Social Determinants and Outbreak Dynamics of the 2025 Measles Epidemic in Mexico: A Nationwide Analysis of Linked Surveillance Data
Judith Carolina De Arcos-Jiménez, Pedro Martínez-Ayala, Oscar Francisco Fernández-Diaz, Sergio Sánchez-Enríquez, Patricia Noemi Vargas-Becerra, Ana María López-Yáñez, Roberto Damian-Negrete, Sofía Gutierrez-Perez, Jaime Briseno-Ramírez

TL;DR
The 2025 measles outbreak in Mexico was driven by social factors and low vaccination rates in marginalized communities, leading to severe outcomes among indigenous populations.
Contribution
This study integrates nationwide surveillance data with social determinants to reveal how measles spreads through vulnerable populations and identifies actionable vaccination pathways.
Findings
Measles cases were concentrated in Chihuahua, with 47 hot-spot municipalities accounting for 64.4% of cases.
Molecular surveillance identified two virus strains, with the majority linked to the North American outbreak.
Late outbreak phase and being under one year old were significant risk factors for complications.
Abstract
Measles resurgence threatens elimination achievements in the Americas. We conducted a nationwide analysis of Mexico’s 2025–2026 measles outbreak, integrating individual-level surveillance data from the Special Surveillance System for Febrile Exanthematous Diseases with municipal-level social determinants from eight national databases, complemented by molecular surveillance data. We analyzed 6892 confirmed cases using spatial autocorrelation (Moran’s I and LISA), effective reproduction number estimation, logistic regression models for municipal case presence, and multivariable logistic regression for risk factors for complications. Cases concentrated in Chihuahua (65.2%), with 47 LISA hot-spot municipalities containing 64.4% of cases. Molecular surveillance confirmed two independent introductions: D8/MVs/Ontario.CAN/47.24 (98.1%), linked to the North American outbreak, and B3 (1.9%) in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
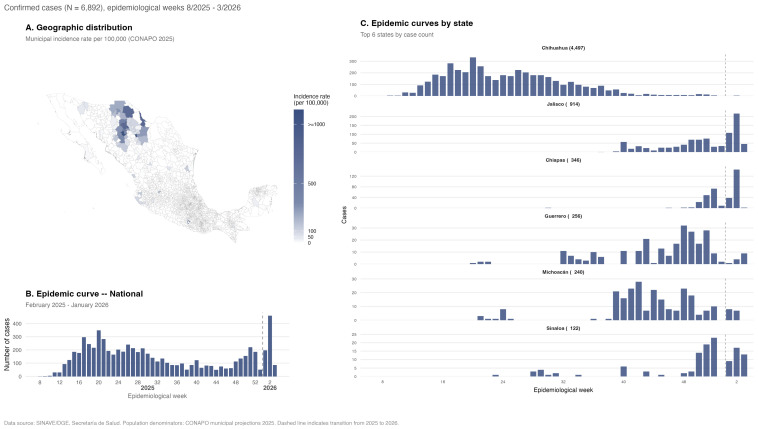 Figure 1
Figure 1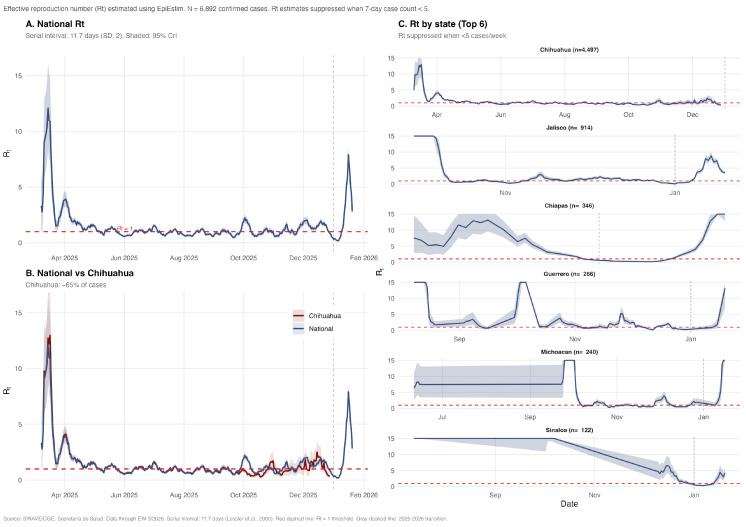 Figure 2
Figure 2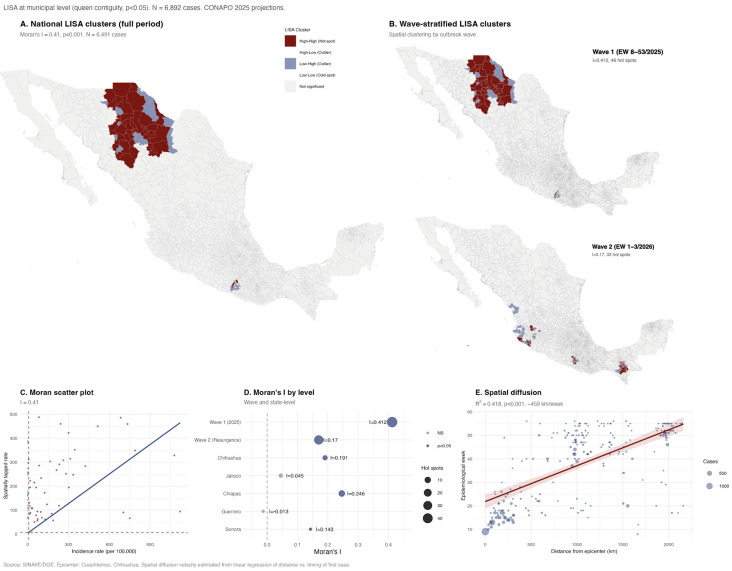 Figure 3
Figure 3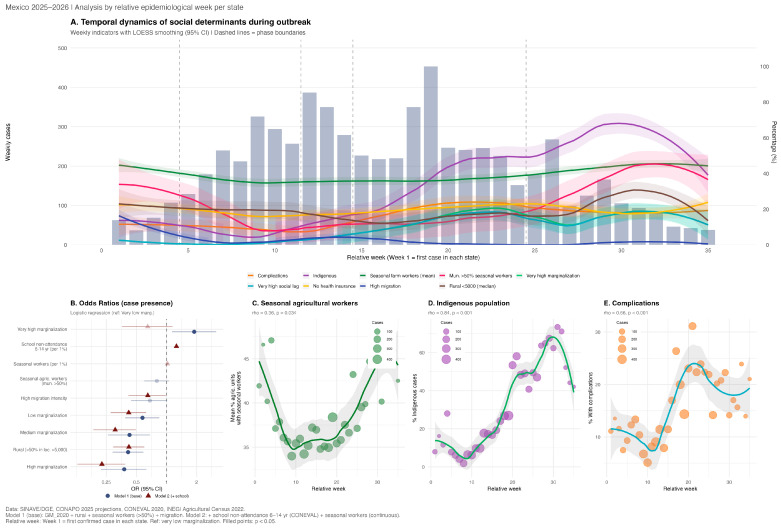 Figure 4
Figure 4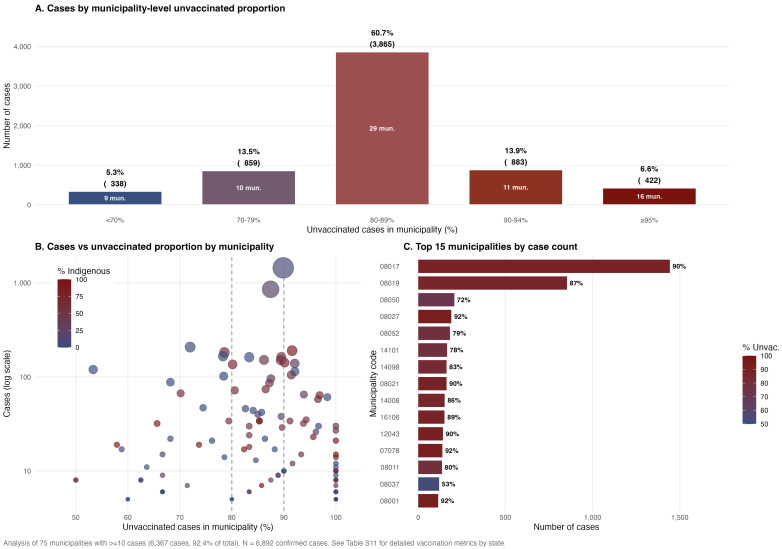 Figure 5
Figure 5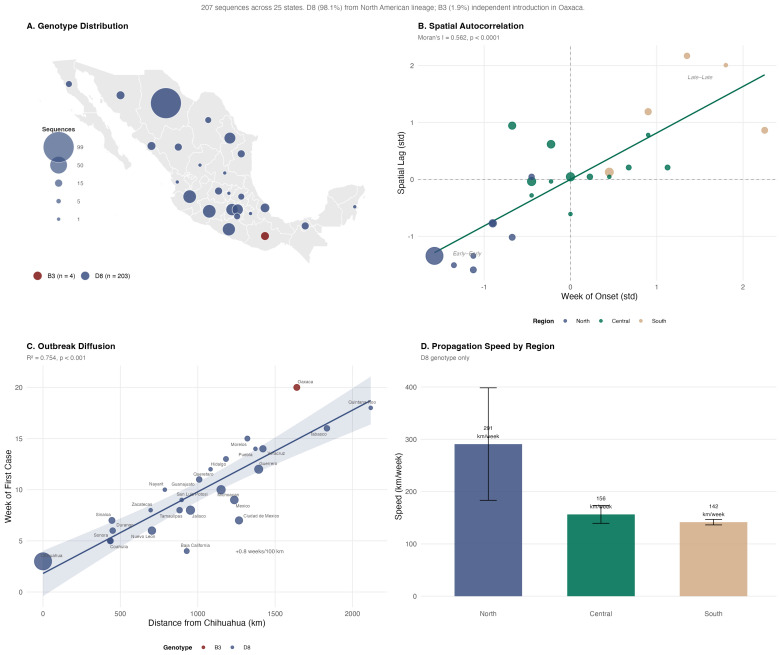 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVirology and Viral Diseases · Vaccine Coverage and Hesitancy · Mosquito-borne diseases and control
1. Introduction
Measles remains a significant global public health concern due to its high transmissibility and disease burden, particularly among children in low- and middle-income countries [1,2]. Despite a highly effective vaccine available for over five decades, periodic resurgences persist due to immunization gaps and health system disruptions [3,4].
In 2024, 395,521 laboratory-confirmed cases were reported globally, with 16,147 additional cases in early 2025—likely underestimating true burden due to surveillance limitations [1,5]. Over half required hospitalization, reflecting severe complications, including pneumonia and encephalitis, among unvaccinated and immunocompromised individuals [1,5]. Children under 5 years face the highest burden, with mortality amplified by measles-induced immune amnesia that increases susceptibility to other infections for months to years post-recovery [6,7,8]. Large outbreaks incur substantial economic costs—projected at US$90 million for 2025—while vaccination programs yield an estimated 58:1 return on investment through averted healthcare expenditures and productivity losses [9,10].
Global measles incidence and mortality declined over 90% between 1990 and 2021 through widespread vaccination [5]. Nevertheless, in 2021, measles caused approximately 4.1 million cases, 48,100 deaths, and 4.2 million disability-adjusted life years among children aged <5 years [5]. Burden remains disproportionately concentrated in regions with low sociodemographic indices and suboptimal coverage, such as sub-Saharan Africa and South Asia [5,11,12].
With a basic reproduction number (R_0_) of 12–18, measles requires ≥95% two-dose coverage to prevent outbreaks [3,13]. However, global first-dose (MCV1) coverage stagnated at 83% in 2022–2023—exacerbated by COVID-19 disruptions—with only 64% coverage in low-income countries [1,5,14]. These gaps fueled major outbreaks in 2019 and ongoing crises in 2024–2025, including Europe’s highest case counts in over 25 years and a large outbreak across Texas and New Mexico driven by undervaccinated communities [1,15,16,17].
In the Americas, 18 countries experienced outbreaks between 2019–2023, temporarily disrupting elimination status in two nations; however, no country had endemic transmission by end-2023 [18,19]. Mexico interrupted endemic transmission in 1997 through high coverage and robust surveillance, with subsequent cases primarily importation-driven [20,21].
In post-elimination settings, resurgence is increasingly driven by subnational “pockets of susceptibility”—geographic and social clusters where structural determinants (inequitable access, rurality, marginalization, and mobility networks) concentrate low immunity despite high national coverage [22,23,24,25]. This study provides a comprehensive analysis of Mexico’s 2025 measles epidemic, integrating individual-level surveillance and immunization registry data with municipality-level sociodemographic indicators. We characterize outbreak dynamics, identify high-risk populations, elucidate social determinants driving resurgence, and generate evidence to guide targeted interventions in post-elimination settings.
2. Materials and Methods
2.1. Study Design and Setting
We conducted a population-based retrospective observational study of Mexico’s 2025–2026 measles epidemic from 19 February 2025 to 18 January 2026 (epidemiological weeks 8–53 of 2025 and weeks 1–3 of 2026). The outbreak included an initial wave (February–December 2025) and an early resurgence in January 2026. Analyses were performed at the individual (confirmed cases) and ecological (municipal) levels across all 32 states and 2469 municipalities.
2.2. Data Sources
We integrated eight national databases spanning epidemiological surveillance, hospital discharges, socioeconomic marginalization, migration, agricultural labor, and vaccination coverage. Additional analytic variables were derived a priori based on biological plausibility, including state-relative outbreak week, epidemic phase, binary indicators for high-marginalization and high-migration municipalities, and municipal vaccination gap estimates (data dictionary in Supplementary Table S1).
2.2.1. Primary Data Source: Epidemiological Surveillance
Case-level data were obtained from Mexico’s Special Surveillance System for Febrile Exanthematous Diseases (EFE), maintained by the General Directorate of Epidemiology (DGE) [26]. This open-access database includes 24 clinical–epidemiological variables per case (demographics, clinical manifestations, vaccination and indigenous status, and diagnostic classification) updated weekly through epidemiological week 3 of 2026. Cases were defined per Mexican Official Standard NOM-017-SSA2-2012 [27] and InDRE laboratory guidelines [28] as: (a) laboratory-confirmed (measles-specific IgM or RT-PCR positive) or (b) clinically–epidemiologically confirmed (compatible illness epidemiologically linked to a laboratory-confirmed case). Only confirmed cases were analyzed; suspected, discarded, and rubella cases were excluded.
2.2.2. Hospital Discharge Data
Hospitalization records (ICD-10 code B05) were obtained from the Automated Hospital Discharge Subsystem (SAEH), maintained by the General Directorate of Health Information (DGIS) [29], capturing diagnosis codes, complications, length of stay, and discharge status from Ministry of Health facilities.
2.2.3. Sociodemographic and Marginalization Data
Municipal-level socioeconomic data were obtained from complementary sources: Marginalization Index 2020 (CONAPO) [30]: composite index incorporating nine social deprivation indicators (illiteracy, incomplete basic education, inadequate housing, overcrowding, income below minimum wage, and rurality); Social Lag Index 2020 (CONEVAL) [31]: municipal-level index based on education, health access, housing quality, and basic services; Migration Intensity Index 2020 (CONAPO) [32]: based on households with U.S. emigrants, remittances, circular migrants, and return migrants, characterizing international migration activity; and Indigenous population: proportion self-identifying as indigenous from EFE individual records and CONAPO estimates [26,30].
2.2.4. Agricultural Census Data
To identify municipalities with high agricultural migrant worker activity, we analyzed the 2022 Agricultural Census (INEGI) [33], using variables on agricultural labor force and seasonal worker presence.
2.2.5. Vaccination Coverage Data
Historical vaccination coverage estimates by state and year, including first and second MMR doses, were obtained from the National Center for Child and Adolescent Health (CENSIA) BIO-SIS database (1990–2023; latest state-level annual data were available at time of analysis) [34]. BIO-SIS database coverage data are administrative estimates from CENSIA and subject to the inherent limitations of this data source.
2.2.6. Molecular Surveillance Data
Measles virus genomic sequencing data were obtained from GenBank (NCBI) [35], yielding 26 historical N-450 nucleoprotein gene sequences from Mexico (2003–2021) and from Epidemiological Bulletins (DGE) [36], providing 2025 outbreak data aggregated by state (number of sequences, genotype, and lineage) through epidemiological week 48 of 2025.
2.2.7. Population Denominators
Population denominators by municipality and age group were obtained from CONAPO mid-year 2025 projections [37] for calculating age- and municipality-specific incidence rates throughout the 2025–2026 outbreak period.
2.3. Data Linkage
All databases were linked at the municipal level using the official five-digit INEGI geographic code (clave geoestadística), consisting of two-digit state and three-digit municipality identifiers. This standardized coding enabled deterministic linkage across all sources. Complete socioeconomic indicators were available for 99.5% of municipalities (n = 2457).
2.4. Statistical Analysis
2.4.1. Descriptive Analysis
We characterized the epidemic through temporal (epidemic curves by epidemiological week), geographic (state and municipality distribution), and demographic (age, sex, indigenous status, and vaccination status) dimensions. Incidence rates were calculated per 100,000 population using CONAPO 2025 projections as denominators: case fatality rates as deaths divided by confirmed cases [37].
Epidemic phase classification. Two temporal frameworks were employed: (1) absolute epidemiological weeks for national-level summaries and (2) state-relative weeks (from each state’s index case) for social determinants analysis. Relative phases were: introduction (weeks 1–4), growth (weeks 5–11), peak (weeks 12–14), decline (weeks 15–24), and late (weeks 25+). Cases from the January 2026 resurgence (weeks 54–56) were classified separately as “Resurgence (Wave 2)” to distinguish them from the primary outbreak. National absolute phases were: Introduction (weeks 8–16), Peak (weeks 17–28), Decline (weeks 29–40), Late (weeks 41–53), and Resurgence (weeks 54–56).
2.4.2. Transmission Dynamics
The time-varying effective reproduction number (Rt) was estimated using Bayesian inference with a gamma-distributed serial interval (mean 11.7 days, SD 2.0 days) derived from measles contact-tracing studies [38]. Rt was calculated with a 7-day sliding window, reporting posterior means and 95% credible intervals (CrI) at each time point. National Rt was estimated from aggregated daily case counts, and state-level Rt was computed for the six highest-burden states (Chihuahua, Jalisco, Chiapas, Guerrero, Michoacán, Sinaloa). Sustained transmission was defined as Rt > 1. Estimates were obtained using the EpiEstim R package, which implements a Bayesian framework to infer the instantaneous reproduction number from incidence data while incorporating the serial interval distribution and its uncertainty [39,40]. Interpretation follows standard thresholds: Rt > 1 indicates exponential growth, Rt = 1 stable transmission, and Rt < 1 declining transmission. Additional methodological details are provided in the Supplementary Materials (Supplementary Note S1).
2.4.3. Spatial Analysis
Spatial autocorrelation of measles incidence was assessed at the municipal level (n = 2457) using exploratory spatial data analysis. Incidence rates per 100,000 were calculated using CONAPO 2025 population projections [37]. A spatial weights matrix was constructed using queen contiguity with row-standardized weights. Global spatial autocorrelation was evaluated using Moran’s I statistic [41], with significance assessed through conditional permutation (p < 0.05).
Local Indicators of Spatial Association (LISA) were calculated using local Moran’s I [42] to identify significant clusters (p < 0.05): hot spots (High-High: high incidence surrounded by high incidence), cold spots (Low-Low), and spatial outliers (High-Low or Low-High). To capture changes in clustering across epidemic phases, LISA was estimated separately for Wave 1 (2025) and Wave 2 (January 2026), and within individual states with ≥100 cases to detect intra-state patterns not visible at the national scale. To assess temporal–spatial spread, we calculated the Euclidean distance from each affected municipality to the outbreak epicenter (Cuauhtémoc, Chihuahua) and examined its relationship with case detection timing using linear regression. The epicenter was defined as the municipality reporting the earliest sustained case cluster. A detailed description of the mathematical formulation, spatial weight specification, and multi-level analytical approach is provided in the Supplementary Materials [43,44] (Supplementary Figure S1).
2.4.4. Social Determinants and Introduction Mechanism
We conducted a municipal-level ecological analysis comparing 266 municipalities with confirmed cases versus 2203 without cases, linking municipal indicators (by residence municipality) from four national sources: the CONAPO Marginalization Index 2020 (0–100; degree categories) [30], CONAPO Migration Intensity Index 2020 (null to very high) [32], CONEVAL Social Lag Index 2020 (including proportion without health insurance) [31], and INEGI 2022 Agricultural Census (percentage employing seasonal workers) [33]. These indicators reflect municipal context rather than individual attributes. Bivariate comparisons used Mann–Whitney U tests for continuous variables and chi-squared tests for categorical variables. Multivariable analyses used negative binomial regression of case counts with log(population) as an offset, reporting incidence rate ratios (IRRs) with 95% CIs. Analyses were stratified by (1) outbreak phase using state-relative weeks (introduction, 1–4; growth, 5–11; peak, 12–14; decline, 15–24; late, 25+; resurgence, January 2026), (2) the six states with ≥50 cases (95.5% of total), and (3) a sensitivity comparison of the first vs. last 50 affected municipalities chronologically. Temporal trends were assessed using Spearman rank correlation and LOESS smoothing (span = 0.4) with 95% CIs to visualize non-linear patterns.
2.4.5. Vaccination and Case Incidence
Vaccine effectiveness (VE) was estimated using the Farrington screening method [45]: VE = 1 − [(PCV × (1 − PPV))/(PPV × (1 − PCV))], and VE = 1 − [(PCV × (1 − PPV))/(PPV × (1 − PCV))], where PCV is the proportion of cases vaccinated (EFE data), and PPV is the population proportion vaccinated (mean MCV1 coverage 2014–2023, CENSIA/CNI). Ninety-five percent CIs were calculated using the Orenstein method [46,47]. State-level VE was estimated for states with ≥20 cases using state-specific coverage. Additional metrics included relative risk (RR) and population attributable fraction (PAF). To characterize focal transmission, municipalities with ≥10 cases were classified by the proportion of unvaccinated cases (<70%, 70–79%, 80–89%, 90–94%, and ≥95%) to identify “pockets of susceptibles” where transmission concentrated. Further details on VE estimation and interpretation in outbreak settings are provided in the Supplementary Materials (Supplementary Figure S2), consistent with prior outbreak investigations and immunization guidelines [48,49,50,51].
2.4.6. Complications and Risk Factors for Severity
Risk factors for complications (pneumonia, otitis media, encephalitis, and others) were assessed using logistic regression. Individual-level covariates included age group (<1, 1–4, 5–19, and ≥20 years), sex, vaccination status, and indigenous self-identification. Municipal-level contextual variables included marginalization degree; rurality (>50% of the population living in localities of <5000); and access to piped water, drainage, and health services (CONAPO 2020 [30] and CONEVAL 2020 [31]). All plausible predictors were screened in bivariable analyses; variables with p < 0.20 were considered for multivariable modeling [52,53]. The final model prioritized parsimony, control of collinearity (VIF < 5), and model fit (AIC, pseudo-R^2^). Results are reported as crude (OR) and adjusted odds ratios (aOR) with 95% CIs. Due to the low number of deaths (n = 25), mortality analyses were descriptive only.
2.4.7. Molecular Epidemiology
Molecular analysis was based on 207 sequences; data for weeks 49–56 (through January 2026) were unavailable. Spatial autocorrelation of the week of symptom onset was assessed using Moran’s I with a k = 4 nearest-neighbor spatial weight matrix. Linear regression fitted the distance from the epicenter (Chihuahua) as the predictor and the week of first case as the outcome to estimate propagation speed. Distances were calculated using the Haversine formula, with speed estimated as a distance/time ratio, stratified by region (North, Central, and South). States with non-predominant genotypes were evaluated separately to identify independent introductions.
2.4.8. Software and Reproducibility
All analyses used R version 4.4.1. Data management and visualization: tidyverse (v2.0.0), ggplot2 (v4.0.1), and dplyr (v1.1.4); spatial analysis: sf (v1.0.23) and spdep (v1.3.13); Rt estimation: EpiEstim (v2.2.5); additional packages: broom (v1.0.9), gtsummary (v2.3.0), flextable (v0.9.9), and patchwork (v1.3.2). Scripts are available in a reproducible pipeline with centralized configurations, enabling re-execution with updated data, upon request to the corresponding author.
3. Results
3.1. Descriptive Analysis
A total of 6892 confirmed measles cases (6432 laboratory-confirmed; 460 epidemiologically linked) were reported across all 32 states during epidemiological weeks 8–53/2025 and 1–3/2026, with marked geographic concentration in northern Mexico (Figure 1).
Chihuahua accounted for 65.2% of cases (n = 4497). Cases in Chihuahua were older (median 20 years, IQR 4–31 vs. 12 years, IQR 4–24; p < 0.001), with adults of 20–39 years comprising 41.5% of the population versus 27.8% in other states (Table 1). Overall, 85.5% of cases were unvaccinated, and 29.1% were indigenous. Import-related transmission predominated in Chihuahua (78.2% vs. 20.0% in other states; p < 0.001). Complications occurred in 15.5% of cases, significantly higher in Chihuahua (19.1% vs. 8.7%; p < 0.001). Twenty-five deaths were recorded (CFR 0.4%), predominantly in Chihuahua (n = 23; p = 0.009).
Cases with complications (n = 1069, 15.5%) were younger (median 4 years, IQR 1–18 vs. 19 years; p < 0.001), with children <5 years accounting for 51.4% of complications (Supplementary Table S2). Indigenous individuals represented 51.3% of complicated cases versus 25.0% of uncomplicated cases (p < 0.001) (Table S3).
Five outbreak phases were identified: Introduction (weeks 8–16, n = 647), Peak (17–28, n = 2770), Decline (29–40, n = 1398), Late (41–53, n = 1336), and Resurgence (54–56, n = 741) (Supplementary Table S4). The most notable temporal shift was in indigenous status, rising from 0.8% during Introduction to 54.6% during Decline (p < 0.001). Complication rates peaked during Decline (25.1%), and deaths occurred primarily during Peak (n = 10) and Decline (n = 12, CFR 0.9%).
3.2. Transmission Dynamics
The national effective reproduction number peaked at 12.1 (95% CrI: 8.5–16.4) on 15 March 2025 during the early growth phase, indicating intense initial transmission (Figure 2A). Rt declined progressively, crossing below 1 on 8 May, approximately 8 weeks after the peak. National Rt remained below 1 from May through December 2025, consistent with controlled transmission during the Decline and Late phases. In Chihuahua (~65% of cases), the Rt trajectory closely paralleled national estimates, peaking at 13.0 on 17 March (Figure 2B). Chihuahua’s Rt first declined below 1 on 6 May, and the last reliable estimate was 0.38 (28 December 2025), indicating controlled transmission in the epicenter by year-end.
State-level Rt estimates showed distinct temporal patterns reflecting heterogeneous outbreak timing (Figure 2C). States with a later epidemic onset (Jalisco, Chiapas, Guerrero, and Michoacan) exhibited elevated Rt values during their respective introduction phases, with wider credible intervals due to smaller case counts. By January 2026, the national Rt had risen to 2.81, driven by the resurgence wave predominantly affecting states outside Chihuahua. Table S5 reports state-specific exact values (peak Rt, peak dates, number of days with Rt > 1, and last status) and the extended methodology for reference.
3.3. Spatial Analysis
Measles incidence showed strong national spatial autocorrelation (Moran’s I = 0.41, z = 35.3; p < 0.001), confirming marked geographic clustering (Figure 3A). LISA identified 47 hot-spot municipalities—predominantly in Chihuahua—accounting for 4441 cases, with highest incidence in Sierra Tarahumara areas characterized by large indigenous populations and limited healthcare access. Spatial clustering shifted across epidemic phases (Figure 3B): Wave 1 (2025) was highly localized (I = 0.412; 46 hot spots, mostly in Chihuahua), whereas during the Wave 2 resurgence (January 2026), clustering weakened (I = 0.17) and dispersed across multiple states (33 hot spots) (Supplementary Table S6). State-level analyses revealed significant intra-state clustering in Chiapas, Chihuahua, and Sonora, while Jalisco and Guerrero showed no spatial autocorrelation, consistent with metropolitan rather than contiguous spread (Figure 3D; Supplementary Figure S1). Distance from the outbreak epicenter (Cuauhtémoc, Chihuahua) was strongly associated with the timing of first case arrival (R^2^ = 0.418; p < 0.001), supporting radial diffusion at approximately 459 km/week through population mobility networks (Figure 3E).
3.4. Social Determinants and Introduction Mechanism
Municipalities with confirmed measles cases (n = 266, 10.4%) differed significantly from those without cases across multiple social indicators (Table 2). Affected municipalities had lower rurality (median 19.3% vs. 48.8%; p < 0.001) and a bimodal marginalization distribution—with overrepresentation of both very low and very high categories. In a base logistic regression for case presence (Model 1, Figure 4B), only very high marginalization was positively associated with having cases (OR 1.89, 95% CI 1.13–3.14; p = 0.014), while low-, medium-, and high-marginalization categories were less likely affected than the very low (urban) reference (OR 0.38–0.58; all p < 0.01), and rural municipalities showed lower odds (OR 0.41, 95% CI 0.29–0.59; p < 0.001). When school non-attendance among children aged 6–14 years (CONEVAL) was added to the model (Model 2), it emerged as the strongest predictor of municipal case presence (OR 1.26 per 1% increase, 95% CI 1.21–1.31; p < 0.001), while very high marginalization lost significance (OR 0.65; p = 0.14), suggesting that vaccination gaps in communities where children do not attend school mediate the marginalization effect (Table S7). Model 2 showed a substantially improved fit (AIC 1380 vs. 1522; AUC 0.795 vs. 0.716). The inverse association with migration intensity (OR 0.65, 95% CI 0.42–0.98; p = 0.049) reflects that the CONAPO index measures emigration to the USA—distinct from the cross-border agricultural worker mobility that introduced measles into northern Mexico.
Temporal analysis by outbreak phase revealed pronounced shifts in the social profile of cases (Table 3; Figure 4). Early transmission (weeks 1–4; n = 274) was concentrated in periurban areas, with municipalities comprising high seasonal agricultural workers (40.4%) and with fewer indigenous cases (17.9%) and complications (10.2%). By the late phase (weeks ≥ 25; n = 1080), spread had shifted to remote, marginalized communities, with indigenous cases rising to 65.8%, complications to 23.1%, and median age decreasing from 22 to 11 years, indicating involvement of younger, unvaccinated cohorts (Figure 4C–E). The share of cases from high agricultural seasonal worker municipalities followed a U-shaped pattern—declining to 7.1% at peak and rebounding to 29.9% at late outbreak—suggesting seasonal re-activation of agricultural labor networks.
State-level stratification (Table S8) confirmed heterogeneous social profiles: Chihuahua (4497 cases; 73.4% average vaccine coverage) was characterized by periurban transmission, while Guerrero (256 cases) and Chiapas (346 cases) showed predominantly indigenous and rural case profiles. Sensitivity analysis comparing the first and last 50 municipalities by temporal sequence (Table S9) reinforced the introduction-to-dispersion gradient: early-affected municipalities had an older median age, lower indigenous proportion, and lower seasonal worker concentration than late-affected municipalities. State-level vaccine coverage (2019–2023 average) was lower in states with ≥100 confirmed cases (mean, 80.8%) compared to states with fewer or no cases (mean 86.5%), though Chihuahua exhibited the lowest coverage among all affected states (73.4%), consistent with long-standing vaccination gaps (Table S8).
3.5. Vaccination and Vaccine Effectiveness (VE)
Among 6892 confirmed cases, 5895 (85.5%) were unvaccinated. National vaccine effectiveness estimated by the Farrington screening method (Supplementary Figure S2) was 98.1% (95% CI: 98.0–98.2%), with a PCV of 14.5% and population vaccination coverage (PPV) of approximately 90% (Supplementary Figure S2B, Table S10). VE ranged from 93.5% in Jalisco to 99.1% in Guerrero among states with ≥50 cases (Table S9). The relative risk for unvaccinated individuals was 53.2, and the population attributable fraction was 97.8%, indicating that an estimated 6741 of 6892 cases were attributable to lack of vaccination. Municipality-level analysis revealed pronounced pockets of susceptibles: among 75 municipalities with ≥10 cases (representing 92.4% of all cases), 60.7% of cases (3865) concentrated in 29 municipalities where 80–89% of cases were unvaccinated, and an additional 20.5% in municipalities with ≥90% unvaccinated (Figure 5A). The two highest-burden municipalities (Cuauhtemoc and Guachochi, Chihuahua) each exceeded 1000 cases with 87–90% unvaccinated, and municipalities with the highest indigenous population proportions showed the most extreme vaccination gaps (Figure 5B,C).
State-level MCV1 coverage (2014–2023 average) showed no significant correlation with 2025 incidence rates (Spearman rho = −0.13; p = 0.46), consistent with the focal, sub-state nature of the outbreak. Chihuahua had the lowest mean historical coverage (77.6%), with those 15 to 34 years below 80% and a 2023 coverage of only 65.6%—far below the 95% herd immunity threshold (Supplementary Figure S2A). Proportions of vaccinated cases were lowest among infants < 1 year (5.4%), reflecting a pre-vaccination age, and highest among ages 1–4 (16.4%) and 20–39 years (16.2%), suggesting immunity gaps in these cohorts (Supplementary Figure S2C). Birth-cohort analysis revealed case accumulation across cohorts born between 1993 and 2022, with peaks among children born in 2020–2022 (ages 3–5) and adults born in 1993–2000 (ages 25–32), corresponding to periods of lower national MCV1 coverage in the late 1990s (Supplementary Figure S2D). The absence of a correlation between cohort-level coverage and case counts (rho = 0.17; p = 0.36) underscores that susceptibility accumulated through multiple mechanisms beyond routine immunization failures, including missed booster doses and waning immunity in adults.
3.6. Complications and Risk Factors for Severity
Overall, 1069 of 6892 cases (15.5%) developed complications. In the bivariable analysis, 26 of 28 variables tested were associated with complications at p < 0.20 (Table S12). The multivariable model (Table 4) identified six independent risk factors. Age < 1 year carried the highest risk (aOR 3.36, 95% CI 2.72–4.15), followed by ages 1–4 (aOR 2.58, 2.14–3.13), while adults ≥ 20 years had lower risk (aOR 0.64, 0.53–0.77) compared to the 5–19 reference group. Indigenous status (aOR 1.89, 1.61–2.22), lack of vaccination (aOR 1.96, 1.53–2.51), late outbreak phase (aOR 1.68, 1.42–2.00), and rural municipality of residence (aOR 1.73, 1.48–2.03) were all significantly associated with complications (all p < 0.001). The resurgence wave (January 2026) was not independently associated with increased complications (aOR 0.81, 0.60–1.09; p = 0.164), despite the overall lower complication rate (8.5% vs. 16.4% in Wave 1), suggesting that observed differences reflect changes in the geographic and demographic composition of cases rather than a secular trend.
Complication rates showed stark gradients: rural (27.5%) vs. urban (12.3%) municipalities, indigenous (27.4%) vs. non-indigenous (10.7%) cases, and late-phase (22.5%) vs. early-phase (9.2%) cases. These gradients converge geographically—as the outbreak migrated from urban periurban areas into remote indigenous communities during late phases (Section 3.4), the cumulative burden of complications concentrated in the most marginalized populations.
Hospital Discharge Analysis
Linkage with the Automated Hospital Discharge Subsystem (SAEH) identified 663 measles-coded hospitalizations in Ministry of Health facilities during February–November 2025 (Table S13). Among hospitalized patients, 308 (46.5%) had documented complications, predominantly pneumonia (n = 270, 87.7% of complications), followed by other complications (n = 18, 5.8%), encephalitis (n = 10, 3.2%), and otitis media (n = 3, 1.0%). Pneumonia was most frequent among children aged 5–9 years (49.4%) and 1–4 years (47.9%). Median length of stay was 4 days (IQR: 2–6). Indigenous patients had significantly higher complication rates (50.3% vs. 41.6%; p = 0.033) and longer hospitalizations (p < 0.001). Six in-hospital deaths occurred (case fatality rate among hospitalized: 0.9%), with three (50%) associated with pneumonia. Twenty-five total deaths were recorded during the epidemic (overall CFR: 0.36%).
3.7. Molecular Epidemiology
Genomic surveillance based on 207 sequences across 25 states (Table S14) identified two co-circulating genotypes. Genotype D8 (lineage MVs/Ontario.CAN/47.24) predominated with 203 sequences (98.1%) in 24 states, while B3 (lineage MVs/New South Wales.AUS/10.24) comprised 4 sequences (1.9%) restricted to Oaxaca, indicating at least two independent introductions. Spatial analysis of onset timing demonstrated significant autocorrelation (Moran’s I = 0.562; p < 0.0001), consistent with contagious diffusion from the Chihuahua epicenter (Figure 6A,B). Distance from Chihuahua explained 75.4% of the variance in the week of first case arrival (R^2^ = 0.754; p < 0.001), with each 100 km adding approximately 0.80 weeks (5.6 days) to the arrival time (Figure 6C). The mean D8 propagation speed was 194.8 km/week (27.8 km/day), being fastest in the North (290.9 km/week), intermediate in the Center (156.5 km/week), and slowest in the South (141.6 km/week) (Figure 6D). Oaxaca’s B3 genotype arrived 5.1 weeks later than predicted by the D8 diffusion model (observed: week 20 vs. predicted: week 14.9), confirming an independent introduction unrelated to the northern epicenter. Historically, only 26 measles sequences from Mexico were deposited in GenBank (2003–2021), representing sporadic importations of genotypes D4, D8, D9, and H1 (Supplementary Figure S3). The 2025 outbreak represents an unprecedented scale of measles genomic detection in the country.
4. Discussion
This nationwide analysis of 6892 confirmed measles cases concerns Mexico’s largest epidemic since interruption of endemic transmission in 1997 and provides comprehensive insight into the outbreak dynamics, social determinants, and molecular origins of this resurgence [20]. Six principal findings emerge. First, the outbreak was highly focal: Chihuahua accounted for 65.2% of cases, and 47 LISA hot-spot municipalities concentrated 64.4% of the national burden. Second, molecular surveillance confirmed two independent introductions—a predominant D8 genotype (98.1%, lineage MVs/Ontario.CAN/47.24) linked to the 2024–2025 North American outbreak, and a separate B3 genotype (1.9%) imported into Oaxaca from Australia. Third, transmission followed a three-stage pattern: introduction through seasonal agricultural worker networks, amplification in undervaccinated communities, and subsequent diffusion toward marginalized indigenous populations. Fourth, a dual-model analysis revealed that school non-attendance among children aged 6–14 years mediated the effect of very high marginalization on municipal case presence, identifying a previously unreported and potentially actionable pathway for targeted vaccination. Fifth, vaccine effectiveness remained high (98.1%), confirming that the outbreak was driven by accumulation of susceptible individuals rather than vaccine failure. Sixth, independent risk factors for complications—age of <1 year (aOR 3.36), age of 1–4 years (aOR 2.58), unvaccinated status (aOR 1.96), indigenous status (aOR 1.89), rural residence (aOR 1.73), and late outbreak phase (aOR 1.68)—converged geographically in the most marginalized communities reached last by the epidemic.
The predominance of D8/MVs/Ontario.CAN/47.24 places Mexico’s outbreak within the broader 2024–2025 North American measles resurgence. The lineage was first characterized in Canada (Ontario, epidemiological week 47 of 2024), but the same genotype circulated across at least eight countries, linked by Mennonite community travel networks [54]. In the United States, 2267 confirmed measles cases were reported in 2025—the highest annual total since elimination was declared in 2000—including a Southwest US outbreak of 933 cases across Texas (646), New Mexico (65), and neighboring states, with three deaths [55,56,57,58]. Mathematical modeling of the Texas–New Mexico cluster estimated effective reproduction numbers of 1.65–3.82 depending on model structure (exponential growth, SIR, or SEIR), with basic reproduction numbers of 30–40 [17]. Our peak national Rt of 12.1 (95% CrI: 8.5–16.4) is not directly comparable, as it represents instantaneous Bayesian estimations (EpiEstim), capturing peak transmission intensity, whereas the Texas–New Mexico estimates reflect growth-phase averages from mechanistic models; nevertheless, both outbreaks confirm above-threshold transmission in undervaccinated communities. Given Chihuahua’s shared border with Texas and New Mexico, and the well-documented cross-border mobility of Mennonite communities between these regions, transmission through the US–Mexico border represents a plausible and likely route of introduction—consistent with, rather than contradicting, the Canadian lineage designation [59]. Notably, the high proportion of import-related cases in Chihuahua (78.2% vs. 20.0% in other states) reflects this extensive inter-municipal and cross-border mobility, as SINAVE classifies any infection acquired outside the municipality of residence as “imported,” encompassing inter-municipal, inter-state, and international movement. Indeed, the temporal and geographic overlap of the Gaines County, Texas, outbreak (January–August 2025) with Mexico’s epidemic (February 2025 onward) and the shared D8 genotype across both countries support a connected North American transmission chain. Collectively, these outbreaks contributed to the loss of measles elimination status in the Region of the Americas, declared by PAHO in November 2025 after Canada documented >12 months of sustained transmission [60]. In January 2026, PAHO invited both the United States and Mexico for a formal review of their individual elimination status [61], underscoring the regional dimension of this resurgence [18].
Spatial analysis at two complementary scales provided evidence of contagious diffusion. At the municipal level, incidence showed strong spatial autocorrelation (Moran’s I = 0.41; p < 0.001), with distance from the Cuauhtémoc epicenter explaining 42% of the variance in timing of first case arrival at approximately 459 km/week through population mobility networks. At the state level, molecular surveillance showed even stronger spatial structuring (R^2^ = 0.754), with a propagation speed of 194.8 km/week, reflecting the broader North-to-South diffusion pattern, consistent with contagious rather than strongly hierarchical diffusion [62]. Wave-stratified LISA analysis revealed a transition from concentrated clustering during Wave 1 (I = 0.412, 46 hot spots in Chihuahua) to more dispersed patterns during the January 2026 resurgence (I = 0.17, 33 hot spots across multiple states), consistent with epidemic maturation from focal introduction to wider geographic dissemination.
A novel finding is the identification of school non-attendance as a mediator of the relationship between very high marginalization and municipal case presence. In the base model, very high marginalization was the only category positively associated with case presence (OR 1.89; p = 0.014), with a bimodal pattern reflecting simultaneous vulnerability in urban hubs (very low marginalization) and remote indigenous communities. When school non-attendance among children aged 6–14 years was added, it emerged as the strongest predictor (OR 1.26 per 1% increase; p < 0.001), while very high marginalization lost significance (OR 0.65; p = 0.14), and model fit improved substantially (AIC 1380 vs. 1522; AUC 0.795 vs. 0.716; Table S7). This mediation pattern suggests that the excess risk in very highly marginalized municipalities operates, at least partly, through children not enrolled in or attending school—and therefore not exposed to school-based vaccination campaigns and coverage verification. These findings extend evidence that multiple deprivations drive suboptimal childhood vaccination in Latin America [22] by identifying a specific, actionable indicator that could prioritize municipalities for supplementary vaccination activities.
The progressive concentration of cases among indigenous populations—from 0.8% during introduction to 54.6% during the decline phase (p < 0.001)—mirrors patterns observed in Ecuador, where measles odds were fourfold higher in parishes with larger indigenous populations [22]. PAHO reported that most measles deaths in Mexico’s 2025 outbreak occurred among indigenous individuals [63], consistent with our finding that indigenous persons comprised 29.1% of cases but 76% of the 25 deaths. The convergence of indigenous status, young age, lack of vaccination, rural residence, and late outbreak phase in the multivariable model underscores that complications concentrated where the epidemic arrived last: remote communities with multiple structural barriers to vaccination and timely healthcare [56,64]. Indigenous patients in the SAEH hospital discharge data experienced higher complication rates (50.3% vs. 41.6%; p = 0.033) and longer hospitalizations, reinforcing surveillance-based findings across an independent data source.
Vaccine effectiveness of 98.1% (95% CI: 98.0–98.2%) is consistent with expected values for measles-containing vaccines [3] and with estimates from recent outbreaks in the United States [58]. The population attributable fraction of 97.8%—indicating that an estimated 6741 of 6892 cases were attributable to lack of vaccination—underscores that this was an outbreak of access, not of vaccine failure. The absence of correlation between state-level vaccination coverage and incidence (rho = −0.13; p = 0.46) reflects the focal nature of transmission in “pockets of susceptibles”, where 81.2% of cases in municipalities with ≥10 cases occurred in settings where ≥80% of cases were unvaccinated (Figure 5). This pattern has been documented in US outbreaks where city-wide or state-wide coverage exceeded 90%, but sub-county pockets of undervaccination sustained transmission [11,24,25]. Chihuahua’s chronically low coverage (mean, 77.6%; only 65.6% in 2023; 15 years below 80%) illustrates how sustained coverage deficits create conditions for explosive transmission once the virus is introduced [1].
Wave segmentation—separating the January 2026 resurgence from the 2025 epidemic—revealed that late outbreak phase was independently associated with complications (aOR 1.68, 95% CI: 1.42–2.00; p < 0.001), a finding obscured in models without wave stratification (previously aOR 1.09; p = 0.366). This suggests that as the epidemic migrated from periurban areas into remote communities, affected populations faced compounding disadvantages: younger unvaccinated cohorts, limited healthcare access, and delayed case detection. The resurgence wave was not independently associated with increased complications (aOR 0.81; p = 0.164), and its lower complication rate (8.5% vs. 16.4% in Wave 1) likely reflects shifts in the geographic and demographic composition of cases—predominantly affecting Jalisco, Chiapas, and Sinaloa—rather than a secular trend in virulence. The marked differences in effective reproduction number between Chihuahua (peak Rt, 13.0) and other states also reflect differences in the size and density of susceptible populations: decades of coverage deficits in communities created large susceptible pools sustaining prolonged transmission, whereas later-affected states experienced smaller, more contained chains seeded by imported cases, with Rt declining more rapidly.
This study has several limitations that should be acknowledged. First, vaccination status was self-reported by patients or caregivers and may be subject to recall bias; misclassification could lead to underestimation of vaccine effectiveness if vaccinated cases were incorrectly classified as unvaccinated, and municipal averages may mask important intra-municipal heterogeneity. Second, the screening method for vaccine effectiveness estimation assumes that cases are representative of the population with respect to vaccination status, an assumption that may not hold in outbreak settings where vaccination campaigns modify coverage differentially across populations. Third, our ecological analyses linking individual cases to municipal-level social determinants may be subject to ecological fallacy; municipal averages may mask important intra-municipal heterogeneity, particularly in large or diverse municipalities. Fourth, the cross-sectional design limits causal inference regarding social determinants and transmission; we cannot definitively establish whether marginalization or school non-attendance caused increased transmission, or whether common underlying factors explain both. While statistically supported, this requires prospective confirmation. Fifth, genomic data covered only 207 sequences (~3% of cases); although geographic representation was adequate across 25 states, limited public data availability precluded phylogenetic analysis of intra-state transmission chains. Sixth, the SAEH hospital discharge data cover only Ministry of Health facilities through December 2025, excluding private-sector and social-security hospitals as well as the January 2026 resurgence, and therefore underestimates total hospitalization burden. Finally, complication rates may be underestimated, as the surveillance system captures acute complications but may miss delayed sequelae.
Despite these limitations, this study integrates individual-level surveillance data with municipal-level social determinants from eight national databases, enabling analysis of structural factors at a granularity rarely achieved in outbreak investigations. The dual-scale spatial analysis, wave-stratified LISA clustering, and dual logistic regression models provide methodological approaches transferable to other post-elimination settings. The identification of school non-attendance as a mediator of the marginalization–susceptibility pathway and the documentation of a three-stage transmission model generate evidence with direct policy relevance for outbreak response in Latin America and globally.
Several research directions emerge from our findings. First, prospective seroprevalence studies are needed to accurately quantify population immunity at subnational levels, particularly in communities identified as “pockets of susceptibles” to guide targeted vaccination campaigns. Second, qualitative research exploring vaccine decision-making in Mennonite, indigenous, and agricultural worker communities would inform culturally appropriate interventions. Third, mathematical modeling studies could estimate the vaccination coverage required to prevent future outbreaks in the specific demographic and spatial context of northern Mexico, accounting for population mobility and community structure. Fourth, implementation research should evaluate strategies for reaching mobile populations, including seasonal agricultural workers, with vaccination services. Fifth, enhanced molecular surveillance with real-time whole-genome sequencing could enable more precise tracking of transmission chains and earlier identification of new importations. Sixth, economic analyses quantifying the direct and indirect costs of this outbreak would strengthen the case for investment in elimination maintenance. Finally, cross-border surveillance coordination mechanisms between Mexico, the United States, and Canada warrant systematic evaluation to develop sustainable approaches for the post-elimination era.
5. Conclusions
Mexico’s 2025–2026 measles epidemic—6892 confirmed cases across 266 municipalities in all 32 states—represents the largest resurgence since interruption of endemic transmission and underscores that national coverage averages can mask focal vulnerability. Transmission was driven by accumulation of susceptible individuals in municipal pockets rather than by vaccine failure (VE, 98.1%). Molecular evidence confirmed at least two independent introductions dominated by a D8 lineage shared with the 2024–2025 North American outbreak, highlighting the role of cross-border mobility networks. The epidemic trajectory was socially patterned: introduction compatible with agricultural worker networks, amplification in undervaccinated communities, and progressive concentration among indigenous populations where the burden of complications was disproportionate. A key finding was that school non-attendance among children mediated the effect of very high marginalization on municipal case presence, identifying children not reached by school-based vaccination as a critical target population. Wave segmentation revealed that late outbreak phase was an independent risk factor for complications (aOR, 1.68), highlighting the compounding disadvantages faced by remote communities reached last by the epidemic. Sustaining elimination in Mexico requires shifting from coverage averages to precision public health: identifying municipal susceptibility pockets through linked sociodemographic surveillance, deploying catch-up vaccination targeting school non-attenders and mobile agricultural populations, and strengthening cross-border genomic surveillance in the post-elimination era.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Do L.A.H. Mulholland K. Measles 2025 N. Engl. J. Med.20253932447245810.1056/NEJ Mra 250451640561553 · doi ↗ · pubmed ↗
- 2Stoneman E.K. Measles JAMA 20253341665166610.1001/jama.2025.1421040811055 · doi ↗ · pubmed ↗
- 3Hübschen J.M. Gouandjika-Vasilache I. Dina J. Measles Lancet 202239967869010.1016/S 0140-6736(21)02004-335093206 · doi ↗ · pubmed ↗
- 4Bester J.C. Measles and Measles Vaccination: A Review JAMA Pediatr.20161701209121510.1001/jamapediatrics.2016.178727695849 · doi ↗ · pubmed ↗
- 5Chen W. Du M. Deng J. Liu M. Liu J. Global, Regional, and National Trends of Measles Burden and Its Vaccination Coverage among Children under 5 Years Old: An Updated Systematic Analysis from the Global Burden of Disease Study 2021 Int. J. Infect. Dis.202515610790810.1016/j.ijid.2025.10790840246060 · doi ↗ · pubmed ↗
- 6Peltola H. The History of Measles and Vaccine Development Acta Paediatr.202510.1111/apa.7037041246889 · doi ↗ · pubmed ↗
- 7Laksono B.M. de Vries R.D. Verburgh R.J. Visser E.G. de Jong A. Fraaij P.L.A. Ruijs W.L.M. Nieuwenhuijse D.F. van den Ham H.-J. Koopmans M.P.G. Studies into the Mechanism of Measles-Associated Immune Suppression during a Measles Outbreak in the Netherlands Nat. Commun.20189494410.1038/s 41467-018-07515-030470742 PMC 6251901 · doi ↗ · pubmed ↗
- 8Mina M.J. Kula T. Leng Y. Li M. de Vries R.D. Knip M. Siljander H. Rewers M. Choy D.F. Wilson M.S. Measles Virus Infection Diminishes Preexisting Antibodies That Offer Protection from Other Pathogens Science 201936659960610.1126/science.aay 648531672891 PMC 8590458 · doi ↗ · pubmed ↗