Dentipellicula hainanensis (Hericiaceae, Agaricomycetes) and Phylloporia rigida (Hymenochaetaceae, Agaricomycetes), two new species of wood-inhabiting fungi from Hainan Province, South China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
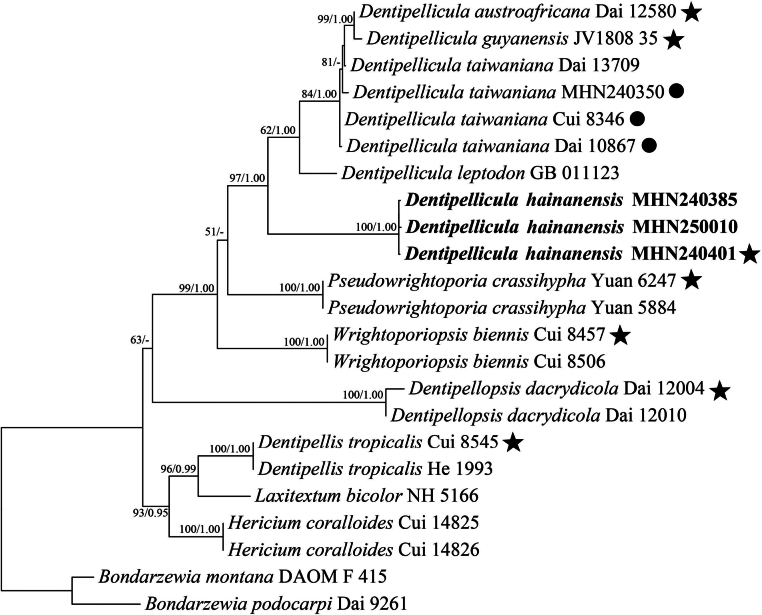 Figure 1
Figure 1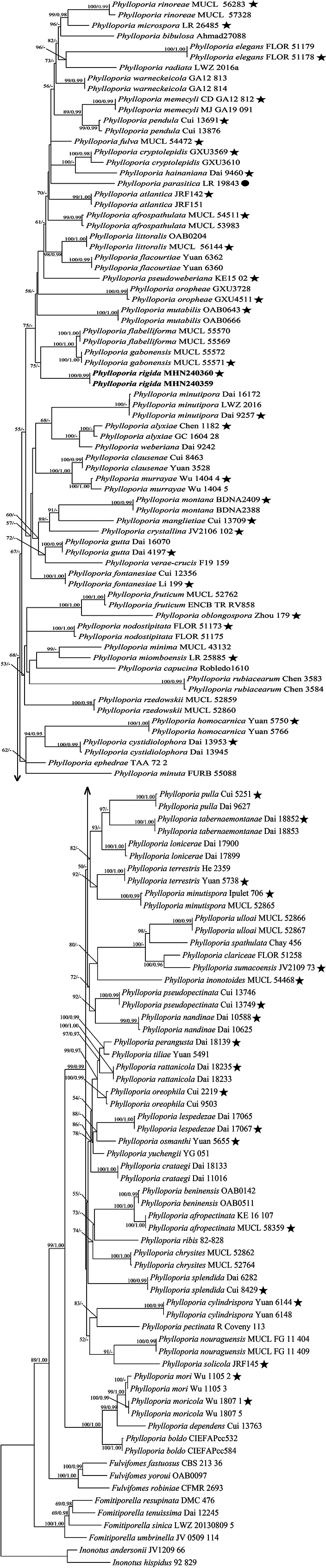 Figure 2
Figure 2 Figure 3
Figure 3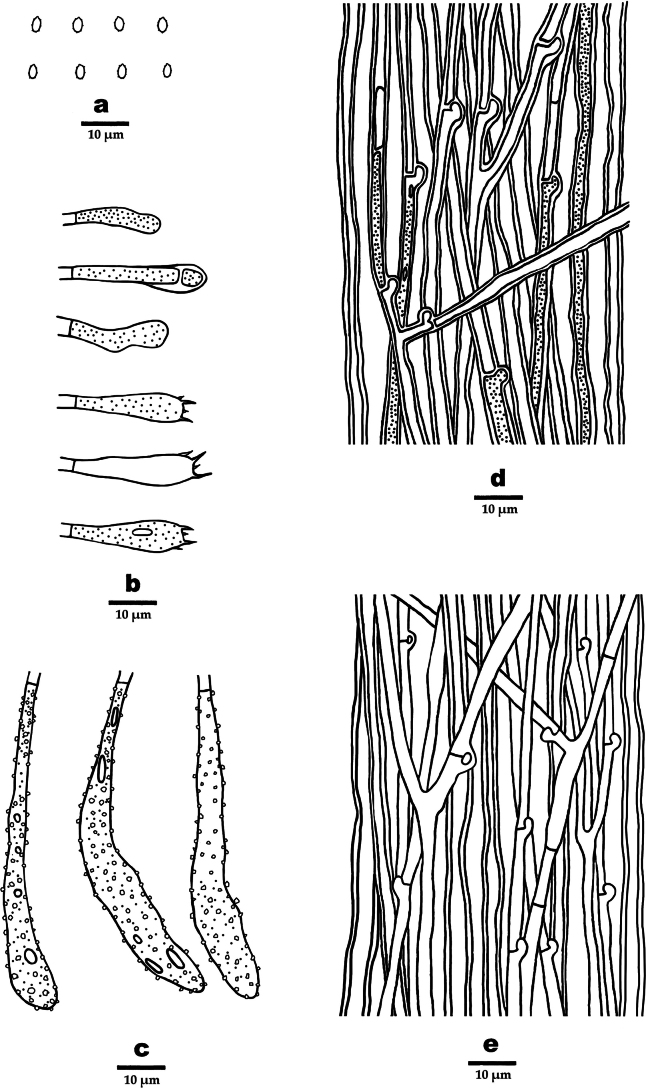 Figure 4
Figure 4 Figure 5
Figure 5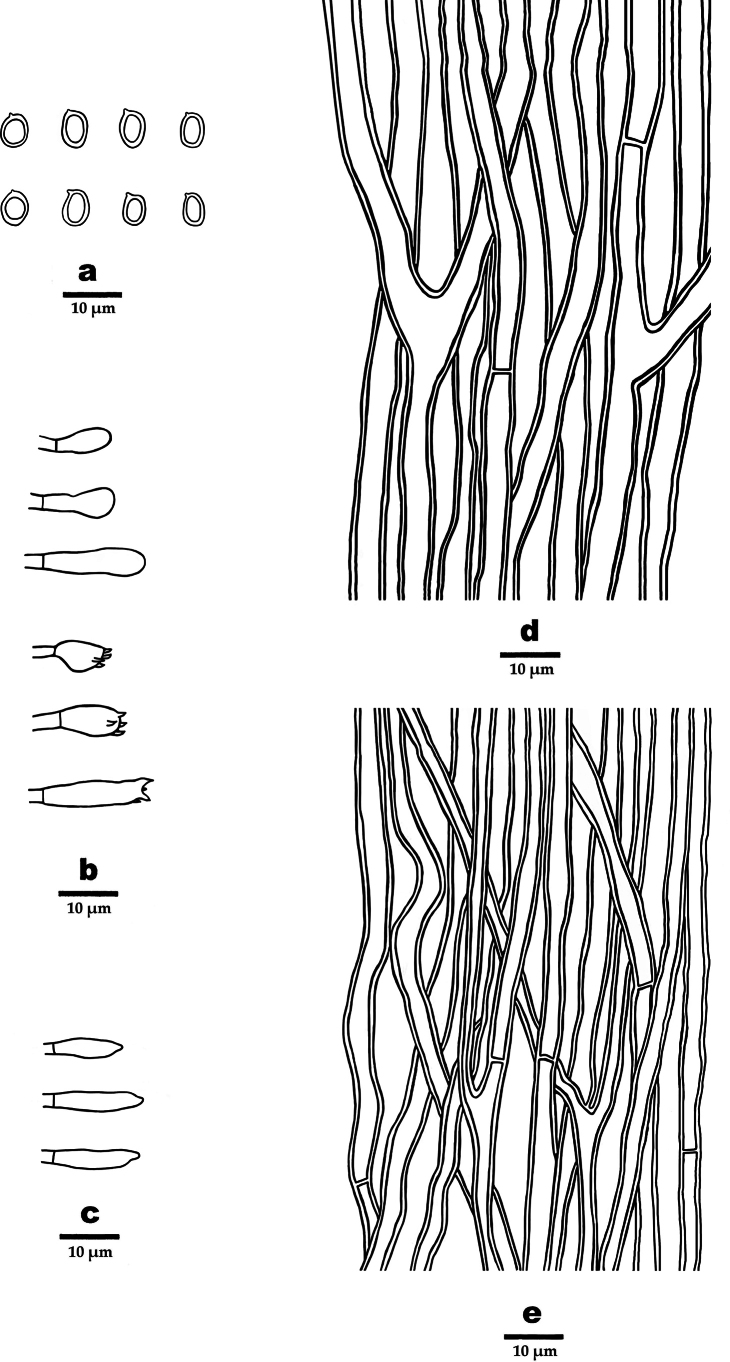 Figure 6
Figure 6| Species | Specimen no. | Locality | GenBank accession no. | Reference | |
|---|---|---|---|---|---|
| ITS | nLSU | ||||
|
| DAOM F-415 | Canada | DQ 200923 | DQ 234539 |
|
|
| Dai 9261 | China | KJ 583207 | KJ 583221 |
|
|
| Dai 12580 | South Africa | KJ 855274 | KJ 855275 |
|
|
| JV1808/35 | Uganda |
|
|
|
|
| MHN240385 | China |
|
| This study |
|
| MHN240401 | China |
|
| This study |
|
| MHN250010 | China |
| - | This study |
|
| GB 011123 | China | EU 118625 | EU 118625 |
|
|
| Cui 8346 | China | JQ 349114 | JQ 349100 |
|
|
| Dai 10867 | China | JQ 349115 | JQ 349101 |
|
|
| Dai 13709 | China | MH 085941 | MH 085957 |
|
|
| MHN240350 | China |
|
| This study |
|
| Cui 8545 | China | KR 108236 | KR 108240 |
|
|
| He 1993 | China | KR 108237 | KR 108241 |
|
|
| Dai 12004 | China | JQ 349104 | JQ 349089 |
|
|
| Dai 12010 | China | - | JQ 349090 |
|
|
| DMC 476 | Cameroon |
|
|
|
|
| LWZ 20130809-5 | China |
|
|
|
|
| Dai 12245 | China |
|
|
|
|
| JV 0509/114 | USA |
|
|
|
|
| CBS 213.36 | Philippines |
|
|
|
|
| CFMR 2693 | USA |
|
|
|
|
| OAB0097 | Benin |
|
|
|
|
| Cui 14825 | China | MH 085948 | MH 085962 |
|
|
| Cui 14826 | China | MH 085949 | MH 085963 |
|
|
| JV1209_66 | USA |
|
|
|
|
| 92-829 | Germany |
|
|
|
|
| NH 5166 | Sweden | AF 310102 | AF 310102 |
|
|
| KE 16 107 | Kenya | - |
|
|
|
| MUCL 58359 | Kenya | - |
|
|
|
| MUCL 54511 | Gabon | - |
|
|
|
| MUCL 53983 | Gabon | - |
|
|
|
| Chen 1182 | China | - |
|
|
|
| GC 1604-28 | China | - |
|
|
|
| JRF142 | Brazil | - |
|
|
|
| JRF151 | Brazil | - |
|
|
|
| OAB0142 | Brazil |
|
|
|
|
| OAB0511 | Benin | - |
|
|
|
| Ahmad27088 | Pakistan | - |
|
|
|
| CIEFAPcc532 | Chile | - |
|
|
|
| CIEFAPcc584 | Chile | - |
|
|
|
| Robledo1610 | Argentina | - |
|
|
|
| MUCL 52862 | Mexico | - |
|
|
|
| MUCL 52764 | Mexico | - |
|
|
|
| FLOR:51258 | Brazil | - |
|
|
|
| Yuan 3528 | China | - |
|
|
|
| Cui 8463 | China |
|
|
|
|
| Dai 18133 | China |
|
|
|
|
| Dai 11016 | China | - |
|
|
|
| GXU3569 | China |
|
|
|
|
| GXU3610 | China |
|
|
|
|
| JV2106/102 | Ecuador | - |
|
|
|
| Yuan 6144 | China | - |
|
|
|
| Yuan 6148 | China | - |
|
|
|
| Dai 13953 | China | - |
|
|
|
| Dai 13945 | China | - |
|
|
|
| Cui 13763 | China |
|
|
|
|
| FLOR:51179 | Brazil | - |
|
|
|
| FLOR:51178 | Brazil | - |
|
|
|
| TAA 72-2 | Turkmenistan |
|
|
|
|
| MUCL 55570 | Gabon |
|
|
|
|
| MUCL 55569 | Gabon |
|
|
|
|
| Yuan 6362 | China | - |
|
|
|
| Yuan 6360 | China | - |
|
|
|
| Cui 12356 | China |
|
|
|
|
| Li 199 | China | - |
|
|
|
| MUCL 52762 | Mexico | - |
|
|
|
| ENCB TR&RV858 | Mexico | - |
|
|
|
| MUCL 54472 | Gabon | - |
|
|
|
| MUCL 55572 | Gabon |
|
|
|
|
| MUCL 55571 | Gabon |
|
|
|
|
| Dai 16070 | China |
|
|
|
|
| Dai 4197 | China | - |
|
|
|
| Dai 9460 | China | - |
|
|
|
| Yuan 5766 | China | - |
|
|
|
| Yuan 5750 | China |
|
|
|
|
| MUCL 54468 | China | - |
|
|
|
| Dai 17065 | China |
|
|
|
|
| Dai 17067 | China |
|
|
|
|
| MUCL: 56144 | Gabon | - |
|
|
|
| OAB0204 | Benin |
|
|
|
|
| Dai 17900 | China |
|
|
|
|
| Dai 17899 | China |
|
|
|
|
| Cui 13709 | China |
|
|
|
|
| CD-GA12-812 | Gabon | - |
|
|
|
| MJ-GA19-091 | Gabon | - |
|
|
|
| LR 26485 | Zimbabwe | - |
|
|
|
| MUCL 43132 | Australia | - |
|
|
|
| FURB 55088 | Brazil | - |
|
|
|
| Dai 16172 | China | - |
|
|
|
| LWZ-2016 | China | - |
|
|
|
| Dai 9257 | China | - |
|
|
|
| Ipulet 706 | Uganda | - |
|
|
|
| MUCL 52865 | Congo | - |
|
|
|
| LR 25885 | Zimbabwe | - |
|
|
|
| BDNA2409 | Brazil | - |
|
|
|
| BDNA2388 | Brazil | - |
|
|
|
| Wu 1105 2 | China | - |
|
|
|
| Wu 1105 3 | China | - |
|
|
|
| Wu 1807 1 | China | - |
|
|
|
| Wu 1807 5 | China | - |
|
|
|
| Wu 1404-4 | China | - |
|
|
|
| Wu 1404-5 | China | - |
|
|
|
| OAB0643 | Benin |
|
|
|
|
| OAB0666 | Benin |
|
|
|
|
| Dai 10588 | China | - |
|
|
|
| Dai 10625 | China | - |
|
|
|
| FLOR:51173 | Brazil |
|
|
|
|
| FLOR:51175 | Brazil | - |
|
|
|
| MUCL/FG-11-404 | Guyana | - |
|
|
|
| MUCL/FG-11-409 | Guyana | - |
|
|
|
| Zhou 179 | China |
|
|
|
|
| Cui 2219 | China |
|
|
|
|
| Cui 9503 | China | - |
|
|
|
| GXU3728 | China |
|
|
|
|
| GXU4511 | China |
|
|
|
|
| Yuan 5655 | China | - |
|
|
|
| LR 19843 | Argentina |
| - |
|
|
| R.Coveny 113 | Australia | - |
|
|
|
| Cui 13691 | China | - |
|
|
|
| Cui 13876 | China | - |
|
|
|
| Dai 18139 | China |
|
|
|
|
| Cui 13746 | China | - |
|
|
|
| Cui 13749 | China | - |
|
|
|
| KE15-02 | Kenya | - |
|
|
|
| Cui 5251 | China | - |
|
|
|
| Dai 9627 | China | - |
|
|
|
| LWZ-2016a | China | - |
|
|
|
| Dai 18233 | China | - |
|
|
|
| Dai 18235 | China |
|
|
|
|
| 82-828 | Germany | - |
|
|
|
| MHN240359 | China |
| - | This study |
|
| MHN240360 | China |
| - | This study |
|
| MUCL: 56283 | Gabon | - |
|
|
|
| MUCL: 57328 | Gabon | - |
|
|
|
| Chen 3583 | China | - |
|
|
|
| Chen 3584 | China | - |
|
|
|
| MUCL 52859 | Mexico | - |
|
|
|
| MUCL 52860 | Mexico | - |
|
|
|
| JV2109/73 | Ecuador | - |
|
|
|
| JRF145 | Brazil | - |
|
|
|
| Chay 456 | Mexico | - |
|
|
|
| Dai 6282 | China | - |
|
|
|
| Cui 8429 | China | - |
|
|
|
| Dai 18852 | China | - |
|
|
|
| Dai 18853 | China | - |
|
|
|
| Yuan 5738 | China | - |
|
|
|
| He 2359 | China |
|
|
|
|
| Yuan 5491 | China | - |
|
|
|
| MUCL 52866 | Mexico | - |
|
|
|
| MUCL 52867 | Mexico | - |
|
|
|
| F19-159 | Mexico | - |
|
|
|
| GA12-813 | Gabon | - |
|
|
|
| GA12-814 | Gabon | - |
|
|
|
| Dai 9242 | China | - |
|
|
|
| YG 051 | Uzbekistan | - |
|
|
|
| Yuan 6247 | China | KM 107873 | KM 107892 |
|
|
| Yuan 5884 | China | KM 107872 | KM 107891 |
|
|
| Cui 8457 | China | KJ 807066 | KJ 807074 |
|
|
| Cui 8506 | China | KJ 807067 | KJ 807075 |
|
| Specimen no. | BLAST results (based on ITS) | Query Cover | Per. Ident |
|---|---|---|---|
| MHN240385 |
| 95% | 83.31% |
| MHN240401 |
| 95% | 83.31% |
| MHN250010 |
| 85% | 90.68% |
| MHN240359 | 100% | 87.71% | |
| MHN240360 | 99% | 87.08% |
| Species | Ecological habits | Basidiomata | Pores | Context | Hyphal system | Cystidioles | Basidiospores (µm) |
|
|---|---|---|---|---|---|---|---|---|
|
| on living | perennial, pileate | 6–8 per mm | duplex with a black line | dimitic | present | (2.8−)3–3.2(−3.5) × (2−)2.2–2.7 |
|
|
| on living | annual, pileate | 8–9 per mm | duplex with two black zones | monomitic | absent | 3–3.5 × 2–3 |
|
|
| on living | perennial, pileate | 7–9 per mm | duplex with a black line | monomitic | absent | 2.5–3.3 × 2–2.8 |
|
|
| on the branch of living | annual, pileate | 5–6 per mm | duplex with a black line | monomitic | absent | (3.2–)3.6–4.6(–4.7) × (2.2 )2.5–3.5(–3.9) |
|
|
| on living angiosperm | annual, pileate | 6–8 per mm | duplex with a black line | monomitic | absent | 3.5–4 × 1.5–2 |
|
|
| on living angiosperm | annual, pileate | 8–10 per mm | duplex with a black line | monomitic | present | 2.5–3 × 2.1–2.8 |
|
|
| on living angiosperm | perennial, pileate to pendent | 7–9 per mm | homogenous | monomitic | present | 3–3.4 × 2.7–3 |
|
|
| on living | annual, pileate | 5–7 per mm | duplex without black line | monomitic | absent | 3.7–4.4 × 2.5–3.2 |
|
|
| on living | annual, pileate | 10–12 per mm | duplex with a black line | monomitic | absent | 2.5–3 × 2–2.5 |
|
|
| on dead | perennial, pileate | 7–9 per mm | duplex with a black line | monomitic | absent | 3–4 × 2–2.5 |
|
|
| on living angiosperm | annual, pileate | 4–6 per mm | duplex with a black line | monomitic | absent | 4.6–5.6 × 3–3.6 |
|
|
| on dead angiosperm | annual, pileate | 4–6 per mm | homogenous | monomitic | absent | 4–4.9 × 2.5–3 |
|
|
| on living | annual, pileate | 8–9 per mm | duplex with a black line | monomitic | present | 3.1–3.7 × 2.2–2.7 |
|
|
| on living | perennial, pileate | 6–8 per mm | duplex with a black line | dimitic | absent | 3–3.3 × 2.1–2.5 |
|
|
| on living | perennial, pileate | 6–8 per mm | duplex with a black line | monomitic | absent | 3–3.5 × 2–2.5 |
|
|
| on living angiosperm | annual, pileate | 12–15 per mm | duplex with a black line | dimitic | absent | 2.5–3 × 2–2.4 |
|
|
| on living angiosperm shrub | annual, pileate | 12–13 per mm | duplex with a black line | monomitic | absent | (2.9−)3 3.5(−3.6) × (1.8−)2–2.3(−2.5) |
|
|
| on fallen trunk of | perennial, resupinate to effused reflexed | 7–9 per mm | homogeneous | dimitic | present | (3.8−)4–4.7(−5) × 3.1–3.6(−4) |
|
|
| on living tree of | perennial, resupinate to effused reflexed | 7–9 per mm | duplex with a black line | dimitic | absent | (3.2–)3.5–4 × (2.5 )2.8–3.1(−3.3) |
|
|
| on living | perennial, pileate | 8–10 per mm | duplex without black line | dimitic | absent | (3−)3.2–3.7 × (2.5−)2.8–3(−3.2) |
|
|
| on living | annual, pileate | 5–6 per mm | duplex with a black line | monomitic | absent | 3.5–4.2 × 2–2.5 |
|
|
| on living angiosperm | annual, pileate | 2–4 per mm | homogenous | monomitic | absent | 4–4.8 × 2–2.5 |
|
|
| on living angiosperm | annual, pileate | 7–9 per mm | duplex with a black line | monomitic | absent | 3–3.7 × 2–3 |
|
|
| on branch of living | annual, pileate | 3–4 per mm | duplex with a black line | dimitic | absent | (4.1–) 4.2–5.3(–5.7) × (2.5–)2.6–3.8(–3.9) |
|
|
| on living | annual, pileate | 7–9 per mm | duplex with a black line | monomitic | present | 2.9–3.4 × 2–2.6 |
|
|
| on living angiosperm | perennial, pileate | 9–11 per mm | duplex with a black line | dimitic | present | 2.5–2.9 × 1.8–2 |
|
|
| on living angiosperm | annual, pileate | 7–9 per mm | duplex with a black line | dimitic | absent | 3.5–4 × 2.5–3 |
|
|
| on living angiosperm | annual, pileate | 8–9 per mm | duplex with a black line | dimitic | absent | 3–3.5 × 2–3 |
|
|
| unknown substrate | Annual, pileate | 11–12 per mm | duplex | dimitic | absent | 2.8–3.3 × 2.3–2.8 |
|
|
| on living lianas | annual, pileate | 8–10 per mm | duplex with a black line | monomitic | absent | 2.5–3.5 × 2–2.5 |
|
|
| on lianas | perennial, pileate | 9–11 per mm | duplex with a black line | dimitic | present | 2.9–3.1 × 2–2.1 | This study |
|
| on living | annual, pileate | 5–7 per mm | duplex with a black line | monomitic | present | 2.3–4.5 × 2.2–3.8 |
|
|
| on living trees of | perennial, pileate | 7–9 per mm | duplex with a black line | dimitic | present | 3–3.5 × (2.2−)2.3–2.8(−2.9) |
|
|
| on living angiosperm | annual, pileate | 9–10 per mm | duplex with a black line | monomitic | present | 3.2–4 × 2.1–2.8 |
|
|
| on living angiosperm | perennial, pileate | 13–16 per mm | duplex with a black line | dimitic | absent | 2.7–3 × 1.9–2.1 |
|
|
| on ground | annual, stipitate | 10–14 per mm | duplex with a black line | monomitic | absent | 2.5–3.3 × 1.8–2.5 |
|
|
| on living | perennial, pileate | 9–12 per mm | duplex with a black line | monomitic | absent | 3–3.4 × 2–2.5 |
|
|
| on living angiosperm | annual, pileate | 6–8 per mm | duplex with a black line | monomitic | absent | 3.4 4.1 × 2.2–3 |
|
| 1 | Pileal surface buff yellow to pinkish-buff |
|
| – | Pileal surface with different colour |
|
| 2 | Basidiospores > 3.5 µm long |
|
| – | Basidiospores < 3.5 µm long |
|
| 3 | Spines 8–10 per mm |
|
| – | Spines 5–7 per mm |
|
| 4 | Basidiocarps resupinate; gloeoplerous hyphae present |
|
| – | Basidiocarps effused-reflexed; gloeoplerous hyphae absent |
|
| 1 | Basidiomata stipitate |
|
| – | Basidiomata sessile |
|
| 2 | Hyphal system dimitic |
|
| – | Hyphal system monomitic |
|
| 3 | Pores > 12 per mm |
|
| – | Pores < 12 per mm |
|
| 4 | Basidiomata perennial; basidiospores 1.9–2.1 μm wide, basidia 11–13 μm long |
|
| – | Basidiomata annual; basidiospores 2–2.5 μm wide, basidia 5–7 μm long |
|
| 5 | Basidiomata perennial |
|
| – | Basidiomata annual |
|
| 6 | Cystidioles absent |
|
| – | Cystidioles present |
|
| 7 | Basidiocarps resupinate to effused reflexed |
|
| – | Basidiocarps pileate |
|
| 8 | Basidiomata pendent, basidia 7–18 μm long |
|
| – | Basidiomata not pendent, basidia 13–15 μm long |
|
| 9 | Basidiocarps resupinate to effused reflexed |
|
| – | Basidiocarps pileate |
|
| 10 | Pores > 9 per mm |
|
| – | Pores < 9 per mm |
|
| 11 | Upper tomentum up to 2 mm thick; basidiospores 2.5–2.9 μm long |
|
| – | Upper trichoderm up to 0.5 mm thick; basidiospores 2.9–3.1 μm long |
|
| 12 | Pores 6–8 per mm |
|
| – | Pores 7–9 per mm |
|
| 13 | Pores > 9 per mm |
|
| – | Pores < 9 per mm |
|
| 14 | Pores > 4 per mm |
|
| – | Pores < 4 per mm |
|
| 15 | Basidiomata pendent; basidiospores 3.5–4 μm long |
|
| – | Basidiomata not pendent; basidiospores 2.8–3.5 μm long |
|
| 16 | Context homogeneous |
|
| – | Context duplex |
|
| 17 | Basidiospores broadly ellipsoid to subglobose |
|
| – | Basidiospores ellipsoid to oblong-ellipsoid |
|
| 18 | Pores 4–6 per mm; basidiospores 2.5–3 μm wide |
|
| – | Pores 2–4 per mm; basidiospores 2–2.5 μm wide |
|
| 19 | Black line absent in context |
|
| – | Black line present in context |
|
| 20 | Basidiospores 2–2.5 μm wide |
|
| – | Basidiospores 2.5–3.5 μm wide |
|
| 21 | Basidiospores broadly ellipsoid to subglobose |
|
| – | Basidiospores ellipsoid, oblong-ellipsoid to cylindrical |
|
| 22 | Cystidioles present |
|
| – | Cystidioles absent |
|
| 23 | Basidiospores broadly ellipsoid, 3.1–3.7 μm long |
|
| – | Basidiospores subglobose to broadly ellipsoid, 2.5–3 μm long |
|
| 24 | Basidia > 15 μm long |
|
| – | Basidia < 15 μm long |
|
| 25 | Pores > 10 per mm |
|
| – | Pores < 10 per mm |
|
| 26 | Basidia > 5.5 μm long |
|
| – | Basidia < 5.5 μm long |
|
| 27 | Tramal hyphae interwoven |
|
| – | Tramal hyphae subparallel |
|
| 28 | Pileal surface not radially striate, two black zones present in context |
|
| – | Pileal surface radially striate, one black zone present in context |
|
| 29 | Cystidioles present |
|
| – | Cystidioles absent |
|
| 30 | Pores > 9 per mm; basidia > 13 μm long |
|
| – | Pores < 9 per mm; basidia < 13 μm long |
|
| 31 | Pores > 7 per mm |
|
| – | Pores < 7 per mm |
|
| 32 | Basidia > 19 μm long |
|
| – | Basidia < 19 μm long |
|
| 33 | Basidiospores ellipsoid |
|
| – | Basidiospores cylindrical |
|
| 34 | Basidiomata perennial |
|
| – | Basidiomata annual |
|
| 35 | Pores > 9 per mm |
|
| – | Pores < 9 per mm |
|
| 36 | Pores > 6 per mm |
|
| – | Pores < 6 per mm |
|
| 37 | Basidiospores 3–3.5 μm long |
|
| – | Basidiospores 3.4–4.1 μm long |
|
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases · Fungal Biology and Applications
Introduction
China harbours high diversity of wood-inhabiting fungi, with more than 1700 species documented to date (Dai 2010, 2012; Cui et al. 2019; Wu et al. 2022a, b; Liu et al. 2023; Yuan et al. 2023), the majority of which are distributed in south-western China (Wu et al. 2022b).
Dentipellicula Y.C. Dai & L.W. Zhou, a genus of Hericiaceae Donk, Russulales Kreisel ex P.M. Kirk, P.F. Cannon & J.C. David, is a kind of hydnoid fungi. It was established by Zhou and Dai (2013) and typified by Dentipellicula taiwaniana (Sheng H. Wu) Y.C. Dai & L.W. Zhou. Dentipellicula is characterised by hydnoid, resupinate basidiocarps with soft spines, a monomitic hyphal system with clamp connections and cyanophilous hyphae; and rough amyloid basidiospores (Ginns 1986; Zhou and Dai 2013; Chen et al. 2015; Zhou et al. 2021).
Dentipellicula was originally a member of Dentipellis. Zhou and Dai (2013) conducted a study of wood-inhabiting hydnoid species in Russulales and found that Dentipellis taiwaniana and D. leptodon (Mont.) Y.C. Dai & L.W. Zhou separated from Dentipellis, but phylogenetically clustered with Wrightoporia Pouzar. Morphologically, different from Dentipellis species, the generative hyphae of D. taiwaniana and D. leptodon showed a cyanophilic reaction. Meanwhile, the two species differ from Wrightoporia by their hydnoid hymenium and monomitic hyphal structure. Therefore, they argued that these two species did not belong to Dentipellis and Wrightoporia and established a new genus Dentipellicula. Subsequently, Chen et al. (2015) described Dentipellicula austroafricana Jia J. Chen, L.L. Shen & Y.C. Dai from South Africa, based on morphological characters and molecular data. Zhou et al. (2021) described Dentipellicula guyanensis Yuan Yuan, Meng Zhou, Jia J. Chen & Vlasák, based on morphological characters and ITS and nLSU sequences data from French Guiana. To date, the genus Dentipellicula comprises four recognised species, D. austroafricana, D. guyanensis, D. leptodon and D. taiwaniana.
The genus Phylloporia Murrill belongs to Hymenochaetaceae, Hymenochaetales, typified by P. parasitica Murrill, which was initially described from South America (Colombia) and grows on Bignonia sp. (Murrill 1904). Phylloporia species are characterised by annual or perennial, soft corky to hard corky basidiomata, tomentose to velutinate pileal surface, generative hyphae with simple septa and subglobose, ellipsoid or cylindrical, hyaline to yellowish, fairly thick-walled basidiospores. Phylloporia species are widely distributed in the Tropics and mostly parasitise living angiosperm trees, causing white rot (Wu et al. 2022b; Zhou et al. 2022).
With the development of molecular systematics, a series of phylogenetic studies on Phylloporia have emerged in recent years. Wagner and Ryvarden (2002) conducted the first monographic revision of this genus, incorporating phylogenetic data and ultimately confirmed 12 species within the genus. Olou et al. (2021) described a new species and a new record species from Benin, based on morphological characters and molecular data. Zhou et al. (2022) described two new species from the Neotropics, based on 28S ribosomal RNA phylogeny, morphology, host and geographic distribution. Subsequently, Olou et al. (2023) discovered and described a new species Phylloporia in Benin. Jerusalem et al. (2025) conducted a comprehensive multigene phylogeny of Phylloporia and described six new species from tropical Africa. To date, approximately 84 species have been recognised in Phylloporia worldwide, with 37 species reported from China (Wu et al. 2019a, 2020; Chamorro-Martínez et al. 2022; Zhou et al. 2022; Castro Hernández et al. 2023; Olou et al. 2023; Jerusalem et al. 2025; Shao et al. 2025).
Wood-inhabiting fungi are a morphologically, phylogenetically and ecologically diverse group, playing an integral role in wood degradation and the matter cycle within the ecological system and recognised as pivotal contributors to the intricate balance of forest ecosystems, these fungi being renowned as “key players” due to their enzymatic prowess, effectively breaking down woody components like lignin, cellulose and hemicellulose (Larsson et al. 2006; Dai et al. 2015; Dong et al. 2024, 2025; Yang et al. 2025). During our investigations of macrofungi from Hainan Province, three specimens of Dentipellicula and two of Phylloporia, exhibiting distinct morphological characteristics, were collected. The morphological observations and phylogenetic analyses, based on ITS + nLSU combined matrices were conducted to confirm the affinity of the undescribed species corresponding to Dentipellicula and Phylloporia. Accordingly, two new species are formally described and illustrated in the present study.
Materials and methods
Morphological studies
Specimen collection and preservation followed the methods described by Song et al. (2025). The specimens used in this study were gathered during the annual growing season of macrofungi (June to October). The specimens were dried on-site using a portable oven, to prevent putrefaction and contamination; the drying temperature was controlled at 40 °C–45 °C and the process was continued until the specimens were completely desiccated. Macromorphological descriptions were based on field notes and herbarium specimens. Detailed specimen information, such as host trees, ecological habits, geographic coordinates, location, altitude, collector and date, was recorded (Rathnayaka et al. 2025). Meanwhile, photos of the fruiting bodies and growth environment were taken by iPhone 15 Pro Max. All samples examined in this study were deposited in the Fungarium of the College of Life Sciences, Liaocheng University (LCUF). Micro-morphological data were obtained from dried specimens and observed under an Olympus BX53 compound microscope following the methods of Song et al. (2022). Basidiospores were measured from sections cut from the spines. The following abbreviations were used: IKI = Melzer’s reagent, IKI– = negative in Melzer’s reagent, KOH = 5% potassium hydroxide, CB = Cotton Blue, CB+ = cyanophilous, CB− = acyanophilous, L = mean spore length (arithmetic average of all spores), W = mean spore width (arithmetic average of all spores), Q = variation in the L/W ratios between the specimens studied and n = number of spores measured from a given number of specimens. All new fungal taxa described in this study have been formally registered in MycoBank (https://www.mycobank.org/) and the corresponding MycoBank registration numbers are provided in the taxonomic treatment section of the main text.
Molecular study
The CTAB plant genome rapid extraction Hi-DNA-secure Plant Kit (Tiangen, Beijing, China) was used to extract total genomic DNA from dried herbarium specimens. The extracted DNA was used to perform the polymerase chain reaction (PCR) according to the manufacturer’s instructions with some modifications (Li et al. 2023; Song et al. 2025). The primer pairs, ITS5/ITS4 and LR0R/LR7, were used to amplify ITS and nLSU sequences (Song et al. 2022, 2023). The PCR process for ITS was as follows: initial denaturation at 95 °C for 3 min, followed by 35 cycles at 94 °C for 40 s, 56 °C for 45 s and 72 °C for 1 min and a final extension of 72 °C for 10 min. The PCR process for nLSU was as follows: initial denaturation at 94 °C for 1 min, followed by 35 cycles at 94 °C for 30 s, 50 °C for 1 min and 72 °C for 1.5 min and a final extension of 72 °C for 10 min. The PCR products were purified and sequenced at Beijing Genomics Institute, China, with the same primers. All newly-generated sequences were submitted to GenBank (Table 1). Other sequences in the dataset for phylogenetic analysis were downloaded from GenBank.
Phylogenetic analyses
The phylogenetic relationships of Dentipellicula and Phylloporia were inferred, based on the combined ITS + nLSU sequence datasets. The datasets were aligned in MAFFT 7 (Katoh and Standley 2013) and manually adjusted in BioEdit (Hall 1999). The sequences of Bondarzewia podocarpi Y.C. Dai & B.K. Cui and B. occidentalis Jia J. Chen, B.K. Cui & Y.C. Dai were used as the outgroups for Dentipellicula, according to Zhou and Dai (2013). The sequences of Inonotus andersonii (Ellis & Everh.) Nikol. and I. hispidus (Bull.) P. Karst. were used as the outgroups for Phylloporia, according to Olou et al. (2023). Alignments were concatenated in Mesquite v. 3.2. The best-fit model of nucleotide evolution for the datasets was selected with AIC (Akaike Information Criterion) using MrModelTest 2.3 (Posada and Crandall 1998).
Maximum Likelihood (ML) analysis was performed in RAxML v.7.2.8 with a GTR + G + I model (Stamatakis 2006). All model parameters were estimated by the programme, but only the best Maximum-Likelihood tree from all searches was kept. MrModelTest 2.3 (Posada and Crandall 1998; Nylander 2004) was used to determine the best-fit evolution model for each dataset for Bayesian Inference (BI).
BI was performed using MrBayes 3.2.6 with two independent runs, each one beginning from random trees with four simultaneous independent chains, performing 5 million generations, sampling every 100 generations (Ronquist and Huelsenbeck 2003). The first 25% of the sampled trees were discarded as burn-in and a majority rule consensus tree of all remaining trees was calculated.
Branches that received bootstrap support for Maximum Likelihood (ML) and Bayesian posterior probabilities (BPP) greater than or equal to 50% (ML) and 0.95 (BPP) were regarded as prominently supported. Phylogenetic trees were visualised using FigTree v.1.4.2 (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 18 November 2025).
Results
BLAST search results
BLAST searches of the ITS sequences for all specimens (Table 2) showed that MHN240385, MHN240401 and MHN250010 matched Wrightoporiopsis amylohypha with query cover of 95%, 95% and 85% and percentage identity of 83.31%, 83.31% and 90.68%, respectively. Specimens MHN240359 and MHN240360 had the top hits to Phylloporia sp., with 100%/99% query cover and 87.71%/87.08% percentage identity. These results provided preliminary molecular evidence for the taxonomic assignment of the specimens, supporting subsequent phylogenetic analysis.
Phylogenetic analyses
The combined ITS + nLSU dataset of Hericiaceae included sequences from 23 fungal strains representing 13 taxa. The best-fit model for each partition was TN+F+G4 for ITS and TN+F+I+G4 for nLSU. Bayesian analysis yielded a topology similar to that from the ML analysis. The Bayesian analysis yielded a concordant topology with an average standard deviation of split frequencies of 0.009999. Only the ML tree is provided in Fig. 1 and the ML (50%) and BI (0.95) are shown at the nodes.
Maximum Likelihood tree illustrating the phylogeny of Dentipellicula, based on the combined sequence dataset of ITS + nLSU. Branches are labelled with Maximum Likelihood bootstrap higher than 50% and Bayesian posterior probabilities more than 0.95, respectively. Bold names = new species. Black stars: represent type sequences. Filled circles: represent type species.
The combined ITS + nLSU dataset of Phylloporia included sequences from 131 fungal strains representing 85 taxa. The best models for each region of the combined ITS + nLSU sequence dataset, as estimated and applied in the Bayesian analysis, were both GTR + I + G models. Bayesian analysis yielded a topology similar to that from the ML analysis. The Bayesian analysis resulted in a concordant topology with an average standard deviation of split frequencies = 0.009999. Only the ML tree is provided in Fig. 2 and the ML (50%) and BI (0.95) are shown at the nodes.
Maximum Likelihood tree illustrating the phylogeny of Phylloporia, based on the combined sequence dataset of ITS+nLSU. Branches are labelled with Maximum Likelihood bootstrap higher than 50% and Bayesian posterior probabilities more than 0.95, respectively. Bold names = new species. Black stars: represent type sequences. Filled circles: represent type species.
Phylogenetic analysis, based on the combined ITS + nLSU dataset (Fig. 1), provided more robust and accurate taxonomic placement of the specimens compared to preliminary BLAST searches. While BLAST results initially matched specimens MHN240385, MHN240401 and MHN250010 to Wrightoporiopsis, the phylogenetic tree clearly resolved the three specimens as a distinct, well-supported lineage (100%ML/1.00BI) representing the new species Dentipellicula hainanensis, which clustered within Dentipellicula rather than Wrightoporiopsis.
For specimens MHN240359 and MHN240360, BLAST searches targeting their ITS sequences initially identified Phylloporia sp. as the top homologous matches. This initial taxonomic assignment was robustly supported by our phylogenetic analysis of the combined ITS + nLSU dataset (Fig. 2), which confidently positioned both specimens within Phylloporia. Notably, the two specimens clustered together in a well-supported, monophyletic clade (100% ML/0.99 BI), indicating that they merit recognition as a new species in Phylloporia.
Taxonomy
Dentipellicula
hainanensis
Taxon classificationFungiRussulalesHericiaceae
C.G. Song & Z.F. Jia sp. nov.
E954190B-1775-597C-857C-9F2A85F5EA77
MycoBank No: 861880
Diagnosis.
Differs from other Dentipellicula species by its resupinate, soft corky and tomentose, basidiomata, uneven pileal surface and the presence of clamp connections in generative hyphae.
Basidiocarps of Dentipellicula hainanensis. Scale bars: 2 cm (a–d); 0.3 cm (e).
Microscopic structures of Dentipellicula hainanensis (drawn from the holotype). a. Basidiospores; b. Basidia and basidioles; c. Gloeocystidia; d. Hyphae from context; e. Hyphae from spines.
Type.
China • Hainan Province, Yinggeling National Nature Reserve, on fallen wood, alt. 670 m, 19°02'24"N, 109°34'12"E, 31 December 2024, MHN240401.
Etymology.
Hainanensis (Lat.) refers to the holotype locality of the species in Hainan Province.
Fruiting body.
Basidiomata annual, resupinate, without odour or taste. Pileus fan-shaped to nearly circular, up to 2.5 cm in diam., 0.3 cm thick at centre. Pileal surface greyish-orange (5B5) to light orange (6A5) to pinkish-buff, uneven, azonate, tomentose; margin darker than the middle, greyish-orange (6B5) buff, up to 0.35 cm wide. Fresh spines soft, light orange (6A5) to greyish-orange (6B5) when fresh, becoming fragile, upon drying, up to 2 mm long, 7–9 per mm across base. Subiculum soft corky, concolorous with pileal surface, up to 0.2 cm thick.
Microstructure.
Hyphal system monomitic; generative hyphae with clamp connections; IKI–, CB–; tissue unchanged in KOH. Generative hyphae in context hyaline, thick-walled, occasionally branched, interwoven to regularly arranged and 3–6 µm in diam. Generative hyphae in spines hyaline, thin- to thick-walled, occasionally branched, regularly arranged and 3–5 µm in diam. Gloeocystidia present, rooting deep from the trama, often encrusted, 8–11.6 µm in diam., cystidioles absent. Basidia clavate, 4-spored, 16–23 × 3.8–7.5 µm; basidioles similar to basidia in shape. Basidiospores, hyaline, ellipsoid, thick-walled, echinulate, IKI+, CB–, (2.8–)3–4.7(–5.2) × 3.2–4.2(–4.4) µm, L = 4 µm, W = 3.6 µm, Q = 1.03–1.32 (n = 90/3).
Additional specimens (paratypes) examined.
• Hainan Province, Changjiang County, Bawangling, Yajia Scenic Area, on a branch, alt. 630 m, 19°05'06"N, 109°07'22.8"E, 30 December 2024, MHN240385; Yinggeling National Nature Reserve, on fallen wood, alt. 670 m, 19°03'10.8"N, 109°33'50.4"E, 1 January 2025, MHN250010.
Phylloporia
rigida
Taxon classificationFungiHymenochaetalesHymenochaetaceae
C.G. Song & Z.F. Jia sp. nov.
6478CB6D-50D3-53DA-9B32-7E5EB664CCF5
MycoBank No: 861881
Diagnosis.
Differs from other Phylloporia species by its pileate, sessile basidiomata, fawn to reddish-brown and fan-shaped pileal surface, salmon to peach pileal margin, cream to buff yellow tubes and monomitic hyphal system and ellipsoid, thick-walled basidiospores.
Basidiocarps of Phylloporia rigida. Scale bars: 2 cm (a–d); 0.3 cm (e).
Microscopic structures of Phylloporia rigida (drawn from the holotype). a. Basidiospores; b. Basidia and basidioles; c. Cystidioles; d. Hyphae from context; e. Hyphae from spines.
Type.
China • Hainan Province, Changjiang County, Bawangling Yajia Scenic Area, on the living tree of Garcinia oblongifolia Champ. ex Benth., alt. 495 m, 19°05'06"N, 109°07'22.8"E, 30 December 2024, MHN240360.
Etymology.
Rigida (Lat.) refers to the rigid basidiomata.
Fruiting body.
Basidiomata annual, epixylous, pileate, sessile, without odour or taste. Pileus applanate to nearly half round, up to 4 cm in diam., 2 cm thick at centre. Pileal surface greyish-orange (6B5), uneven, azonate, fibrous to tomentose; margin greyish-orange (6B4), greyish-red (7B6) to dull red (8B3), wave, up to 1.2 cm wide. Tube light orange (6A4) when fresh, tough upon drying, up to 1 mm long, 5–7 per mm. Context, rigid, duplex, with a lower corky context separated from an upper trichoderm by a thin black line, up to 0.4 cm thick.
Microstructure.
Hyphal system monomitic; generative hyphae with simple septa; IKI–, CB–; tissue darker in KOH. Generative hyphae in context yellowish-brown, slightly thick-walled, occasionally branched, interwoven to regularly arranged and 3–8 µm in diam. Generative hyphae in tubes yellowish-brown, thick-walled, often branched, regularly arranged and 2–6 µm in diam. Gloeocystidia absent. Cystidioles present, 10.5–13.5 × 1–3.5 µm in diam. Basidia clavate, 4-spored, 9–13 × 2.5–5 µm; basidioles similar to basidia in shape. Basidiospores ellipsoid, oblong-ellipsoid to cylindrical, yellow-brown, ellipsoid, thick-walled, IKI–, CB–, 2.3–4.5 × 2.2–3.8 µm, L = 3.31 µm, W = 2.83 µm, Q = 1–1.32 (n = 60/2).
Additional specimen (paratype) examined.
China • Hainan Province, Changjiang County, Bawangling Yajia Scenic Area, on a living tree of Garcinia oblongifolia, alt. 495 m, 19°05'06"N, 109°07'22.8"E, 30 December 2024, MHN240359.
Discussion
In the present study, phylogenetic analysis of the combined ITS and nLSU sequences revealed that Dentipellicula hainanensis is distantly related to Dentipellis, Hericium and Laxitextum, but clusters within the Dentipellicula clade (Fig. 1).
Dentipellicula hainanensis formed a single lineage, different from other species of Dentipellicula in our phylogenetic analyses (Fig. 1). Morphologically, D. hainanensis might be confused with D. leptodon in having cottony pileus, pale yellow spines and the presence of clamp connections in generative hyphae (Ginns 1986). However, D. leptodon is distinguished by its white pileus margin, longer spines measuring 3–5 mm, longer basidia measuring 20–26 × 4–5 µm and narrower basidiospores measuring 3.7 × 2.8 µm (Ginns 1986). The prevalence of newly-described species in subtropical and tropical areas indicates a remarkable richness of Phylloporia in these regions (Anonymous 1997; Decock et al. 2015; Olou et al. 2021, 2023; Wu et al. 2022b; Zhou et al. 2022; Jerusalem et al. 2025). In the present study, one new species of Phylloporia is described from Hainan Province, based on morphological characters and phylogenetic analyses of the combined ITS + nLSU sequences. The main morphological characteristics of each Phylloporia species from China were summarised in Table 3.
Phylloporia rigida grouped with Phylloporia flabelliformis Decock & Yombiy. and Phylloporia gabonensis Decock & Yombiy. in our phylogenetic analyses (Fig. 2). Morphologically, P. flabelliformis and P. gabonensis are similar to P. rigida in having pileate, sessile basidiomata and cork-coloured pileal surface and similar basidia. However, P. flabelliformis is distinguished by its shiny, smooth pileal surface, thin context, white pileal margin and smaller basidiospores, measuring (3–)3.3–4 × (2.2–)2.5–2.8(–3) µm (Decock et al. 2015). Phylloporia gabonensis is distinguished by its dull, smooth pileal surface, thin, irregular, pale yellow to pale greyish-orange pileal margin and slightly smaller basidiospores, measuring 3.7–4.2(–5) × 2.7–3(–3.3) µm (Decock et al. 2015).
For ecological traits, Dentipellicula species predominantly grow on angiosperms and are frequently found on fallen trunks and dead trees (Chen et al. 2015; Wang 2018; Zhou et al. 2021). However, the genus Phylloporia displays marginally distinct ecological traits. Most Phylloporia species are parasitic on living hardwood hosts; however, a few species grow on deadwood and speciation in this genus is likely promoted by the colonisation of new hosts and the adaptive evolution that follows (Wu et al. 2019a). High levels of host specificity are displayed by the majority of Phylloporia species that parasitise living hardwood hosts (Table 2) (Wagner and Ryvarden 2002). It indicated that species in Phylloporia are host-biased, which can be used as an auxiliary basis for species discovery and identification.
Key to accepted species of Dentipellicula
**: **
Key to species of Phylloporia from China
**: **
Supplementary Material
XML Treatment for Dentipellicula hainanensis
XML Treatment for Phylloporia rigida
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bittencourt F, Stürmer SL, Reck MA, Drechsler-Santos ER (2018) Phylloporia minuta sp. nov. (Basidiomycota, Hymenochaetales): a remarkable species discovered in a small protected urban area of Atlantic Forest. Phytotaxa 348: 199–210. 10.11646/phytotaxa.348.3.3 · doi ↗
- 2Castro Hernández L, Camino Vilaró M, Herrera Figueroa S (2023) Revisión taxonómica del género Phylloporia (Hymenochaetaceae, Basidiomycota) en Cuba. Acta Botánica Mexicana 130: e 2149. 10.21829/abm 130.2023.2149 · doi ↗
- 3Chamorro-Martínez HA, Raymundo T, Martínez González CR, Acosta EA, Valenzuela R (2022) Two new stipitate species of Phylloporia (Basidiomycota, Hymenochaetaceae) from Chamela Biology Station, U.N.A.M. in Jalisco, Mexico. Lilloa 59: 359–375. 10.30550/j.lil/2022.59.S/2022.09.28 · doi ↗
- 4Chen JJ, Shen LL, Dai YC (2015) Dentipellicula austroafricana sp. nov. supported by morphological and phylogenetic analyses. Mycotaxon 130: 17–25. 10.5248/130.17 · doi ↗
- 5Chen YY, Zhu L, Xing JH, Cui BK (2017) Three new species of Phylloporia (Hymenochaetales) with dimitic hyphal systems from tropical China. Mycologia 109: 951–964. 10.1080/00275514.2017.141069229474112 · doi ↗ · pubmed ↗
- 6Cui BK, Yuan HS, Dai YC (2010) Two new species of Phylloporia (Basidiomycota, Hymenochaetaceae) from China. Mycotaxon 113: 171–178. 10.5248/113.171 · doi ↗
- 7Cui BK, Li HJ, Ji X, Zhou JL, Song J, Si J, Yang ZL, Dai YC (2019) Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China. Fungal Diversity 97(1): 137–392. 10.1007/s 13225-019-00427-4 · doi ↗
- 8Dai YC (2010) Hymenochaetaceae (Basidiomycota) in China. Fungal Diversity 45(1): 131–343. 10.1007/s 13225-010-0066-9 · doi ↗