CO2-driven biosurfactant synthesis by bacteria within CCUS
Amanda Pasinato Napp, William Lautert Dutra, Lovaine Silva Duarte, Eduarda Vargas Abati, Francine Melise dos Santos, Clarissa Lovato Melo

TL;DR
This paper explores how bacteria can capture CO2 and produce biosurfactants, offering a sustainable way to reduce greenhouse gases and create useful products.
Contribution
The paper introduces the integration of CO2 capture with biosurfactant synthesis using microbial systems and synthetic biology.
Findings
Microbial CO2 capture can be coupled with biosurfactant production for sustainable biomanufacturing.
Anaerobic and CO2-enriched systems offer new routes for biosurfactant synthesis.
Synthetic biology can link carbon fixation to biosurfactant biosynthesis pathways.
Abstract
Microbial CO2 capture coupled with biosurfactant production represents a promising strategy for greenhouse gas mitigation and sustainable biomanufacturing. This review examines the metabolic and engineering aspects of microbial carbon capture, focusing on both anaerobic and CO2-enriched systems within the Microbial-CCUS framework. The structural diversity, physicochemical properties, and industrial applications of microbial biosurfactants are discussed, along with emerging evidence of anaerobic biosurfactant synthesis linked to CO2 metabolism. Advances in genetic and synthetic biology, pathway modularization, and systems-level modeling are reshaping the potential to coordinate CO2 fixation with biosurfactant biosynthesis. Integrating artificial intelligence with metabolic engineering may further optimize productivity, scalability, and energy efficiency. Despite technical and economic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
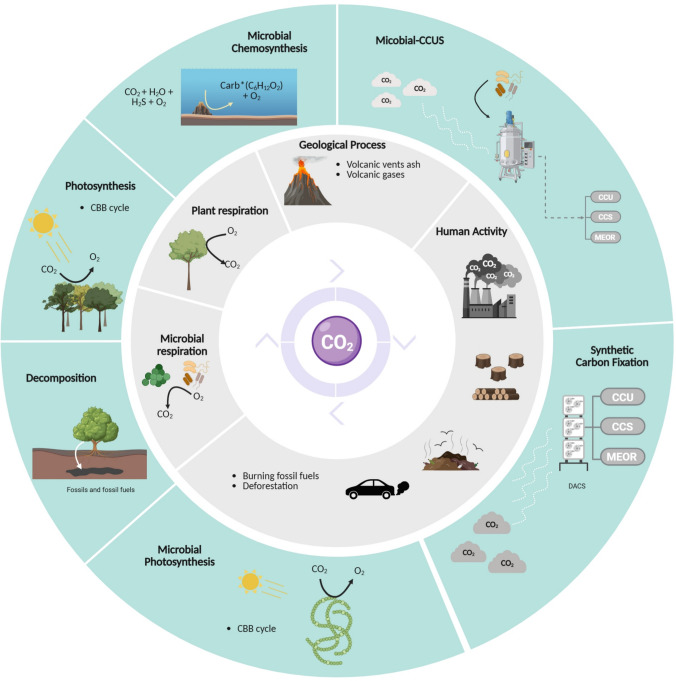 Figure 1
Figure 1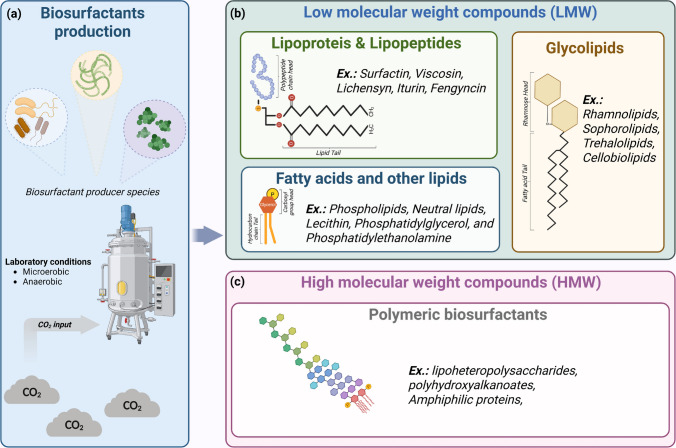 Figure 2
Figure 2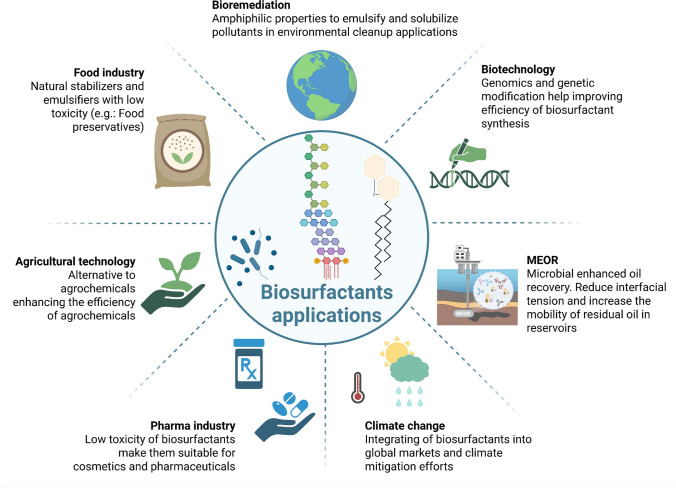 Figure 3
Figure 3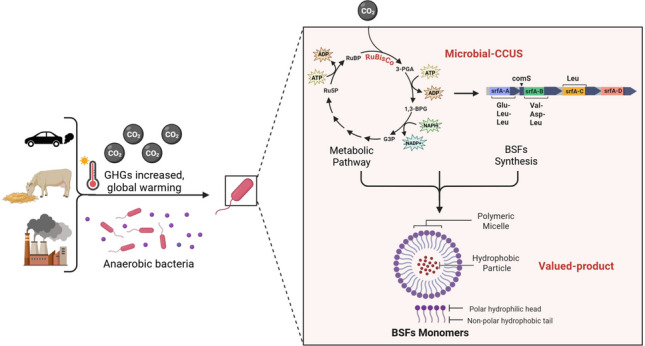 Figure 4
Figure 4- —Petronas Petróleo Brasil Ltda
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial bioremediation and biosurfactants · CO2 Sequestration and Geologic Interactions · Microbial Fuel Cells and Bioremediation
1. Introduction
Large-scale anthropogenic emissions of greenhouse gases (GHGs), initiated during the Industrial Revolution, have significantly disrupted Earth’s climate system (Kammerer et al. 2023). Key GHGs, including carbon dioxide (CO_2_), methane (CH_4_), nitrous oxide (N_2_O), hydrofluorocarbons (HFCs), perfluorocarbons (PFCs), and sulfur hexafluoride (SF_6_), absorb and re-emit infrared radiation, enhancing the greenhouse effect and contributing to global warming (Al‐Ghussain 2019; Hanifa et al. 2023). The CO_2_ remains the dominant contributor, rising from historically stable levels below 250 to 420 ppm in 2024, now representing about 76% of atmospheric GHGs (Ritchie et al. 2023). This unprecedented rise is linked to ocean acidification, biodiversity loss, altered precipitation patterns, and threats to food security (Rising et al. 2022).
The Intergovernmental Panel on Climate Change (IPCC) reports a 1.1 °C rise in global temperatures since pre-industrial times, with a projected 0.2 °C increase per decade (Calvin et al. 2023). Moreover, current actions are insufficient to meet the Paris Agreement strategies (Tollefson 2021). The International Energy Agency emphasizes that net-negative emissions are unlikely without large-scale Carbon Capture, Utilization, and Storage (CCUS) deployment (IEA 2021). In this context, conventional CCUS, including amine washing and mineral carbonation, faces economic and scalability constraints, as well as secondary environmental impacts (Hanifa et al. 2023; Thiedemann & Wark 2025).
In contrast, biological CO_2_ sequestration offers sustainable and low-energy alternatives using microorganisms as natural catalysts for carbon fixation (Bardi et al. 2025; Ruan et al. 2025). Microbes employ metabolic pathways such as the Calvin–Benson–Bassham cycle, the Wood–Ljungdahl pathway, and the 3-hydroxypropionate cycle (Duarte et al. 2017; Mistry et al. 2019). These systems not only fix CO_2_ but can generate high-value biomaterials, particularly biosurfactants (Kumar et al. 2018; Sundaram and Thakur 2015). These amphiphilic molecules have broad industrial applications in remediation, cosmetics, agriculture, and pharmaceuticals (Eras-Muñoz et al. 2022; Kugaji et al. 2025; Sarubbo et al. 2022). Their production through microbial CO_2_ assimilation provides a dual advantage, reducing GHG emissions and generating sustainable biomolecules with high added value (Rajkumar et al. 2022). In addition to their role in carbon capture, the contribution of CO_2_-derived biosurfactants to the transition toward greener industries must be demonstrated using quantitative sustainability metrics. Future studies should therefore evaluate microbial-CCUS platforms through indicators such as net CO_2_ balance (kg CO_2_-eq captured or avoided per kg product), energy demand and aeration intensity, carbon utilization efficiency, and life-cycle GHG emissions in direct comparison with petrochemical surfactants and conventional CCUS technologies. Additional parameters, including water and nutrient intensity, biodegradability, ecotoxicity, and biosurfactant performance indicators, will be essential to demonstrate whether microbial-CCUS systems provide genuine environmental advantages, rather than simply transferring impacts between process steps. Integrating its development into sustainability certifications and circular economy frameworks will not only strengthen economic viability but also accelerate the widespread adoption of microbial-CCUS technologies (Kugaji et al. 2025; Sharma et al. 2025).
This review examines anaerobic microorganisms as key agents in CO_2_ fixation and biosurfactant production, highlighting their metabolic pathways, microbial diversity, and recent industrial applications. We further synthesize advances in genetic and metabolic engineering, as well as emerging AI-assisted approaches to optimize yields. By examining technical, economic, and environmental challenges along with scalability and regulatory frameworks, we position anaerobic microbial systems as next-generation platforms that integrate carbon management with sustainable biotechnology.
2. CO2 capture technologies
The global carbon cycle includes interconnected natural and anthropogenic processes that regulate CO_2_ exchanges among the atmosphere, biosphere, and lithosphere (Fig. 1). Natural pathways such as photosynthesis and chemosynthesis operate as biological sinks. In contrast, plant and microbial respiration, decomposition, and geological activity (e.g., volcanic emissions) constitute major sources of atmospheric CO_2_. These biogeochemical mechanisms maintain atmospheric CO_2_ balance but are increasingly disrupted by anthropogenic emissions from fossil fuel combustion, industrial processes, and deforestation (Ángeles et al. 2021).Fig. 1Global cycle of CO_2_ production and fixation. The figure illustrates the dual role of CO_2_ as a central chemical compound in metabolic, industrial, and environmental processes. The inner sections depict CO_2_-releasing processes, including anthropogenic activities such as fossil fuel combustion, industrial operations, and deforestation, which are the primary drivers of climate change. Geological processes, including volcanic activity and tectonic emissions, also contribute to atmospheric CO_2_. Plant respiration and microbial respiration release CO_2_ as by-products of aerobic metabolism and organic matter decomposition, respectively. The outer ring represents pathways for CO_2_ fixation. Natural mechanisms include photosynthesis and microbial chemosynthesis, while artificial approaches involve synthetic and synthetic biological carbon fixation technologies, including CCU, CCS, and MEOR. These engineered approaches are increasingly critical for advancing carbon sequestration and mitigating climate change
In response to rising emissions, several CO_2_ capture and storage (CCS) technologies based on engineered physical and chemical principles have been developed to mitigate point-source pollution (Yousaf et al. 2022). CCS systems isolate CO_2_ from flue gases or process streams through post-combustion, pre-combustion, or oxyfuel methods, followed by compression and injection into deep geological formations such as saline aquifers or depleted reservoirs (Karimi et al. 2023).
An evolution of this principle, Direct Air Carbon Capture and Storage (DACCS), removes CO_2_ directly from ambient air using solid sorbents or liquid solvents, offering potential for diffuse and hard-to-abate emissions (Brazzola et al. 2025; Cobo et al. 2022). Similarly, Bioenergy with Carbon Capture and Storage (BECCS) integrates biomass-based energy generation with CO_2_ capture and storage, simultaneously producing renewable energy and achieving net negative emissions (Ataeian et al. 2019; Oh et al. 2025).
Beyond storage-based approaches, Carbon Capture and Utilization (CCU) and Carbon Capture, Utilization, and Storage (CCUS) expand carbon management into industrial and circular pathways. CCU technologies convert captured CO_2_ into fuels, polymers, chemicals, or construction materials, displacing fossil-derived feedstocks and promoting a circular carbon economy. CCUS combines utilization and long-term storage, linking industrial valorization with permanent sequestration and creating an economic bridge toward large-scale mitigation (Cuéllar-Franca & Azapagic 2015; Khandelwal et al. 2021; Rajput & Keshavkant 2025; Ruan et al. 2025). A concise summary of the main carbon management pathways discussed in this section is provided in Box 1.
In parallel, Microbial-CCUS represents an emerging biological strategy that complements engineered systems. Through autotrophic pathways such as the CBB and Wood–Ljungdahl cycles, microorganisms can fix CO_2_ into valuable biomolecules including biosurfactants, bioplastics, and biofuels. This biological capture approach is characterized by low energy demand, scalability, and compatibility with waste gas streams, aligning biotechnological innovation with sustainable carbon management.
Despite their potential, the widespread deployment of CCS, DACCS, BECCS, and CCUS faces economic, technical, and environmental challenges. High energy and water demand, solvent degradation, corrosion, leakage risks, and material limitations remain key obstacles (Ángeles et al., 2021a; Oh et al. 2025). Overcoming these barriers requires continued research to develop cost-effective, durable, and scalable systems. The integration of engineered technologies with natural carbon sinks and biological approaches is essential for advancing sustainable and resilient carbon management. Box 1. Glossary of carbon management pathways****CCS: Capture of CO_2_ from large point sources (e.g., power plants, industries), followed by permanent storage in geological reservoirs (saline aquifers, depleted oil and gas fields).DACCS: Direct removal of atmospheric CO_2_ using chemical sorbents or solvents, coupled with geological sequestration.BECCS: Combines biomass-based energy production with CO2 capture and storage, achieving simultaneous low-carbon energy generation and removal of biogenic CO2 from the atmosphere.CCUS: Integration of CO2 utilization and permanent storage, bridging economic and mitigation goals.CCU: Conversion of captured CO2 into industrial products (fuels, chemicals, or materials), promoting circular carbon use.Microbial-CCUS: Biological fixation or assimilation of CO2 by microorganisms (e.g., CBB, Wood–Ljungdahl cycles), producing high-value biochemicals.
3. Biological CO2 fixation
Microbial CO_2_ fixation represents a frontier in carbon capture research, providing solutions to the limitations of conventional CCS and chemical methods. Microorganisms employ diverse biochemical pathways and enzymatic strategies to convert CO_2_ into biomass or value-added compounds, often with greater sustainability and adaptability (Choi et al. 2022; Correa et al. 2025; de Oliveira Maciel et al. 2022).
These strategies are broadly classified into photosynthetic and non-photosynthetic systems (Table 1). Photoautotrophs, including algae and cyanobacteria, rely on light-driven energy conversion, whereas chemoautotrophs and anaerobic archaea exploit chemical energy from inorganic substrates such as hydrogen, sulfur compounds, or ammonia. Although heterotrophs are not primary CO_2_ fixers, some exhibit ancillary pathways that contribute marginally to carbon assimilation (Gong et al. 2018; Maheshwari et al. 2022). Together, these organisms demonstrate the ecological and metabolic diversity of microbial carbon fixation and its biotechnological potential. Table 1. Summary of key microbial CO_2_ fixation pathwaysPathwayMicroorganismsSubstrateKey EnzymesProductsKey ReactionsEnvironmentReferencesCBB cyclePlants, microalgae, cyanobacteria, chemolithotrophic bacteria3 CO_2 9 ATP 6 NADPHRuBisCOG3PCO_2 + RuBP → 3-PGA3-PGA + ATP → 1,3-BPG + ADP 1,3-BPG + NADPH → G3P + NADP + PiG3P + ATP → RuBP + ADPPhotosynthetic environments, terrestrial and aquatic(Bassham et al., 1954)rTCA cycleChlorobium limicola, Proteobacteria, acetogens, sulfur bacteria2 CO_2 2 ATP 4 NADPH2-oxoglutarate synthase, Isocitrate dehydrogenaseAcetyl-CoACO_2 + H_2_O → 2-oxoglutarateCitrate → acetyl-CoA + oxaloacetateDeep-sea vents, anoxic environments(Evans et al. 1966)Wood-LjungdahlAcetogens (Clostridium thermoaceticum), methanogens2 CO_2 1 ATP 4 NADPHFormate dehydrogenase, CODHAcetyl-CoACO_2 → formate → formyl-THF → methyl-THF + CO + CoA → acetyl-CoAAnaerobic environments(Ljungdhal, 1986)3HP bicycleChloroflexus aurantiacus, Chloroflexichemoautotrophic bacteria, archaea3 HCO_3_⁻ 5 ATP 5 NADPHAcetyl-CoA carboxylase, Propionyl-CoA carboxylasePyruvateAcetyl-CoA or Propionyl-CoA + HCO_3_⁻ + ATP → malonyl-CoA or Methylmalonyl-CoA +ADP + Pi → Beta-Methylmalyl-CoA → Pyruvate + Other IntermediatesAerobic conditions, nutrient-limited habitats(Strauss and Fuchs, 1993)3HP/4HB cycleMetallosphaera sedula, Acidianus infernus2 HCO_3_⁻ 4 ATP 4 NADPHAcetyl-CoA-Propionyl-CoA carboxylaseAcetyl-CoACO_2_ + Acetyl-CoA + ATP → malonyl-CoA + ADP + Pi → 3HP + CoA → Propionyl-CoA + CO_2_ → 4HB → Acetyl-CoAAnaerobic, acidic, thermophilic environments(Berg et al., 2007)Di/4HB cycleIgnicoccus hospitalis, Thermoproteus neutrophilus1 CO_2 1 HCO_3⁻ 3 ATP 4 NADPHPyruvate synthase, PEP carboxylaseAcetyl-CoACO_2_ + Acetyl-CoA + ATP → malonyl-CoA + ADP + PiSuccinyl-CoA + H_2_ → 4-HB → Acetyl-CoA + H_2_O/Pyruvate + CO_2_ + ATP → Oxaloacetate + ADP + PiGeothermal, hydrothermal vents(Huber et al., 2008)
3.1 Photosynthetic CO2 fixation
Photosynthesis remains the dominant natural mechanism for CO_2_ assimilation. In photoautotrophic organisms (plants, algae, cyanobacteria), light-dependent reactions generate ATP and NADPH, which power the Calvin–Benson–Bassham (CBB) cycle (Table 1) (Bassham et al. 1954; Karishma et al. 2024). The ribulose-1,5-bifosfato carboxilase/oxygenase (RuBisCO) catalyzes CO_2_ fixation into ribulose-1,5-bisphosphate, producing 3-phosphoglycerate and eventually carbohydrates (Kajla et al. 2022; Li et al. 2024; Liu et al. 2024).
The main photoautotrophic microbial chassis explored for biotechnological CO₂ capture include cyanobacteria such as Synechocystis sp. PCC 6803, Synechococcus elongatus PCC 7942, and Nostoc spp., as well as microalgae such as Chlorella, Botryococcus, and Arthrospira (Spirulina), which are widely used as model systems to investigate and rationally engineer photosynthetic carbon fixation pathways (Angermayr et al. 2009; Della Valle et al. 2024; Ighalo et al. 2022; Mills et al. 2020; Mohapatra et al. 2022; Rajkumar et al. 2022).
Microalgae such as Chlorella and Botryococcus employ Carbon Concentration Mechanisms (CCMs), actively transporting bicarbonate (HCO_3_⁻) and compartmentalizing it in pyrenoids to enhance RuBisCO efficiency (Duarte et al. 2017). Cyanobacteria use analogous carboxysomes. These adaptations improve carbon capture under limiting conditions (Ataeian et al. 2019; Correa et al. 2025; Duarte et al. 2017). Microalgae stand out when compared to terrestrial plants in CO_2_ sequestration due to their rapid growth rates, high photosynthetic efficiency, and ability to utilize concentrated CO_2_ sources such as flue gases. Unlike terrestrial crops, which generally exhibit lower carbon capture efficiency per unit of biomass, microalgae can fix up to 1.83 kg of CO_2_ per kg of dry biomass, making them particularly attractive biological platforms for CCUS-oriented bioprocesses (Razzak et al. 2024); Mohapatra et al. 2022).
In photosynthetic systems, most reported surface-active compounds correspond to extracellular polysaccharides (EPS) and glycolipid-rich fractions with emulsifying activity, rather than canonical biosurfactants such as rhamnolipids or lipopeptides typically produced by heterotrophic or anaerobic bacteria. These EPS-based materials exhibit surface and interfacial activity and have been proposed as functional biosurfactant analogues in CO_2_-driven phototrophic platforms (de Morais et al., 2019; Han et al. 2013; Laroche 2022; Zhou et al. 2024).
Genetic strategies to improve carbon fixation and product formation include carboxysome engineering, overexpression of RuBisCO and carbonic anhydrase, and optimization of CCM-related genes to enhance intracellular CO_2_ availability. At the technological level, advances include high-density photobioreactors, improved light distribution systems, and CO₂-enriched cultivation platforms aimed at increasing biomass productivity and carbon capture efficiency (Rajkumar et al. 2022).
However, quantitative data on biosurfactant titers under fully CO_2_-driven phototrophic conditions remain scarce. Reported productivities of surface-active EPS in cyanobacteria typically range from tens to a few hundreds of mg·L⁻^1^, which remain substantially lower than those achieved in anaerobic or heterotrophic biosurfactant systems (Laroche 2022).
Despite their ecological diversity, photosynthetic systems face challenges related to scalability, resource demand, and sensitivity to controlled operational parameters, such as light distribution, temperature, and CO_2_ concentration, under industrial cultivation conditions (Occhipinti et al. 2024). Compared to anaerobic CCUS platforms, photosynthetic systems are constrained by light availability, lower volumetric productivity, and limited biosurfactant diversity. In contrast, anaerobic chemoautotrophs and facultative anaerobes operate independently of light, reach higher cell densities in conventional bioreactors, and have demonstrated biosurfactant titers in the g·L⁻^1^ range.
These limitations create a clear rationale for exploring anaerobic microbial CO_2_ fixation and Microbial-CCUS (Box 1), which may overcome phototrophic bottlenecks while simultaneously enabling the production of high-value biomolecules. However, the absence of harmonized quantitative metrics across studies, particularly regarding biosurfactant titers, productivities, and growth-associated parameters under CO_2_-driven phototrophic conditions, currently prevents a robust and quantitative comparison with anaerobic CCUS platforms. This challenge reflects a broader methodological gap in the literature rather than a lack of technological potential, underscoring the need for standardized reporting frameworks in future studies.
3.2 Non-photosynthetic CO2 fixation
Non-photosynthetic microorganisms provide a complementary approach to CO_2_ fixation that is often associated with lower operational energy demand and simplified cultivation requirements, as these systems avoid the restrictions related to light distribution, photolimitation, and reactor geometry that typically restrict volumetric productivity in phototrophic platforms. In this context, energy efficiency can be assessed through indicators such as energy demand per mass of CO_2_ captured or converted, energy consumption per unit of biosurfactant produced, and life-cycle energy intensity. However, these metrics remain rarely standardized across the literature, since most studies emphasize metabolic capability or product titers while detailed evaluations of agitation, gas transfer, and downstream energy inputs are reported inconsistently.
These organisms exploit chemical energy from substrates such as hydrogen, reduced sulfur compounds, or ammonia, which makes them particularly relevant in anaerobic and extreme environments. Their metabolic versatility underpins several biochemical pathways that channel CO_2_ into central metabolites and value-added products, offering significant potential for carbon capture technologies (Gong et al. 2018). The major pathways are summarized in Table 1., while their functional significance is outlined below.
Reductive tricarboxylic acid (rTCA) cycle
First described in Chlorobium limicola, this reverse TCA cycle incorporates CO_2_ into intermediates such as acetyl-CoA and pyruvate via ATP-citrate lyase and related enzymes. The rTCA cycle is pivotal in light-limited environments like hydrothermal vents, exemplifying metabolic adaptation to extreme conditions (Evans et al. 1966; Kajla et al. 2022).
Wood–Ljungdahl (WL) pathway
Found in acetogens and methanogens such as Clostridium thermoaceticum, this pathway reduces CO_2_ to formate and carbon monoxide, which then combine with a methyl group to form acetyl-CoA. Its minimal ATP requirement makes it highly energy-efficient and suitable for industrial CO_2_ conversion into acetate, methane, or other precursors (Ljungdahl 1986).
3-Hydroxypropionate (3HP) bicycle
Discovered in Chloroflexus aurantiacus, this dual-cycle process incorporates CO_2_ into glyoxylate via intermediates such as malonyl-CoA and 3-hydroxypropionate, involving multiple enzymatic steps. It is particularly advantageous for microorganisms in nutrient-limited or extreme habitats (Kajla et al. 2022; Strauss and Fuchs 1993).
3-Hydroxypropionate/4-hydroxybutyrate (3HP/4HB) pathway
Identified in autotrophic archaea such as Metallosphaera sedula, this pathway carboxylates acetyl-CoA to malonyl-CoA and regenerates acetyl-CoA through intermediates including 4-hydroxybutyrate. It is highly effective under anaerobic, thermophilic, and acidic conditions (Berg et al. 2007; Choi et al. 2022).
Dicarboxylate/4-hydroxybutyrate (Di/4HB) cycle
Observed in extremophiles like Ignicoccus hospitalis, this cycle combines CO_2_ with acetyl-CoA to form pyruvate and oxaloacetate, which are subsequently recycled into succinyl-CoA and acetyl-CoA. Its energy-efficient design supports robust carbon fixation under resource-limited conditions (Gong et al. 2018; Huber et al. 2008).
Compared to photosynthetic organisms, non-photosynthetic microorganisms maintain the capacity to thrive in light-limited or extreme habitats while remaining fully compatible with conventional bioreactor infrastructures. Importantly, integrating phototrophic and chemotrophic strategies may provide complementary advantages, enabling resilient Microbial-CCUS frameworks that couple carbon assimilation with the production of value-added metabolites and expand opportunities for industrial biomanufacturing.
4. Microbial surfactants
Microorganisms capable of assimilating CO_2_ can channel fixed carbon into high-value biomolecules, offering a sustainable strategy for climate change mitigation. Among these products, biosurfactants stand out as amphiphilic metabolites capable of lowering surface and interfacial tension, solubilize hydrophobic substrates, and modulate cell–surface interactions. Their biodegradability, renewable origin, and eco-friendly properties make them attractive alternatives to synthetic surfactants (Kugaji et al. 2025; Markande et al. 2021). These molecules align with Microbial-CCUS concepts, where CO_2_ fixation is directly coupled to metabolite synthesis (Fig. 2a). Compared to synthetic surfactants, biosurfactants offer ecological advantages, including enhanced microbial motility, biofilm formation, competitive interactions, and resistance to toxic compounds (Gong et al. 2018; Hu et al. 2019).Fig. 2Biosurfactants by classes. Biosurfactants are naturally occurring compounds produced by microorganisms. a) Environmental and laboratory-cultured microbes serve as model systems for biosurfactant production under specific laboratory conditions, where CO_2_ can be utilized as a carbon source. CO_2_ can be utilized as a carbon source for generating biosurfactants. b) Microorganisms synthesize a variety of biosurfactant classes, typically composed of a hydrophilic “head” and a hydrophobic “tail,” with differences in their chemical composition. Based on their molecular weight and complexity, biosurfactants are categorized as low-molecular-weight (LMW) or c) high-molecular-weight (HMW)
4.1 Structural diversity and classification of biosurfactants
Biosurfactants are composed of a hydrophilic “head” (sugars, peptides, phosphates) and a hydrophobic “tail” (lipids or fatty acids). They are broadly classified into low-molecular-weight (LMW) and high-molecular-weight (HMW) compounds (Fig. 2b, c) (Qin et al. 2025). LMW biosurfactants (< 1500 Da) include glycolipids (e.g., rhamnolipids, sophorolipids), lipopeptides (surfactins, iturins, fengycins), fatty acids, and phospholipids. These molecules rapidly adsorb to interfaces, efficiently reducing water surface tension from 72 to ~ 27 mN·m^−1^ (Lakatos et al. 2022). Lipopeptides produced by Bacillus and Pseudomonas exhibit both surface-active and antimicrobial properties. HMW biosurfactants include polysaccharides, lipopolysaccharides, amphiphilic proteins, and polymeric complexes such as emulsan. These macromolecules act as potent emulsifiers, stabilizing biofilms and enhancing the bioavailability of hydrophobic substrates (Ibrahim et al. 2022).
The type and yield of biosurfactants obtained from bioprocesses are influenced by both the producing microorganism and the environmental conditions, including nutrient availability, oxygen levels, and salinity (Araújo et al., 2020; Moro et al. 2018; Napp et al. 2018). Understanding these parameters is essential for optimizing production and tailoring the physicochemical properties of biosurfactants to specific industrial applications.
4.2 Biosurfactant applications
The collective properties of biosurfactants make them sustainable alternatives to synthetic surfactants across multiple industries (Fig. 3). In environmental remediation, surfactins and glycolipids lower surface tension, promoting hydrocarbon solubilization and pollutant degradation, while polysaccharide-based compounds such as emulsan (Acinetobacter calcoaceticus) stabilize emulsions and increase the bioavailability of hydrophobic substrates, a key for oil recovery and bioremediation (Moro et al. 2018). In agriculture, lipopeptides such as iturins and fengycins act as antifungal agents and can trigger systemic resistance in plants, whereas phospholipids like lecithin and phosphatidylethanolamine serve as natural wetting and emulsifying agents, enhancing agrochemical efficiency while reducing synthetic inputs (Zhang et al. 2022). In the pharmaceutical and cosmetic sectors, surfactins support drug delivery and personal care formulations through strong surface activity and antimicrobial action, while phosphatidylglycerol enhances formulation stability under variable conditions, reinforcing the value of biosurfactants as safe multifunctional excipients (Sultan et al. 2025).Fig. 3Applications of biosurfactants. Multisectoral applications of microbial biosurfactants and their relevance in environmental and industrial contexts. Due to their amphiphilic nature, biosurfactants play a key role in bioremediation, facilitating the emulsification and solubilization of hydrophobic pollutants. In agriculture, they serve as sustainable alternatives to agrochemicals, enhancing efficacy and reducing toxicity. In the food industry, biosurfactants are applied as natural preservatives and stabilizers. Their low toxicity and biodegradability enable their use in the pharmaceutical and cosmetic sectors. In energy systems, biosurfactants contribute to MEOR by improving oil mobilization. Advances in genomics and metabolic engineering further support biotechnological innovations to enhance biosurfactant yield and performance. Integration of biosurfactants into global markets may also support climate mitigation strategies by replacing synthetic surfactants with bio-based alternatives
Despite this wide applicability, most biosurfactant production systems remain aerobic, and anaerobic pathways are largely unexplored. This knowledge gap reflects technical challenges related to the limited availability of oxygen-independent metabolic routes, low production yields, and constraints in energy and redox balance. Expanding biosurfactant synthesis to anaerobic or CO_2_-enriched systems could enhance process sustainability by reducing aeration costs, minimizing foaming, and enabling operation in closed bioreactors. Furthermore, coupling biosurfactant production with CO_2_ utilization offers the prospect of developing microbial platforms compatible with CCUS frameworks, where carbon capture and metabolite synthesis occur simultaneously, thereby broadening the scope of sustainable microbial biotechnology. The feasibility of anaerobic biosurfactant production has been demonstrated under specific environmental and physiological conditions.
4.3 Anaerobic biosurfactant production
The earliest evidence of biosurfactant synthesis under oxygen deprivation dates back to 1955, when Desulfovibrio desulfuricans was shown to produce surface-active compounds in the absence of oxygen (La Rivière 1955). In this pioneering study, cells were cultivated in a defined mineral medium containing sodium lactate as the sole carbon source, and anaerobiosis was achieved by boiling and cooling the medium before inoculation. Since this initial report, only a limited number of microbial taxa have been confirmed as capable of biosurfactant production under oxygen-limited or strictly anaerobic conditions (Eras-Muñoz et al. 2022; Hu et al. 2019).
Table 2 summarizes the bacterial isolates reported as anaerobic biosurfactant producers with demonstrated potential for CO_2_ fixation. Genera including Bacillus, Pseudomonas, and Clostridium stand out for their ability to reduce surface tension and interfacial tension and to produce structurally diverse biosurfactants under anoxic conditions. These organisms typically use organic acids, hydrocarbons, or CO_2_-derived intermediates as carbon sources, depending on their metabolic strategy. Table 2 highlights the substantial heterogeneity in experimental design and reporting across studies. Quantitative metrics such as productivity, growth rates, and biomass accumulation are inconsistently reported and often measured under non-comparable cultivation conditions. This lack of standardized performance indicators currently limits robust cross-study comparisons and represents a major bottleneck for evaluating the true potential of CO_2_-driven biosurfactant production within Microbial-CCUS frameworks. Table 2. Overview of bacterial growth and biosurfactant production under oxygen-limited and anaerobic conditionsMicroorganismBiosurfactant typeGrowthCulture conditionsCarbon sourceGrowth-related metricsBiosurfactant yieldBiosurfactant evidenceReferencesD. desulfuricans DSM 1926UnknownAnaerobicMineral medium30 °CLactateNRNR44–46 mN·m^−1^ STOil-spreadingEOR 35%(La Riviere et al., 1955)C. pasteurianum NRRL B-598Neutral lipidAnaerobicModified Mallette mediumRoom temperatureSucroseNRNR30 mN·m^−1^ STFoaming activityRelative CMC(Cooper et al. 1980)B. mojavensis JF-2Lichenysin BAnaerobicMineral medium40 °CGlucoseA_480_ 0.5–1.05100–700 mg/L27–31 mN·m^−1^ STFoaming activityRelative CMC(Javaheri et al. 1985)B. licheniformis BAS50Lichenysin AAnaerobicMineral medium40–45 °CGlucoseSucroseNR160 mg/L35 mN·m^−1^ STCMC 12 mg/L(Yakimov et al. 1995)B. licheniformis BNP29; BNP36; Mep132LipopetideEPSAnaerobicCNY medium50 °CCrude oilNR50 mg/L80–1600 mg/LEOR 22%(Yakimov et al. 1997)B. subtilis ATCC 21332SurfactinAnaerobicMineral medium32 °CGlucoseBiomass 5.8–6 g/L439 mg/LHPLC(Davis et al. 1999)B. licheniformis**B. mojavensisLipopetideAnaerobicMineral medium37 °CSucroseNRNROil-spreading(Youssef et al. 2005)Bacillus sp. RS-1B. subtilis subsp. spizizenii NRRL B-23049LipopetideAnaerobicMineral medium37 °CGlucoseNR90 mg/LHPLCRelative STOil-spreading(Youssef et al. 2007)B. licheniformis VKM B-511LipopetideAnaerobicMineral medium30 °CKeroseneA_600_ 0.4552590 mg/L35.3 mN·m^−1^ STE24 46.1%(Gogotov and Miroshnikov 2009)B. mojavensis GMTB-C1-2LipopetideAnaerobicMedium E42 °CPeptidesAmino acidsA_600_ 0.650NR27 mN·m^−1^ STE24 56–70%(Ghojavand et al., 2011)B. subtilis#191; #309; #311; #552; #573LipopetideSurfactin-likeAnaerobicMineral medium40 °CHexadecaneBiomass 0.176–0.238 g/LNR30–41.5 mN·m^−1^ STCMC 20–30 mg/LE24 13.3–34.2%(Gudina et al., 2012)B. amyloliquefaciens S499SurfactinsIturinsFengycinsOxygen-limitedRE medium30° CGlucoseA_600_ 0.9252 mg/L;108 mg/L;10 mg/LLC–ESI–MS(Nihorimbere et al. 2012)Bacillus sp. ISTS2LipopeptidesFree fatty acidsOxygen-limitedMSM medium30 °CCO_2_NRNRGC–MS33 mN·m^−1^ STE24 53%(Sundaram and Thakur 2015)Bacillus subtilis DSM 10^ T^SurfactinAnaerobicMSM medium30 °CGlucoseCDWmax 0.856 g/L150 mg/LHPLC(Willenbacher et al. 2015)B. licheniformis 421Lichenysin GAnaerobicSynthetic seawater50 °CMolassesA_600_ 1.29.60 mg/LUHPLC-MS0.60 mN·m^−1^ IFT(Halim et al. 2017)Bacillus sp. SS105LipopetideElevated CO_2_ conditionsMSM medium30 °CNaHCO_3_MolassesBiomass 2.78 g/L2650 mg/LFTIR and NMROil-spreadingE24 63%(Masheshwari et al., 2017)B. subtillis ATCC 6633SurfactinAnaerobicMineral medium37 °CGlucoseNR62 mg/LFTIRIFT reduction(Park et al. 2017)B. subtilis JABs24; MG1; MG5SurfactinAnaerobicMineral medium37 °CGlucoseCDWmax 0.332 g/L–0.492 g/L85.6–190 mg/LHPLC(Hoffmann et al. 2020)Bacillus subtilis AnPL-1SurfactinAnaerobicMineral medium39 °CSucroseA_600_ 3.953182.9 mg/L28.5 mN·m^−1^ STCMC 30 mg/LE24 70.5%(Zhao et al. 2021a)B. licheniformis EL3LipopetideOxygen-limitedMSM medium37 °CGlucoseBiomass 0.108 g/L775 mg/g biomassFTIR29 mN·m^−1^ STCMC 27 mg/L(Leal et al. 2024)Pseudomonas sp. BS2201Pseudomonas sp. BS2203UnknownAnaerobicPPGA medium28 °CPetroleumNRNR38 mN·m^−1^ ST34.7 mN·m^−1^ ST(Grishchenkov et al. 2000)P. aeruginosa PAO1TRhamnolipidAnaerobicModified Davis medium30 °CGlycerolNRNRTLC(Nozawa et al. 2007)Pseudomonas sp. ANBIOSURF-1GlycolipidAnaerobicMineral medium30 °CCoconut oilNRNRFTIR and TLCCMC 52 mg/L(Albino and Nambi 2010)P.aeruginosa E03-40RhamnolipidOxygen-limitedMineral medium34 °CGlycerolNR17 mg/g (dry cells)-hColorimetric analysis(Pinzon et al. 2013)P. stutzeri RhlRhamnolipidAnaerobicMineral medium42 °CGlycerolNR3120 mg/LColorimetric analysisEOR 15.7%(Zhao et al. 2014)P. aeruginosa SGRhamnolipidAnaerobicMineral medium37 °CGlycerolNR1080 mg/LFTIR and TLC33.3 mN·m^−1^ STCMC 80 mg/LE24 > 80%, EOR 8.33%(Zhao et al. 2015a; 2015c; 2021b)P. aeruginosa PrhlAB; PoprABRhamnolipidAnaerobicMineral medium37 °CGlycerolNR2420–3560 mg/L37.2 mN·m^−1^ ST(Zhao et al. 2015c)P. stutzeri WJ-1RhamnolipidAnaerobicGN medium37 °CGlycerolBiomass 5.1 g/L390 mg/LColorimetric analysis33 mN·m^−1^ ST(Zhao et al. 2015b)P. stutzeri CX3GlycolipidLipopeptidesAnaerobicMineral medium30 °CGlycerolBiomass 2.6 g/LNRFTIR, TLC and GC-FID30.4 mN·m^−1^ STCMC 35 mg/L(Fan et al. 2018)P. stutzeri DQ3RhamnolipidAnaerobicMineral medium42 °CGlycerolNR228 mg/L33.8 mN·m^−1^ STOil-spreadingE24 58%(Zhao et al. 2018)P. putida KT2440RhamnolipidAnaerobicModified Wilms KPi medium30 °CAcetateA_600_ 2–2.5366–414 mg/LColorimetric analysis(Widberger et al. 2024)P. aeruginosa SGhmRhamnolipidAnaerobicGNP medium35 °CGlycerolA_600_ 0.5–0.61540 mg/LE24 89.4%(Zhao et al. 2025)T. pseudethanolicus 39EUnknownAnaerobicMineral medium65 °CGlucoseTryptoneA_660_ 0.8–1.2NRTLC(Yen et al. 1991)Brevibacillus sp. BS2202UnknownAnaerobicPPGA medium28 °CPetroleumNRNR31.6 mN·m^−1^ STOil-spreading(Grishchenkov et al. 2000)A. thermohalophila Fru22UnknownAnaerobicMineral medium50 °CHexadecaneGlucoseNRNRSDS-PAGERelative ST reduction(Denger et al. 2002)G. pallidus H9GlycolipopeptideAnaerobicMinimal medium65 °CCrude oilBiomass 9.11 g/L2160 mg/L32.25 mN·m^−1^ STCMC 22 mg/LE24 10–35%(Wenjie et al., 2012)R. ruber Z25UnknownAnaerobicMineral medium37 °CParaffinBiomass 0.11 g/L530 mg/L29.54 mN·m^−1^ ST1.0 mN·m^−1^ IFTCMC 57–133 mg/LEOR 25.8%(Zheng et al. 2012)Fusant strain FA-2LipopeptideAnaerobicMedium E37 °CSucroseA_660_ 1.8382 mg/LTLC and LC–MS31.2 mN·m^−1^ STCMC 60 mg/LE24 50%, EOR 5.22%(Liang et al. 2017)L. huabeiensis HB-2LipopeptideAnaerobicFermentation medium40 °CGlucoseA_660_ 0.830300–400 mg/LFTIR and TLCE24 40%, EOR 11%(Ke et al. 2018)C. daeguensis HB-4UnknownAnaerobicMineral medium37 °CGlucoseNRNREOR 20.5%(Ke et al. 2019)H. zhaodongensis 253UnknownOxygen-limitedMineral medium28 °CCO_2_NRNRE24 68.5%(Santos et al. 2024)E. alkaliphilum 283UnknownOxygen-limitedMineral medium28 °CCO_2_NRNRE24 67.3%(Santos et al. 2024)M. hydrocarbonocalsticus and Thalssopspira sp. (consortia)LipopeptideAnaerobicSeawater37 °CDieselNR4200 mg/L66.9 mN·m^−1^ STCMC 3300 mg/L(Wahby et al. 2025)E. proteobaterium and Desulfovibrio sp.(consortia)LipopeptideAnaerobicSeawater37 °CDiesel-sodium lactateNR4200 mg/L66.5 mN·m^−1^ STCMC 3400 mg/LE24 45.8%(Wahby et al. 2025)Agarivorans sp. and MVP-15 unclassified (consortia)LipopeptideAnaerobicSeawater37 °CDiesel-glucoseNR5300 mg/L47.3 mN·m⁻^1^ STCMC 4200 mg/LE24 58.4%(Wahby et al. 2025)Marinobacter sp. and Vibrio sp. (consortia)LipopeptideAnaerobicSeawater37 °CDieselNR4400 mg/L33.4 mN·m^−1^ STCMC 3500 mg/L(Wahby et al. 2025)Desulfocurvus thunnarius and D. hontreensis(consortia)LipopeptideAnaerobicSeawater37 °CDiesel-sodium lactateNR6400 mg/L55.5 mN·m^−1^ STCMC 3000 mg/LE24 46.8%(Wahby et al. 2025)Agarivorans sp. and *D. hontreensis *(consortia)LipoproteinAnaerobicSeawater37 °CDiesel-glucoseNR6600 mg/L49.8 mN·m^−1^ STCMC 3000 mg/LE24 45.7%(Wahby et al. 2025)NR: not reported. Where quantitative metrics were unavailable. CDWmax: maximum cell dry weight, EOR: enhanced oil recovery, E24: emulsification index.
Bacillus
Among facultative anaerobes, Bacillus represents the most extensively documented genus for biosurfactant production under oxygen deprivation. Well-characterized isolates such as B. mojavensis JF-2 (formerly B. licheniformis) produce the lipopeptide lichenysin under strictly anaerobic conditions, achieving surface tension values below 30 mN·m^−1^ (Folmsbee et al. 2006; Javaheri et al. 1985). Subsequent studies on B. licheniformis strains (e.g., BAS50 and the BNP series) confirmed anaerobic lipopeptide synthesis with efficiencies approaching those of commercial surfactants (Yakimov et al. 1995, 1997). In some cases, anaerobic cultivation yielded higher biosurfactant titers than aerobic conditions (2.59 g/L vs. 0.69 g/L), although these comparisons remain study-specific and dependent on cultivation parameters (Gogotov & Miroshnikov 2009).
Oxygen-limited cultivation has also shown reduced excessive foaming, a critical bottleneck in industrial-scale-up biosurfactant production (Hoffmann et al. 2020). Recent evidence further indicates that oxygen limitation, rather than strict anaerobiosis, can represent an optimal metabolic window for Bacillus. Leal et al. (2024) demonstrated that Bacillus isolates maintain high lipopeptide productivity and achieve surface tension values below 30 mN·m^−1^ under oxygen-limited conditions, combining reduced foaming with enhanced process stability and extracellular biosurfactant accumulation.
Nevertheless, robust biosurfactant production under strictly anaerobic conditions remains uncommon. A large screening involving 207 bacterial strains revealed that only 35 tolerated anaerobiosis in the presence of 5% NaCl, with biosurfactant production under strictly anaerobic conditions restricted to B. mojavensis and B. licheniformis (Youssef et al. 2005). Subsequent field-scale evidence demonstrated that selected Bacillus strains not only tolerate but actively produce lipopeptide biosurfactants under reservoir anaerobic conditions. In a limestone petroleum reservoir, Bacillus strain RS-1 and B. subtilis subsp. spizizenii produced lipopeptides at average concentrations of 90 mg L^−1^, exceeding the critical micelle concentration (CMC) required for oil mobilization, with confirmed nitrate-respiring metabolism and carbon-balanced anaerobic fermentation (Youssef et al. 2007). These findings emphasize both the challenge of identifying robust anaerobic producers and the unique ecological and biotechnological value of Bacillus as an effective production chassis.
Mechanistic and genomic evidence further support a functional coupling between CO_2_ metabolism and biosurfactant synthesis in Bacillus. At the physiological level, anaerobic cultivation of B. licheniformis 421 revealed that despite low lichenysin G titers in the aqueous phase, pronounced reductions in interfacial tension and stable emulsion formation were achieved through bacterial accumulation at the oil–water interface, highlighting the importance of localized and cell-bound biosurfactant activity under oxygen deprivation (Halim et al. 2017). Consistent with this physiological behavior, enzymatic activities such as carbonic anhydrase and RuBisCO, together with biosynthetic markers (e.g., licA3, lipP1, C23O), suggest integration of carbon assimilation pathways and lipopeptide production (Phetcharat et al. 2019; Sundaram & Thakur 2015). Moreover, halotolerant and thermotolerant isolates, including oilfield-derived B. subtilis strains and B. mojavensis GMTB-C1-2, thrive under elevated salinity and temperature, while reference strains (ATCC 21332/6633) demonstrate anaerobic surfactin synthesis applied to interfacial control in CO_2_–brine systems (Davis et al. 1999; Ghojavand et al., 2011; Guez et al. 2008; Park et al. 2017). Process innovations reported in Table 2 include nitrate-respiring media (e.g., B. subtilis AnPL-1 producing around 150 mg/L surfactin on sucrose), foam-free bioreactor operation (B. subtilis JABs24), and oxygen-limited production of multiple lipopeptide families (B. amyloliquefaciens S499, producing surfactins, iturins, and fengycins) (Gudiña et al., 2012; Hoffmann et al. 2020; Maheshwari et al. 2017; Nihorimbere et al. 2012; Willenbacher et al. 2015; Zhao, et al. 2021a). Collectively, these findings establish Bacillus as a priority microbial chassis for anaerobic biosurfactant-CO_2_ coupling, with actionable optimization levers including oxygen and electron-acceptor management, CO_2_/bicarbonate supply, and fatty-acid composition.
Pseudomonas
Species of Pseudomonas combine metabolic versatility with the ability to synthesize glycolipid biosurfactants under anaerobic conditions, primarily through nitrate respiration (Zhao, et al. 2015a). Strains isolated from petroleum-contaminated environments (e.g., BS2201 and BS2203) reduce surface tension to 34–38 mN·m^−1^ under nitrate-reducing conditions (Grishchenkov et al. 2000), while P. aeruginosa produces rhamnolipids during hydrocarbon catabolism (Chayabutra et al. 2001). Anaerobic growth has also been reported under high-CO_2_ atmospheres, where biosurfactants contribute directly to cellular adaptation and membrane stabilization (Nozawa et al. 2007), with production demonstrated in strains such as ANBIOSURF-1 cultivated on vegetable oils (Albino & Nambi 2010). Similarly, the environmental isolate CX3 exhibits rhamnolipid production under anaerobic, nitrate-reducing conditions, further supporting the ecological relevance of glycolipid biosurfactant synthesis by naturally occurring Pseudomonas strains in oxygen-depleted environments (Fan et al. 2018).
Advances in bioprocess engineering have significantly improved anaerobic rhamnolipid production, including immobilized-cell systems for continuous operation (Pinzon et al. 2013), nitrate-respiring bioreactors operated at elevated temperatures (Zhao et al. 2018), and targeted genetic modification of biosynthetic pathways (Zhao et al. 2015a, c, 2021b). These approaches have achieved biosurfactant titers above 1.6 g/L under oxygen-limited conditions (Zhao et al. 2014). Moreover, co-culture systems sustain production in sulfide-rich oilfield environments (Zhao et al. 2015b), highlighting the resilience of Pseudomonas under geologically relevant stresses.
Notably, genetic studies in Pseudomonas aeruginosa indicate that efficient anaerobic rhamnolipid production is not an inherent physiological trait but rather the result of targeted metabolic engineering. In the SG strain, the wild-type background exhibits limited biosurfactant synthesis under nitrate-respiring conditions, whereas substantial yield improvements are achieved only after overexpression of key biosynthetic operons (rhlAB and rmlBDAC). These findings identify transcriptional regulation and precursor supply as primary bottlenecks for anaerobic secondary metabolism, while also demonstrating that enhanced biosurfactant titers can be attained without proportional increases in biomass, reinforcing the decoupling between growth and product formation under oxygen limitation (Zhao et al. 2015a, c; Zhao et al. 2025).
As summarized in Table 2, these attributes position Pseudomonas as a promising platform for integrating anaerobic biosurfactant production with CO_2_-rich industrial streams.
Clostridium and other related taxa
Beyond facultative anaerobes, several obligate anaerobic and extremophilic taxa further expand the phylogenetic and metabolic scope of anaerobic biosurfactant production. Early studies identified Clostridium pasteurianum and Thermoanaerobacter pseudethanolicus 39E as pioneer models secreting low-molecular-weight surface-active compounds under thermophilic, halotolerant, and strictly anaerobic conditions (Cooper et al. 1980; Yen et al. 1991). In contrast to fermentative anaerobes, taxa with distinct metabolic strategies have demonstrated even more pronounced interfacial performance. In this context, Rhodococcus ruber Z25 achieved exceptional interfacial performance (IFT 1.0 mN·m^−1^), enhanced oil recovery up to 25.8% during MEOR trials (Zheng et al. 2012). However, the thermophilic bacterium Geobacillus pallidus H9 exhibited a pronounced dependence on oxygen availability, with biosurfactant yields decreasing from 9.8 to 2.8 g/L under oxygen-limited conditions across temperatures of 45–80 °C and salinities up to 15% (Wenjie et al., 2012). Rather than representing an anaerobic advantage, this response highlights oxygen sensitivity as a critical constraint in certain thermophilic systems. Conversely, other extremophiles retain functional biosurfactant activity precisely under oxygen limitation. Complementarily, the alkaliphilic isolate Exiguobacterium alkaliphilum 283 exhibited pronounced emulsification activity under oxygen-limited conditions at elevated pH, underscoring the role of surface-active compounds in interfacial stabilization and stress adaptation in extreme environments (Santos et al. 2024).
Among halophilic systems, Halomonas stevensii efficiently coupled CO_2_ fixation with biosurfactant production, removing up to 98% of inlet CO_2_ under thiosulfate-driven metabolism and growing across broad salinity, pH, and temperature ranges (Mishra et al. 2017). Similarly, Halomonas zhaodongensis strain 253 sustained biosurfactant production under CO_2_-enriched (5–10%) and oxygen-limited conditions, maintaining emulsification indices around 60% and encoding carbonic anhydrase genes (γ-CA and CsoSCA) that facilitate CO_2_ hydration and bicarbonate utilization (Santos et al. 2024). Additional taxa, including Brevibacillus sp. BS2202, Anaerophaga thermohalophila Fru22, Luteimonas huabeiensis HB-2, and Chelatococcus daeguensis HB-4, further demonstrate biosurfactant-mediated interfacial modification and oil recovery under oxygen-deprived conditions (Denger et al. 2002; Grishchenkov et al. 2000; Ke et al. 2018, 2019).
Extending the single-strain perspective, Wahby et al. (2025) demonstrated that anaerobic biosurfactant functionality can also emerge robustly at the community level. Microbial consortia cultivated in diesel–sodium lactate and diesel–glucose systems produced biosurfactants with emulsification indices (E24) of 45.7–46.8%, CMC around 3.0 g/L, and relatively high biosurfactant titers (6.4–6.6 g·L⁻^1^), while achieving surface tension values of 49.8–55.5 mN·m⁻^1^ under oxygen-depleted conditions. Community composition analyses revealed mixed groups dominated by fermentative, hydrocarbon-degrading, and sulfate-reducing taxa, including Marinobacter hydrocarbonoclasticus, Thalassospira sp., Epsilon proteobacteria, Desulfovibrio sp., Desulfocurvus thunnarius, Agarivorans sp., and Vibrio sp., alongside several previously unreported biosurfactant-producing genera. Importantly, the functional interfacial performance of these consortia did not depend on exceptional surface tension reduction by individual members but rather on metabolic complementarity and cooperative interactions, highlighting community-level resilience and sustained emulsification capacity under reservoir-like anaerobic conditions.
Recent methodological advances, such as microplate-based anaerobic screening of oilfield samples, have accelerated the identification of new biosurfactant producers with surface tension values below 45 mN·m^−1^ (Zhao et al. 2022). At the process level, integrated systems coupling chemolithoautotrophic CO_2_ fixation (e.g., Acetobacterium woodii) with heterotrophic rhamnolipid synthesis by Pseudomonas putida illustrate emerging strategies for linking CO_2_ capture and biosurfactant production within unified platforms (Widberger et al. 2024). Overall, these taxa collectively demonstrate that anaerobic biosurfactant-CO_2_ coupling is a broadly distributed and mechanistically versatile trait, spanning facultative and obligate anaerobes as well as extremophilic lineages.
5. Challenges and future directions
Despite increasing industrial demand for sustainable surfactants, anaerobic biosurfactant production linked to CO_2_ fixation remains constrained by high costs, scale-up challenges, and downstream processing bottlenecks (Farias et al. 2021). In anoxic systems, slower microbial growth, inefficient substrate conversion, and yield variability further exacerbate these barriers. A major limitation lies in the scarcity of direct evidence for CO_2_/HCO_3_^−^ incorporation into biosurfactant precursors. Only a few isolates, such as Halomonas species, have shown promising emulsification under CO_2_-enriched conditions, though metabolic fluxes remain poorly resolved (Mishra et al. 2017; Santos et al. 2024). Addressing this knowledge gap requires not only improving biosurfactant yield but also mechanistically linking CO_2_ fixation pathways to surfactant biosynthesis within a Microbial-CCUS framework.
Recent studies have started to bridge this divide through genetic and process innovations. Hybrid or synthetic strains, such as the FA-2 protoplast fusion (Bacillus mojavensis × Pseudomonas stutzeri), have demonstrated lipopeptide production under anaerobiosis (382 mg/L), suggesting that engineered systems can channel inorganic carbon into amphiphilic molecules (Liang et al. 2017). Advances in high-throughput screening methods (e.g., microplate assays, oil displacement tests) now enable rapid strain discovery and preliminary tracking of carbon flux into glycolipid and lipopeptide backbones (Zhao et al. 2022; Widgerber et al., 2024).
At the technological level, bioreactor innovations are essential for ensuring efficient gas transfer, maintaining redox balance, and replicating subsurface pressures to sustain continuous CO_2_ uptake in biosurfactant-producing cultures. In addition, prolonged operation under high cell density and complex feed streams may promote biofilm formation and biofouling on reactor surfaces, potentially affecting mass transfer efficiency and maintenance requirements. Although not intrinsic to biosurfactant metabolism, such phenomena represent practical engineering limitations that must be addressed to ensure stable long-term Microbial-CCUS performance (Hoffmann et al. 2020; Zhao et al. 2018).
Functional characterization of CO_2_-derived biosurfactants, evaluating their CMC, stability, toxicity, biodegradability, and performance under saline or high-pressure conditions will ultimately determine their competitiveness with synthetic analogs and conventional aerobic products (Zhang et al. 2022).
Furthermore, computational tools and AI-driven metabolic design are redefining microbial carbon utilization optimization. Genetic algorithm-based AI and sequence-based predictors, such as Evo and ProGen, can analyze vast genomic datasets to reveal functional gene networks and optimize pathways for biosurfactant biosynthesis and CO_2_ fixation (Januszewski & Jain 2024; Nguyen et al. 2024; Sasse et al. 2024). Integrating AI-guided predictions with microbial genetics expertise offers a powerful route for targeted pathway design, reduced experimental effort, and accelerated strain optimization (Goodswen et al. 2021; Simon et al. 2024).
Ultimately, techno-economic and life-cycle assessments must determine whether CO_2_-derived biosurfactants can compete not only with synthetic surfactants but also with conventional microbial ones (Sarubbo et al. 2022). By coupling the discovery of novel isolates with verified CO_2_ incorporation, advanced genetic engineering, and tailored cultivation systems, this emerging field can expand our understanding of microbial adaptation to anoxic niches while creating innovative Microbial-CCUS strategies that deliver both economic and environmental value. Collectively, these integrative efforts position CO_2_-derived biosurfactants as pivotal components of a low-carbon bioeconomy, embedding them within global climate mitigation and sustainable industrial frameworks (Eras-Muñoz et al. 2022; Kugaji et al. 2025; Sarubbo et al. 2022). Achieving this integration at scale ultimately depends on advances at the molecular level.
5.1 Genetic and synthetic biology approaches to integrate CO2 fixation and biosurfactant production
At the molecular level, recent progress in genetic and synthetic biology provides the foundation for directly coupling CO_2_ assimilation with biosurfactant biosynthesis. Although experimental demonstrations remain limited, the combination of pathway engineering, modular design, and genome editing is enabling microbial platforms that coordinate carbon fixation with surfactant production within the Microbial-CCUS paradigm (Coimbra et al. 2025; Lea-Smith et al. 2025).
Synthetic biology now allows the construction of artificial operons linking CO_2_-fixing modules to reconfigured metabolic routes, channeling assimilated carbon toward the synthesis of multicarbon compounds. For example, engineered Escherichia coli strains expressing the RuBisCO–PRK cycle have shown enhanced CO_2_ assimilation and increased flux toward succinate and amino acid precursors (Zhou et al. 2025). Similarly, modular systems with optimized promoters, ribosome-binding sites, and artificial carboxysomes enable spatial coordination between carbon fixation and downstream biosynthesis. The inclusion of RuBisCO activator proteins in recombinant carboxysomes significantly improves enzyme activation and CO_2_ capture efficiency, extending the applicability of metabolic engineering for tailored compound production (Chen et al. 2022; Liu et al. 2022).
In addition to autotrophic models, the application of CRISPR-Cas systems to organisms such as Cupriavidus necator and anaerobic acetogens like Clostridium autoethanogenum has expanded the potential for carbon fixation through genetically tractable bacterial chassis. In particular, facultative or obligate anaerobes represent ideal platforms for anaerobic Microbial-CCUS. These genome-scale interventions allow for precise redirection of metabolic fluxes toward amphiphilic and lipidic products, including biosurfactant precursors (Yu et al. 2022). Extending these strategies to anoxic or microaerophilic bacteria remains a promising yet underexplored direction for true CO_2_-driven biosurfactant production.
Complementary computational frameworks are accelerating this progress. Machine learning platforms such as the Automated Recommendation Tool (ART) assist in predicting optimal genetic modifications and pathway rewiring for targeted compound synthesis (Chen & Xia 2023). Furthermore, recent reviews emphasize that robust microbial cell factories designed for sustainable manufacturing depend on finely tuned metabolic engineering to enhance the synthesis of multicarbon and lipophilic metabolites (Yan et al. 2024). Together, these genetic, synthetic, and computational strategies delineate a realistic roadmap toward next-generation microbial cell factories capable of converting inorganic carbon into high-value amphiphilic molecules, aligning biomanufacturing innovation with climate change mitigation and circular carbon economy goals.
6. Conclusions
The exploration of biosurfactants derived from microbial CO_2_ capture represents a promising opportunity to address urgent environmental challenges while advancing sustainable industrial practices. These compounds offer eco-friendly and biodegradable alternatives to synthetic surfactants, with potential applications in environmental remediation, agriculture, pharmaceuticals, and cosmetics. Both photosynthetic and non-photosynthetic bacteria provide suitable model organisms for microbial CCUS strategies. Advances in genetic engineering and systems biology enable the optimization of biosynthetic pathways, improved yields, and greater economic feasibility. Furthermore, integrating artificial intelligence into biosurfactant research holds additional potential to enhance process efficiency and resource utilization.
To date, quantitative data comparing CO_2_ fixation rates and biosurfactant yields under integrated microbial-CCUS systems remain scarce, as most studies focus either on CO_2_ assimilation or on biosurfactant productivity in isolation. Despite these advances, microbial CO_2_ capture and biosurfactant synthesis technologies remain at an early stage of development, typically between Technological Readiness Levels (TRL) 3 and 4. Most studies have been conducted under laboratory or small pilot conditions, where process efficiency, energy balance, and product recovery costs still represent major uncertainties. Achieving economic viability will require improving CO_2_ fixation rates and biosurfactant yields through metabolic engineering, as well as ensuring access to low-cost CO_2_ sources and renewable feedstocks. Integrating microbial CCUS processes with existing industrial emitters offers a promising route to reduce operational costs and accelerate scale-up. In parallel, comprehensive life cycle and techno-economic assessments will be essential to benchmark these systems against conventional petrochemical surfactants. Progressing toward TRL 6–7 will demand demonstration-scale facilities capable of continuous operation, validation of technical performance, and the implementation of policy instruments—such as carbon credits and green certification schemes—to foster market acceptance.
Future integration of CO_2_ assimilation with industrial infrastructures, alongside process optimization, is expected to facilitate progressive scale-up and greater economic feasibility of microbial CCUS platforms. However, widespread deployment remains constrained by challenges related to scalability, process optimization, and cost competitiveness. Overcoming these limitations will require coordinated efforts among academia, industry, and policymakers, supported by targeted incentives, regulatory frameworks, and multidisciplinary collaboration. Achieving these advancements could ultimately enable the deployment of sustainable microbial CCUS platforms, establish economically viable biosurfactant production at scale, drive greener industrial practices, contribute to global carbon reduction, and promote a circular bioeconomy that aligns environmental stewardship with economic growth.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Araújo WJ, Oliveira JS, Araújo SCS, Minnicelli CF, Silva-Portela RCB, da Fonseca MMB, Freitas JF, Silva-Barbalho KK, Napp AP, Pereira JES, Peralba MCR, Passaglia LMP, Vainstein MH, Agnez-Lima LF (2020) Microbial Culture in Minimal Medium With Oil Favors Enrichment of Biosurfactant Producing Genes. Front Bioeng Biotechnol Vol 8-2020. https://www.frontiersin.org/journals/bioengineering-and-biotechnology/articles/10.3389/fbioe.2020.00962
- 2Bardi MJ, Müller F, Polag D, Gabbiye Habtu N, Koch K (2025) The intriguing effect of CO 2 enrichment in anaerobic digestion. Bioresour Technol 416. 10.1016/j.biortech.2024.131743
- 3International Energy Agency (2021) Technology perspectives—special report on carbon capture, utilisation and storage: CCUS in clean energy transitions. www.iea.org/t&c/. Accessed 10 Nov 2025
- 4Mills LA, Mc Cormick AJ, Lea-Smith DJ (2020) Current knowledge and recent advances in understanding metabolism of the model cyanobacterium Synechocystis sp. PCC 6803. Biosci Rep. 10.1042/BSR 20193325
- 5Rajkumar R, Takriff MS, Veeramuthu A (2022) Technical insights into CO 2 sequestration by microalgae: a biorefinery approach. Biomass Convers Biorefin 12. 10.1007/s 13399-022-02446-9
- 6Ritchie H, Rosado P, Roser M (2023) CO 2 and greenhouse gas emissions. Our World in Data. https://ourworldindata.org/co 2-and-greenhouse-gas-emissions. Accessed 18 Nov 2025
- 7Santos D, Napp AP, Aguiar CP, Dutra WL, Gonçalves BX, Vecchia D, Tewari RDB (2024) Carbon dioxide as a carbon source for biosurfactant production. SSRN preprint 5066224. https://ssrn.com/abstract=5066224
- 8Yakimov MM, Timmis KN, Wray V, Fredrickson HL (1995) Characterization of a new lipopeptide surfactant produced by thermotolerant and halotolerant subsurface Bacillus licheniformis BAS 50. Appl Environ Microbiol 61:1706–1713. 10.1128/AEM.61.5.1706-1713.1995