Co-occurrence of mcr-9 and blaNDM-1 in carbapenem-resistant Enterobacter hormaechei from burn patients
Xingchen Tao, Lingyi Zeng, Xinyi Sun, Yuhai Du, Liping Zhong

TL;DR
This study identifies carbapenem-resistant Enterobacter hormaechei strains from burn patients that carry two resistance genes, posing a risk of spreading antibiotic resistance.
Contribution
The co-occurrence of mcr-9 and blaNDM-1 in Enterobacter hormaechei is reported, with insights into plasmid mobility and genetic diversity.
Findings
Three CR-E. hormaechei strains co-harboring blaNDM-1 and mcr-9 were isolated from burn patients.
The mcr-9 gene was found on IncHI2 plasmids, and blaNDM-1 on IncX3 or IncHI2 plasmids.
All strains belonged to sequence type 97 and showed high genetic relatedness.
Abstract
Carbapenem-resistant Enterobacter (CRE) has emerged as a critical clinical concern due to its broad multidrug resistance. This study aims to characterize the features of clinical CRE strains co-harboring the blaNDM-1 and mcr-9 genes from three burn patients. This study collected 110 non-repetitive carbapenem-resistant Enterobacteriaceae (CRE) from clinical settings. blaNDM-1 and mcr-9 genes were identified by PCR, and strains were identified via MALDI-TOF MS and 16S rRNA sequencing. Minimum inhibitory concentrations (MICs) of common antimicrobial agents were determined by the broth microdilution method. The conjugation experiment was used to verify the transfer of resistance plasmids. Whole-genome sequencing (WGS) was performed using NovaSeq and PacBio_HIFI platforms, and analyzed through bioinformatics to characterize the resistance genes, virulence factors, plasmid profiles, and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
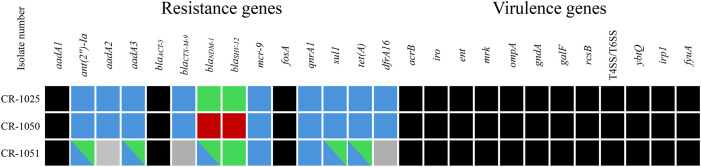 Figure 1
Figure 1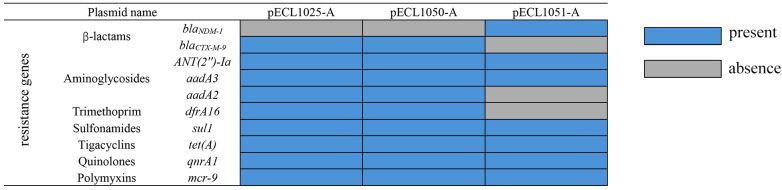 Figure 2
Figure 2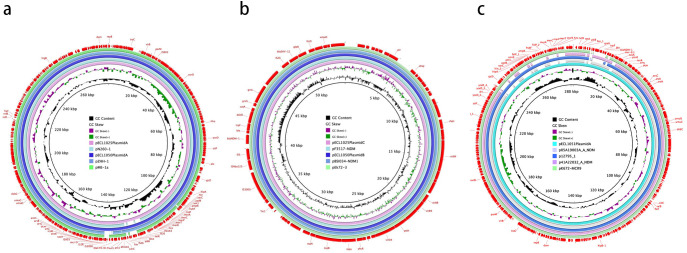 Figure 3
Figure 3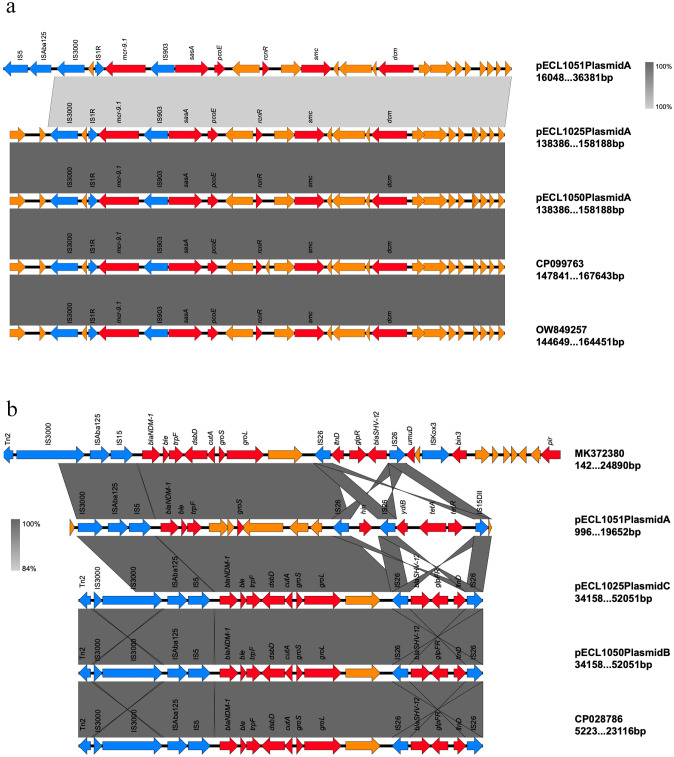 Figure 4
Figure 4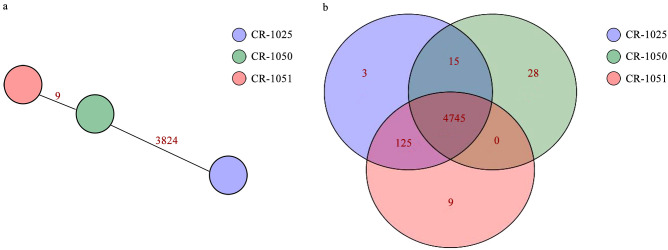 Figure 5
Figure 5| Isolate number | CR-1025 | CR-1050 | CR-1051 |
|---|---|---|---|
| Ward | ICU | Department of Burns and Plastic Surgery | ICU |
| Gender | male | male | male |
| Age(years) | 62 | 70 | 60 |
| Specimen | wound | blood | wound |
| Clinical diagnosis | burn | burn | burn |
| Outcome | died | improved | improved |
| Invasive operation | yes | yes | yes |
| Usage of colistin | yes | no | yes |
| Usage of other antibiotics | PTZ MFX CFS CAZ–AVI AZT | PTZ CFS TGC AMK | PTZ CFS TGC MEM |
| Isolate number | MIC (µg/mL) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CLS | PB | IMP | MEM | ETP | TGC | TC | CAP | CIP | LEV | AMK | GEM | TOB | CFS | PTZ | EFP | CRO | CAZ | AZT | |
| Carbapenem-resistant | |||||||||||||||||||
| CR-1025 | ≤1(I) | 4(R) | >8(R) | >8(R) | >2(R) | 2(R) | >8(R) | ≤4(S) | >4(R) | >8(R) | ≤8(S) | 4(S) | 4(S) | >32/8(R) | >64/4(R) | >16(R) | >32(R) | >32(R) | >32(R) |
| CR-1050 | ≤1(I) | 2(I) | >8(R) | >8(R) | >2(R) | 2(R) | >8(R) | ≤4(S) | >4(R) | >8(R) | ≤8(S) | 4(S) | 4(S) | >32/8(R) | >64/4(R) | >16(R) | >32(R) | >32(R) | >32(R) |
| CR-1051 | ≤1(I) | 4(R) | >8(R) | >8(R) | >2(R) | 2(R) | >8(R) | ≤4(S) | >4(R) | >8(R) | ≤8(S) | 8(I) | 8(I) | >32/8(R) | >64/4(R) | >16(R) | >32(R) | >32(R) | >32(R) |
| | <0.5(I) | <0.5(I) | <0.5(S) | 0.5(S) | <0.5(S) | <0.5(S) | ≤1(S) | ≤2(S) | <0.5(S) | <0.5(S) | ≤2(S) | ≤1(S) | ≤1(S) | ≤8/4(S) | ≤8/4(S) | <0.5(S) | <0.5(S) | <0.5(S) | ≤1(S) |
| J1025 | ≤1(I) | 1(I) | >8(R) | >8(R) | >2(R) | 2(R) | >8(R) | ≤4(S) | >4(R) | >8(R) | ≤8(S) | ≤2(S) | ≤2(S) | >32/8(R) | >64/4(R) | >16(R) | >32(R) | >32(R) | >32(R) |
| J1050 | ≤1(I) | 1(I) | >8(R) | >8(R) | >2(R) | ≤1(S) | ≤2(S) | ≤4(S) | ≤0.5(S) | ≤1(S) | ≤8(S) | ≤2(S) | ≤2(S) | >32/8(R) | >64/4(R) | >16(R) | >32(R) | >32(R) | >32(R) |
| J1051 | ≤1(I) | 1(I) | 8(R) | >8(R) | >2(R) | ≤1(S) | >8(R) | ≤4(S) | ≤0.5(S) | ≤1(S) | ≤8(S) | 4(S) | 8(I) | >32/8(R) | >64/4(R) | >16(R) | >32(R) | >32(R) | >32(R) |
| Isolate number | Plasmid numbers | Plasmid name | Colistin resistance gene | Carbapenem resistance genes | Plasmid size (bp) | Plasmid replicon type | G+C content (%) |
|---|---|---|---|---|---|---|---|
| CR-1025 | 4 | pECL1025-A |
| – | 278,515 | IncHI2 | 46.3 |
| pECL1025-C | – |
| 54,035 | IncX3 | 49.03 | ||
| CR-1050 | 3 | pECL1050-A |
| – | 278,515 | IncHI2 | 46.3 |
| pECL1050-B | – |
| 54,035 | IncX3 | 49.03 | ||
| CR-1051 | 4 | pECL1051-A |
|
| 281,800 | IncHI2 | 46.66 |
- —Medical Science and Technology Project of Zhejiang Province10.13039/501100017594
- —Science and Technology Bureau of Jiaxing City10.13039/501100019976
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotic Use and Resistance · Nosocomial Infections in ICU
Introduction
The Enterobacter cloacae complex (ECC) is an important component of Gram-negative Enterobacteriaceae, comprising six subspecies, namely, Enterobacter asburiae, Enterobacter hormaechei, Enterobacter kobei, Enterobacter ludwigii, Enterobacter nimipressuralis, and Enterobacter cloacae (Sanders et al., 1997; Mezzatesta et al., 2012). As an opportunistic pathogen, ECC has been associated with various clinical infections, such as bacteremia, respiratory tract infections, wound infections, and urinary tract infections (Huang et al., 2021).
Carbapenem-resistant Enterobacteriaceae (CRE) bacteria pose a serious threat to global public health (Ma et al., 2023), and production of carbapenemases is the main mechanism in CRE clinical isolates (Peng et al., 2022). New Delhi Metallo-β-lactamase (NDM) is a type of Metallo-β-lactamase (MBL) able to hydrolyze most β-lactams (including carbapenems), blaNDM-1 is often carried in carbapenem-resistant Enterobacter cloacae complex (CR-ECC) (Cai et al., 2024; Hu et al., 2022). The emergence of CR-ECC-carrying blaNDM-1 greatly limits the selection of clinical antibiotics and increases the difficulty of clinical anti-infective treatment. Colistin was once regarded as the last-resort antibiotic for treatment caused by CR-ECC (Halder et al., 2025). However, unfortunately, the activity and efficacy of colistin have been challenged by the global spread of plasmid-mediated colistin resistance genes (mcr) (Wu et al., 2023). Currently 10 mcr variants (mcr-1 to mcr-10) have been discovered, among them, mcr-1 and mcr-9 are the most widespread (Liu et al., 2022; Hussein et al., 2021). Since mcr-9 was first identified from Salmonella enterica in the United States (Carroll et al., 2019), it has been successively detected in 40 countries on six continents (Li et al., 2020), and has become a widely reported variant among CR-ECC (Wu et al., 2023).
This study isolated three ST97 strains of carbapenem-resistant Enterobacter hormaechei (CR-E. hormaechei) carrying both blaNDM-1 and mcr-9 from burned patients, and was conducted to explore the molecular characteristics, antimicrobial resistance (AMR) gene profile and potential transmission mechanism. Furthermore, the genetic background of plasmids was investigated by whole-genome sequencing (WGS).
Materials and methods
Sample collection
CRE is defined as resistance to any carbapenem drug, we totally collected 110 CRE from the clinical patients in Zhejiang province, China. The presence of carbapenemase genes and mcr genes was screened by PCR amplification using primers described previously (Supplementary material Table S1), and the positive products were sequenced using Sanger sequencing. Three strains were detected to carry both blaNDM-1 and mcr-9 simultaneously, and they were named as CR-1025, CR-1050, and CR-1051 respectively. Preliminary species identification was achieved by MALDI-TOF MS (Bruker Daltonik GmbH, Bremen, Germany) and 16s rRNA sequencing, all three strains were identified as ECC. And the identification of E. hormaechei species was confirmed by WGS.
Antimicrobial susceptibility testing
The minimum inhibitory concentration (MIC) values of strains against common antibiotics were determined according to the broth dilution method. In accordance with Clinical and Laboratory Standards Institute (CLSI) guidelines, the Escherichia coli (E. coli) ATCC 25922 was the quality control strain used for the MIC measurement. The results interpreted according to CLSI instructions, while tigecycline resistance was defined according to EUCAST clinical breakpoints.
Conjugation experiment
The conjugation experiment was carried out using a membrane filter mating experiment as previously described (Zeng et al., 2021; Wang et al., 2015). E. hormaechei as the donor strain and EC600 as recipient strain were mixed on Luria-Bertani agar at a ratio of 1:3, and the mixtures were incubated for 24 h at 35 °C. Transconjugants were selected on MHA plates supplemented with rifampicin (600 µg/ml) and meropenem (1 µg/ml). Colonies that grew on the selective medium were identified by the MALDI-TOF MS and 16S rRNA sequencing. Strains that harbored mcr-9 and exhibited higher MICs of resistance to carbapenems than EC600 were defined as the transconjugants. The carrier status of other drug resistance genes and MIC value of the transconjugants were determined by PCR and antimicrobial susceptibility testing.
Whole-genome sequencing and analysis
Whole-genome sequencing (WGS) was carried out via a combination of the Illumina NovaSeq (Illumina, USA) and PacBio HiFi (Pacific Biosciences, USA) platforms. Concomitantly, sequence analysis yielded high-quality genome assemblies and fully closed genomes of the isolate as well as all its associated plasmids. This entire process was conducted by Majorbio Co., Ltd. (Shanghai, China). In brief, genomic DNA was isolated using a commercial DNA extraction kit (Qiagen, Hilden, Germany). Sequencing libraries were constructed with Illumina Nextera XT kits and subjected to paired-end 150-base sequencing on an Illumina NovaSeq X Plus platform. Furthermore, a dedicated library for PacBio sequencing was prepared using the Rapid Barcoding Sequencing kit and loaded onto an R9.4 flow cell.
For Illumina sequencing data assembly, SOAPdenovo 2.04 ((https://github.com/aquaskyline/SOAPdenovo2) was employed; whereas PacBio data were assembled using Unicycler v0.4.8 (https://github.com/rrwick/Unicycler/releases), Canu v2.2 (https://github.com/marbl/canu), Flye v2.9.2 (https://github.com/mikolmogorov/Flye), NextDenovo v2.5.2 (https://github.com/Nextomics/NextDenovo) and Hifiasm v0.16.1 (https://github.com/chhylp123/hifiasm) to obtain complete whole-genome and plasmid sequences. Genomic sequence annotation was performed using the Pfam Database 33.1 (http://pfam.xfam.org/). Plasmid replicons were identified via PlasFlow 1.1 (https://github.com/smaegol/PlasFlow) and annotated with the PLSDB Database 202106 (https://ccb-microbe.cs.uni-saarland.de/plsdb/). In silico multilocus sequence typing (MLST) was assigned through the PUBMLST database (https://pubmlst.org/databases/). The ChewBBACA method was utilized to identify the core genome, and cg-MLST results were visualized with PHYLOViZ based on the goeBURST algorithm. Acquired antibiotic resistance genes were detected using ResFinder 4.5.0 (https://bitbucket.org/genomicepidemiology/resfinder/src/master/), while virulence genes were predicted via the VFDB Database 20240301 (http://www.mgc.ac.cn/VFs/main.htm). Nucleotide sequences of plasmids or chromosomes carrying blaNDM-1 and mcr-9 were compared with homologous sequences using BLAST, and the results were visualized by BRIG (http://brig.sourceforge.net) or Easyfig v2.2.3 (https://github.com/mjsull/Easyfig).
Nucleotide sequence accession numbers
All sequencing data generated in this study have been deposited in the NCBI Sequence Read Archive under the BioProject accession number PRJNA 1327493 (Biosample: SAMN51284745), PRJNA 1327495 (Biosample: SAMN51284820), PRJNA 1327550 (Biosample: SAMN51284833).
Results
Clinical characteristics
Three strains of carbapenem-resistant E. hormaechei (CR-E. hormaechei) were isolated from elderly male patients, and all three patients were hospitalized with different degrees of burns, two of which were transferred to ICU for treatment due to serious injuries. Two patients (66.7%) received colistin for anti-infection during the treatment period, and all patients (100%) had a history of invasive surgical procedures and multiple antibiotic exposures. Please refer to Table 1 for specific details.
Antibiotic resistance phenotype
MICs of 19 antimicrobial agents are shown in Table 2. In addition to being resistant to all three carbapenems, CR-1025, CR-1050, and CR-1051 also exhibited resistance to tigecycline, tetracycline, ciprofloxacin, levofloxacin, cefoperazone-sulbactam, piperacillin-tazobactam, cefepime, ceftriaxone, ceftazidime, aztreonam, while remaining but still susceptible to chloramphenicol and amikacin. In contrast, CR-1050 exhibited intermediate susceptibility to both colistin and polymyxin B, whereas CR-1025 and CR-1051 displayed intermediate susceptibility to colistin and full resistance to polymyxin B. All experimental strains have a broad drug resistance spectrum and were classified as multidrug-resistant bacteria.
Results of plasmid conjugation test
The conjugation success rate was 100% (3/3). Both blaNDM-1 and mcr-9 were able to transfer from donor to recipient, confirming the horizontal transmission of the plasmid. After conjugation, the transconjugants showed a significant increase in resistance to carbapenems, while the MIC values for the other drugs exhibited varying degrees of changes. The specific results are detailed in Table 2.
Results of WGS
Resistome and virulence genes in isolates and location
In addition to co-harbored mcr-9 and blaNDM-1 genes, CR-1025 and CR-1050 also carried 12 other antimicrobial resistance genes mediating resistance to aminoglycosides (aadA1, ant(2’’)-Ia, aadA2b, aadA3), β-lactams (blaACT-5, blaCTX-M-9, blaSHV-12), fosfomycin (fosA), quinolones (qnrA1), sulphonamides (sul1), tetracycline (tet(A)) and trimethoprim (dfrA16). By contrast, CR-1051 harbored 9 other resistance genes. These resistance genes were distributed on different genetic carriers of bacteria (one chromosome and multiple plasmids). Some plasmids even carry multidrug-resistant genes, while certain drug-resistant genes were present on multiple plasmids.
In addition to resistance genes, CR-1025, CR-1050 and CR-1051 also carried many virulence genes on the chromosome. The profiles of their virulence genes are complex and are associated with such as bacterial adhesion, colonization, transcriptional regulation, and other biological processes. Please refer to Figure 1 for specific details.
Profile of resistance and virulence genes in CR- E. hormaechei. Black: Present and located on chromosome; Blue: Present and locate on plasmid A; Red: Present and locate on plasmid B; Green: Present and locate on plasmid C; Grey: Absent; Two colors: both of the two different plasmids corresponding to the different colors carry this gene. T4SS/T6SS: Type IV/VI secretion system.
Genetic characteristics of the plasmid-borne mcr-9
Whole-genome sequencing revealed that isolates CR-1025, CR-1050, and CR-1051 harbored 4, 3, and 4 plasmids, respectively. Plasmids in each isolate were designated alphabetically (A to D) in descending order of size; thus, CR-1025 and CR-1051 contained plasmids A–D, while CR-1050 contained plasmids A–C. In all three isolates, the largest plasmid (designated A) carried the mcr-9 gene and corresponded to pECL1025-A, pECL1050-A, and pECL1051-A, respectively. These mcr-9-positive plasmids belonged to the IncHI2 replicon type and carried genes involved in replication (repB), stability (parM), and conjugative transfer (tra and virB). CR-1025 and CR-1050 shared two pairs of homologous plasmids: one pair (pECL1025-A and pECL1050-A) carried mcr-9 and was 278,515 bp in size with a GC content of 46.30%; the other pair (pECL1025-C and pECL1050-B) carried NDM-1 and measured 54,035 bp with a GC content of 49.03%. Notably, pECL1051-A—also an IncHI2 plasmid—harbored mcr-9 along with six additional resistance genes, including NDM-1. Key characteristics of these plasmids are summarized in Table 3; Figure 2, and schematic maps are provided in Figure 3.
Distribution of antimicrobial resistance genes in plasmids carrying mcr-9. Blue: Resistance gene present in the corresponding mcr-9-harboring plasmid; Gray: Resistance gene absent in the corresponding mcr-9-harboring plasmid.
Schematic maps of plasmids harboring blaNDM-1 and (or) mcr-9. (a). Map of the plasmids pECL1025-A (this study), pECL1050-A (this study), pN260-1 (accession no. AP023448.1), pzHH-1 (accession no. CP059712.1), and pME-1a (accession no. CP041734.1). (b). Map of the plasmids pECL1025-C (this study), pECL1050-B (this study), pF3517-NDM (accession no. CP137176.1), pBSI034-NDM1 (accession no. MN937240.1), and pEk72-3 (accession no. CP088232.1). (c). Map of the plasmids pECL1051-A (this study), p05A19003A_A_NDM (accession no. PV023031.1), p12795_1 (accession no. CP083854.1), p41A22032_A_NDM (accession no. PV023161.1), and pK672-MCR9 (accession no. CP183850.1).
Analysis of the genetic environment of mcr-9
The genetic environments of mcr-9 in this study were highly similar and mcr-9 was located in an ~4-kb region surrounded by two insertion sequences IS903 and IS1R, there was also an IS3000 downstream. The region upstream of mcr-9 included the conserved gene structure, rcnR- pcoE- sasA, but lacked the downstream structure wbuC. These genes may be involved in the regulation of plasmid-related functions, such as metal ion transport regulation and plasmid replication regulation, and might influence the expression and function of mcr-9 to some extent, also reflecting, also reflecting the cooperative and interactive relationships between genes on the plasmid. These elements may play a crucial role in transferring mcr-9 (Figure 4a). A similar structure has been found in some IncHI2 plasmids such as p123 (Accession no. CP099763) and p1 (Accession no. OW849257).
Linearized analyses for genetic environment of mcr-9 and blaNDM-1. (a). mcr-9 harbored by pECL1051-A, pECL1025-A, and pECL1050-A in this study. (b). blaNDM-1 harbored by pECL1051-A, pECL1025-C, and pECL1050-B in this study.
Analysis of the plasmids and genetic environment of blaNDM-1
The genetic structures of pECL1025-C and pECL1050-B were highly homologous, and both belong to the IncX3 type. The blaNDM-1 gene, accompanied by bleMBL (bleomycin resistance gene), was located downstream of an ISAba125 element. This ISAba125 was disrupted by the insertion of IS5; furthermore, an IS3000 and a truncated Tn2 element were found upstream of the element. They also harbored blaSHV-12, which was flanked by two IS26 elements. However, pECL1051-A—an IncHI2 plasmid approximately five times the size of pECL1050-B—does not carry blaSHV-12. Its downstream structure varied greatly, and there was no Tn2 upstream. For specific details, refer to Figure 4b.
Homology analysis comparison
MLST analysis identified all strains as sequence type ST97. Core-genome MLST (cg-MLST) further resolved their genetic relatedness (Figure 5a), revealing 9 and 3,824 allelic differences between CR-1050/CR-1051 and CR-1050/CR-1025, respectively. Based on homologous gene clusters (Figure 5b), the three strains shared 4,745 core gene families, while harboring 3 (CR-1025), 28 (CR-1050), and 9 (CR-1051) unique families. These results demonstrate a high overall genetic similarity among the E. hormaechei strains, with CR-1050 and CR-1051 being most closely related.
Results of cg-MLST and homologous gene analysis. (a). Minimum spanning tree based on the core-genome MLST. The allelic distance between strains was represented by numbers. (b). Homologous gene analysis results (Venn). Each colored ellipse in the figure represents a strain; the numbers in the intersecting parts among strains indicate the number of common gene families, and the numbers in the individual parts of each strain indicate the number of unique gene families of that strain.
Discussion
bla_NDM-1_ and mcr-9 have been detected in various bacteria worldwide (Hem et al., 2024; Li et al., 2023; Cahill et al., 2023). However, few reports have performed a detailed genomic analysis of CR-E. hormaechei with blaNDM-1 and mcr-9, simultaneously. In this study, three CR-E. hormaechei strains carrying both blaNDM-1 and mcr-9 were isolated from three patients with varying degrees of burns.
mcr-9 has been identified in several CRE backgrounds since it was described in 2019 (Macesic et al., 2021), and its prevalence in clinical isolates from Asia and Europe has increased annually suggesting the potential for a global outbreak. mcr-9 is located on IncHI2 plasmids (Chen et al., 2022), consistent with the findings of this study. pECL-1025 plasmid A, pECL-1050 plasmid A and pECL-1051 plasmid A share a similar mcr-9 gene background, the conserved region encodes nickel-cobalt efflux transporters (rcnR) and sensory protein kinases (pcoE). Sequence alignment showed that there was a similar structure to the reference plasmid p123 (Accession no. CP099763) from a Salmonella enterica in 2023 from Belgium and plasmid p1 (Accession no. OW849257) from a Citrobacter freundii in 2022 from Spain. This indicates that the abundance of mobile genetic elements endows the gene structure in the mcr-9 region with high dynamism and plasticity, providing favorable mobility and dissemination conditions for the mcr-9 gene. Similarly, the mcr-9-like structure is not only identified in the Enterobacter cloacae complex (ECC) but also detected in other species such as Salmonella and Citrobacter. This observation further highlights the high risk of mcr-9 gene transfer among different plasmids or across the genomes of distinct bacterial species. This underscores the urgent need for close monitoring to determine the detection rate of mcr-9 and for effective action to control its further spread.
The blaNDM-1 gene is commonly found on high-frequency transfer plasmids such as IncX3 and IncC, the IS5 transposon has made a significant contribution to its dissemination (Uz Zaman et al., 2018), and a 100% conjugation success rate further validates this conclusion. The emergence of blaNDM-1 is one of the primary drivers of carbapenem-resistant Enterobacteriaceae (CR-ECC). The co-expression of aminoglycoside resistance, trimethoprim resistance, sulfonamide resistance, quinolone resistance, and β-lactamase genes confers a multidrug-resistant (MDR) phenotype on the strain, exerting pressure on clinical antibiotic selection. Interestingly, in the present study, blaCTX-M-9 and mcr-9, as well as blaSHV-12 and blaNDM-1, exhibit a strong association respectively. blaCTX-M-9 has previously been identified as the most prevalent extended-spectrum β-lactamase (ESBL) gene among hospital-acquired CRE in China (Zhou et al., 2018). blaSHV-12 has also been frequently detected in the Enterobacter cloacae complex (ECC) (Houkes et al., 2025). This indicates that genetic elements can form “antibiotic resistance gene clusters” through homologous recombination or co-integration events. The simultaneous transmission of such clusters across strains and species would constitute a significant public health threat.
Many Enterobacter hormaechei strains are characterized by high antimicrobial resistance but low virulence (Davin-Regli et al., 2019). In this study, the virulence genes carried by E. hormaechei include acrB, T4SS/T6SS, and rcsb. These genes are associated with bacterial colonization and efflux systems, enabling the bacteria to develop resistance to cytotoxic substances and thereby reduce the impact of antibiotics on them. Although these virulence factors are located on chromosomes and have limited transmission rates compared to mobile elements such as plasmids, their synergistic effects on drug resistance could potentially enable E. hormaechei to possess both high drug resistance and high virulence simultaneously. The emergence of such strains would significantly increase treatment challenges. Therefore, the potential pathogenicity risk of ECC also warrants attention.
This study reports the first detection of CR-ECC with ST97 suggests that this type is likely the predominant clonal group in this region. Furthermore, all three patients received treatment in the burn unit, indicating a probable nosocomial outbreak within that department. However, this requires further investigation incorporating temporal and epidemiological data.
Conclusions
In summary, we first report the genomic characteristics of three ST97 clinical E. hormaechei strains carrying both bla_NDM-1_ and mcr-9. The CR-E. hormaechei strains have a broad drug resistance spectrum, posing a challenge to clinical treatment. Whether it is blaNDM-1 or mcr-9, if no measures are taken to control them, they will be spread widely and significantly, in a matter of time. We should conduct routine genomic surveillance to effectively curb the spread of drug-resistant bacteria in the region.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cahill N. Hooban B. Fitzhenry K. Joyce A. O’Connor L. Miliotis G. . (2023). First reported detection of the mobile colistin resistance genes, mcr-8 and mcr-9, in the Irish environment. Sci. Total. Environ. 876, 162649. doi: 10.1016/j.scitotenv.2023.162649, PMID: 36906027 · doi ↗ · pubmed ↗
- 2Cai S. Quan J. Wang Z. Hu H. Han X. Jiang Y. . (2024). High prevalence of carbapenem-resistant Enterobacter cloacae complex in a tertiary hospital over a decade. Microbiol. Spectr. 12, e 0078024. doi: 10.1128/spectrum.00780-24, PMID: 39475294 PMC 11619405 · doi ↗ · pubmed ↗
- 3Carroll L. M. Gaballa A. Guldimann C. Sullivan G. Henderson L. O. Wiedmann M. (2019). Identification of Novel Mobilized Colistin Resistance Gene mcr-9 in a Multidrug-Resistant, Colistin-Susceptible Salmonella enterica Serotype Typhimurium Isolate. m Bio 10, e 00853–e 00819. doi: 10.1128/m Bio.00853-19, PMID: 31064835 PMC 6509194 · doi ↗ · pubmed ↗
- 4Chen C. Xu H. Liu R. Hu X. Han J. Wu L. . (2022). Emergence of neonatal sepsis caused by MCR-9- and NDM-1-co-producing enterobacter hormaechei in China. Front. Cell. Infect. Microbiol. 12. doi: 10.3389/fcimb.2022.879409, PMID: 35601097 PMC 9120612 · doi ↗ · pubmed ↗
- 5Davin-Regli A. Lavigne J.-P. Pagès J.-M. (2019). Enterobacter spp.: update on taxonomy, clinical aspects, and emerging antimicrobial resistance. Clin. Microbiol. Rev. 32, e 00002–e 00019. doi: 10.1128/cmr.00002-19, PMID: 31315895 PMC 6750132 · doi ↗ · pubmed ↗
- 6Halder G. Chaudhury B. N. Denny P. Chakraborty M. Mandal S. Dutta S. (2025). Emergence of concurrently transmissible mcr-9 and carbapenemase genes in bloodborne colistin-resistant Enterobacter cloacae complex isolated from ICU patients in Kolkata, India. Microbiol. Spectr. 13, e 0154224. doi: 10.1128/spectrum.01542-24, PMID: 39912656 PMC 11878022 · doi ↗ · pubmed ↗
- 7Hem S. Cummins M. L. Wyrsch E. R. Drigo B. Hoye B. J. Maute K. . (2024). Genomic analysis of Citrobacter from Australian wastewater and silver gulls reveals novel sequence types carrying critically important antibiotic resistance genes. Sci. Total. Environ. 909, 168608. doi: 10.1016/j.scitotenv.2023.168608, PMID: 37977387 · doi ↗ · pubmed ↗
- 8Houkes K. M. G. Weterings V. Verweij J. J. Murk J. L. van den Bijllaardt W. Stohr J. J. J. M. (2025). Molecular characterization and prevalence of plasmids co-harbouring mcr and ESBL genes. Microb. Genom. 11, 1458. doi: 10.1099/mgen.0.001458, PMID: 40728876 PMC 12306943 · doi ↗ · pubmed ↗