Phylogeny, divergence time and historical biogeography of Hyphoderma (Hyphodermataceae, Polyporales): Introducing five new species from China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
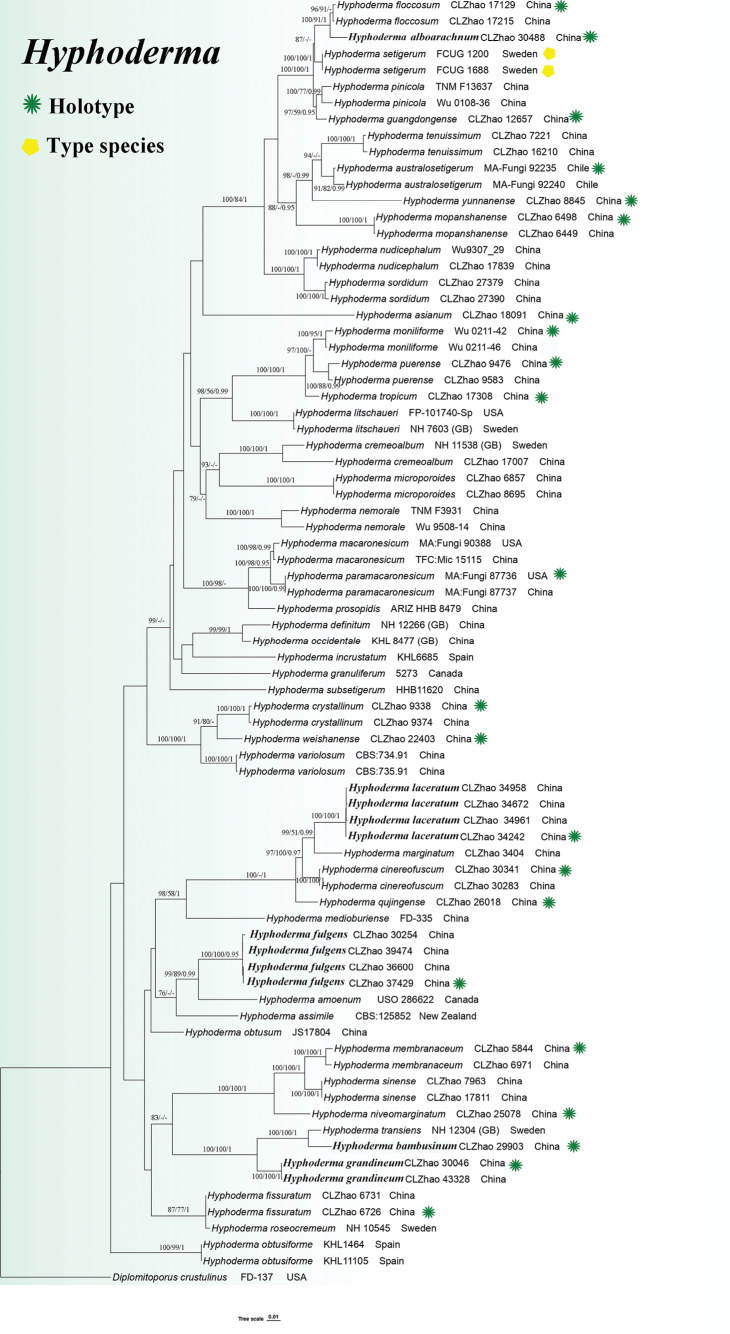 Figure 1
Figure 1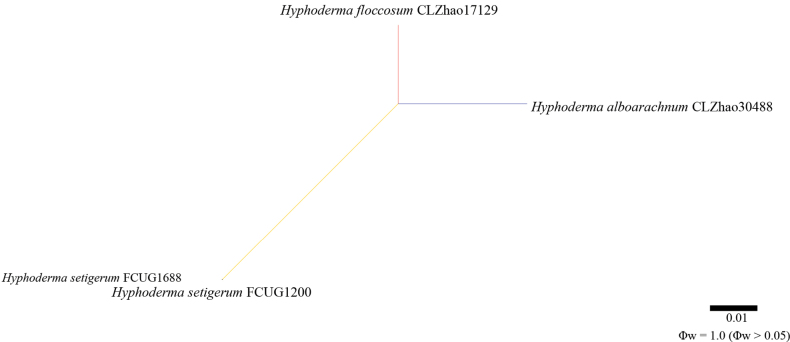 Figure 2
Figure 2 Figure 3
Figure 3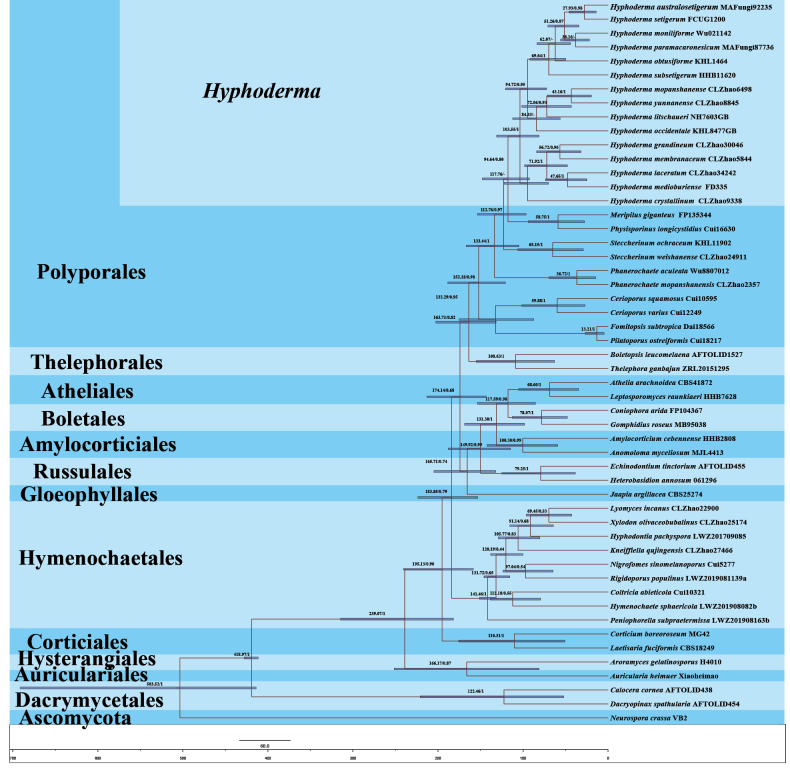 Figure 4
Figure 4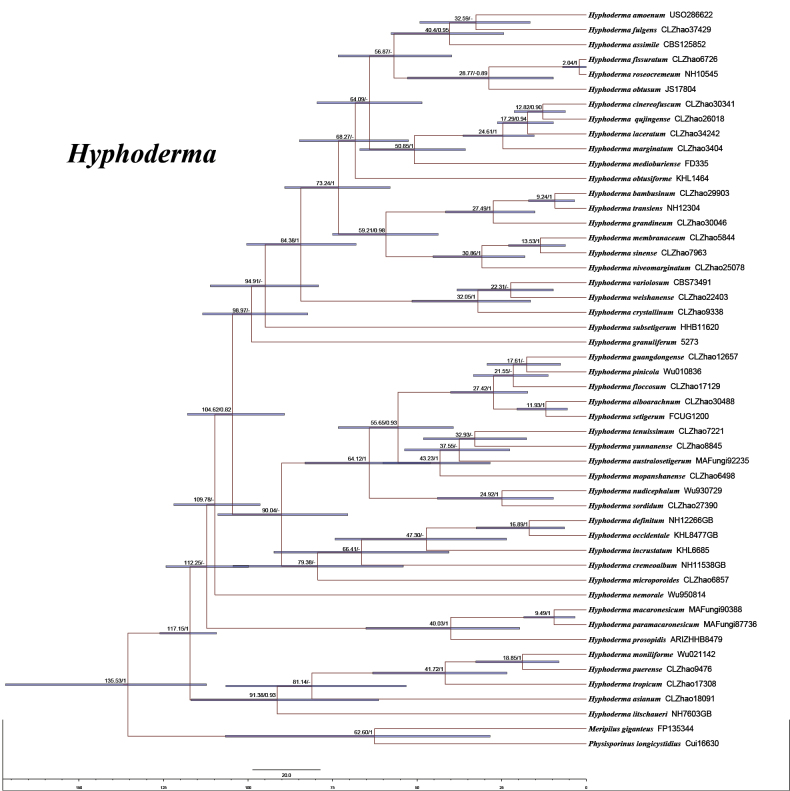 Figure 5
Figure 5 Figure 6
Figure 6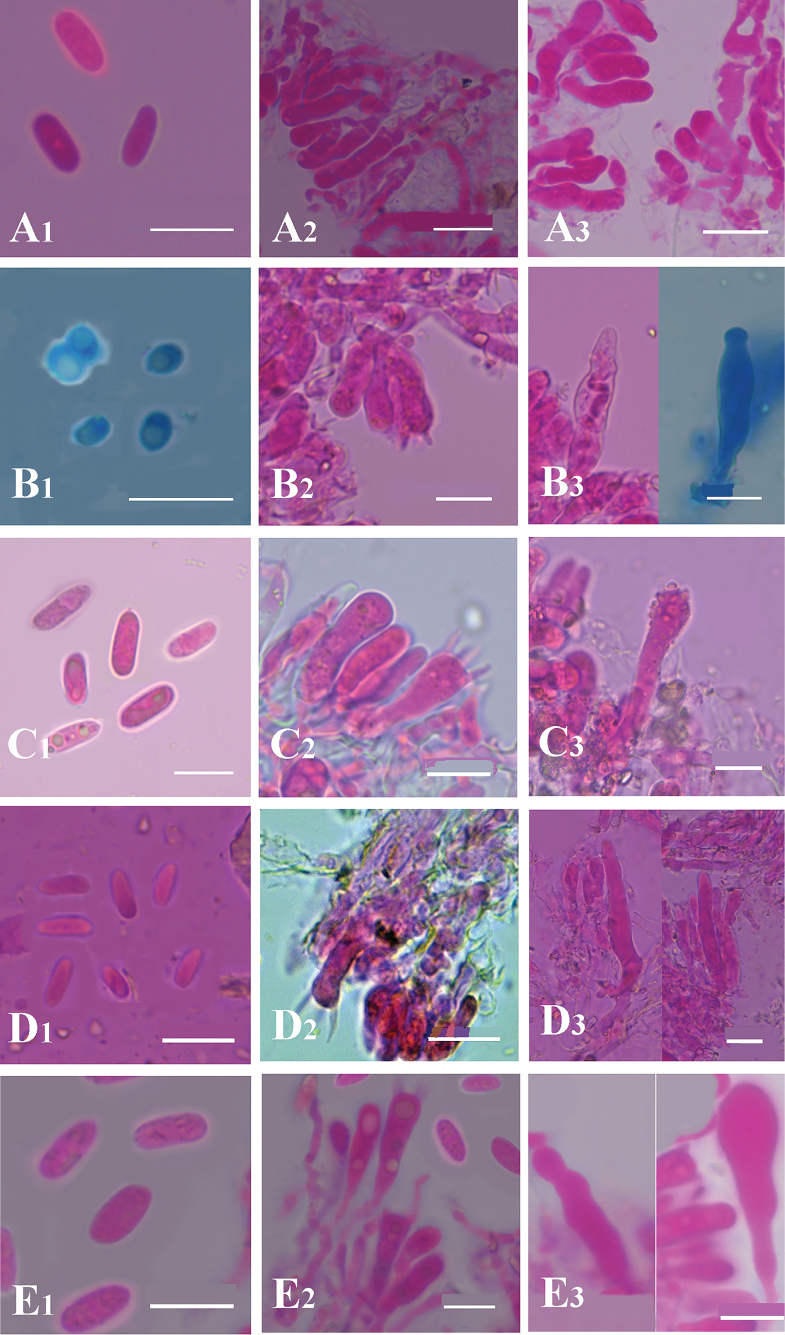 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10 Figure 11
Figure 11 Figure 12
Figure 12 Figure 13
Figure 13 Figure 14
Figure 14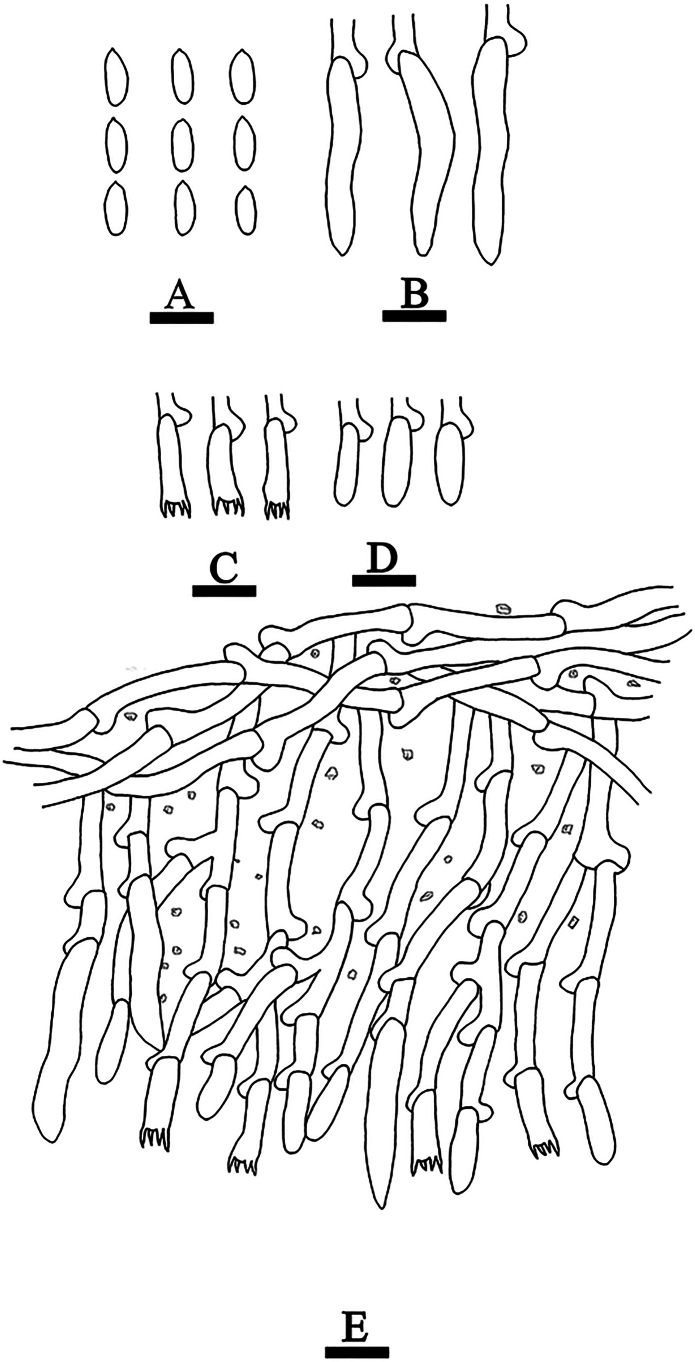 Figure 15
Figure 15 Figure 16
Figure 16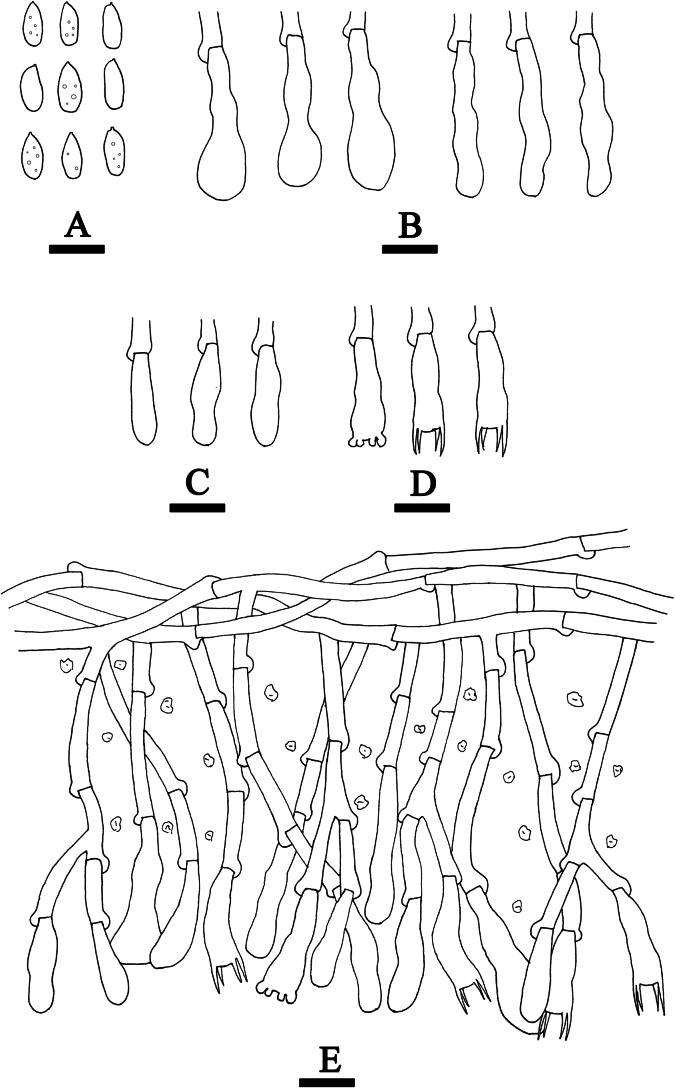 Figure 17
Figure 17| Species Name | Sample No. | GenBank Accession No. | References | Country | |
|---|---|---|---|---|---|
|
| nLSU | ||||
|
| HHB-2808 |
|
|
| USA |
|
| MJL-4413 |
|
|
| USA |
|
| H4010 | – |
|
| USA |
|
| CBS 418.72 |
|
|
| Netherlands |
|
| Xiaoheimao |
|
|
| China |
|
| AFTOL-ID 1527 |
|
|
| USA |
|
| AFTOL 438 |
|
|
| USA |
|
| Cui 10321 |
|
|
| China |
|
| Dai 13320 |
|
|
| China |
|
|
|
|
| USA | |
|
| MG 42 |
|
|
| Sweden |
|
| AFTOL 454 |
|
|
| USA |
|
| FD-137 |
|
|
| USA |
|
| AFTOL-ID 455 |
|
|
| USA |
|
| Cui 18217 |
|
|
| China |
|
| Dai 18566 |
|
|
| China |
|
| MB 95-038 |
|
|
| Germany |
|
| 06129/6 |
|
|
| China |
|
|
|
|
|
| |
|
| USO 286622 |
| — |
| Canada |
| CLZhao 18091 |
|
|
| China | |
|
| CBS:125852 |
|
|
| New Zealand |
| MA-Fungi 92235 |
|
| Chile | ||
|
| MA-Fungi 92240 |
|
| Chile | |
|
|
|
|
|
|
|
| CLZhao 30341 |
|
|
| China | |
|
| CLZhao 30283 |
|
|
| China |
|
| NH 11538 (GB) |
|
|
| Sweden |
|
| CLZhao 17007 |
|
|
| China |
| CLZhao 9338 |
|
|
| China | |
|
| CLZhao 9374 |
|
|
| China |
|
| NH 12266 (GB) |
|
|
| China |
|
| CLZhao 6731 |
| — |
| China |
| CLZhao 6726 |
|
|
| China | |
| CLZhao 17129 |
|
|
| China | |
|
| CLZhao 17215 |
|
|
| China |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| 5273 |
|
|
| Canada |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CLZhao 12657 |
|
|
| China | |
|
| KHL6685 | — |
|
| Spain |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
| NH 7603 (GB) |
|
|
| Sweden |
|
| FP-101740-Sp |
|
|
| USA |
|
| MA:Fungi 90388 |
| — |
| USA |
|
| TFC:Mic 15115 |
| — |
| China |
| CLZhao 3404 |
|
|
| China | |
|
| FD-335 |
|
|
| China |
| CLZhao 5844 |
|
|
| China | |
|
| CLZhao 6971 |
|
|
| China |
|
| CLZhao 6857 |
|
|
| China |
|
| CLZhao 8695 |
|
|
| China |
|
| Wu 0211-42 |
| — |
| China |
|
| Wu 0211-46 |
| — |
| China |
| CLZhao 6498 |
|
|
| China | |
|
| CLZhao 6449 |
|
|
| China |
|
| TNM F3931 |
|
|
| China |
|
| Wu 9508-14 |
|
|
| China |
| CLZhao 25078 |
|
|
| China | |
|
| Wu9307_29 |
| — |
| China |
|
| CLZhao 17839 |
|
|
| China |
|
| KHL1464 |
| — |
| Spain |
|
|
| — |
| Spain | |
|
| JS17804 | — |
|
| China |
|
| KHL 8477 (GB) |
|
|
| China |
| MA:Fungi 87736 |
| — |
| USA | |
|
| MA:Fungi 87737 |
| — |
| China |
|
| Wu 0108-32 |
|
|
| China |
|
| Wu 0108-36 |
|
|
| China |
|
| ARIZ HHB 8479 |
| — |
| China |
| CLZhao 9476 |
| — |
| China | |
|
| CLZhao 9583 |
|
|
| China |
| CLZhao 26018 |
|
|
| China | |
|
| NH 10545 |
|
| Sweden | |
|
| FCUG 1200 |
| — |
| Sweden |
|
| FCUG 1688 |
| — |
| Sweden |
| CLZhao 7963 |
|
|
| China | |
|
| CLZhao 17811 |
|
|
| China |
|
| CLZhao 27379 |
| — |
| China |
| CLZhao 27390 |
|
|
| China | |
|
| HHB11620 |
| — |
| China |
| CLZhao 7221 |
|
|
| China | |
|
| CLZhao 16210 |
|
|
| China |
|
| NH 12304 |
|
|
| Sweden |
| CLZhao 17308 |
|
|
| China | |
|
| CBS:734.91 |
|
|
| China |
|
| CBS:735.91 |
|
|
| China |
| CLZhao 22403 |
|
|
| China | |
|
| CLZhao 8845 |
|
|
| China |
|
| LWZ 20170908-5 |
|
|
| China |
|
| LWZ20190808-2b |
|
|
| China |
|
| CBS 252.74 |
|
|
| Netherlands |
|
| CBS:182.49 |
|
|
| Netherlands |
|
| HHB-7628 |
|
|
| USA |
|
| CLZhao 22900 |
|
|
| China |
|
| FP 135344 |
|
|
| USA |
|
| VB2 |
|
|
| India |
|
| Cui 5277 |
|
|
| China |
|
| Wu 880701-2 |
|
|
| China |
|
| CLZhao 2357 * |
|
|
| China |
|
| Cui 16630 |
|
|
| China |
|
| Cui 10595 |
|
|
| China |
|
| Cui 12249 |
|
|
| China |
|
| LWZ 20190816-3b |
|
|
| China |
|
| LWZ 20190811-39a |
|
|
| China |
|
| CLZhao 24911 * |
|
|
| China |
|
| KHL 11902 |
|
|
| Sweden |
|
| ZRL20151295 |
|
|
| China |
|
| CLZhao 25174 |
|
|
| China |
| Species name | Basidiomata | Hyphal system | Cystidia | Basidiospores | References |
|---|---|---|---|---|---|
|
| Membranaceous, hymenial surface arachnoid. | Monomitic; generative hyphae with clamp connections, colorless, thin-walled. | Absent | Oblong-ellipsoid to cylindrical; 5–6 × 2–3 µm. | Present study |
|
| Ceraceous to membranaceous, hymenial surface smooth, floccose. | Monomitic; generative hyphae with clamp connections, colorless, thin-walled. | Moniliform; 20–50 × 4–6 μm. | Ellipsoid; 6.5–10 × 4–6 μm. |
|
|
| Membranaceous, hymenial surface smooth. | Monomitic; generative hyphae with clamp connections, colorless, thin–to thick. | Tubular; 21.5–26.5 × 5–6.5 µm. | Ellipsoid; 4.5–6.5 × 3–4.5 µm. | Present study |
|
| Coriaceous, hymenial surface smooth. | Monomitic; generative hyphae with simple-septa, colorless, thin-walled. | 1) Tubular; 25.9–37.1 × 1.8–2.5 μm 2) Capitate; 12.5–15.5 × 6.2–7 μm. | Cylindrical; 9–11.5 × 4.5–5 μm. |
|
|
| Membranaceous, scattered nubby crystals. | Monomitic; generative hyphae with clamp connections, colorless, thin-walled. | 1) Tubular 32–51 × 5–10 μm. 2) Encrusted 14–46 × 4–11 μm. | Allantoid; 11–14.5 × 4–5.5 μm. |
|
|
| Ceraceous, hymenial surface smooth. | monomitic, generative hyphae with clamp connections. | Absent | Cylindrical; 8.5–10.3 × 3–4 μm. |
|
|
| Ceraceous, hymenial surface farinaceous. | Monomitic; generative hyphae with simple-septa, colorless, thin-walled. | 1) septate 60–161 × 5.5–10 μm. 2) tubular 37.5–100 × 4–8.5 μm. | Ellipsoid to allantoid; 6–9.5 × 3–4.5 μm. |
|
|
| Membranaceous, hymenial surface smooth. | Monomitic; generative hyphae with simple-septa, colorless, thin-walled. | Clavate 37.1 × 5.5–7.5 µm. | Subcylindrical; 7.5–11.5 × 2.5–4 µm. | Present study |
|
| Membranaceous, hymenial surface grandinioid. | Monomitic; generative hyphae with simple-septa, colorless, thin-walled. | Leptocystidia 29.5–33.5 × 4–5 µm. | Cylindrical;7–9 × 2.5–4 µm. | Present study |
|
| Membranaceous, hymenial surface smooth. | Monomitic; generative hyphae with simple-septa, colorless, thin-walled. | 1) Clavate 27–29 × 6.5–8 µm. 2) Capitate 26–30 × 7.5–8.5 µm. | Subellipsoid; 6.5–9 × 3–4.5 µm. | Present study |
|
| Membranaceous, hymenial surface smooth. | Monomitic; generative hyphae with simple-septa, colorless, thin-walled. | Cylindrical 30–48.5 × 7.5–11.5 µm. | Cylindrical; 9–10 × 3.5–4.5 µm. |
|
|
| Membranaceous, hymenial surface tuberculate | Monomitic, generative hyphae with clamp thin-walled. | Moniliform; 28–60 × 6.5–10.5 μm. | Ellipsoid to cylindrical 11–13.5 × 4.5–5.5 μm. |
|
|
| Cottony hymenial surface smooth. | Monomitic, generative hyphae with clamp connections, colorless, thin-walled | Capitate; 18–51 × 4.5–7 μm. | Cylindrical to allantoid, 8.5–10 × 2.5–3.5 μm. |
|
|
| Ceraceous, hymenial surface porulose to pilose. | monomitic, generative hyphae bearing clamp connections | Fusiform; 86–171 × 10.5–13 μm | Cylindrical 7.8–9.7 × 2.6–3.3 μm. |
|
|
| Ceraceous, hymenial surface smooth. | Monomitic, generative hyphae with clamp connections, colorless, thin-walled | Moniliform; 29–55.5 × 5–7 µm. | Ellipsoid; 7–9 × 3.5–5 μm. |
|
|
| Membranaceous, hymenial surface chalky. | Monomitic, generative hyphae with clamp connections, colorless, thin-walled | (1) Scattered; 65–180 × 7–11 μm. (2) Aseptate; 35–45 × 6–7 μm. | Cylindrical to allantoid; 13–16 × 4–4.5 μm. |
|
|
| Byssoid, hymenial surface smooth, floccose. | Monomitic; generative hyphae with clamp connections, colorless, thick-walled. | Tubular; 25–97 × 5.5–9.5 µm. | Ellipsoid; 6–7.5 × 3–4.5 μm. |
|
|
| Membranaceous, hymenial surface smooth, floccose. | Monomitic; generative hyphae with clamp connections, colorless, thin-walled. | Capitate; 20–40 × 9–13 μm. | Ellipsoid to cylindrical; 7–11.5 × 3.5–5 μm. |
|
|
| Membranaceous, hymenial surface smooth. | Monomitic; generative hyphae with clamp connections, colorless, thick-walled. | 1) Encrusted; 18.5–38 × 6–11 μm. 2) Moniliform; 30–60.5 × 3–5 μm. | Cylindrical to slightly allantoid; 8–11.5 × 2.6–3.3 μm |
|
|
| Membranaceous, hymenial surface smooth. | Monomitic; generative hyphae with clamp connections, colorless, thin-walled. | Tubular; 42–72.5 × 6–11 μm. | Ellipsoid; 3–4.5 × 2–3 μm. |
|
|
| Membranaceous, hymenial surface tuberculate to minutely-grandinioid | Monomitic; generative hyphae with clamp connections, colorless, thick-walled. | Cylindrical; 50–220 × 6.5–13 µm. | Cylindrical; 7–10.5 × 3–4.5 µm. |
|
|
| Coriaceous hymenial surface tuberculate. | Monomitic; generative hyphae with clamp connections, colorless, thin-walled. | Moniliform; 60–102.5 × 5.5–7.5 μm. | Ellipsoid to cylindrical; 6.5–7.5 × 3–4 μm. |
|
|
| Membranaceous, hymenial surface smooth. | Monomitic; generative hyphae with clamp connections, colorless, thin-walled. | Absent | Broadly ellipsoid; 4.5–8.5 × 4–7 μm. |
|
|
| Corneus, hymenial surface tuberculate. | Monomitic; generative hyphae with clamp connections, colorless, thin-walled. | Cylindrical; 63–124 × 7–10 µm | Ellipsoid to cylindrical; 10–11.5 × 4–5.5 μm. |
|
| Taxa | Means of stem age (Ma) | 95% |
|---|---|---|
|
| 11.93 | 5.57–20.46 |
|
| 32.59 | 16.57–49.20 |
|
| 81.14 | 53.20–106.54 |
|
| 40.40 | 24.36–57.71 |
|
| 37.55 | 22.63–53.73 |
|
| 9.24 | 3.42–17.02 |
|
| 12.82 | 6.16–21.28 |
|
| 66.41 | 40.63–92.33 |
|
| 32.05 | 16.41–51.53 |
|
| 16.89 | 6.40–32.47 |
|
| 2.04 | 0.02–6.92 |
|
| 21.55 | 11.21–33.35 |
|
| 32.59 | 16.57–49.20 |
|
| 98.97 | 82.33–113.44 |
|
| 27.49 | 15.22–41.59 |
|
| 17.61 | 7.59–29.35 |
|
| 47.30 | 23.50–74.28 |
|
| 17.29 | 9.71–26.31 |
|
| 91.38 | 61.37–116.85 |
|
| 9.49 | 3.33–18.47 |
|
| 24.61 | 15.35–36.42 |
|
| 50.85 | 35.70–66.97 |
|
| 13.53 | 6.11–22.98 |
|
| 79.38 | 54.13–104.43 |
|
| 18.85 | 8.07–32.66 |
|
| 43.23 | 28.40–60.09 |
|
| 112.25 | 96.42–121.98 |
|
| 30.86 | 18.16–45.31 |
|
| 24.92 | 9.68–43.99 |
|
| 66.27 | 52.57–84.85 |
|
| 28.77 | 9.77–52.86 |
|
| 16.89 | 6.40–32.47 |
|
| 9.49 | 3.33–18.47 |
|
| 17.61 | 7.59–29.35 |
|
| 40.03 | 19.71–65.11 |
|
| 18.85 | 8.07–32.66 |
|
| 12.82 | 6.16–21.28 |
|
| 2.04 | 0.02–6.92 |
|
| 11.93 | 5.57–20.46 |
|
| 13.53 | 6.11–22.98 |
|
| 24.92 | 9.68–43.99 |
|
| 94.91 | 79.13–111.14 |
|
| 32.93 | 17.60–48.15 |
|
| 9.24 | 3.42–17.02 |
|
| 41.72 | 23.45–63.12 |
|
| 22.31 | 9.77–38.21 |
|
| 22.31 | 9.77–38.21 |
|
| 32.93 | 17.60–48.15 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases · Slime Mold and Myxomycetes Research
Introduction
The class Agaricomycetes is a diverse class of fungi within the Basidiomycota, encompassing mushrooms, bracket fungi, puff balls, and other fruiting body-forming species, which are found on all continents, including Antarctica, in habitats ranging from tropical rainforests to arctic and alpine ecosystems, and they have important ecological roles as decayers, mycorrhizal symbionts, and parasites of plants and fungi (Nagy et al. 2025). Saprotrophic Agaricomycetes are broadly categorized into wood decayers and litter decomposers, in which wood-decayer Agaricomycetes are further classified into white rot and brown rot based on the chemical and anatomical characteristics of the decayed wood (Floudas 2021). The genus Hyphoderma Wallr. (Hyphodermataceae, Polyporales), typified by H. setigerum (Fr.) Donk, represents one of the most speciose and taxonomically challenging groups of wood-decaying fungi (Donk 1957; Nilsson et al. 2003; Kirk et al. 2008; Yurchenko and Wu 2015). Species of Hyphoderma typically produce resupinate to effuse-reflexed basidiomata with a ceraceous consistency, smooth to tuberculate or hydnoid hymenophores, a monomitic (rarely dimitic) hyphal system with clamp connections, with or without cystidia, suburniform to subcylindrical basidia, and smooth, thin-walled, ellipsoid to subglobose basidiospores (Bernicchia and Gorjón 2010). Species in this genus are white rot fungi (Wu 1997; Floudas et al. 2012; Guan et al. 2021; Duan et al. 2023; Yang et al. 2023). Currently, 217 specific and infraspecific names are recorded in Index Fungorum (accessed 7 February 2026), of which 125 are accepted worldwide, and 41 species are accepted in China (Nakasone 2008; Wu et al. 2010; Martín et al. 2018; Ma et al. 2021; Yang et al. 2023; Su et al. 2024; Li et al. 2025).
The traditionally circumscribed Hyphoderma was shown to be polyphyletic, and 20 species were transferred to Peniophorella within the Hymenochaetales (Larsson 2007). Subsequent studies further examined the delimitation of Hyphoderma and related genera, revealing that eight Peniophorella species form a distinct, well-supported clade separate from Hyphoderma (Tellería et al. 2012; Justo et al. 2017). Molecular systematic analyses have since provided a family-level phylogenetic framework for the order Polyporales and demonstrated that Hyphodermataceae contains only a single genus, Hyphoderma. A recent identification key to Chinese Hyphoderma species was published by Guan et al. (2021). Furthermore, multi-marker phylogenetic analyses based on ITS, nLSU, mtSSU, rpb1, and rpb2 sequence data revealed that Hyphoderma comprises several well-supported clades (Yang et al. 2023, 2025b).
Wood-decay fungi, as a group of macrofungi, possess significant economic and ecological value beyond their taxonomic importance. Biogeographical studies that address the origin, diversification, and distribution patterns of organisms are essential at the genus level for wood-decay fungi (Savinova et al. 2019; He et al. 2024; Hyde et al. 2024; Zhao et al. 2025). Recent advances have substantially improved our understanding of species diversity and divergence times in fungi, including many genera within the Agaricomycetes (He et al. 2019; Varga et al. 2019; Wang et al. 2023; Cui et al. 2025). Currently, more than 10,000 macrofungi species have been described (Hawksworth and Lücking 2017; Niskanen et al. 2023; Hibbett et al. 2025). Elucidating their origins, reconstructing evolutionary histories, and clarifying geographical distributions provide a strong foundation for forest ecosystem research and management (He et al. 2024; Zhao et al. 2025). Although the origin, evolution, and biogeography of certain genera, such as Porodaedalea and Laetiporus, have been well studied (Song and Cui 2017; Zhao et al. 2025), comparable research remains limited for many other genera, including Hyphoderma.
This research continues our long-term investigation into the taxonomy, phylogeny, and fungal diversity of wood-decaying fungi (Dai 2012; Mao et al. 2023; Cho et al. 2023; Pagin-Cláudio and Gugliotta 2024). The study aims to characterize novel and rare fungi, refine phylogenetic relationships within Hyphoderma, and provide new insights into the divergence times and biogeographical history of the genus. It is based on an integrative approach combining morphology and molecular phylogenetics. In addition, five new Hyphoderma species were discovered in Yunnan Province, China. This research expands the taxonomic framework of Hyphoderma through modern taxonomic approaches and enhances our understanding of fungal evolution and geography.
Materials and methods
Sample collection and herbarium specimen preparation
Fresh basidiomata of fungi growing on angiosperm branches were collected from Yunnan Province, China. The samples were photographed in situ, metadata were recorded (Rathnayaka et al. 2025), and fresh macroscopic details were documented following the guidelines provided by Dong et al. (2024). All photographs were stacked and merged using Helicon Focus Pro 7.7.5 software. Specimens were dried in an electric food dehydrator at 40 °C, then sealed and stored in envelope bags (Dong et al. 2024, 2025), and deposited in the herbarium of Southwest Forestry University (SWFC), Kunming, Yunnan Province, P.R. China.
Morphological examinations
Macro-morphological descriptions were based on field notes and photographs captured in the field and laboratory. The descriptions followed Petersen (1996) for color terminology. Micro-morphological characters were obtained from dried specimens observed under a light microscope at 1000× oil immersion (Dong et al. 2024, 2025). Sections were mounted in 5% KOH, 1% Congo Red solution, and 1% phloxine B (C_20_H_2_Br_4_Cl_4_Na_2_O_5_). Other reagents, including Cotton Blue and Melzer’s reagent, were also used to observe micromorphology following the methods of Wu et al. (2019, 2022). To show variation in spore sizes, 5% of measurements were excluded from each end of the range and are shown in parentheses. At least 30 basidiospores from each specimen were measured. Stalks were excluded from basidia measurements, and the hilar appendage was excluded from basidiospore measurements (Yuan and Zhao 2024). The following abbreviations were used: KOH = 5% potassium hydroxide aqueous solution; CB = Cotton Blue; CB– = acyanophilous; CB+ = cyanophilous; IKI = Melzer’s reagent; IKI– = both inamyloid and indextrinoid; L = mean spore length (arithmetic average of all spores); W = mean spore width (arithmetic average of all spores); Q = variation in the L/W ratios between the specimens studied; and n = a/b (number of spores (a) measured from a given number (b) of specimens).
DNA extraction, PCR amplification, sequencing
The CTAB rapid fungal genome extraction kit DN14 (Aidlab Biotechnologies Co., Ltd., Beijing, China) was used to obtain genomic DNA from dried fungal specimens according to the manufacturer’s instructions. The extracted DNA was maintained at −20 °C for long-term storage. Two molecular markers were investigated: the internal transcribed spacer (ITS) region and the nuclear large subunit ribosomal RNA (LSU) gene. PCR primers and conditions are shown in Table 1. PCR products were purified and sequenced at Kunming Tsingke Biological Technology Limited Company (Yunnan Province, China). All newly generated sequences were deposited in NCBI GenBank (Sayers et al. 2025) (Table 1).
Phylogenetic analyses
ITS and nLSU sequences were aligned using MAFFT version 7 (Katoh et al. 2019) with the G-INS-i strategy. Single-gene alignments were manually adjusted using AliView version 1.27 (Larsson 2014; Caboň et al. 2019). The ITS and nLSU sequences were then combined using Mesquite version 3.51. The final sequence alignment, together with related information, was deposited in Figshare (doi: https://doi.org/10.6084/m9.figshare.30939239). Diplomitoporus crustulinus (Bres.) Domanski was selected as the outgroup for phylogenetic analyses following Justo et al. (2017). Maximum parsimony (MP), maximum likelihood (ML), and Bayesian inference (BI) analyses were performed using the combined dataset. Phylogenetic analytical procedures followed Sun et al. (2022). MP analysis was conducted in PAUP* version 4.0b10 (Swofford 2002). All characters were equally weighted, and gaps were treated as missing data. Trees were inferred using a heuristic search with TBR branch swapping and 1,000 random sequence additions. Maxtrees were set to 5,000, branches of zero length were collapsed, and all most-parsimonious trees were saved.
Clade robustness was assessed using bootstrap (BT) analysis with 1,000 replicates (Felsenstein 1985; Guan et al. 2023). Descriptive tree statistics, including tree length (TL), consistency index (CI), retention index (RI), rescaled consistency index (RC), and homoplasy index (HI), were calculated for each most-parsimonious tree. ML analysis was conducted using RAxML-HPC2 through the CIPRES Science Gateway (www.phylo.org) (Miller et al. 2012). Branch support (BS) for ML analysis was estimated using 1,000 bootstrap replicates under the gamma model.
MrModeltest 2.3 (Nylander 2004) was used to determine the best-fit evolutionary model for each dataset. BI was performed in MrBayes 3.2.7a using the GTR+I+G model of DNA substitution with gamma-distributed rate variation across sites (Ronquist et al. 2012). Four Markov chains were run for two independent runs from random starting trees for four million generations for the ITS+nLSU dataset (Fig. 1), with trees and parameters sampled every 1,000 generations. The first one-fourth of all generations was discarded as burn-in. A majority-rule consensus tree was calculated from the remaining trees. Branches were considered significantly supported if they received a maximum likelihood bootstrap value (BS) ≥ 70%, a maximum parsimony bootstrap value (BT) ≥ 50%, or a Bayesian posterior probability (BPP) ≥ 0.95.
Phylogenetic tree generated from an ML analysis based on ITS+nLSU sequences. Branches are labeled with maximum likelihood bootstrap values equal to or higher than 70%, maximum parsimony bootstrap values equal to or higher than 50%, and Bayesian posterior probabilities equal to or higher than 0.95. Novel sequences are printed in bold.
Estimation of divergence times
The divergence-time analysis followed methodologies outlined in previous studies (Wang et al. 2023; Cui et al. 2025). Using a secondary calibration approach, two independent analyses were conducted to estimate divergence times within Hyphoderma.
BEAST v2.6.0 was used to perform molecular clock analyses on the combined alignment derived from Table 1 (Sun et al. 2022; Wang et al. 2023; Cui et al. 2025). A lognormal relaxed molecular clock model and a Yule speciation prior were selected to estimate divergence times and their associated credibility intervals (Zhao and Wu 2017; Spirin et al. 2024). Two calibration points were applied. The first calibration used an offset age with a gamma distribution prior (scale = 20, shape = 1) for Basidiomycota at 400 Mya, based on the divergence between Ascomycota and Basidiomycota inferred from the fossil Paleopyrenomycites devonicus, discovered in Great Britain (Floudas et al. 2012). Trees were sampled every 1,000 generations over 200 million generations, with the first 10% discarded as burn-in. The resulting log files were examined for convergence using Tracer v1.5 (Wang et al. 2023).
Similarly, divergence times of species within Hyphoderma were estimated based on the results obtained above. This analysis included 48 samples representing a single genus of Hyphoderma, with Meripilus giganteus and Physisporinus longicystidius designated as outgroup taxa. Sequence data used in this analysis are provided in Table 2. These analyses were used to estimate the origin and biogeographical history of Hyphoderma species.
Divergence-time analyses were further conducted using BEAST v1.10.4 (He et al. 2024). XML input files were generated in BEAUti v1.10.4 by importing separate NEXUS files for each gene partition. Gene partitions were unlinked for substitution and molecular-clock models but linked for gene trees. Nucleotide substitution models were selected using jModelTest v2, with GTR+I+G applied to ITS and nrLSU. An uncorrelated lognormal relaxed clock model and a Yule speciation prior were used to assume a constant speciation rate per lineage. The prior distribution for the ucld.mean parameter was gamma-distributed (shape = 1.0, scale = 0.001, offset = 0.0) for all genes. Secondary calibrations were implemented using a normally distributed prior on the treeModel.rootHeight parameter (SD = 1), with mean values determined by fossil-based node calibrations. Four independent Markov chain Monte Carlo (MCMC) runs were performed, each consisting of 100,000,000 generations, with parameters sampled every 10,000 generations. Log files were assessed for convergence and mixing using Tracer v1.7.1 (Wang et al. 2023) (http://tree.bio.ed.ac.uk/software/tracer/) to confirm effective sample size (ESS) values exceeded 200. An ultrametric maximum clade credibility (MCC) tree, with mean node ages and 95% highest posterior density (HPD) intervals and per-clade posterior probabilities, was summarized using TreeAnnotator v1.10.4, applying a 25% burn-in and a posterior probability threshold of 0.8. Resulting trees were visualized using iTOL v5 (Wang et al. 2023; Zhao et al. 2023; Cui et al. 2025).
Biogeographic analysis of Hyphoderma
Biogeographic analysis of the genus Hyphoderma was conducted using Reconstruct Ancestral State in Phylogenies (RASP) v4.2 with the Dispersal–Extinction–Cladogenesis (DEC) model, estimated through the BioGeoBEARS package (Cui et al. 2025). Ancestral area reconstruction was performed using a posterior distribution of the aligned dataset 4, which was estimated in BEAST v2.6.5 with 10 million generations. The geographical distribution of Hyphoderma was divided into six regions: A) Asia, B) Europe, C) North America, D) South America, E) Africa, and F) Oceania.
Results
Phylogeny of Hyphoderma
The ITS+nLSU dataset comprised sequences from 79 fungal specimens representing 48 species. The aligned dataset contained 2,040 characters, of which 1,414 were constant, 107 were variable but parsimony-uninformative, and 519 (25%) were parsimony-informative. Maximum parsimony analysis yielded 4,898 equally parsimonious trees (TL = 3.063, CI = 0.3389, HI = 0.6611, RI = 0.7269, and RC = 0.2463). Bayesian inference and ML analyses produced topologies largely congruent with the MP results, with an average standard deviation of split frequencies of 0.019743 for the BI analysis.
The phylogenetic tree inferred from ITS+nLSU sequences (Fig. 1) revealed that the five new taxa were grouped within the genus Hyphoderma. Hyphoderma alboarachnum was closely related to H. floccosum C.L. Zhao and Q.X. Guan. Hyphoderma bambusinum was closely related to H. transiens (Bres.) Parmasto. The species H. fulgens was closely related to H. amoenum (Burt) Donk. Hyphoderma grandineum was closely related to both H. transiens and H. bambusinum. Hyphoderma laceratum was closely related to H. marginatum Z.Y. Duan and C.L. Zhao.
This study conducted a comparative analysis of the macro- and micromorphological characteristics of the five new species and compared the morphological features with those of other Hyphoderma species from Yunnan Province, China (Fig. 7; Table 2).
Results of the pairwise homoplasy index (PHI) test for the combined partial ITS sequence data of Hyphoderma alboarachnum and closely related taxa, using the LogDet transformation and splits decomposition. PHI test results (Φw ≤ 0.05) indicate significant recombination within the dataset.
Results of the pairwise homoplasy index (PHI) test for the combined partial ITS sequence data of Hyphoderma bambusinum and closely related taxa, using the LogDet transformation and splits decomposition. PHI test results (Φw ≤ 0.05) indicate significant recombination within the dataset.
Maximum-clade-credibility chronogram and estimated divergence times of families within Hyphoderma inferred from the combined dataset of ITS and nLSU regions. The estimated divergence times of 95% highest posterior density are indicated as node bars for all clades and are also provided in the upper left of the tree as exact numbers for families within Hymenochaetales. Bayesian posterior probabilities above 0.7 and mean divergence times of clades (crown ages) are labeled before and after the slashes, respectively, at the nodes.
Estimated divergence times for the genus Hyphoderma, derived from molecular clock analyses using a combined dataset of internal transcribed spacers (ITS) and nuclear large ribosomal subunit (nLSU) sequences. Mean divergence times (Ma) and posterior probabilities (PP) > 0.8 are annotated at the internodes, with horizontal blue bars representing the 95% highest posterior density (HPD) intervals for divergence time estimates.
Ancestral state reconstruction of Hyphoderma conducted to determine (A) the origin center and (B) the origin host trees using a dataset comprising internal transcribed spacers (ITS) and nuclear large ribosomal subunit (nLSU) sequences. At each node, a pie chart represents the possible ancestral distributions inferred from dispersal–extinction–cladogenesis (DEC) analyses, as implemented in Reconstruct Ancestral State in Phylogenies (RASP).
Comparison of micromorphological characteristics among the five new Hyphoderma species. A1–A3. Hyphoderma alboarachnum; B1–B3. Hyphoderma bambusinum; C1–C3. Hyphoderma fulgens; D1–D3. Hyphoderma grandineum; E1–E3. Hyphoderma laceratum. Scale bars: 10 μm (A1–E3).
Application of the pairwise homoplasy index (PHI) test to the combined partial ITS sequence dataset revealed no evidence of recombination among phylogenetically related species. No significant recombination events were observed between H. alboarachnum and H. bambusinum and H. floccosum, H. setigerum, H. grandineum, H. transiens, and other phylogenetically closely related species (Figs 2, 3). The test results for the combined partial ITS dataset showed that Φw = 1.0 (Φw > 0.05).
Divergence time estimation for Hyphoderma
The results of divergence time estimation show (Figs 4, 5) that Polyporales emerged earlier, with a mean stem age of 163.71 Mya [95% highest posterior density (HPD) of 131.76–202.70 Mya], which is consistent with previous studies (Song and Cui 2017; Cui et al. 2025). Within Polyporales, Hyphoderma is closely related to the genera Meripilus and Physisporinus, with a mean stem age of 117.76 Mya (95% HPD of 92.38–147.74 Mya) and full support (1.0 PP; Figs 4, 5; Table 3).
The historical biogeography of Hyphoderma
The historical biogeography scenarios inferred using RASP are shown in Fig. 6. Results of the RASP analysis suggested that Asia is likely the center of origin for Hyphoderma species. Among these species, 39 are found in Asia, 18 in Europe, 14 in North America, four in South America, four in Africa, and six in Oceania, suggesting that Asia remains the center of Hyphoderma species. In addition, the ancestral state reconstruction dataset also suggested that Asia is the ancestral region for this genus (Fig. 6).
Taxonomy
Hyphoderma
alboarachnum
Taxon classificationFungiPolyporalesHyphodermataceae
W. Li and C.L. Zhao sp. nov.
928F33EF-A88C-555B-943E-BBB9832BCBA6
859913
Typification.
China • Yunnan Province: Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, GPS coordinates 24°30'N, 97°30'E, altitude 1006 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 19 July 2023, CLZhao 30488 (SWFC 00030488), GenBank: ITS = PV470563.
Basidiomata of Hyphoderma alboarachnum (holotype CLZhao 30488). Scale bars: 1 cm (A); 1 mm (B).
Microscopic structures of Hyphoderma alboarachnum (holotype CLZhao 30488). A. Basidiospores; B. Basidia; C. Basidioles; D. Part of a vertical section of the hymenium. Scale bars: 10 μm (A–D).
Etymology.
alboarachnum (Lat.): refers to the white basidiomata with an arachnoid hymenial surface of the type specimen.
Description.
Basidiomata. Annual, resupinate, adnate, membranaceous, without odor or taste when fresh, up to 12 cm long, 1.5 cm wide, and 100 μm thick. Hymenial surface arachnoid, white when fresh, becoming white to cream when dry. Sterile margin narrow, white, up to 1 mm.
Hyphal system. Monomitic; generative hyphae with clamp connections, colorless, thin-walled, branched, interwoven, 2.5–3.5 µm in diameter, IKI–, CB–, tissues unchanged in KOH.
Hymenium. Cystidia and cystidioles absent. Basidia cylindrical, colorless, thin-walled, smooth, 23.5–30 × 3.5–4.5 µm; basidioles numerous, similar to basidia in shape, but smaller.
Spores. Basidiospores oblong-ellipsoid to cylindrical, colorless, thin-walled, smooth, with inner oil droplets, IKI–, CB–, 5–6(–6.5) × 2–3 µm, L = 5.59 µm, W = 2.66 µm, Q = 2.25 (n = 30/1).
Notes.
In the phylogenetic analysis (Fig. 1), Hyphoderma alboarachnum (CLZhao 30488) was closely related to and formed a sister lineage with H. floccosum (CLZhao 17129, CLZhao 17215), with 100% ML, 91 MP bootstrap support, and a 1.00 BYPP value. However, morphologically, H. floccosum differs from H. alboarachnum by having a farinaceous hymenial surface and larger basidiospores (5–6 × 2–3 µm vs. 6–9.5 × 3–4.5 µm; Guan and Zhao 2021b). In addition, the latter was found in the Wenshan National Nature Reserve at an altitude of 2,480 m. Morphologically, H. alboarachnum is similar to H. mopanshanense and H. membranaceum in having subellipsoid to cylindrical basidiospores (Guan and Zhao 2021a; Ma et al. 2021). However, H. mopanshanense is distinguished from H. alboarachnum by its ceraceous hymenial surface and longer basidiospores (7.8–9.7 × 2.6–3.3 µm vs. 5–6 × 2–3 µm; Ma et al. 2021). The species H. membranaceum differs from H. alboarachnum by its tuberculate hymenial surface and larger basidiospores (11–13.5 × 4.5–5.5 µm vs. 5–6 × 2–3 µm; Guan and Zhao 2021a). Thus, based on morphological and phylogenetic evidence, we introduce our collection as a new species, Hyphoderma alboarachnum.
Hyphoderma
bambusinum
Taxon classificationFungiPolyporalesHyphodermataceae
W. Li and C.L. Zhao sp. nov.
923F16F1-2C18-571F-8500-8187FA307E3D
859914
Typification.
China • Yunnan Province: Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, GPS coordinates 24°30'N, 97°30'E, altitude 1006 m asl., on dead bamboo, leg. C.L. Zhao, 18 July 2023, CLZhao 29903 (SWFC 00029903), GenBank: ITS = PV469674; nLSU = PV819428.
Basidiomata of Hyphoderma bambusinum (holotype CLZhao 29903). Scale bars: 1 cm (A); 1 mm (B).
Microscopic structures of Hyphoderma bambusinum (holotype CLZhao 29903). A. Basidiospores; B. Basidia; C. Basidioles; D. Cystidia; E. Part of a vertical section of the hymenium. Scale bars: 2 μm (A); 10 μm (B–E).
Etymology.
bambusinum (Lat.) refers to the host bamboo on which the fungal species grows.
Description.
Basidiomata. Annual, resupinate, adnate, membranaceous, without odor or taste when fresh, up to 7 cm long, 2.5 cm wide, and 100 μm thick. Hymenial surface smooth, white to cream when fresh, becoming cream to buff when dry. Sterile margin narrow, white to cream, up to 1 mm.
Hyphal system. Monomitic; generative hyphae with clamp connections, colorless, thick, branched, interwoven, 2.5–3 µm in diameter, IKI–, CB–, tissues unchanged in KOH.
Hymenium. Cystidia tubular, colorless, thin-walled, smooth, 21.5–26.5 × 5–6.5 µm. Basidia subclavate to subcylindrical, thin-walled, smooth, slightly flexuous, with four sterigmata and a simple septum at the base, 16–17.5 × 3.5–5.5 µm; basidioles similar to basidia in shape but slightly smaller.
Spores. Basidiospores ellipsoid, colorless, thin-walled, smooth, with oil droplets inside, IKI–, CB–, 4.5–6.5 × 3–4.5(–5) µm, L = 5.55 µm, W = 3.78 µm, Q = 1.47 (n = 30/1).
Notes.
Hyphoderma bambusinum (CLZhao 29903) was found to be phylogenetically closely related to H. transiens (NH 12304 [GB]), with 100% ML, 100 MP bootstrap support, and a 1.00 BYPP value. However, morphologically, H. transiens differs from H. bambusinum by having an odontioid hymenial surface and longer basidiospores (9–13 × 3–4.5 µm vs. 4.5–6.5 × 3–4.5 µm; Parmasto 1968); additionally, the latter was found in Portugal and growing on the bark of Quercus. Morphologically, H. bambusinum is similar to H. cremeoalbum (Höhn. and Litsch.) Jülich. and H. floccosum by having ellipsoid basidiospores (Bernicchia and Gorjón 2010; Guan and Zhao 2021b). However, H. cremeoalbum is distinguished from H. bambusinum by its larger basidiospores (10–14 × 5–6.5 µm vs. 4.5–6.5 × 3–4.5 µm; Bernicchia and Gorjón 2010). The species H. floccosum differs from H. bambusinum by its ceraceous basidiomata and longer, tubular cystidia (37.5–100 × 4–8.5 µm vs. 21.5–26.5 × 5–6.5 µm; Guan and Zhao 2021b). These morphological differences, together with phylogenetic analyses, support the conclusion that our taxon represents a new species, Hyphoderma bambusinum.
Hyphoderma
fulgens
Taxon classificationFungiPolyporalesHyphodermataceae
W. Li and C.L. Zhao sp. nov.
C69F8956-7D84-5290-B5AE-3266E1367A48
859915
Typification.
China • Yunnan Province: Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, GPS coordinates 24°30'N, 97°30'E, altitude 1006 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 3 July 2024, CLZhao 37429 (SWFC 00037429), GenBank: ITS = PV829544; nLSU = PV810095.
Basidiomata of Hyphoderma fulgens (holotype CLZhao 37429). Scale bars: 1 cm (A); 1 mm (B).
Microscopic structures of Hyphoderma fulgens (holotype CLZhao 37429). A. Basidiospores; B. Cystidia; C. Basidioles; D. Basidia; E. Part of a vertical section of the hymenium. Scale bars: 10 μm (A–E).
Etymology.
fulgens (Lat.): refers to the shiny color of the hymenial surface of the specimens.
Description.
Basidiomata. Annual, resupinate, adnate, membranaceous, without odor or taste when fresh, up to 9 cm long, 1.8 cm wide, and 100 μm thick. Hymenial surface smooth, white to cream when fresh, white when dry. Sterile margin narrow, white, up to 1 mm.
Hyphal system. Monomitic; generative hyphae with clamp connections, colorless, thin-walled, branched, interwoven, 2.5–3.2 µm in diameter, IKI–, CB–, tissues unchanged in KOH.
Hymenium. Cystidia clavate, colorless, thin-walled, smooth, encrusted, 30.4–37.1 × 5.5–7.5 µm. Basidia barreled, thin-walled, smooth, slightly flexuous, with four sterigmata and a simple septum at the base, 21.5–26 × 5.5–8 µm; basidioles similar to basidia in shape but slightly smaller.
Spores. Basidiospores subcylindrical, colorless, thin-walled, smooth, with oil droplets inside, IKI–, CB–, (7–)7.5–11.5 × (2–)2.5–4 µm, L = 9.55 µm, W = 3.39 µm, Q = 2.74–2.89 (n = 120/4).
Additional specimens examined (paratypes).
China • Yunnan Province: Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, GPS coordinates 24°30'N, 97°30'E, altitude 1006 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 19 July 2023, CLZhao 30254 (SWFC 00030254); 2 July 2024, CLZhao 37266 (SWFC 00037266); • Mang City, Tongbiguan Provincial Nature Reserve, GPS coordinates 24°42'N, 97°54'E, altitude 1006 m asl., on fallen angiosperm branch, leg. C.L. Zhao, 29 June 2024, CLZhao 36600 (SWFC 00036600); 8 July 2024, CLZhao 39677 (SWFC 00039677); CLZhao 39618 (SWFC 00039618); CLZhao 39474 (SWFC 00039474).
Notes.
In the phylogenetic analysis, the specimens of Hyphoderma fulgens (CLZhao 37429, CLZhao 30254, CLZhao 36600, CLZhao 39474) formed a closely related sister relationship to H. amoenum (Burt) Donk (USO 286622), with 99% ML, 89 MP bootstrap support, and a 0.99 BYPP value. However, morphologically, H. amoenum differs from H. fulgens by having wider basidiospores (9–13 × 5–6 µm vs. 7.5–11.5 × 2.5–4 µm; Tellería et al. 2012). Morphologically, H. fulgens is similar to H. guangdongense and H. tenuissimum by having subcylindrical basidiospores (Guan and Zhao 2021a; Su et al. 2024). However, H. guangdongense is distinguished from H. fulgens by its farinaceous hymenial surface and septate cystidia (Su et al. 2024). In addition, H. tenuissimum differs from H. fulgens by its tuberculate to minutely grandinioid hymenial surface and larger cystidia (50–220 × 6.5–13 µm vs. 30.4–37.1 × 5.5–7.5 µm; Guan and Zhao 2021a). These morphological and phylogenetic data indicate that our collections represent a new species, Hyphoderma fulgens.
Hyphoderma
grandineum
Taxon classificationFungiPolyporalesHyphodermataceae
W. Li and C.L. Zhao sp. nov.
CBC6C098-6CC3-59C5-BD14-178642221919
859916
Typification.
China • Yunnan Province: Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, GPS coordinates 24°30'N, 97°30'E, altitude 1006 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 18 July 2023, CLZhao 30046 (SWFC 00030046), GenBank: ITS = PV470561, nLSU = PV819429.
Basidiomata of Hyphoderma grandineum (holotype CLZhao 30046). Scale bars: 1 cm (A); 1 mm (B).
Microscopic structures of Hyphoderma grandineum (holotype CLZhao 30046). A. Basidiospores; B. Cystidia; C. Basidia; D. Basidioles; E. Part of a vertical section of the hymenium. Scale bars: 10 μm (A–E).
Etymology.
grandineum (Lat.): refers to the grandinioid hymenial surface of the type specimen.
Description.
Basidiomata. Annual, resupinate, adnate, membranaceous, without odor or taste when fresh, up to 7.5 cm long, 2 cm wide, and 100 μm thick. Hymenial surface grandinioid, cream when fresh, becoming cream to yellowish when dry. Sterile margin narrow, white, up to 1 mm.
Hyphal system. Monomitic; generative hyphae with clamp connections, colorless, thin-walled, branched, interwoven, 3–3.6 µm in diameter, IKI–, CB–, tissues unchanged in KOH.
Hymenium. Leptocystidia, colorless, thin-walled, smooth, 29.5–33.5 × 4–5 µm. Basidia clavate, thin-walled, smooth, slightly flexuous, with four sterigmata and a simple septum at the base, 18–19.5 × 4.5–6 µm; basidioles similar to basidia in shape but slightly smaller.
Spores. Basidiospores cylindrical, colorless, thin-walled, smooth, IKI–, CB–, (6–)7–9 × 2.5–4 µm, L = 7.9 µm, W = 3.2 µm, Q = 2.48–2.5 (n = 60/2).
Additional specimen examined (paratype).
China • Yunnan Province: Dehong, Ruili City, Tongbiguan Provincial Nature Reserve, GPS coordinates 23°22'N, 97°18'E, altitude 976 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 2 December 2024, CLZhao 43328(SWFC 00043328), GenBank: ITS = PV470562, nLSU = PV819430.
Notes.
In the phylogenetic analysis, Hyphoderma grandineum (CLZhao 30046, CLZhao 43328) formed a closely related sister relationship with the clade consisting of H. transiens (Bres.) Parmasto (NH 12304 [GB]) and H. bambusinum (CLZhao 29903), with 100% ML, 100 MP bootstrap support, and a 1.00 BYPP value. However, morphologically, H. transiens differs from H. grandineum by having a whitish to ochraceous hymenial surface and longer basidiospores (9–13 × 3–4.5 µm vs. 7–9 × 2.5–4 µm; Parmasto 1968); in addition, the latter was found in Portugal. Morphologically, H. grandineum is similar to H. tropicum and H. tenuissimum by having a tuberculate to odontoid hymenial surface (Guan et al. 2021; Duan et al. 2023). However, H. tropicum is distinguished from H. grandineum by its coriaceous hymenial surface and moniliform cystidia (Duan et al. 2023). In addition, H. tenuissimum differs from H. grandineum by its larger cystidia (50–220 × 6.5–13 µm vs. 29.5–33.5 × 4–5 µm; Guan et al. 2021). These morphological and phylogenetic evidences show that our collections represent a new species, Hyphoderma grandineum.
Hyphoderma
laceratum
Taxon classificationFungiPolyporalesHyphodermataceae
W. Li and C.L. Zhao sp. nov.
244B55E2-697E-5829-87E3-95218FA73B9B
861118
Typification.
China • Yunnan Province: Diqing, Weixi County, Weideng Town, Fuchuan Village, GPS coordinates 27°17'N, 99°16'E, altitude 1700 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 12 October 2023, CLZhao 34242 (SWFC 00034242), GenBank: ITS = PV829552, nLSU = PV810101.
Basidiomata of Hyphoderma laceratum (holotype CLZhao 34242). Scale bars: 1 cm (A); 1 mm (B).
Microscopic structures of Hyphoderma laceratum (holotype CLZhao 34242). A. Basidiospores; B. Cystidia; C. Basidioles; D. Basidia; E. Part of a vertical section of the hymenium. Scale bars: 10 μm (A–E).
Etymology.
laceratum (Lat.): refers to the lacerate hymenial surface of the type specimens.
Description.
Basidiomata. Annual, resupinate, adnate, membranaceous, without odor or taste when fresh, up to 10.5 cm long, 2.5 cm wide, and 150 μm thick. Hymenial surface smooth, white to cream when fresh, becoming cream when dry. Sterile margin narrow, white, up to 1 mm.
Hyphal system. Monomitic; generative hyphae with clamp connections, colorless, thin-walled, branched, interwoven, 2.5–3.3 µm in diameter, IKI–, CB–, tissues unchanged in KOH.
Hymenium. Cystidia, two types: (1) Clavate, colorless, thin-walled, smooth, 27–29 × 6.5–8 µm; (2) Capitate, colorless, thin-walled, smooth, 26–30 × 7.5–8.5 µm. Basidia subcylindrical, thin-walled, smooth, slightly flexuous, with four sterigmata and a simple septum at the base, 18–20.5 × 5–7 µm; basidioles similar to basidia in shape but slightly smaller.
Spores. Basidiospores sub-ellipsoid, colorless, thin-walled, smooth, with guttulate, IKI–, CB–, (6–)6.5–9 × (2.5–)3–4(–4.5) µm, L = 7.72 µm, W = 3.49 µm, Q = 2.18–2.35 (n = 330/11).
Additional specimens examined (paratypes).
China • Yunnan Province: Diqing, Weixi County, Weideng Town, Fuchuan Village, GPS coordinates 27°17'N, 99°16'E, altitude 1700 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 12 October 2023, CLZhao 34163 (SWFC 00034163), CLZhao 34138 (SWFC 00034138); • Zhonglu Town, GPS coordinates 27°17'N, 99°16'E, altitude 1811 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 15 October 2023, CLZhao 34961 (SWFC 00034961), CLZhao 35040 (SWFC 00035040), CLZhao 34718 (SWFC 00034718), CLZhao 34672 (SWFC 00034672), CLZhao 34958 (SWFC 00034958); • Qujing, Zhanyi District, Lingjiao Town, Xiajia Village, GPS coordinates 25°8'N, 103°6'E, altitude 2040 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 6 March 2023, CLZhao 27226 (SWFC 00027226); • Dehong, Yingjiang County, Tongbiguan Provincial Nature Reserve, GPS coordinates 24°30'N, 97°30'E, altitude 1006 m asl., on the fallen angiosperm branch, leg. C.L. Zhao, 19 July 2023, CLZhao 30414 (SWFC 00030414), CLZhao 30552 (SWFC 00030552).
Notes.
In the phylogenetic analysis, Hyphoderma laceratum (CLZhao 34958, CLZhao 34672, CLZhao 34961, CLZhao 34242) formed a closely related sister relationship with H. marginatum (CLZhao 3404), with 99% ML bootstrap support and a 0.99 BYPP value. However, morphologically, H. marginatum differs from H. laceratum by having a cream hymenial surface and longer basidia (21–31.5 × 5–7 µm vs. 18–20.5 × 5–7 µm; Duan et al. 2023). Morphologically, H. laceratum is similar to H. niveomarginatum by having a cream hymenial surface (Yang et al. 2023). Further, H. niveomarginatum is distinguished from H. laceratum by its moniliform cystidia (Yang et al. 2023). Based on these morphological differences and phylogenetic analysis, we propose it as a new species, Hyphoderma laceratum.
Discussion and conclusion
Many recently described wood-decaying fungal taxa have been reported from subtropical and tropical regions, and the genus Hyphoderma represents one of the most rapidly expanding groups in these areas (Nilsson et al. 2003; Wu et al. 2010; Martín et al. 2018; Guan and Zhao 2021a, b). Molecular systematics and taxonomy are essential for resolving diversity within Hyphoderma because species in this genus exhibit substantial variation in basidiomata morphology, including farinaceous, coriaceous, and membranaceous forms, as well as hymenophore types ranging from smooth to grandinioid (Guan et al. 2023; Su et al. 2024; Li et al. 2025; Yang et al. 2025b). Despite this variation, the hymenial surface predominantly ranges from cream to whitish (Yurchenko and Wu 2014a, b; Guan et al. 2023; Wang et al. 2025). The morphological delineation of Hyphoderma species is challenging, and continuous descriptions of new taxa further reduce the diagnostic value of the limited morphological characteristics available for distinguishing species within each morpho-ecological group (Yang et al. 2025b). In the present study, we described five new species, viz., H. alboarachnum, H. bambusinum, H. fulgens, H. grandineum, and H. laceratum. These taxa represent distinct hymenophore types and highlight the necessity of integrative taxonomy for accurate species identification.
Wood-decaying fungi occupy a broad range of substrates, including living trees, decorticated wood of dead branches and trunks, and processed wood materials (Dai et al. 2015; Cui et al. 2019; Zhao et al. 2023; Zhang et al. 2023; He et al. 2024; Hyde et al. 2024; Luo et al. 2024; Deng et al. 2025; Yang et al. 2025a). Recent biogeographical studies, particularly in Polyporaceae and Hymenochaetaceae, have substantially improved our understanding of diversification patterns among wood-decaying fungi (Yang et al. 2023; Zhao et al. 2025). Consistent with these findings, our results show that the geographic distribution of Hyphoderma follows a distinct and well-structured biogeographical pattern, similar to that observed in other major groups of wood-decaying fungi (Song and Cui 2017; Yang et al. 2023).
Molecular dating analyses have provided important insights into the evolution of wood-decaying fungi (Zhao et al. 2025). According to our divergence-time estimates based on ITS and nLSU sequence datasets, Hyphoderma likely originated during the Cretaceous period. The mean stem age is 117.76 Mya (95% HPD = 92.38–147.74 Mya), with strong support (PP = 1.0). This deep evolutionary origin is consistent with the long-term ecological continuity of Cretaceous forest ecosystems (Martín et al. 2018; Cui et al. 2025). Our biogeographical reconstruction further suggests that Hyphoderma most likely originated in Asia, a recognized global biodiversity hotspot (Cui et al. 2025). Dispersal between East Asia and North America may have occurred through Beringia (Cui et al. 2025; Zhao et al. 2025). In contrast, later vicariance events, such as the opening of the Bering Strait, probably restricted gene flow between the Old and New Worlds and contributed to the present distribution of the genus (Hibbett 2001; Cai et al. 2014; Zhao et al. 2025).
In summary, by combining extensive field collections, detailed morphological observations, and multi-locus phylogenetic analyses, we reconstructed the evolutionary relationships within Hyphoderma and uncovered five previously unknown species. Our divergence-time and biogeographical analyses provide a refined understanding of the evolutionary origin and historical distribution of the genus, thereby contributing valuable insights into the broader evolutionary history of wood-decaying fungi. The present study fills knowledge gaps regarding wood-inhabiting fungi by reporting new taxa and providing detailed morphological descriptions and phylogenetic analyses while contributing to the enrichment of fungal diversity in Asia.
Supplementary Material
XML Treatment for Hyphoderma alboarachnum
XML Treatment for Hyphoderma bambusinum
XML Treatment for Hyphoderma fulgens
XML Treatment for Hyphoderma grandineum
XML Treatment for Hyphoderma laceratum
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bernicchia A, Gorjón SP (2010) Fungi Europaei 12: Corticiaceae s.l. Edizioni Candusso, Lomazzo, 1–1007.
- 2Boonmee S, Wanasinghe DN, Calabon MS, Huanraluek N, Chandrasiri SKU, Jones GEB, Rossi W, Leonardi M, Singh SK, Rana S, Singh PN, Maurya DK, Lagashetti AC, Choudhary D, Dai YC, Zhao CL, Mu YH, Yuan HS, He SH, Phookamsak R, Jiang HB, Martín MP, Dueñas M, Tellería MT, Kałucka IL, Jagodziński AM, Liimatainen K, Pereira DS, Phillips AJL, Suwannarach N, Kumla J, Khuna S, Lumyong S, Potter TB, Shivas RG, Sparks AH, Vaghefi N, Abdel-Wahab MA, Abdel-Aziz FA, Li GJ, Lin WF, Singh U, Bhatt RP, Lee HB, Nguyen · doi ↗ · pubmed ↗
- 3Caboň M, Li GJ, Saba M, Kolařík M, Jančovičová S, Khalid AN, Moreau PA, Wen HA, Pfister DH, Adamčík S (2019) Phylogenetic study documents different speciation mechanisms within the Russula globispora lineage in boreal and arctic environments of the Northern Hemisphere. IMA Fungus 10: 5. 10.1186/s 43008-019-0003-9PMC 732566732647614 · doi ↗ · pubmed ↗
- 4Cai Q, Tulloss RE, Tang LP, Tolgor B, Zhang P, Chen ZH, Yang ZL (2014) Multi-locus phylogeny of lethal amanitas: Implications for species diversity and historical biogeography. BMC Evolutionary Biology 14: 143. 10.1186/1471-2148-14-143PMC 409491824950598 · doi ↗ · pubmed ↗
- 5Cho Y, Kim D, Lee Y, Jeong J, Hussain S, Lim YW (2023) Validation of Fuscoporia (Hymenochaetales, Basidiomycota) ITS sequences and five new species based on multi-marker phylogenetic and morphological analyses. IMA Fungus 14: 12. 10.1186/s 43008-023-00117-6PMC 1030879337381063 · doi ↗ · pubmed ↗
- 6Cui BK, Li HJ, Ji X, Zhou JL, Song J, Si J, Yang ZL, Dai YC (2019) Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China. Fungal Diversity 97: 137–392. 10.1007/s 13225-019-00427-4 · doi ↗
- 7Cui YJ, Wang CG, Dai YC, Liu S, Ren YH, Schultes NP, Kaishian PO, Paine E, Yuan Y, Li DW, Zhao H (2025) Phylogeny, divergence times, and biogeography of the phytopathogenic fungal genus Phaeolus (Basidiomycota, Polyporales). Journal of Systematics and Evolution 00: 1–15. 10.1111/jse.13187 · doi ↗
- 8Dai YC (2012) Polypore diversity in China with an annotated checklist of Chinese polypores. Mycoscience 53: 49–80. 10.1007/s 10267-011-0134-3 · doi ↗