Identification of endophytic Trichoderma species (Hypocreaceae, Hypocreales) and their application against leaf blight of host Platycladus orientalis

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
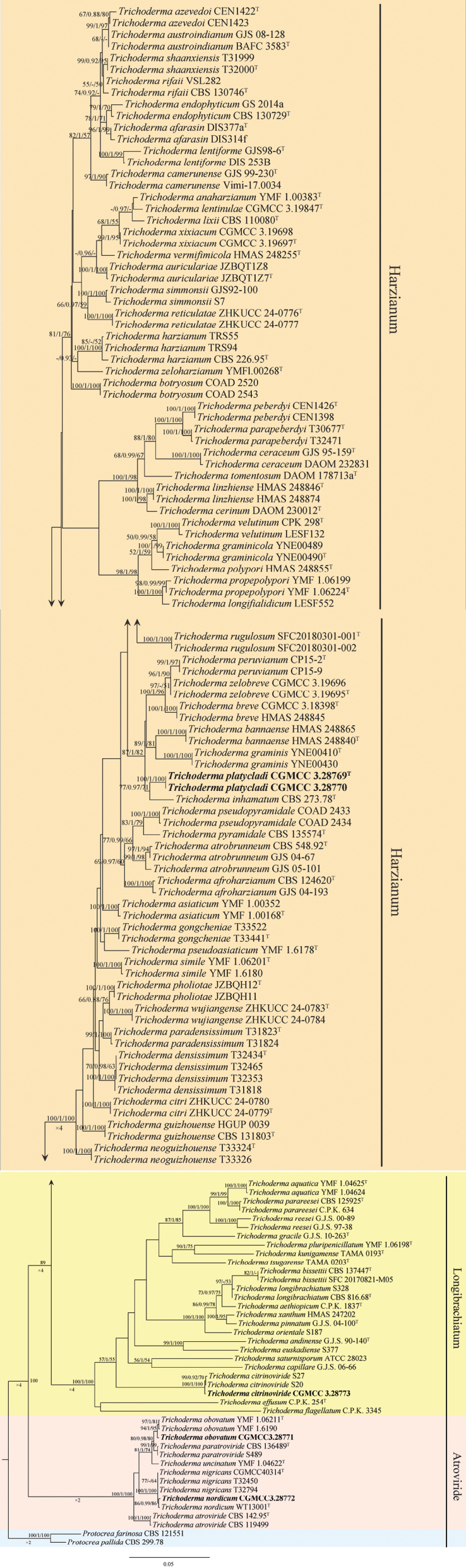 Figure 1
Figure 1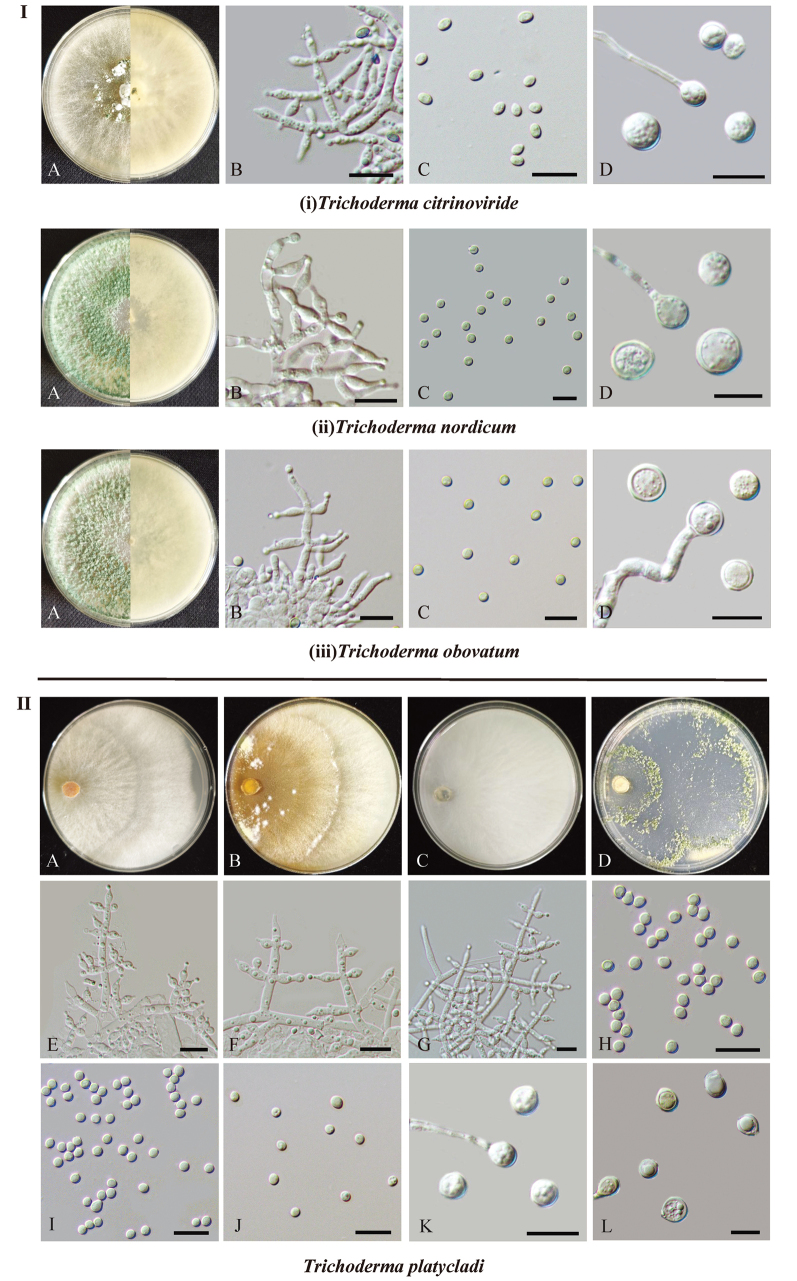 Figure 2
Figure 2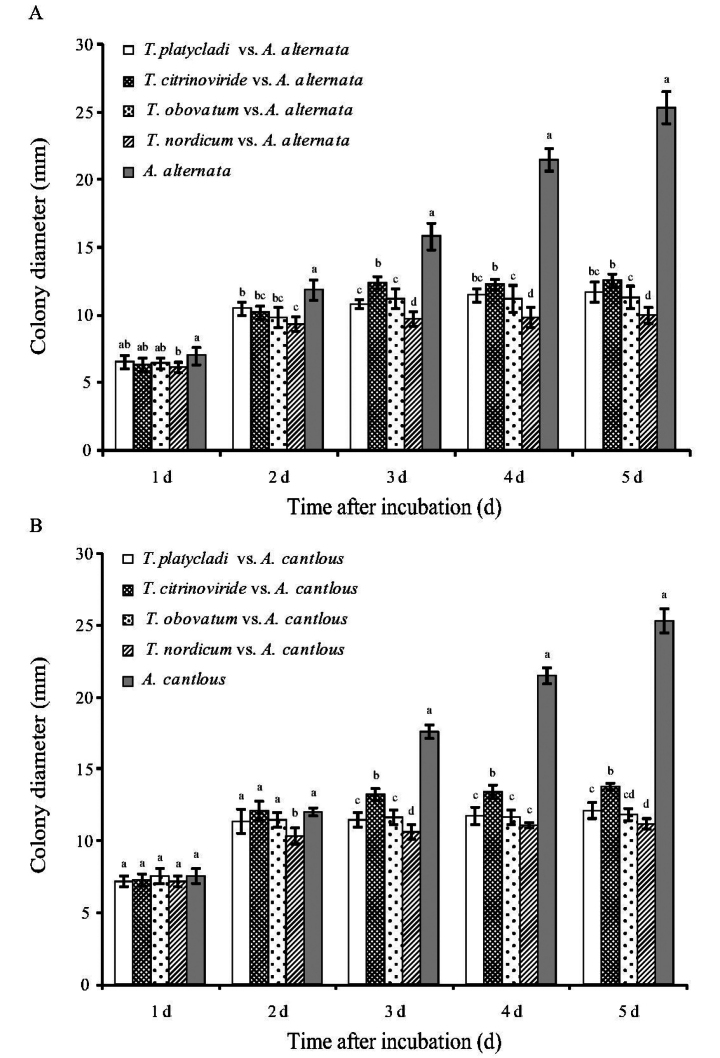 Figure 3
Figure 3| Disease grade | Grading standard | Representative value |
|---|---|---|
| 0 | Asymptomatic | 0 |
| I | The area of disease spots occupies 1 to 5% of the foliar area | 1 |
| II | The area of disease spots occupies 5 to 10% of the foliar area | 2 |
| III | The area of disease spots occupies 10 to 20% of the foliar area | 3 |
| IV | The area of disease spots occupies 20 to 50% of the foliar area | 4 |
| V | Leaf disease spots area > 50% | 5 |
| Species | Strain | Colony character on | Conidiophores | Phialides | Conidia | Chlamydospores | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Description of colonial morphology | Diameters (mm/3 days) | Character | Size (μm) | Character | Size (μm) | Character | Size (μm) | Character | ||
|
| CGMCC3.28773 | White at first, quickly shading to green, occasionally forming compact, hemispherical pustules up to 10 mm in diameter, odor indistinct. | 59–62 | Irregularly shaped tufts, main branches long and relatively straight, side branches arising at near right angles, usually arising singly and at irregular intervals, or occasionally in opposite pairs, rarely branched. | 3.6–6.8 × 2.4–3.0 (n = 30) | lageniform to ampulliform | 2.9–3.7 × 1.8–2.4 (n = 50) | dilute green, more or less ellipsoidal | 5.8–7.0 × 3.5–5.0 (n = 30) | single or in pairs, globose to subglobose |
|
| CGMCC3.28772 | Grey-white at first and slowly turning green, diffusing pigment or distinctive odour absent. | 67–71 | Branches narrow and flexuous, typically forming a pyramidal structure with regularly whorled branches, each branch terminating in a cruciate whorl of up to 5 phialides. | 7.5–11.2 × 2.8–3.3 (n = 30) | lageniform | 4.0–4.8 × 4.0–4.3 (n = 50) | green, globose to obovoidal | 8.5–10.0 × 8.5–9.7 (n = 30) | sometimes present, globose to subglobose |
|
| CGMCC3.28771 | Pale green to green, margin dense, sparse in the middle, floccose, homoge neous, indistinctly zonate, aerial hyphae abundant, especially pleasant odor noted. | 80–85 | Straight or curved, emerging in right angles or oriented towards to the conidiophore axis, typically with 1–2 branched levels, side branches simple. | 5.0–9.0 × 2.8–4.0 (n = 30) | ampuliform to lageniform, sometimes nearly round | 3.2–4.0 × 3.0–3.5 (n = 50) | green, globose, oval to obovate | 7.0–8.8 × 6.8–8.2 (n = 30) | single, terminal or intercalary, globose to subglobose |
|
| CGMCC3.28769, CGMCC3.28770 | Radial, white, fluffy, aerial hyphae dense, abundant, yellow-green and white conidial pustules form after 7 days, no distinct odor or diffusing pigment observed. | 62–65 | Pyramidal, with opposite branches slightly inclined upward or at right angles, sometimes arising at irregular intervals along the main axis, closely spaced and not re-branching. | 6.0–8.1 × 3.0–3.8 (n = 30) | lageniform to subulate | 2.6–3.4 × 2.4–2.8 (n = 50) | green, globose to obovoidal | 5.1–7.0 × 4.9–6.8 (n = 30) | often terminal, globose to subglobose |
| Inoculated strains | Inoculation condition | Incidence rate (%) | Disease index | Relative control effect (%) | |||
|---|---|---|---|---|---|---|---|
| Before pathogens | 10.05 ± 0.63 H | 10.34 ± 1.46 g | 14.67 ± 2.07 FG | 12.67 ± 4.68 g | 83.38 ± 2.69 AB | 85.32 ± 6.60 a | |
| Simultaneously | 15.57 ± 0.59 DE | 15.94 ± 1.35 ef | 21.33 ± 4.13 E | 22.00 ± 4.20 ef | 76.63 ± 3.13 CD | 75.11 ± 5.26 bc | |
| After pathogens | 32.15 ± 1.74 C | 34.10 ± 2.62 bc | 34.67 ± 4.84 BC | 36.67 ± 3.01 bc | 60.64 ± 6.40 FG | 58.50 ± 6.07 ef | |
| Before pathogens | 14.03 ± 0.84 EF | 10.54 ± 1.89 g | 19.33 ± 3.01 EF | 15.33 ± 3.01 fg | 78.15 ± 3.24 BCD | 82.54 ± 4.32 ab | |
| Simultaneously | 13.22 ± 0.71 FG | 12.42 ± 1.28 fg | 28.67 ± 3.01 D | 20.67 ± 5.89 f | 67.37 ± 5.20 E | 76.37 ± 8.12 b | |
| After pathogens | 30.14 ± 0.90 C | 30.24 ± 0.92 cd | 37.33 ± 4.13 B | 34.67 ± 3.27 cd | 57.56 ± 6.52 G | 60.74 ± 6.17 de | |
| Before pathogens | 11.45 ± 1.02 GH | 10.31 ± 0.94 g | 15.33 ± 3.01 FG | 16.67 ± 3.93 fg | 82.53 ± 4.03 ABC | 81.33 ± 4.27 ab | |
| Simultaneously | 16.57 ± 0.79 D | 18.79 ± 0.57 e | 22.67 ± 5.47 E | 28.67 ± 3.01 de | 74.50 ± 5.22 D | 67.67 ± 4.53 cd | |
| After pathogens | 39.54 ± 0.74 B | 36.24 ± 3.77 b | 38.67 ± 4.84 B | 42.67 ± 4.13 b | 56.42 ± 3.91 G | 52.13 ± 3.10 f | |
| Before pathogens | 9.49 ± 1.33 H | 10.49 ± 0.35 g | 10.67 ± 3.26 G | 11.33 ± 3.93 g | 86.64 ± 4.54 A | 86.84 ± 5.80 a | |
| Simultaneously | 11.74 ± 1.77 FGH | 12.24 ± 1.17 fg | 18.67 ± 3.27 EF | 17.33 ± 4.84 fg | 78.96 ± 3.28 BCD | 80.39 ± 5.81 ab | |
| After pathogens | 31.58 ± 1.37 C | 29.52 ± 0.51 d | 30.67 ± 3.27 CD | 32.00 ± 3.58 cd | 65.33 ± 3.71 EF | 63.56 ± 7.38 de | |
| Alone | 50.65 ± 1.68 A | 48.29 ± 4.82 a | 88.67 ± 6.89 A | 89.33 ± 9.35 a | — | — | |
| — | Sterile water | 0 I | 0 h | 0 H | 0 h | — | — |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal Plant Pathogen Control · Plant-Microbe Interactions and Immunity · Plant Pathogens and Fungal Diseases
Introduction
Ancient trees are valuable natural resources and historical relics that sustain biodiversity and ecosystem functions, as well as provide social and cultural benefits to people (Huang et al. 2023). Platycladus orientalis (Cupressaceae), native to China and North Korea, is a robust, long-lived, evergreen coniferous tree widely distributed in China (Li et al. 2016; Dong et al. 2023). The ancient P. orientalis, symbolizing auspiciousness and longevity, is commonly found in prominent historical and cultural sites in Beijing, including palaces, temples, and parks. Beijing hosts the largest number of ancient trees in China, with 22,570 ancient P. orientalis, accounting for 55.69% of the city’s total ancient tree population (Zhao 2022; Zhang 2024).
Ancient P. orientalis, remaining in an over-mature stage for extended periods, experience declining physiological function and stress resistance, as well as urban human disturbances, which significantly increase the risk of diseases (Lindenmayer and Laurance 2017; Jiao et al. 2024). Leaf blight caused by Alternaria spp. is the most common disease of P. orientalis. As early as 1922, P. orientalis leaf blight caused by A. pruni was identified in northern China (Zhu 1922). Leaf blight of ancient P. orientalis caused by A. tenuis was reported in Beijing in 1996, resulting in extensive leaf and branch death (Lei et al. 1996). Subsequently, A. alternata was identified as the causal agent of leaf blight of ancient P. orientalis at the Mausoleum of the Yellow Emperor in Shaanxi (Li et al. 2021). In addition, A. alternata and A. infectoria were reported to cause leaf blight and dieback of P. orientalis around Tbilisi (Danelia et al. 2021). We have recently confirmed that leaf blight affecting ancient P. orientalis in Beijing is caused by Alternaria species, including A. alternata, A. subcucurbitae, A. cantlous, and A. platycladi (Jiao et al. 2025). Of these, A. alternata and A. cantlous were the most dominant causal agents (Jiao et al. 2025).
Trichoderma (Hypocreaceae, Hypocreales), originally introduced by Persoon in 1794, is an ecologically and economically important genus (Persoon 1794; Ye et al. 2023). Species of Trichoderma are widely distributed in various ecosystems, including natural soils, decaying wood, plant leaves, bark, and root systems, and also live as endophytes in plant tissues (Samuels 2006; Zheng et al. 2021). To date, more than 500 species have been described and recognized (Zhao et al. 2025). Many Trichoderma species are extensively used in agriculture, industry, and medicine, serving as bio-fungicides to control plant diseases, regulators of plant growth, fortifiers of soil fertility, and producers of antibiotics and enzymes (Saravanakumar and Kathiresan 2014; Bischof et al. 2016; Ye et al. 2023). Additionally, some members show great potential for applications in soil and water pollution remediation, as well as in the production of gold or silver nanoparticles (Harman et al. 2004; Anand et al. 2006; Mazyar et al. 2010). Endophytic Trichoderma species exhibit enhanced adaptation and colonization in their native hosts, thereby providing more effective disease control as biocontrol agents (Joseph 2025). For example, the endophytic T. koningiopsis T2, isolated from Liriodendron chinense × tulipifera, can effectively suppress black spot disease caused by Alternaria alternata and Colletotrichum gloeosporioides (Kong et al. 2022). In addition, a wettable powder formulation developed from the endophytic T. zelobreve T20 effectively controlled apple canker caused by Cytospora cincta and Neoscytalidium dimidiatum (Ranjbar et al. 2024). However, screening of endophytic Trichoderma species from P. orientalis for controlling leaf blight has not yet been studied.
In this study, several Trichoderma isolates were obtained from ancient P. orientalis in Beijing. The aim of this study was to (1) identify the Trichoderma isolates based on morphological characteristics and multi-locus phylogenetic analysis using ITS, tef1-α, and rpb2 loci, (2) determine the resistance of these representative Trichoderma strains against Alternaria pathogens through plate antagonism tests, and (3) evaluate the control effect of these Trichoderma strains against Alternaria leaf blight of P. orientalis through in situ inoculation in a greenhouse.
Materials and methods
Specimen collection and fungal isolation
Healthy branch and trunk tissues of ancient P. orientalis were collected from three locations (Zhongshan Park, Fahai Temple, and Working People’s Cultural Palace) in Beijing, China, from September 2023 to December 2024. The tissue was cut into 0.5 × 0.5 × 0.2 cm sections, then surface-sterilized with 75% ethanol for 60 s and 1% NaOCl for 60 s, followed by three rinses with sterile distilled water, and cultured on potato dextrose agar (PDA) medium at 25 °C until fungal colonies were observed (Zhou et al. 2019). Pure cultures were obtained from hyphal tips at the margins of single colonies, which were subcultured on fresh PDA and maintained at 25 °C. Based on colony characteristics, several Trichoderma isolates were obtained. The specimens were deposited at Beijing Forestry University (BJFU), whereas the fungal isolates were submitted to the China General Microbiological Culture Collection Center (CGMCC).
Morphological characterization
The purified strains were cultured on PDA (10 g potato extract, 20 g dextrose, 13 g agar, 1 L distilled water), malt extract agar (MEA; 20 g malt extract, 15 g agar, 1 L distilled water), cornmeal dextrose agar (CMD; 40 g cornmeal, 20 g dextrose, 15 g agar, 1 L distilled water), and synthetic low-nutrient agar (SNA; 1 g KH_2_PO_4_, 1 g KNO_3_, 0.5 g MgSO_4_, 0.5 g KCl, 0.2 g glucose, 0.2 g sucrose, 15 g agar, 1 L distilled water) for 7 days in an incubator at 25 °C with alternating 12 h/12 h fluorescent light/darkness (Zhao et al. 2023). Colony diameters were measured after 3 days, and trials were replicated three times. The morphological characteristics of colonies, including colony appearance, color, radial and concentric rings formed by spore production, pigmentation, and odor, were recorded (Zheng et al. 2021). The characteristics of conidiophores, conidia, and phialides were photographed, and at least 30 measurements per structure were documented and examined under a microscope (Nikon Eclipse E600).
DNA extraction, PCR amplification, and sequencing
Mycelia (approximately 0.1 g) were subjected to DNA extraction using a CTAB plant genomic DNA fast extraction kit (Aidlab Biotechnologies Co., Ltd., Beijing, China). The internal transcribed spacer (ITS) region was amplified with primers ITS5 and ITS4 (White et al. 1990), tef1-α with primers EF1-728F (Carbone and Kohn 1999) and EF1-LLErev (Jaklitsch et al. 2005), and rpb2 with primers fRPB2-5f and fRPB2-7cr (Liu et al. 1999). The polymerase chain reaction (PCR) amplification system and reaction conditions were described by Zhao et al. (2025). PCR amplicons were purified and sequenced at Sangon Biotechnology Co., Ltd. (Beijing, China).
Phylogenetic analyses
The newly generated sequences in this study were submitted to GenBank to obtain accession numbers, and ITS, tef1-α, and rpb2 sequences of closely related Trichoderma species were retrieved from GenBank for phylogenetic analysis (Suppl. material 1). Multiple sequence alignment was performed using MAFFT v.7.110 (http://mafft.cbrc.jp/alignment/server/). Ambiguous sequences at the start and end were deleted and manually adjusted using BioEdit.
A combined dataset of ITS, tef1-α, and rpb2 sequences from Trichoderma species was used to perform phylogenetic analyses based on maximum likelihood (ML), Bayesian inference (BI), and maximum parsimony (MP). ML analyses were conducted using RAxML-HPC BlackBox v.8.2.10 under the GTR+GAMMA model with 1,000 bootstrap replicates (Stamatakis 2014). BI analyses were performed in MrBayes v.3.2.6 with four Markov Chain Monte Carlo (MCMC) chains run for 5,000,000 generations, sampling every 1,000 generations and discarding the initial 25% of sampled data as burn-in (Ronquist et al. 2012). MP analyses were performed in PAUP* v.4.0b10 using heuristic searches with tree bisection–reconnection (TBR) branch swapping and 1,000 random addition replicates (Swofford 2002). Phylogenetic trees were visualized using FigTree v.1.4.4 (http://tree.bio.ed.ac.uk/software/figtree).
Plate antagonism tests
According to our recent report (Jiao et al. 2025), representative pathogenic strains of leaf blight on ancient P. orientalis in Beijing, Alternaria alternata CGMCC3.28778 and A. cantlous CGMCC3.28780, were selected for further study. The tested biocontrol strains were representative strains of four Trichoderma species isolated in this study: T. citrinoviride CGMCC3.28773, T. nordicum CGMCC3.28772, T. obovatum CGMCC3.28771, and T. platycladi CGMCC3.28769. All tested strains were deposited at BJFU.
The dual-culture plate method was used for antagonism tests (Ojaghian 2011). Mycelial plugs with an 8 mm diameter of Trichoderma and Alternaria strains were cut from the edges of 5-day-old colonies. The plugs were placed at both ends of PDA plates (50 mm apart) and incubated at 25 °C. Plates inoculated only with the Alternaria strain served as the control. The colony diameter of the pathogen was measured daily, and the inhibition rate was calculated after 5 days. The inhibition rate was calculated as follows (Zhang et al. 2018): mycelial growth inhibition rate (%) = (control pathogen colony diameter − treatment pathogen colony diameter) / (control pathogen colony diameter − mycelial plug diameter) × 100%.
All experiments were carried out three times in parallel and repeated three times.
Evaluation of control efficacy
Leaves were inoculated via spray inoculation with a 10^8^ spores/mL Trichoderma sp. suspension and smear inoculation with a 10^6^ spores/mL Alternaria sp. suspension. Each Trichoderma strain treatment corresponded one-to-one with each Alternaria pathogen inoculation. The pot experiments were divided into four groups as follows (Kong et al. 2022): treatment 1 involved inoculating Trichoderma one day before the pathogen; treatment 2 involved simultaneous inoculation of Trichoderma and the pathogen; treatment 3 involved inoculating Trichoderma one day after the pathogen; and the control consisted of Alternaria pathogen inoculation only. One-year-old P. orientalis seedlings (aboveground height, approximately 25 cm) were selected for inoculation experiments. Six leaf clusters per pot were inoculated, and the selected leaf clusters in each pot had similar areas and numbers of scale leaves (approximately 30 scale leaves per pot). Leaf surfaces were slightly wounded before inoculation. One day after inoculation with Alternaria spore suspensions, slight water-soaked symptoms were observed on the leaves. Infection of the leaves was confirmed by fulfilling Koch’s postulates. The inoculated P. orientalis seedlings were placed in a greenhouse at 14 °C–28 °C, 50%–75% relative humidity, under a 12 h light/dark cycle, and disease development was monitored daily. Each treatment had three repetitions with three pots each.
In the greenhouse experiment to investigate the biological control effect of Trichoderma spp. on P. orientalis leaf blight, incidence and disease index were quantified after 7 days of treatment. Disease was quantified according to diseased leaf area (Table 1) as follows (Kong et al. 2022): incidence rate (%) = number of diseased leaves / total number of leaves × 100%; disease index = (Σ representative value at all levels × leaf number at all levels) / (highest representative value × total leaf number) × 100; relative control effect (%) = (control disease index − disease index of each treatment) / control disease index × 100%.
Table 1.: Grading standard for leaf blight of Platycladus orientalis.
Statistical analysis
Data were analyzed using an analysis of variance and Duncan’s multiple comparison tests with IBM SPSS Statistics 19 (IBM Inc., Armonk, NY). Standard errors of all mean values were calculated (P < 0.05).
Results
Phylogenetic analysis
In the tef1-α phylogenetic tree, strains CGMCC3.28769 and CGMCC3.28770 formed a distinct clade within the Harzianum clade and appeared closely related to T. inhamatum (Suppl. material 2). The strain CGMCC3.28773 clustered with T. citrinoviride in the Longibrachiatum clade. Strains CGMCC3.28771 and CGMCC3.28772 clustered with T. obovatum and T. nordicum, respectively, in the Atroviride clade. In the rpb2 phylogenetic tree, strains CGMCC3.28769 and CGMCC3.28770 formed a distinct clade within the Harzianum clade, clustering close to T. afarasin, T. azevedoi, T. camerunense, T. lentiforme, and T. shaanxiensis (Suppl. material 3). The positions of strains CGMCC3.28773 and CGMCC3.28771 were consistent with those in the tef1-α phylogenetic tree, whereas CGMCC3.28772 clustered with T. nigricans and T. nordicum in the Atroviride clade.
The phylogenetic tree based on the concatenated ITS, tef1-α, and rpb2 datasets, analyzed using three methods (ML, BI, and MP), revealed the classification of one new species in the Harzianum clade, one known species in the Longibrachiatum clade, and two known species in the Atroviride clade (Fig. 1). In the phylogram, T. platycladi formed a distinct clade within the Harzianum clade with strong support (ML/BI/MP = 100/1/100). The strain CGMCC3.28773 within the Longibrachiatum clade clustered with T. citrinoviride with high support (ML/BI/MP = 100/1/100). In the Atroviride clade, strain CGMCC3.28772 formed a well-supported clade with T. nordicum (ML/BI/MP = 86/0.99/86), and strain CGMCC3.28771 clustered with T. obovatum with high support (ML/BI/MP = 94/1/95).
Phylogenetic tree generated by maximum likelihood analysis using the concatenated sequences of the ITS, tef1-α, and rpb2 loci of the genus Trichoderma. RAxML bootstrap support values (ML ≥ 50%), Bayesian posterior probability (PP ≥ 0.90), and MP bootstrap support values (ML ≥ 50%) are shown at the nodes. Strains in this study are highlighted in bold.
Taxonomy
Trichoderma
citrinoviride
Taxon classificationFungiHypocrealesHypocreaceae
Bissett, Canad. J. Bot. 62(5): 926 (1984)
12084BC6-BCDA-5606-AB8C-CEE862C92CBD
Strains examined.
China • Beijing, Shijingshan District, Fahai Temple (39°56'29"N, 116°9'33"E), on the branch of ancient P. orientalis, December 2024, N. Jiao and Z. Zhang (BJFU-FHS354; living culture: CGMCC3.28773).
Notes.
Phylogenetic analysis showed that strain CGMCC3.28773 clustered together with T. citrinoviride in the Longibrachiatum clade, with strong support (ML/BI/MP = 100/1/100). Morphologically, the characteristics and sizes of phialides (3.6–6.8 × 2.4–3.0 μm, n = 30 in this study vs. 3.5–6.6 × 2.0–3.2 μm), conidia (2.9–3.7 × 1.8–2.4 μm, n = 50 in this study vs. 2.2–3.7 × 1.5–2.1 μm), and chlamydospores (5.8–7.0 × 3.5–5.0 μm, n = 30 in this study vs. 4–7 × 3–5 μm) were similar to the description provided by Bissett (1984) (Table 2). Based on phylogeny and morphology, this isolate was identified as T. citrinoviride. Trichoderma citrinoviride has previously been reported as an endophyte of P. orientalis, and in this study, this species was isolated again.
Colonies and microscopic characteristics of Trichoderma species isolated from the ancient P. orientalis. Part I: the morphology of the three known species. A. Colonies after 7 d at 25 °C on PDA; B. Conidiophores and phialides; C. Conidia; D. Chlamydospore. Scale bars: 10 μm (B–D). Part II: the morphology of the novel species T. platycladi. A–D. Colonies after 7 d at 25 °C on media (A: PDA; B: MEA; C: CMD; D: SNA); E–G. Conidiophores and phialides; H–J. Conidia; K, L. Chlamydospore. Scale bars: 10 μm (E–L).
Trichoderma
nordicum
Taxon classificationFungiHypocrealesHypocreaceae
G.Z. Zhang, MycoKeys 87: 144 (2022)
5891F2F5-C1B8-5343-A50A-1F0C7FB3C841
Strains examined.
China • Beijing, Shijingshan District, Fahai Temple (39°56'29"N, 116°9'33"E), on the trunk of ancient P. orientalis, November 2024, N. Jiao and Z. Zhang (BJFU-FHS298; living culture: CGMCC3.28772).
Notes.
Trichoderma nordicum was originally isolated from soil in Hebei, China. Phylogenetic analysis revealed that strain CGMCC3.28772 clustered with the type strain T. nordicum WT13001 in the Atroviride clade. Morphologically, the type strain is characterized by lageniform phialides (7.2–10.3 × 2.9–3.2 μm) and globose to obovoidal conidia (4.4–4.8 × 4.1–4.4 μm), which are similar to those of our strain (Zhang et al. 2022) (Table 2). Based on both phylogenetic and morphological evidence, this strain was identified as T. nordicum. This study reports the first isolation of T. nordicum from P. orientalis.
Trichoderma
obovatum
Taxon classificationFungiHypocrealesHypocreaceae
Z.F. Yu & Y.F. Lv, J. Fungi 7(6):467 (2021)
E3388C88-B9FC-5B7B-A469-069D6D1F3608
Strains examined.
China • Beijing, Dongcheng District, Working People’s Cultural Palace (39°54'45"N, 116°23'54"E), on the trunk of ancient P. orientalis, November 2023, N. Jiao and Z. Zhang (BJFU-TM298; living culture: CGMCC3.28771).
Notes.
Trichoderma obovatum was originally isolated from soil in Yunnan, China. Phylogenetically, strain CGMCC3.28771 clustered with T. obovatum in the Atroviride clade with strong support values of 94/1/95 (ML/BI/MP). Morphologically, our isolate was consistent with the type strain, with phialides (5.0–9.0 × 2.8–4.0 μm, n = 30 in this study vs. 4.8–8.9 × 2.5–3.9 μm) and conidia (3.2–4.0 × 3.0–3.5 μm, n = 50 in this study vs. 3.2–3.8 × 3.0–3.6 μm) showing similar characteristics and sizes (Zheng et al. 2021) (Table 2). Therefore, this strain was identified as T. obovatum. To our knowledge, this is the first report of this species isolated from P. orientalis.
Trichoderma
platycladi
Taxon classificationFungiHypocrealesHypocreaceae
N. Jiao & Y. Zhang sp. nov.
73F77AF7-0B96-590E-8E5D-07D9A97992F3
MycoBank No: 859234
Etymology.
The epithet refers to the host plant genus Platycladus.
Description.
Sexual morph: Unknown. Asexual morph: Conidiophores pyramidal, with opposite branches slightly inclined upward or at right angles, sometimes arising at irregular intervals along the main axis, closely spaced, and not re-branching. Phialides lageniform to subulate, often with a narrow neck, discrete or integrated, solitary or in whorls of 2–4, 5.8–8.1(−8.5) × 2.8–4.0 μm (mean ± SD = 7.5 ± 0.7 × 3.2 ± 0.2 μm, n = 30), length/width ratio of 1.8–2.9 (mean 2.3 ± 0.3), base 1.4–1.8 μm (mean ± SD = 1.5 ± 0.2 μm, n = 30). Conidia globose to obovoidal, green, 2.4–3.5 × 2.2–2.9 μm (mean ± SD = 3.1 ± 0.2 × 2.7 ± 0.1 μm, n = 50), length/width ratio of 1.0–1.3 (mean ± SD = 1.1 ± 0.1). Chlamydospores often terminal, globose to subglobose, 5.0–7.1 × 4.8–7.0 μm (mean ± SD = 6.0 ± 0.5 × 5.5 ± 0.6 μm, n = 30), length/width ratio of 1.0–1.2 (mean ± SD = 1.1 ± 0.1).
Culture characteristics.
Optimal growth at 30 °C. Colony radius on PDA after 3 days: 62–65 mm at 25 °C, covering the plate at 30 °C, and 30–32 mm at 35 °C. Covering the plate after 4 days at 25 °C. Colony radial, white, fluffy, aerial hyphaedense, abundant. Yellow-green and white conidial pustules form after 7 days. No distinct odor or diffusing pigment observed. Colony radius on MEA after 3 days: 56–58 mm at 25 °C, 68–70 mm at 30 °C, and 28–31 mm at 35 °C. Covering the plate after 4 days at 25 °C and at 30 °C. Colony well-defined, radial, mycelia white, forming a broad distinct zonate after 7 days. No distinct odor or diffusing pigment observed. Colony radius on CMD after 3 days: 40–45 mm at 25 °C, 62–65 mm at 30 °C, and 36–38 mm at 35 °C. Covering the plate after 5 days at 25 °C and 4 days at 30 °C. Colony white, radial, aerial hyphae rare. No distinct odor or diffusing pigment observed. Colony radius on SNA after 3 days: 40–43 mm at 25 °C, 62–65 mm at 30 °C, and 25–27 mm at 35 °C. Covering the plate after 5 days at 25 °C and 4 days at 30 °C. Colony hyaline, radial, aerial hyphae sparse. Pale green and white conidial pustules form after 3 days, a small green disk around the inoculums. No distinct odor or diffusing pigment observed.
Type.
China • Beijing, Dongcheng District, Zhongshan Park (39°54'37"N, 116°23'39"E), on the trunk of ancient P. orientalis, September 2023, N. Jiao and Y. Zhang (holotype BJFU-ZS17; paratype BJFU-ZS15, ex-type culture: CGMCC3.28769).
Additional strain examined.
• Living culture: CGMCC3.28770; other information is the same as the type.
Notes.
Trichoderma platycladi is recognized as an endophyte of P. orientalis. Phylogenetic analysis indicated that this species is a new member of the Harzianum clade and is closely related to T. inhamatum. T. platycladi exhibits nucleotide differences from T. inhamatum in the rpb2 region amounting to 3.2% (25/788, one gap) and differences of 1.1% (6/563, two gaps) in tef1-α. Morphologically, the phialides of T. platycladi are obviously longer than those of the type strain of T. inhamatum (mean 7.5 × 3.2 μm vs. 5.0 × 3.2 μm) (Veerkamp and Gams 1983; Chaverri et al. 2015). In addition, the conidia of T. platycladi are slightly larger than those of T. inhamatum (2.6–3.4 × 2.4–2.8 μm vs. 2.5–3.0 × 2.2–2.7 μm) (Veerkamp and Gams 1983; Chaverri et al. 2015). Based on morphological characteristics and multigene phylogenetic analyses, this species is described as a novel taxon.
Plate antagonism test
From the second day after inoculation, four Trichoderma strains showed significant inhibitory effects against A. alternata (P < 0.05). By the third day, these strains also showed significant inhibition against A. cantlous (P < 0.05). The diameter of the pathogens in the Trichoderma strain treatments tended to stabilize from the third day, whereas the diameter of the pathogens in the control continued to increase (Fig. 3). On the fifth day, the inhibition rates of T. platycladi CGMCC3.28769, T. citrinoviride CGMCC3.28773, T. obovatum CGMCC3.28771, and T. nordicum CGMCC3.28772 against A. alternata were 52.30%, 45.39%, 53.29%, and 55.92%, respectively, whereas the inhibition rates against A. cantlous were 53.95%, 50.33%, 55.26%, and 60.53%, respectively. Based on the 5-day antagonism tests, T. nordicum CGMCC3.28772 showed the strongest antagonistic effect against Alternaria pathogens, followed by T. platycladi CGMCC3.28769 and T. obovatum CGMCC3.28771, whereas T. citrinoviride CGMCC3.28773 showed the weakest inhibition.
Plate antagonism test of the four representative Trichoderma strains against Alternaria pathogens. A. Colony diameters of A. alternata CGMCC3.28778 from 1 d to 5 d; B. Colony diameters of A. cantlous CGMCC3.28780 from 1 d to 5 d. Different letters indicate significant differences between the control and inoculation treatments based on Duncan’s post hoc test (P < 0.05).
Evaluation of control efficacy
In situ inoculation on 1-year-old P. orientalis seedlings in the greenhouse demonstrated that the relative control effect against Alternaria leaf blight exceeded 75% when the Trichoderma strain was inoculated before the pathogen and still exceeded 50% when the Trichoderma strain was inoculated after the pathogen (Table 3). The disease index when the Trichoderma strain was inoculated before the pathogen was significantly lower than when the Trichoderma strain was inoculated after the pathogen (P < 0.05), whereas the relative control effect was significantly higher (P < 0.05). The evaluation results showed that application of T. nordicum CGMCC3.28772 resulted in the lowest disease index and the highest relative control effect under the same inoculation conditions compared with the other three Trichoderma strains, indicating that it is the most effective biocontrol strain.
Table 3.: The evaluation of the control efficacy of Trichoderma spp. against Alternaria leaf blight of Platycladus orientalis in the greenhouse. y
Discussion
Four endophytic Trichoderma species were identified from ancient P. orientalis in Beijing, namely T. citrinoviride, T. nordicum, T. obovatum, and T. platycladi. Trichoderma citrinoviride was previously reported as an endophyte from the leaves and branches of P. orientalis during a survey of conifers in Tbilisi and its surroundings (Danelia et al. 2021). Subsequently, endophytic T. harzianum was isolated from the branches of ancient P. orientalis in Shaanxi, China (Yu et al. 2021). In addition, Hosseyni-Moghaddam and Soltani (2014) reported that endophytic T. atroviride and T. koningii from Cupressaceae plants effectively inhibited common cypress fungal pathogens, including Diplodia seriata, Phaeobotryon cupressi, and Spencermartinsia viticola. Bu et al. (2016) isolated 311 endophytic fungal strains from the leaves of ancient P. orientalis in Shaanxi, which were classified into 21 genera, with Trichoderma as the dominant genus. Among these, Trichoderma sp. C-14 exhibited significant inhibitory effects against four plant pathogenic fungi: Cytospora chrysosperma, Fusarium oxysporum, Colletotrichum gloeosporioides, and Botryosphaeria dothidea. In the present study, T. citrinoviride was re-isolated as an endophyte from P. orientalis, and three other Trichoderma species were isolated, enriching the diversity of endophytic Trichoderma associated with P. orientalis.
The representative strains of the four Trichoderma species exhibited strong antagonistic activity against Alternaria pathogens in this study. Many members of Trichoderma can efficiently inhibit Alternaria pathogens. For example, T. harzianum significantly inhibited the growth of A. alternata, the causal agent of tobacco brown spot (Gveroska and Ziberoski 2012). Furthermore, the endophytic T. citrinoviride isolated from Panax ginseng exhibited strong antifungal activity against Alternaria panax (Park et al. 2019). Evaluation of the biocontrol effect demonstrated that the four Trichoderma strains were effective in controlling leaf blight of P. orientalis, with the highest effect observed when the Trichoderma strains were applied one day prior to inoculation with Alternaria pathogens. Comparable results have been reported for the control of tobacco black shank by T. harzianum CGMCC23294 and the control of leaf black spot in L. chinense × tulipifera by T. koningiopsis T2, in which the application of Trichoderma strains before pathogen inoculation achieved the best disease control (Ren et al. 2025; Kong et al. 2022). This may be due to the ability of Trichoderma spp. to colonize leaf surfaces rapidly and establish dominant populations in advance, thereby delaying disease occurrence (Guzmán-Guzmán et al. 2023). Moreover, among the tested strains, T. nordicum CGMCC3.28772 showed the highest effect, highlighting its potential for controlling leaf blight of P. orientalis.
Supplementary Material
XML Treatment for Trichoderma citrinoviride
XML Treatment for Trichoderma nordicum
XML Treatment for Trichoderma obovatum
XML Treatment for Trichoderma platycladi
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Anand P, Isar J, Saran S, Saxena RK (2006) Bioaccumulation of copper by Trichoderma viride. Bioresource Technology 97(8): 1018–1025. 10.1016/j.biortech.2005.04.04616324839 · doi ↗ · pubmed ↗
- 2Bischof RH, Ramoni J, Seiboth B (2016) Cellulases and beyond: The first 70 years of the enzyme producer Trichoderma reesei. Microbial Cell Factories 15(1): 106. 10.1186/s 12934-016-0507-6PMC 490290027287427 · doi ↗ · pubmed ↗
- 3Bissett J (1984) A revision of the genus Trichoderma. I. section Longibrachiatum sect. nov. Canadian Journal of Botany 62: 924–931. 10.1139/b 84-131 · doi ↗
- 4Bu FF, Yu ZD, Hou JJ, Sui YJ (2016) Isolation, identification and antifungal activity of endophytic fungi from ancient Platycladus orientalis. Forest Pest and Disease. 35(03): 12–16.
- 5Carbone I, Kohn LM (1999) A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 91(3): 553–556. 10.1080/00275514.1999.12061051 · doi ↗
- 6Chaverri P, Branco-Rocha F, Jaklitsch W, Gazis R, Degenkolb T, Samuels GJ (2015) Systematics of the Trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia 107(3): 558–590. 10.3852/14-147PMC 488566525661720 · doi ↗ · pubmed ↗
- 7Danelia I, Zaqariashvili N, Amiranashvili L, Badridze G, Kvitsiani S (2021) The mycological study of conifers in Tbilisi and its surroundings. Journal of Forest Science 67(10): 464–476. 10.17221/79/2021-JFS · doi ↗
- 8Dong Y, Xiao WF, Guo W, Liu YF, Nie W, Huang RZ, Tan CC, Jia ZR, Liu JF, Jiang ZP, Chang EM (2023) Effects of donor ages and propagation methods on seedling growth of Platycladus orientalis (L.) Franco in winter. International Journal of Molecular Sciences 24(8): 7170. 10.3390/ijms 24087170 PMC 1013832337108331 · doi ↗ · pubmed ↗