and near psbK. Similar to JLA and JLB, the junctions among SSC, IRA, and IRB regions (JSA and JSB) exhibited a movement to include ycf1 and other genes in the SSC region. For example, the JSA located in the IGS between ycf1 and trnN_GUU (i.e., A2, A7, and A9 of Aroideae) and then found in the coding region of ycf1 (i.e., A4, A6, and A8 of Aroideae). Continuously, the JSA was in the IGS between ycf1 and rps15 (i.e., A3 and A11 of Aroideae) and located within ndhA (i.e., A10 of Aroideae) before being identified in the coding region of rps15 (i.e., A5 and A13 of Aroideae). Notably, in A12 variation (belonging to Zantedeschia rehmannii), the JSA was in the IGS between psaC and ndhE. In the JSB, there were also records of ndhF overlap in A1, A3, A5, and A11 variations, which ranged from 4 to 95 bp. A similar trend of JSA and JSB was found in the chloroplast genomes of other Araceae subfamilies (Figure 5).
Junctions among LSC, SSC, and IR regions of Araceae species. JLB: Junction between LSC and IRB regions. JSB: Junction between SSC and IRB regions. JSA: Junction between SSC and IRA regions. JLA: Junction between LSC and IRA regions. The vertical dashed lines indicate the junction sites. The lines represent the sequences and regions and do not show exact length‐based ratios in the chloroplast genomes. Multiplication signs indicate other contents in the chloroplast genomes.ARO, Aroideae; LAS, Lasioideae; LEM, Lemnoideae; MON, Monsteroideae; ORO, Orontioideae; POT, Pothoideae.
Phylogenetic Relationships Among Examined Areaceae Species
3.3
Phylogenetic analysis inferred from 79 protein‐coding genes using ML and BI methods resulted in identical topology of phylogenetic trees (Figure 6). The monophyly of Araceae subfamilies was highly supported with posterior probability = 1 and bootstrap value = 100. Within Aroideae, there were two distinct clades with strong support values. However, phylogenetic relationships among *Ariseama bockii, Pinellia pedatisecta, Aglaonema commutatum, Anchomanes hookeri, Calla palustris *, and related species in Aroideae exhibited low and moderate support values. Homalomena perplexa formed a clade with H. occulta which was sister to Furtadoa mixta with high support values (PP = 1 and BS = 100).
Phylogenetic tree in cladogram format of 46 Araceae species inferred from 79 plastid protein‐coding genes using maximum likelihood and Bayesian inference methods. Only posterior probability ≤ 1 and bootstrap values ≤ 100 were shown at the nodes. ARO, Aroideae; LAS, Lasioideae; LEM, Lemnoideae; MON, Monsteroideae; ORO, Orontioideae; POT, Pothoideae.
Discussion
4
The complete chloroplast genome of H. perplexa revealed a conserved pattern regarding gene content and genome structure, which was found in other angiosperms (Dobrogojski et al. 2020). Within Araceae, the quadripartite structure and a unique profile of 79 protein‐coding genes, 30 tRNAs, and four rRNAs were commonly found in the published chloroplast genomes (Abdullah, Henriquez, Mehmood, Shahzadi, et al. 2020; Henriquez et al. 2020; Abdullah et al. 2021; Zhang et al. 2021; Li et al. 2022, 2025). However, gene loss has been recorded in the chloroplast genomes of Amorphophallus species (Liu et al. 2019; Li et al. 2024). In particular, the ycf1 gene was absent in Amorphophallus bulbifer, whereas Amorphophallus konjac did not have psbE and trnG_GCC. In addition, trnL‐CAA was lost in Amorphophallus muelleri and A. bulbifer and only A. muelleri had the accD gene. The infA gene was only detected in Amorphophallus titanum (Li et al. 2024). Similarly, within Araceae, the loss of infA was found in Colocasia, Dieffenbachia, Spirodela, Lemna, Wolffiella, and Wolffia (Choi et al. 2017). These results indicate a dynamic pattern of gene loss in the Amorphophallus genus and other Araceae genera. Although gene loss was not observed in the current study, many unexplored Homalomena chloroplast genomes may reveal different genomic events. Furthermore, the impacts of gene loss in Araceae require further investigations.
Although the chloroplast genomes of H. perplexa and H. occulta showed a high similarity in terms of shared repeats (62 SSRs and 21 long repeats), they still had unique repeat sequences. Previously, dynamic repeat contents have been characterized among Araceae chloroplast genomes; however, the shared and unique repeats have not been assessed (Choi et al. 2017; Liu et al. 2019; Abdullah, Henriquez, Mehmood, Carlsen, et al. 2020). The lack of information about shared and unique repeats might be caused by the deficiency of chloroplast genomes among the examined plant genera and families. Due to a dramatic increase in chloroplast genome data recently, the shared and unique repeats in different genera and families should be analyzed to provide a new aspect of genomic evolution in land plants. In chloroplast genomes, repeats are useful sources for exploring population genetics, gene flow, and genetic diversity (George et al. 2015; Feng et al. 2023; Huy et al. 2025; Moosavi et al. 2025). The shared and unique repeats in Homalomena chloroplast genomes are helpful in tracing the genomic evolution within the genus and among its species.
In addition to gene loss and conservation of quadripartite structure, Araceae chloroplast genomes possessed high variations in the junctions among LSC, SSC, and IR regions. A previous study indicated that the LSC/SSC/IR junctions were diverse according to plant groups (i.e., angiosperms, gymnosperms, ferns, lycophytes, hornworts, mosses, liverworts, and green algae) (Zhu et al. 2016). In the current study, the LSC/SSC/IR junctions of Araceae exhibited the features of angiosperms, which expanded to include rps19 genes. Notably, the junction variation occurred in three directions, including the expansion from the IR regions to LSC region, from the IRA region to SSC and from the IRB region to the SSC region. Consequently, different junction variations were observed in Aroideae (13 variations), Pothoideae (two variations), Monsteroideae (two variations), and Lemnoideae (four variations). Although Aroideae has the largest number of variations, it does not indicate that Aroideae chloroplast genomes are more diverse than other subfamilies. The reason for the outstanding Aroideae is the availability of chloroplast genomes (belonging to 32 genera) compared to other subfamilies (fewer than five genera). Therefore, more aroid genera and species should be included to examine the junction variations. Within the Araceae genus, junction variations were also found in 13 Amorphophallus chloropalst genomes (Li et al. 2024). The two chloroplast genomes of Homalomena in the current study did not show a significant variation; however, the unexplored Homalomena species may contain more variations and genomic events that require further investigation.
Previously, a phylogenetic study of 102 aroid genera based on rbcL, matK, partial trnK intron, partial tRNA‐Leu gene, trnL–trnF spacer, and partial tRNA‐Phe gene revealed the backbone phylogeny of Araceae (Cusimano et al. 2011). Specifically, Gymnostachydoideae and Orontoideae formed a basal clade of Araceae. Pothoideae was close to Monsteroideae. Meanwhile, Zamioculcadoideae was embedded in Aroideae. In 2014, 70 plastid protein‐coding genes in 37 genera of Araceae were employed to reconstruct the phylogenetic relationships of which the infrafamilial relationships were similar to the previous result, except for the absence of Gymnostachydoideae (Henriquez et al. 2014). Recently, the plastid data set covering 79 Araceae genera and Angiosperms 353 nuclear data representing 111 Araceae genera were used to confirm the infrafamilial classification of Araceae (Haigh et al. 2023). The results from nuclear and plastid data showed similar relationships among subfamilies, except for the absence of Gymnostachydoideae in the plastid dataset. Consequently, they suggested the expansion of Aroideae to include Zamioculcadoideae and proposed a seven‐subfamily system of Araceae, of which Gymnostachydoideae and Orontoideae was basal clade of Araceae. In the current study, although the phylogenetic analysis did not reveal any alterations in infrafamilial relationships compared to previous studies, the data of Gymnostachydoideae were still absent. Therefore, the complete chloroplast genome of Gymnostachys anceps, a monotypic species of Gymnostachydoideae, is required for fulfilling the phyogenetic relationships of Araceae based on chloroplast genome data.
Author Contributions
Nhat Nam Nguyen: conceptualization (equal), formal analysis (equal), methodology (equal), supervision (equal), writing – original draft (equal). Ngoc Trai Nguyen: formal analysis (equal), writing – original draft (equal). Hoang Dang Khoa Do: conceptualization (equal), methodology (equal), supervision (equal), writing – review and editing (equal).
Funding
This work was supported by Tra Vinh University (TVU) under grant contract number 60/2025/HĐ.HĐKH&ĐT—ĐHTV.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdullah , C. L. Henriquez , F. Mehmood , et al. 2020. “Complete Chloroplast Genomes of Anthurium Huixtlense and Pothos Scandens (Pothoideae, Araceae): Unique Inverted Repeat Expansion and Contraction Affect Rate of Evolution.” Journal of Molecular Evolution 88, no. 7: 562–574. 10.1007/s 00239-020-09958-w.32642873 PMC 7445159 · doi ↗ · pubmed ↗
- 2Abdullah , C. L. Henriquez , F. Mehmood , et al. 2020. “Comparison of Chloroplast Genomes Among Species of Unisexual and Bisexual Clades of the Monocot Family Araceae.” Plants 9, no. 6: 737. 10.3390/plants 9060737.32545339 PMC 7355861 · doi ↗ · pubmed ↗
- 3Abdullah , C. L. Henriquez , F. Mehmood , et al. 2021. “Chloroplast Genome Evolution in the Dracunculus Clade (Aroideae, Araceae).” Genomics 113, no. 1: 183–192. 10.1016/j.ygeno.2020.12.016.33326831 · doi ↗ · pubmed ↗
- 4Capella‐Gutiérrez, S. , J. M. Silla‐Martínez , and T. Gabaldón . 2009. “trim Al: A Tool for Automated Alignment Trimming in Large‐Scale Phylogenetic Analyses.” Bioinformatics 25, no. 15: 1972–1973. 10.1093/bioinformatics/btp 348.19505945 PMC 2712344 · doi ↗ · pubmed ↗
- 5Chan, P. P. , and T. M. Lowe . 2019. “t RN Ascan‐SE: Searching for t RNA Genes in Genomic Sequences.” In Gene Prediction. Methods in Molecular Biology, edited by M. Kollmar , 1–14. Humana. 10.1007/978-1-4939-9173-0_1.PMC 676840931020551 · doi ↗ · pubmed ↗
- 6Choi, K. , K. Park , and S. Park . 2017. “The Chloroplast Genome of Symplocarpus Renifolius: A Comparison of Chloroplast Genome Structure in Araceae.” Genes 8, no. 11: 324. 10.3390/genes 8110324.29144427 PMC 5704237 · doi ↗ · pubmed ↗
- 7Cusimano, N. , J. Bogner , S. J. Mayo , et al. 2011. “Relationships Within the Araceae: Comparison of Morphological Patterns With Molecular Phylogenies.” American Journal of Botany 98, no. 4: 654–668. 10.3732/ajb.1000158.21613165 · doi ↗ · pubmed ↗
- 8Dam, S. M. , and H. T. Van . 2022. “Chemical Profiles and Biological Activities of Essential Oils of Arisaema and Homalomena Species (Araceae) – A Review.” Journal of Phytology 14: 41–49. 10.25081/jp.2022.v 14.7444. · doi ↗