Gene Expression Analysis and Whole Genome Sequencing Reveal the Potential Mechanism of Ciprofloxacin Resistance in a Salmonella Dublin Isolate
Kingsley E. Bentum, Amy Leestemaker-Palmer, Stephanie Nuss, Sophia Ballard, Alexandra Montgomery, Woubit Abebe, Temesgen Samuel, Anthony Pokoo-Aikins, Luiz E. Bemudez

TL;DR
This study explores how a Salmonella Dublin isolate becomes resistant to ciprofloxacin, finding that drug efflux activity may play a key role even without major genetic mutations.
Contribution
The study identifies drug efflux as a potential mechanism of ciprofloxacin resistance in Salmonella Dublin, a serovar where resistance is less commonly reported.
Findings
The ciprofloxacin-resistant Salmonella Dublin isolate showed higher drug efflux activity compared to a susceptible isolate.
An N868S mutation in the GyrA protein was found in the resistant isolate, though it did not alter protein structure significantly.
The resistant isolate had elevated expression of efflux-related genes acrA, acrB, ramA, and soxS.
Abstract
Drug resistance in Salmonella to important antibiotics such as Ciprofloxacin is becoming an increasing public health threat. Ciprofloxacin resistance is, however, well documented in common Salmonella serovars such as Salmonella Typhimurium and Salmonella Enteritidis, while resistance is reported less frequently in isolates like Salmonella Dublin. This study was therefore conducted to investigate potential factors underlying Ciprofloxacin resistance in a Salmonella Dublin isolate. This isolate was identified through antibiotic resistance screening of stored Salmonella bacteria recovered from various samples, using the Kirby-Bauer disk diffusion method, followed by a broth-dilution method. For comparative analysis, the whole genome of this Ciprofloxacin-resistant Salmonella Dublin isolate and another Ciprofloxacin-susceptible isolate from this study cohort were sequenced and screened for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
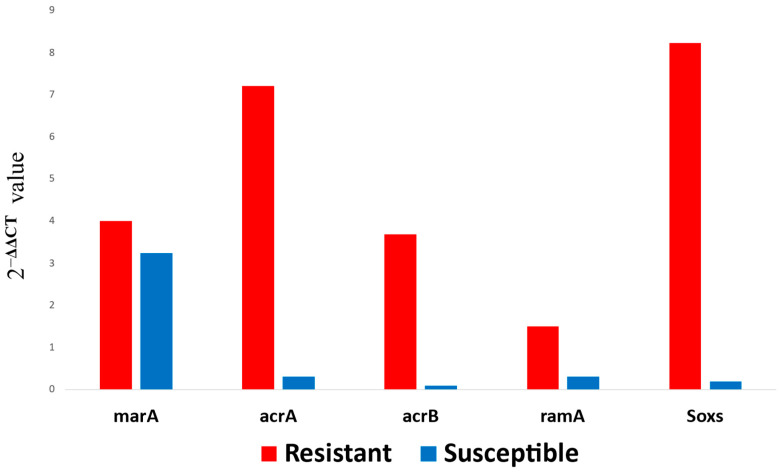 Figure 1
Figure 1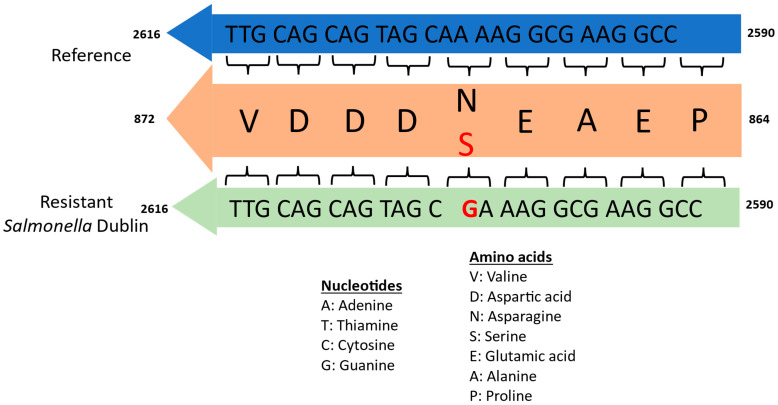 Figure 2
Figure 2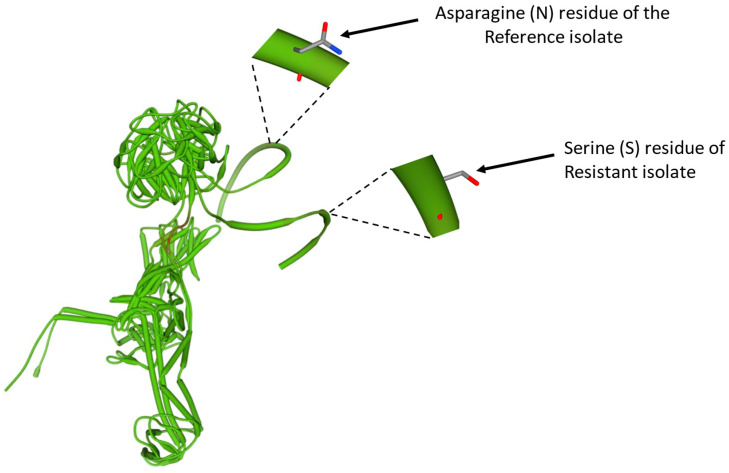 Figure 3
Figure 3- —Carlton College of Veterinary Medicine, Oregon State University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Bacterial Genetics and Biotechnology · Antibiotic Resistance in Bacteria
1. Introduction
Fluoroquinolones (FQs) belong to the second generation of quinolones, and they have enhanced biological activity against microbes [1]. One important member of this class of antibiotics is Ciprofloxacin (CIP), which became clinically available in the mid-1980s [2]. Generally, the enhanced biological activity of FQs allows them to have an extended spectrum against bacteria [3], with drugs like CIP considered the best of treatment options for both invasive and systemic salmonellosis [4].
Sadly, there has been a growing number of quinolone-resistant microbes around the world [3]. Resistance against CIP was practically non-existent when the drug was first introduced [5]. However, the increased use of drugs belonging to the FQ class of antibiotics, especially in livestock production, has greatly contributed to a corresponding high incidence of resistant species against CIP among important pathogens like Salmonella [6]. A typical example was the rise in the proportion of FQ-resistant Salmonella Typhimurium from 1% in 1994 to 12% in 1996 after the drug’s approval for use in veterinary medicine in the U.K. in 1993 [7]. Often, these antibiotics are used among many livestock establishments at sub-inhibitory concentrations for disease prevention or as feed additives [5,8]. This practice has undoubtedly contributed to the development of resistant strains with the potential to infect both humans and animals [9].
Because all FQs have the same mechanism of action, it is easy for resistance to one member of the drug class to be conferred to another [7]. For instance, resistance against enrofloxacin, which is intended for animal use, can translate to resistance against CIP, which is primarily intended for human use. In addition, both drugs are members of the second generation of FQs [1]. The mechanism of action of quinolone drugs against microbial organisms is the inhibition of bacterial DNA replication by blocking the activity of the DNA gyrase and DNA topoisomerase IV enzymes [3]. The gyrA and gyrB genes code for the DNA gyrase enzyme, while the parC and parE genes code for the topoisomerase IV enzyme [3]. Hence, any alterations in the drug targets in these target enzymes within the pathogen can potentially confer resistance against the drug [2].
Since the first report of a CIP-resistant Salmonella in 1990, many other resistant isolates have been identified [10,11,12]. However, compared to the other members of Enterobacteriaceae, CIP-resistance, and for that matter, quinolone-resistance in Salmonella is considered to be an evolving phenomenon [13]. Furthermore, studies discussing quinolone-resistance in less frequent Salmonella isolates, such as Salmonella Dublin, are quite scarce, making it more difficult to assess their public health risk better [14]. This is partly because it is quite uncommon to isolate CIP-resistant Salmonella Dublin. For instance, in a recent study using a global dataset of 1303 Salmonella Dublin genomes, only one isolate was identified to be carrying genetic mutations associated with CIP resistance [15]. Another study involving 74 Salmonella Dublin isolates from ten federal states in Germany reported no CIP resistance except for one strain [16]. Finally, a large collection of Salmonella Dublin isolates collected over decades in Japan had none of the isolates showing resistance to fluoroquinolones (including CIP) [17].
Considering the high morbidity and mortality rates associated with Salmonella Dublin infection [18], and given the potential public health implications of CIP-resistant Salmonella Dublin, this study was conducted to investigate how chromosomal mutations, plasmids, antimicrobial resistance genes, and efflux pumps interplay to contribute to CIP resistance in an identified Salmonella Dublin isolate.
2. Materials and Methods
2.1. Selection of Isolates and Bacterial Culture
A total of 17 Salmonella isolates, biobanked by the Biomedical Research Laboratory at Oregon State University, were selected on 25 June 2023, for initial antibiotic susceptibility screening. The associated metadata for the isolates is presented in Table 1. The isolates were removed from a −80 °C freezer (Thermo Scientific, Asheville, NC, USA). Using a sterile inoculating loop, flakes of the stock cultures of the isolates were then plated on prepared MacConkey agar (BD, Franklin Lakes, NJ, USA) plates. The plates were then incubated at 37 °C for 24 h. Colonies from the stock cultures were used for downstream analysis.
2.2. Antimicrobial Susceptibility Testing
Tryptic Soy Agar culture slants of the 17 isolates were prepared from the revived colonies and sent to the Oregon Veterinary Diagnostic Laboratory on 7 July 2023, for antimicrobial susceptibility testing. All the isolates were tested for their susceptibility to CIP (5 µg) using the Kirby-Bauer disk diffusion method. Disk diffusion zones were measured using the BIOMIC V3 Reader (Giles Scientific, Santa Barbara, CA, USA). For subsequent bacterial challenge with CIP, the susceptibilities of the isolates were also analyzed using the broth dilution method to obtain the minimum inhibitory concentrations (MICs) to determine the appropriate drug exposure concentrations. For both disk diffusion and broth dilution methods, the susceptibility categories and the MIC of the isolates were interpreted according to the established criteria of the Clinical and Laboratory Standards Institute (CLSI) [19].
2.3. Bacteria Challenge with Ciprofloxacin
From the disk diffusion method, CIP-resistant and CIP-susceptible isolates belonging to the same serovar were selected for antibiotic challenge for comparative analysis. The MICs of the selected candidates to CIP were obtained using the broth dilution method, and to also determine the appropriate CIP challenge concentrations for the gene expression analysis. The isolates were tested against various concentrations of CIP (Millipore Sigma, Burlington, MA, USA) prepared using Mueller–Hinton broth (Becton, Dickinson and Company, Franklin Lakes, NJ, USA). Briefly, 900 µL each of CIP concentrations ranging from 250 µg/mL to 0.01 µg/mL, obtained by 2-fold serial dilutions from the highest to the lowest concentration, were prepared. Fresh bacterial cultures grown in Mueller–Hinton broth were centrifuged at 2000 rpm for 10 min. The supernatant was discarded, and the pellet was resuspended in 5 mL of HBSS 1X solution (Gibco Laboratories, Grand Island, NY, USA) to obtain a bacterial stock culture. From the stock culture, a 0.5 McFarland concentration of bacterial culture was prepared using HBSS 1X solution, and 100 μL each of this bacterial culture was inoculated into the various CIP concentrations. The inoculated samples were then incubated for 24 h at 37 °C to determine the broth dilution MICs of our isolates. The next day, to induce a CIP exposure response, rather than a CIP-induced cell death, and to eliminate the possibility of stationary-phase RNA remodeling, 1 mL of freshly prepared 0.5 McFarland concentration of each of the CIP-resistant and CIP-susceptible isolates was inoculated into 9 mL of 1/4 their respective broth dilution MICs for CIP (Supplementary Figure S1). A control experiment was also set up with no CIP supplementation of the media. Both the experimental and control cultures were incubated at 37 °C for 10 h.
2.4. RNA Extraction and Gene Expression Analyses Targeting Efflux Transporter Genes and Regulators
The expression of the acrAB efflux transporter genes and the regulator genes marA, ramA, and soxS was measured in our analysis. Following the incubation of the CIP-challenged and control cultures, the cultures were centrifuged at 14,000 rpm for 10 min, and the supernatant was discarded. Total RNA was then extracted from the culture pellet using the Zymo RNA/DNA Miniprep Kit (Zymo Research, Tustin, CA, USA) according to the manufacturer’s instructions. Residual genomic DNA was removed from the extracted RNA using DNase I Roche (Millipore Sigma, Burlington, MA, USA) following the manufacturer’s protocol. For gene expression analysis, the extracted RNA was first reverse transcribed using the iScript reverse transcription kit (Bio-Rad Laboratories, Hercules, CA, USA) to produce cDNA, which was used as a template for real-time PCR using the qRT–PCR iQ™ SYBR^®^ Green Supermix (Bio-Rad Laboratories, Hercules, CA, USA) according to the manufacturer’s protocol. The qRT–PCR set-ups were conducted in triplicate, and the expression level of each gene was calculated using the average of the three independent set-ups. The primers used for the qRT–PCR are listed in Table 2. The following cycling conditions: an initial denaturation at 95 °C for 3 min, followed by 40 cycles of denaturation and annealing at 95 °C for 10 s and 60 °C for 30 s, respectively, were used as recommended by the manufacturer. Finally, the relative quantities of each expressed gene transcript were measured based on their cycle threshold (CT) values, and the expression of the genes of interest by both treated and untreated cultures was analyzed using the 2^−ΔΔCT^ method [20]. Fold change results were only reported descriptively (with no statistical analysis done) due to prior averaging of the CT values and the small sample size involved.
2.5. Whole Genome Sequencing of CIP-Resistant and CIP-Susceptible Salmonella Isolates
Both the CIP-resistant and CIP-susceptible isolates were freshly cultured on MacConkey agar plates, and DNA was extracted from the cultures for whole-genome sequencing. DNA extraction was done using the Zymo Quick-DNA High Molecular Weight Kit with Magnetic Beads (Zymo Research, Irvine, CA, USA) using modifications for homogenization and lysing, then continued with standard protocols. Careful consideration was taken to reduce mechanical shearing, including minimizing pipetting, to reduce fragmentation as much as possible. Modifications included mixing approximately 10^9^ total bacteria cells in 400 µL of Zymo DNA/RNA Shield buffer and incubating at 37 °C for 4 h, then adding 20 µL of Proteinase K and incubating for an additional hour. The ‘Cultured Bacterial and Fungal Cells’ protocol was then followed from step 4 as listed. Heatless speed-vacuuming was used for 1 h to raise the concentrations of the isolate’s DNA after extraction. Following extraction, genomic DNA libraries were prepared using the Oxford Nanopore Rapid Sequencing Barcoding kit (SQK-RBK-114-24) at 200 ng total input. The respective genomic DNA of the serovars was sequenced using the Oxford Nanopore MinION Mk1D long-read sequencer using a R10.4.1 flow cell and processed using the MinKNOW v5.2.3, with live basecalling performed with the ‘Super-Accurate Basecalling’ model using Dorado with a minimum quality score of 9 (Oxford Nanopore Technologies, Oxford, UK). The raw passing reads were then analyzed with Oxford Nanopore’s Epi2ME v5.2.5 software, using the wf-bacterial-genomes workflow v1.4.2. Generated assembled contigs from the workflow were used for further downstream investigations. Raw reads from both isolates have been deposited in the National Center for Biotechnology Information (NCBI) under the bioproject accession PRJNA1369519.
2.6. Plasmid, Antimicrobial Resistance, and Mutation Analysis
Using ABRicate (version 1.0.1) with a coverage threshold of 95% and an identity threshold of 80%, the resulting assembled contigs were screened against the Comprehensive Antibiotic Resistance Database (CARD) and the PlasmidFinder database, both updated on 17 October 2025. The two databases were used for antimicrobial resistance gene and plasmid detection, respectively. Also, a customized database built from the gyrA, gyrB, parC, and parE genes of the reference isolate—Salmonella enterica subsp. enterica serovar Dublin str. CT_02021853 (NCBI accession NC_011205.1) was constructed, and the assembled contigs were screened against the customized database for the detection of those genes in our isolates’ assembled contigs. For each of the two isolates, contigs that reported hits for each gene target were aligned to the reference gene using Molecular Evolution Genetics Analysis (MEGA—version 12) to detect nucleotide mutations. Furthermore, the ExPASy (Expert Protein Analysis System) translate tool (https://web.expasy.org/translate/), accessed on 10 November 2025, was used to convert the nucleotide sequences into their corresponding amino acid sequences and again aligned to the reference sequence to detect mutations at the amino acid level.
2.7. Protein Modelling
Amino acid sequences were modelled into proteins using the SWISS-MODEL automated protein structure homology-modelling server (https://swissmodel.expasy.org/), accessed on 21 October 2025. The generated protein structures were superimposed and compared in a 3-D format to analyze the effect of mutation on the general structure of the modelled proteins.
3. Results
3.1. Antimicrobial Susceptibility Testing of Isolates Against Ciprofloxacin
Out of the 17 isolates screened, only one isolate, identified as Salmonella Dublin recovered in 2023 from the fetal stomach content of a bovine, was resistant to CIP. The MIC value for this resistant isolate, as determined by the Kirby-Bauer disk diffusion method, was 1.5 μg/mL, as shown in Table 3. For this CIP-resistant isolate and its CIP-susceptible counterpart, the MIC values obtained through the broth dilution method were 1.95 μg/mL and 0.03 μg/mL, respectively (Table 3).
3.2. Gene Expression Analysis for the CIP-Resistant and CIP-Susceptible Salmonella Dublin Isolates
It was observed that, although the expression of the marA gene was almost at equal proportions for both the resistant and the susceptible isolate, the expression levels for the acrA, acrB, and the soxS genes were markedly higher in the CIP-resistant isolate compared to the CIP-susceptible isolate (Figure 1).
3.3. Chromosomal Mutations Identified in the Isolates
No mutations were identified in the CIP-susceptible isolate. However, the gyrase genes (gyrA and gyrB) and the topoisomerase genes (parC and parE) of the CIP-resistant isolate showed several synonymous mutations at the nucleotide level, as shown in Supplementary Table S1. The only non-synonymous mutation occurred as a switch from Adenine (A) to Guanine (G) at position 2603, resulting in a corresponding amino acid codon switch from Asparagine (N) to Serine (S) at position 868 (Figure 2).
3.4. Protein Modeling
The GyrA protein sequences of both the resistant isolate and reference isolate were modeled using the DNA gyrase subunit A of Salmonella as a template, with a sequence identity of 100 and 99.89, respectively. The structures generated for the two proteins were very comparable except for the structure of the single amino acid switch, as shown in Figure 3.
3.5. Plasmid and Antimicrobial Resistance Genes
For resistance genes, both the CIP-resistant and CIP-susceptible isolates carried the mdtK, emrR, emrA, and emrB, which have previously been noted to have activity against fluoroquinolones (Supplementary Data: Tables S2 and S3). No plasmids were detected in the CIP-resistant isolate. However, the IncFII(S) plasmid was identified in the CIP-susceptible isolate (Supplementary Data: Tables S4 and S5).
4. Discussion
This study adds to our understanding of CIP resistance in Salmonella Dublin, which has, over the years, been an underexplored topic. Although it focuses on the less-frequent finding of a single CIP-resistant isolate, the robust combination of two different susceptibility testing, gene expression analysis, whole-genome sequencing, and protein modeling provides a wide-view approach to understanding the potential origin of CIP resistance in this single Salmonella Dublin isolate.
The continuous decrease in the susceptibility of Salmonella Dublin to drugs like CIP has been a growing concern over the years [24]. However, compared to other highly prevalent serovars such as Salmonella Enteritidis and Salmonella Typhimurium, few studies exist investigating this phenomenon because CIP-resistant Salmonella Dublin is less frequently isolated overall [14]. This, undoubtedly, has impeded progress in knowing the repertoire of mechanisms Salmonella Dublin adopts in conferring resistance against CIP, including key genetic mutations. In providing a comprehensive summary of reported mutations in the quinolone resistance-determining region (QRDR) of quinolone-resistant Salmonella isolates, Shaheen et al. noted that continuous research may lead to the expansion of these regions [14]. In addition, the frequency and distribution of these mutations may not necessarily be a Salmonella serovar-specific phenomenon [25].
Although our CIP-resistant Salmonella Dublin isolate carried a GyrA Asp868Ser mutation, this mutation was outside the defined QRDR for Salmonella [14]. In Salmonella, some commonly identified mutations in the GyrA protein have included Ser83Phe, Arg47Ser, Asp82Asn, Asp87Asn/Gly/Tyr/Val, Glu133Gly, and Asp147Gly [8,14,26,27]. Considering the GyrA QRDR, which spans approximately amino acid positions 67 to 106 [14], these notable mutations: Arg47Ser, Glu133Gly, and Asp147Gly, just like the one identified in this study, are clearly outside the QRDR, further supporting the suggestion that the QRDR could be potentially expanded, especially where the mutations are confirmed to confer resistance. We hypothesize that our identified mutation may not have any significant impact on the function of the GyrA protein and perhaps may not have contributed to the observed CIP-resistant phenotype of our isolate. This is mainly because no significant change in the structure of the GyrA protein was observed. This claim about the non-functionality of our observed mutation, however, is a subject we intend to investigate further in future studies. It is also important to note that not all quinolone-resistant Salmonella isolates harbor mutations in the gyrA gene, and resistance may not always correlate with such mutations [28,29].
In this study, it was also observed that both the CIP-resistant and CIP-susceptible isolates carried the multidrug-resistant (MDR) genes mdtK, emrR, emrA, and emrB, which act against notable drugs like FQs. Nevertheless, an active expression of multi-drug efflux-pump-related genes in the CIP-resistant isolate in this study may have informed the observed resistance even in the absence of a potentially non-impactful mutation. In fact, upregulation of MDR efflux genes is key in mediating quinolone resistance [8]. The importance of these efflux pumps to producing resistant isolates cannot be overemphasized. This is because only a target site mutation in the all-important gyrA gene may not necessarily translate to clinical resistance if no efflux pump activities exist [30]. This underscores the significant synergy of both mutations and efflux activity in producing drug resistance in microbes.
As earlier stated in previous research, the action of multidrug efflux transporters in extruding several chemotherapeutic agents greatly helps in conferring resistance [31]. The overexpression of acrA and acrB genes has been strongly correlated with FQ and MDR in Salmonella and other pathogens [12,32]. Hence, the well-expressed acrAB genes in the CIP-resistant isolate in this study suggest their potential significance in contributing to the observed resistant phenotype. Furthermore, the acrAB in Salmonella is part of the acrAB-tolC multidrug efflux system, which is also mainly regulated by the ramRA locus [21]. The ramA gene is responsible for the activation of the transcription of acrAB and tolC genes, and the gene is, in turn, repressed by the ramR gene [21]. The ramA gene, together with the marA and the soxS genes, components of the mar box operon [8], are essential in drug protective responses as their upregulation increases efflux activity for cellular detoxification [30,33]. This detoxification is ultimately achieved through the activation of acrAB transcription [8].
The ramA gene was, however, the least expressed in the CIP-resistant isolate, and this may have been due to expression at levels just enough to sustain drug efflux activity. This is because a high expression of the ramA gene activates a feedback reaction from the ramR gene to repress its activity [34]. The ramA gene activates the acrAB efflux pump. Hence, its inhibition by ramR due to overexpression will lead to a counter-survival action. On the other hand, the soxS gene was among the least expressed genes in the CIP-susceptible isolate, and this may have contributed to its susceptible phenotype. The superoxide (soxS) gene, as the name suggests, is upregulated whenever bacteria are under a threat of oxidative stress, as it is essential in initiating the transcription of genes aimed at reducing superoxide and nitric oxide stress in the bacteria [30]. CIP induces the production of superoxide anion and other reactive oxygen species in microbes [35], and a reduced ability to extrude these from the bacterial cell can compromise survival. Despite the cogent inferences made regarding our comparative gene expression results for these genes, future studies aim to adopt protein quantification and functional inhibition assays to elucidate these observations.
Finally, the mechanisms of quinolone resistance in many bacterial pathogens can be summed up as: (i) mutations in the QRDRs of the gyr and par genes on the chromosome leading to low-affinity binding of the topoisomerase enzymes; (ii) plasmid-mediated quinolone resistance (PMQR), where pathogens acquire the qnr, the aac(6′)-lb-cr, or oqxAB and qepA genes, thus decreasing FQ activity and increasing their efflux; and (iii) the downregulation and upregulation of multidrug efflux pumps to ultimately reduce cellular concentrations of FQs [3,4,36,37]. Although many CIP-resistant Salmonella rely on plasmid-mediated quinolone resistance (PMQR) [6], no plasmid was detected in our CIP-resistant isolate.
This study was limited by the analysis of only one identified CIP-resistant isolate among the cohort of Salmonella isolates screened. Hence, all findings and discussions were made based on this isolate, limiting the generalization of our findings to the entire serovar. Nevertheless, the strength of this study lies in reporting the rare isolation of a CIP-resistant Salmonella Dublin isolate and adopting a combination of robust analyses to investigate the observed resistant phenotype. Our result highlights the potentially significant role of an efficient efflux system in contributing to CIP resistance in this Salmonella Dublin isolate, even when no impactful mutations were identified.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rusu A. Lungu I.A. Moldovan O.L. Tanase C. Hancu G. Structural Characterization of the Millennial Antibacterial (Fluoro) Quinolones—Shaping the Fifth Generation Pharmaceutics 202113128910.3390/pharmaceutics 1308128934452252 PMC 8399897 · doi ↗ · pubmed ↗
- 2Strahilevitz J. Jacoby G.A. Hooper D.C. Robicsek A. Plasmid-Mediated Quinolone Resistance: A Multifaceted Threat Clin. Microbiol. Rev.20092266468910.1128/CMR.00016-0919822894 PMC 2772364 · doi ↗ · pubmed ↗
- 3Mahapatra A. Patro A.R.K. Khajuria A. Dhal S. Praharaj A.K. Ciprofloxacin-Resistant Gram-Negative Isolates from a Tertiary Care Hospital in Eastern India with Novel Gyr A and Par C Gene Mutations Med. J. Armed Forces India 202278243110.1016/j.mjafi.2019.10.00235035040 PMC 8737103 · doi ↗ · pubmed ↗
- 4Cuypers W.L. Jacobs J. Wong V. Klemm E.J. Deborggraeve S. van Puyvelde S. Fluoroquinolone Resistance in Salmonella: Insights by Whole-genome Sequencing Microb. Genom.20184 e 00019510.1099/mgen.0.00019529975627 PMC 6113872 · doi ↗ · pubmed ↗
- 5Das T. Nath C. Das P. Ghosh K. Logno T.A. Debnath P. Dash S. Devnath H.S. Das S. Islam M.Z. High Prevalence of Ciprofloxacin Resistance in Escherichia coli Isolated from Chickens, Humans, and the Environment: An Emerging One Health Issue P Lo S ONE 202318 e 029404310.1371/journal.pone.029404337983240 PMC 10659180 · doi ↗ · pubmed ↗
- 6Chen K. Dong N. Chan E.W.C. Chen S. Transmission of Ciprofloxacin Resistance in Salmonella Mediated by a Novel Type of Conjugative Helper Plasmids Emerg. Microbes Infect.2019885786510.1080/22221751.2019.162619731169081 PMC 6566993 · doi ↗ · pubmed ↗
- 7Hopkins K.L. Davies R.H. Threlfall E.J. Mechanisms of Quinolone Resistance in Escherichia coli and Salmonella: Recent Developments Int. J. Antimicrob. Agents 20052535837310.1016/j.ijantimicag.2005.02.00615848289 · doi ↗ · pubmed ↗
- 8Gu Y. Huang L. Wu C. Huang J. Hao H. Yuan Z. Cheng G. The Evolution of Fluoroquinolone Resistance in Salmonella under Exposure to Sub-Inhibitory Concentration of Enrofloxacin Int. J. Mol. Sci.2021221221810.3390/ijms 22221221834830098 PMC 8619427 · doi ↗ · pubmed ↗