The Effect of Increased Temperature on Dengue Virus in the Vector Aedes aegypti from New Caledonia
Méryl Delrieu, Olivia O’Connor, Nicolas Pocquet, Kenny Teraiharoa, Anne-Fleur Griffon, Christophe Menkes, Morgan Mangeas, Elvina Viennet, Valérie Burtet-Sarramegna, Myrielle Dupont-Rouzeyrol, Francesca D. Frentiu

TL;DR
This study examines how increased temperature affects dengue virus in Aedes aegypti mosquitoes from New Caledonia.
Contribution
The study reveals that higher temperatures increase viral loads in mosquitoes without affecting transmission rates.
Findings
DENV titres in mosquito bodies and heads were significantly higher at 31.1 °C than 26.6 °C.
No significant differences were observed in infection, dissemination, and transmission rates between the two temperatures.
Elevated temperature increases viral loads but not the proportion of infectious mosquitoes.
Abstract
Dengue virus (DENV) is a major public health concern in tropical and subtropical regions, including the Pacific. Temperature is recognised as a major driver of transmission under climate change. Understanding how higher temperatures may alter DENV transmission is essential to anticipate future dengue risk. Therefore, we assessed the effect of temperature on DENV-1 in Aedes aegypti from New Caledonia. Mosquitoes were orally infected and maintained for 14 days at 26.6 °C (average temperatures during recent outbreaks) or 31.1 °C (SSP5-8.5 scenario projected temperatures). Mosquito bodies, heads, and saliva were analysed separately to determine infection, dissemination, and transmission rates as well as transmission efficiencies. Infectious virus was detected by using a fluorescent focus assay, and viral titres were quantified via TCID50 assays. No significant differences were observed in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
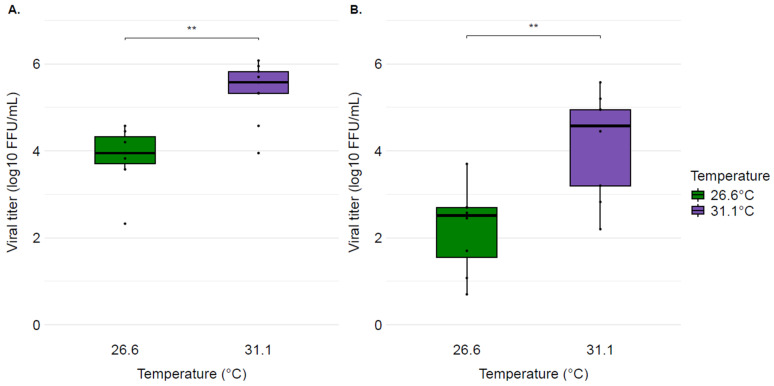 Figure 1
Figure 1- —PACIFIC FUND
- —Prix d’Encouragement à la Recherche
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMosquito-borne diseases and control · Malaria Research and Control · Biological Research and Disease Studies
1. Introduction
Rising temperatures are one of the most noticeable effects of climate change and contribute to the resurgence of vector-borne diseases, including dengue fever, chikungunya, West Nile fever, and Zika, by influencing both vector biology and viral dynamics [1,2,3,4]. Dengue virus (DENV) is one of the most important arboviruses worldwide, with 390 million estimated cases annually [5], and is primarily transmitted by the mosquito Aedes aegypti. Modelling studies predict a geographical expansion of vector ranges and longer seasons of vector-borne pathogen circulation, leading to an overall increased risk of transmission [6,7,8,9]. Virus infection is favoured under higher temperatures, as in vivo experiments show that exposure to warmer temperatures often leads to elevated mosquito infection rates and a shortened extrinsic incubation period [10]. Island territories are particularly vulnerable to the effects of climate change, yet they remain underrepresented in experimental studies exploring its impact on infectious diseases. New Caledonia, a tropical island in the Pacific Ocean, has a long history of DENV circulation [11,12]. However, over the past 15 years, a shift in epidemiological patterns has been observed with the persistence of DENV-1 circulation, the co-circulation of multiple DENV serotypes and other arboviruses, and an increase in the severity of dengue disease [12,13,14,15]. In 2018, a Wolbachia biocontrol strategy was implemented through the World Mosquito Program in four cities of New Caledonia. This intervention led to a decline in DENV cases, with 22 reported for 2025 in New Caledonia, including 9 local cases (https://dass.gouv.nc/votre-sante-maladies/la-dengue-le-chikungunya-et-le-zika; accessed on 7 February 2026) compared to 572 cases between January and April 2018 (https://dass.gouv.nc/sites/default/files/atoms/files/graphedenguedu01092017au06042018.pdf; accessed on 7 February 2026). A recent modelling study predicts an increased risk of DENV epidemics and longer transmission periods in New Caledonia due to rising temperatures, with annual outbreaks anticipated under the SSP5-8.5 scenario, assuming no Wolbachia-based vector control strategies [8]. Among the five climate scenarios proposed by the Intergovernmental Panel on Climate Change (IPCC), SSP5-8.5 represents a worst-case trajectory with increases reaching +4.5 °C (between +3.3 °C and +5.7 °C) by 2100 [16]. Here, we investigated the impact of this scenario on DENV infection in Ae. aegypti mosquitoes from New Caledonia by contrasting present-day mean temperatures with projected end-of-century conditions under SSP5-8.5. Two temperature conditions were assessed: 26.6 °C, which is the current average temperature during recent DENV outbreaks [17], and 31.1 °C, which represents the SSP5-8.5 scenario, with a 4.5 °C increase over the current temperature.
2. Materials and Methods
Seven- to nine-day-old female Ae. aegypti mosquitoes originating from field collections during 2022 in Païta, New Caledonia (F3-F5 generation), were used in experimental infections and confirmed to be free of Wolbachia using established qPCR methods previously described [18,19]. Mosquitoes were deprived of sugar overnight before being fed for 30 min on an infectious blood meal composed of DENV-1 diluted in bovine blood, with a final concentration of 1 × 10^7^ FFU/mL. The DENV-1 genotype I strain used in this study (GenBank: MW315195.1) was isolated from human serum during the 2017 outbreak in New Caledonia [20], and viral stock was produced on Vero E6 cells (ATCC, Manassas, VA, USA). After blood feeding using a Hemotek system (Hemotek Ltd., Blackburn, UK), fully engorged females were incubated for 14 days at 26.6 °C or 31.1 °C, representing present-day and projected future temperatures, respectively, with 80% relative humidity. At 14 days post infection (dpi), 28 and 30 mosquitoes were sampled from the 26.6 °C and 31.1 °C regimes, respectively, to determine infection, dissemination, and transmission (see below for methods). Mosquitoes were anesthetized with 7% CO_2_ and induced to salivate for 30 min into 5 µL of FBS (Foetal Bovine Serum, Gibco, ThermoFisher Scientific, Waltham, MA, USA). Each saliva sample was then added to 45 µL of DMEM (Gibco, ThermoFisher Scientific, Waltham, MA, USA). Mosquitoes were dissected, with heads and bodies stored separately in 350 µL of DMEM (Dulbecco’s Modified Eagle Medium; Gibco, Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 2% FBS and 1% Antibiotic–Antimycotic (Gibco, ThermoFisher Scientific, Waltham, MA, USA). Saliva, heads and bodies were stored at −80 °C, with heads and bodies mechanically ground up with glass beads in a BeadBeater (BioSpec Products Inc., Bartlesville, OK, USA) at the time of analysis [15].
The presence of infectious virus in mosquito bodies, heads, and saliva (the last as a proxy for transmission) was assessed by using a fluorescent focus assay (FFA) as described below. Diluted homogenates (20-fold dilutions of ground-up body and head samples) or undiluted saliva were inoculated on Vero E6 cell monolayers and overlaid after two hours in DMEM supplemented with 5% FBS and 3.2% carboxymethylcellulose (Sigma-Aldrich, Merck, St. Louis, MO, USA). Cells were then maintained for 5 days at 37 °C under 5% CO_2_. Cells were fixed, permeabilized [21], and stained using an anti-dengue complex monoclonal antibody (clone D3-2H2-9-21, Chemicon^®^ Merck Millipore, Burlington, MA, USA) followed by Alexa Fluor 488 goat anti-mouse IgG (Invitrogen, ThermoFisher Scientific, Waltham, MA, USA). The presence of infectious virus was determined by visualising DENV foci under a fluorescent microscope (ZEISS Axiovert 5 TL FL SCB, Carl Zeiss Microscopy GmbH, Jena, Germany) at 100× magnification [15]. The infection rate was calculated as the proportion of mosquitoes with infectious virus detected in the body. The dissemination rate was the proportion of infected mosquitoes showing virus in the head, and the transmission rate was the proportion of mosquitoes with disseminated infection, confirmed with virus detected in the saliva. The transmission efficiency was calculated as the proportion of total tested mosquitoes with virus in the saliva. Viral titres in the bodies and heads of infected mosquitoes were quantified by using TCID_50_ assays with serial 10-fold dilutions inoculated on Vero E6 cell monolayers, incubated for 5 days, and stained and visualised as for FFAs. The presence/absence of infectious virus was scored for each well, and TCID_50_/mL values were calculated using the Spearman–Kärber method and converted to FFU/mL [22].
Pairwise comparisons of infection rate, dissemination rate, transmission rate, and transmission efficiencies between temperatures were performed using Fisher’s exact tests. Viral titres were tested for normality using the Shapiro–Wilk test. Given the small sample sizes, datasets were analysed using non-parametric methods. Overall differences between groups were assessed with Wilcoxon rank sum tests. Data were visualized as boxplots showing medians and interquartile ranges, with analyses performed in R software version 4.5.1 [23].
3. Results
Infection, dissemination, and transmission rates and efficiencies are summarized in Table 1.
Infection rates ranged from 28.6% to 30% for the mosquitoes placed at 26.6 °C and 31.1 °C, respectively. At these temperatures, viral dissemination was observed in all infected mosquitoes (100%). The transmission rates of mosquitoes from 26.6 °C and 31.1 °C were 12.5% and 66.7%, respectively. At 26.6 °C, the transmission efficiency was 3.6%, and at 31.1 °C, the transmission efficiency was 20%. Fisher’s exact tests showed no statistically significant differences in infection rate, dissemination rate, transmission rate, and transmission efficiencies between temperatures.
The median viral titres of infected mosquitoes’ bodies from the 26.6 °C and 31.1 °C regimes were 8.89 × 10^3^ FFU/mL and 3.75 × 10^5^ FFU/mL, respectively (Figure 1A), and the difference was statistically significant (p = 0.004). The median viral titres of infected mosquito heads placed at 26.6 °C and 31.1 °C were 3.28 × 10^2^ FFU/mL and 3.75 × 10^4^ FFU/mL, respectively (Figure 1B). Similarly to bodies, head titres from mosquitoes at 31.1 °C were significantly higher than those at 26.6 °C (p = 0.006).
4. Discussion
In our study, we observed that DENV-1 infection rates were around 30%. Although few mosquitoes were infected, they all disseminated the virus. These observations are in concordance with previous studies from New Caledonia using the standard temperature (28 °C) for mosquito vector competence experiments. Indeed, Ae. aegypti populations orally infected with DENV-1 displayed low-to-moderate infection rates but high dissemination rates reaching 100% [15,24]. These observations suggest that, once midgut infection is established in our local Ae. aegypti, there appears to be no restriction to viral dissemination.
In our experiments, infection, dissemination, transmission rates and dissemination and transmission efficiencies did not significantly differ between 26.6 °C and 31.1 °C. This was somewhat unexpected, as the majority of studies reported an increase in infection, dissemination, and transmission of the tested virus at higher temperatures, as reviewed here [10]. However, some studies have found no clear differences even when using temperature ranges similar to those reported in the studies mentioned above. Limited variation in DENV infection and transmission was observed in Florida populations of Ae. aegypti and Aedes albopictus [25], while others have observed that transmission potential by Ae. albopictus did not systematically increase across all tested temperature conditions [26]. Our findings provide additional useful data, but larger sample sizes will be needed in future studies to confirm these trends.
In our study, infection rates were observed to be low to moderate. While such values are not uncommon, they complicate the interpretation of our results. However, similar patterns have been observed in New Caledonia at 28 °C for DENV-1 and depending on the mosquito population [15,24]. Studies from other regions of the globe have reported highly variable outcomes, ranging from very low values in Kenya and Senegal [27,28] to much higher values in Cuba, Brazil, Ghana, and Japan [29,30,31,32]. Such variability highlights the influence of mosquito population, viral strain, and geographic context. Vector competence may also be shaped by virus–vector compatibilities [33,34,35] and further modulated by environmental factors such as temperature [36,37] or biological factors like the mosquito gut microbiome [38,39,40].
Temperature appears to enhance viral loads in mosquitoes, as significantly higher DENV titres were observed at 31.1 °C than at 26.6 °C. Another study reported that viral genome levels in the bodies and salivary gland tissues of Ae. aegypti were generally higher at 28–34 °C, though significant differences appeared only at certain dpi and did not affect infection rates [41]. More recently, Ae. aegypti mosquitoes infected by DENV-2 or co-infected by DENV-2 and Mayaro virus and maintained at 32 °C displayed significantly higher viral titres than mosquitoes held at 27 °C [42]. In parallel, in vitro experiments using mosquito and mammalian cell lines also demonstrated that DENV replication was enhanced at 32 °C, with faster viral kinetics and higher titres than at 27 °C [41]. These studies reinforce the idea that temperature influences not only viral loads and transmission but also modulates the molecular and physiological response of mosquitoes to infection [43,44]. At the molecular level, high temperatures enhance the activity of viral RNA-dependent enzymes and may destabilize viral RNA structures, while simultaneously affecting host physiology by accelerating metabolism and potentially weakening innate immune defences [45,46]. In addition, temperature has been shown to modulate tissue barriers such as the midgut escape barrier. Importantly, the impact of temperature on this barrier is highly context-dependent, varying according to mosquito population, viral strains, and the thermal conditions experienced during larval and adult stages [47].
Beyond these experimental insights, the impact of temperature on DENV transmission has also been demonstrated at the population level through climate-based models. In New Caledonia, a climate-based statistical model using dengue case data has shown that when daily maximum temperatures exceed 32 °C for 10 consecutive days or more during January to March, the risk of dengue outbreaks increases [48]. Global warming is expected to increase average temperatures worldwide, along with heatwaves that are projected to become more frequent, stronger, and longer. Such conditions may increase the risk of transmission, especially in regions already impacted by dengue. In our study, although infection and dissemination rates did not differ, viral titres were significantly higher at the warmer temperature. These observations support the idea that elevated temperatures can amplify viral replication in mosquitoes and could potentially contribute to increased DENV transmission risk, although a mechanistic link remains unclear. Understanding how increased temperature and heat waves affect mosquito–virus interactions will be important to anticipate future transmission risks and improve mechanistic models for outbreak prediction under climate change.
A major limitation of this study was the very low number of positive saliva samples (n = 1 at 26.6 °C and n = 6 at 31.1 °C), which prevented reliable statistical analysis of transmission potential and limited the interpretation of transmission dynamics. The higher number of positive saliva samples at 31.1 °C suggests that higher viral loads in bodies and heads increase the potential for DENV to reach the salivary glands and, in turn, result in more mosquitoes being infectious. However, this hypothesis needs further testing. A further limitation is that transmission was assessed at a single time point (14 dpi), which provides only a partial view of this complex process. Additionally, at 26.6 °C, viral titres were obtained for seven individuals, as one body sample could not be titrated due to insufficient material, resulting in a slightly reduced number of data points for viral load analysis at this temperature. Nevertheless, analysis of viral titres shows that higher temperatures promoted higher viral replication of DENV, underscoring the importance of further investigating how temperatures influence viral kinetics of DENV and its transmission. While our study was unable to explore the specific viral or host factors contributing to elevated DENV titres, this remains a valuable direction for future research. It should also be noted that our study did not include Wolbachia mosquitoes, despite the use of Wolbachia-based strategies for dengue control in New Caledonia. This choice was deliberate, as we aimed to establish baseline data on temperature effects in wild-type Ae. aegypti from New Caledonia. In addition, our study was conducted at constant temperatures, which does not reflect field conditions where mosquitoes experience daily temperature variation. Realistic field scenarios need to be considered in future studies by applying fluctuating temperature regimes.
5. Conclusions
In conclusion, we explored the effect of temperature on DENV-1 infection in Ae. aegypti from New Caledonia, the primary vector on this island. Despite low-to-moderate infection rates, all infected mosquitoes showed viral dissemination, with significantly higher viral titres at 31.1 °C than at 26.6 °C, indicating that higher temperature can enhance viral amplification in mosquitoes. Further studies are needed to investigate how elevated temperatures influence viral replication over time and the mosquito’s potential for virus transmission and assess the possibility of DENV variant evolution under heat stress. Given that several countries in the Pacific region are currently experiencing DENV outbreaks, these insights will be critical for anticipating the broader consequences of climate change on DENV evolution and epidemiology.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Anikeeva O. Hansen A. Varghese B. Borg M. Zhang Y. Xiang J. Bi P. The impact of increasing temperatures due to climate change on infectious diseases BMJ 2024387 e 07934310.1136/bmj-2024-07934339366706 · doi ↗ · pubmed ↗
- 2Dennington N.L. Grossman M.K. Ware-Gilmore F. Teeple J.L. Johnson L.R. Shocket M.S. Mc Graw E.A. Thomas M.B. Phenotypic adaptation to temperature in the mosquito vector, Aedes aegypti Glob. Change Biol.202430 e 1704110.1111/gcb.1704138273521 · doi ↗ · pubmed ↗
- 3Reinhold J.M. Lazzari C.R. Lahondère C. Effects of the environmental temperature on Aedes aegypti and Aedes albopictus mosquitoes: A review Insects 2018915810.3390/insects 904015830404142 PMC 6316560 · doi ↗ · pubmed ↗
- 4Semenza J.C. Rocklöv J. Ebi K.L. Climate change and cascading risks from infectious disease Infect. Dis. Ther.2022111371139010.1007/s 40121-022-00647-335585385 PMC 9334478 · doi ↗ · pubmed ↗
- 5Bhatt S. Gething P.W. Brady O.J. Messina J.P. Farlow A.W. Moyes C.L. Drake J.M. Brownstein J.S. Hoen A.G. Sankoh O. The global distribution and burden of dengue Nature 201349650450710.1038/nature 1206023563266 PMC 3651993 · doi ↗ · pubmed ↗
- 6Mordecai E.A. Cohen J.M. Evans M.V. Gudapati P. Johnson L.R. Lippi C.A. Miazgowicz K. Murdock C.C. Rohr J.R. Ryan S.J. Detecting the impact of temperature on transmission of Zika, dengue, and chikungunya using mechanistic models P Lo S Neglected Trop. Dis.201711 e 000556810.1371/journal.pntd.0005568 PMC 542369428448507 · doi ↗ · pubmed ↗
- 7Ngonghala C.N. Ryan S.J. Tesla B. Demakovsky L.R. Mordecai E.A. Murdock C.C. Bonds M.H. Effects of changes in temperature on Zika dynamics and control J. R. Soc. Interface 2021182021016510.1098/rsif.2021.016533947225 PMC 8097513 · doi ↗ · pubmed ↗
- 8Ochida N. Mangeas M. Dupont-Rouzeyrol M. Dutheil C. Forfait C. Peltier A. Descloux E. Menkes C. Modeling present and future climate risk of dengue outbreak, a case study in New Caledonia Environ. Health 2022212010.1186/s 12940-022-00829-z 35057822 PMC 8772089 · doi ↗ · pubmed ↗