Correlation Analysis Between HLA Polymorphisms and Immune Response to Hepatitis B Vaccine in Children with Acute Lymphoblastic Leukemia
Rui Zhang, Tian Yang, Yijin Gao, Hua Zhang, Yi Fei, Laibao Yang, Pengfei Deng

TL;DR
This study found that certain HLA gene variations may affect how well children with leukemia respond to the hepatitis B vaccine.
Contribution
The study identifies HLA-B*3501 as a novel genetic factor linked to lower vaccine response in leukemia patients.
Findings
Post-vaccination anti-HBs titers were significantly higher than pre-vaccination levels.
HLA-B*3501 allele was negatively correlated with anti-HBs response levels.
A low-response group showed significantly lower antibody titers compared to the high-response group.
Abstract
Background: The human leukocyte antigen (HLA) is crucial for antigen presentation and vaccine efficacy. This study examined the association between HLA polymorphisms and the immune response to hepatitis B vaccination in children with acute lymphoblastic leukemia (ALL). Methods: 101 pediatric ALL patients at Shanghai Children’s Medical Center affiliated with Shanghai Jiaotong University School of Medicine who tested negative for hepatitis B surface antibody (anti-HBs) and were not infected with hepatitis B received three doses of the hepatitis B vaccine. Anti-HBs titers were measured before and after vaccination. Participants were divided into high- and low-response groups based on post-vaccination anti-HBs titers. Sequence-specific primer polymerase chain reaction (PCR-SSP) was used to genotype HLA-A, -B, -Cw, -DRB1, and -DQB1 alleles. Results: Pre-vaccination anti-HBs titers were 3.38…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
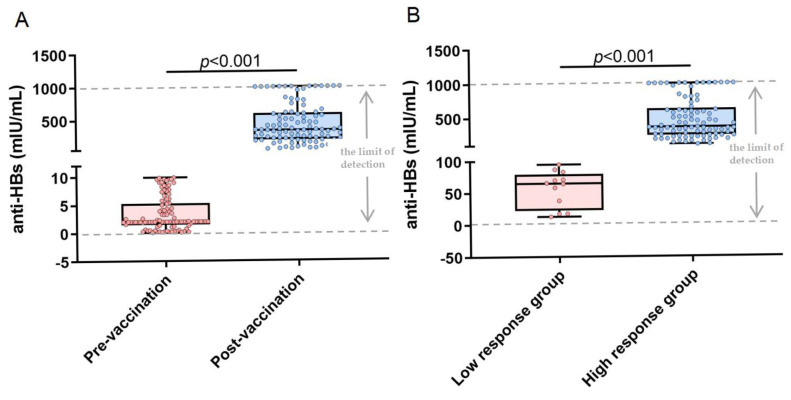 Figure 1
Figure 1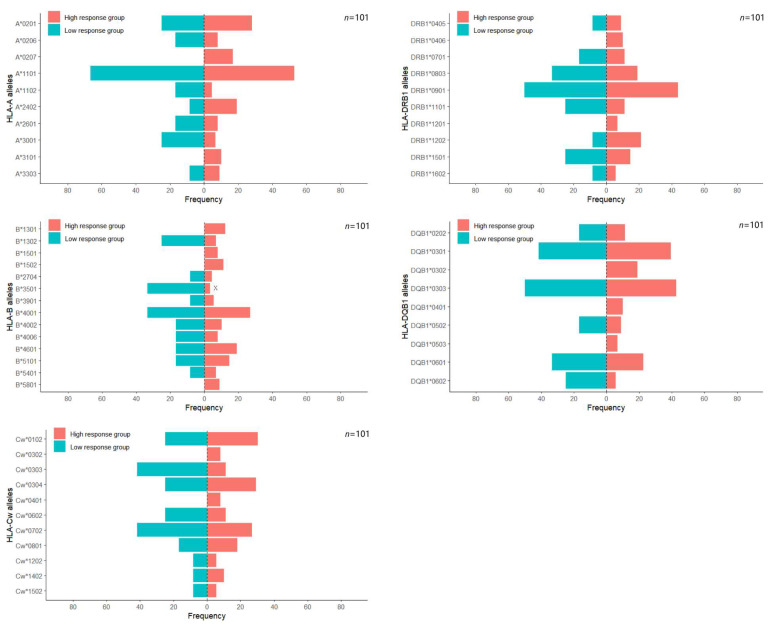 Figure 2
Figure 2- —Key Discipline Program of Pudong New Area Health System
- —Shanghai Three-Year Action Plan for Strengthening Public Health System (2023–2025) Discipline Leader Project
- —Pudong New Area Science and Technology Development Fund Program
- —Pudong New Area Health Commission General Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHepatitis B Virus Studies · Immunotherapy and Immune Responses · Acute Lymphoblastic Leukemia research
1. Introduction
Based on the Global Burden of Disease database, from 1990 to 2021, the global incidence, mortality, and disability-adjusted life years (DALYs) of acute lymphoblastic leukemia (ALL) showed a downward trend. However, 53,485 new cases of childhood ALL were reported globally in 2021, which led to 23,991 deaths and 1,960,922 DALYs [1]. The data therefore demonstrates that the health burden of ALL on children remains substantial and cannot be neglected. The immunocompromised state of children with ALL, arising from both the pathophysiology of the disease and therapeutic measures, significantly heightens their vulnerability to infections compared to immunocompetent children [2,3].
Hepatitis B virus (HBV) infection persists as a significant global public health issue. It is estimated that approximately 296 million individuals are living with chronic HBV infection, and approximately 884,000 deaths each year are attributable to HBV-related diseases [4,5]. Recent research reveals that China shoulders the highest global burden of HBV infection, with 75 million chronic carriers, accounting for one-third of the infected population worldwide [6]. However, for pediatric patients with ALL, chemotherapy or hematopoietic stem cell transplantation may induce immunosuppression, thereby significantly increasing the risk of HBV infection or reactivation. Previous studies have established that HBV reactivation is a significant concern in patients with hematological malignancies. Among the broader group of patients who are hepatitis B surface antigen (HBsAg)-negative, the overall reactivation rate is approximately 3.1%, with patients suffering from ALL being at a particularly elevated risk [7]. It is crucial to recognize that the population of “HBsAg-negative” patients is heterogeneous and includes individuals with a past resolved hepatitis B infection (hepatitis B core antibody-positive, anti-HBc-positive). A prospective cohort study revealed that 39% of untreated HBsAg-negative pediatric ALL patients were anti-HBc-positive [8], and this subgroup accounts for the substantial majority of reactivation events, with a cumulative incidence reaching up to 10.5% within three years of chemotherapy [7]. In contrast, the risk of reactivation is extremely low for patients who are truly uninfected (HBsAg-negative and anti-HBc-negative); however, they remain susceptible to new HBV infection.
The hepatitis B vaccine is widely recognized as the most effective way to prevent HBV infection. It has been proven to reduce the risk of developing acute and chronic hepatitis B, cirrhosis, and hepatocellular carcinoma [9]. Currently, research on hepatitis B vaccines primarily focuses on antibody levels and healthy populations, with extremely limited studies conducted on ALL survivors who have no history of hepatitis B infection. The efficacy of the hepatitis B vaccine varies considerably among individuals, influenced by multiple factors. The major histocompatibility complex (MHC) haplotypes and genetic polymorphisms, which are closely associated with immune regulation, are key genetic factors that affect vaccine effectiveness [10,11]. The human leukocyte antigen (HLA) system, encoded by the MHC on chromosome 6, comprises three main classes: Class I, comprising HLA-A, -B, and -C, is expressed on nucleated cells; Class II, including HLA-DR, -DQ, and -DP, is present on antigen-presenting cells; Class III encompasses immunoregulatory components [12]. HLA Class II molecules play a crucial role in mediating vaccine responsiveness by presenting HBsAg to CD4+ T lymphocytes, thus initiating humoral immune responses. Current evidence suggests that DPB104:01 and DPB104:02 enhance hepatitis B surface antibody (anti-HBs) seroconversion, while DPB105:01 has an inhibitory effect [13]. Specifically, DRB114:54 may reduce anti-HBe responsiveness due to impaired HBeAg peptide binding [14], and DQB103 and DQB105 alleles are associated with persistent HBV infection [15]. Population-based analyses indicate that HLA-DR/DQ haplotypes can account for up to 10% of the variance in vaccine response within the African population [16].
Although extensive research has confirmed an association between HLA polymorphisms and hepatitis B vaccine immunogenicity, there remains a lack of in-depth genetic exploration within the ALL population. Particularly in China, the large size of the ALL population, low vaccination rates, and insufficient awareness and parental initiative regarding vaccination have led to a scarcity of relevant studies. Therefore, this study will explore the potential influence of HLA polymorphism on the antibody response levels following hepatitis B vaccination in pediatric ALL patients.
2. Materials and Methods
2.1. Study Design and Objectives
This prospective study recruited 101 children diagnosed with ALL at Shanghai Children’s Medical Center, affiliated with Shanghai Jiaotong University School of Medicine, from 2021 to 2024. Before their inclusion in the study, written informed consent was procured from all legal guardians. Recombinant hepatitis B vaccine (01190526A, Hissen Vaccine, Dalian, China) was used for the study subjects and administered as 3 doses of 10 μg at 0, 1, and 6 months. Venous blood specimens were collected at two discrete time points: (1) before vaccination, which served as the baseline, and (2) one month after completion of the hepatitis B vaccination program.
Genomic deoxyribonucleic acid (DNA) was extracted from peripheral blood samples. Anti-HBs titers were quantitatively determined via an enzyme-linked immunosorbent assay (anti-HBs ELISA kit from the Shanghai Zheke Biological Engineering Center, Shanghai, China) in conjunction with the EVO automatic enzyme immunoassay system (Tecan Group Ltd., Zürich, Switzerland). The detection limit was >0 mIU/mL, and the quantification range of the anti-HBs ELISA assay was 0–1000 mIU/mL.
China’s Technical Guide for Adult Hepatitis B Immunization defines anti-HBs levels ≥ 10 mIU/mL as protective. In contrast, Public Health England and recent studies suggest that ≥100 mIU/mL may also serve as an alternative threshold [17,18,19]. Therefore, employing 10 mIU/mL and 100 mIU/mL as thresholds, individuals with anti-HBs levels below 10 mIU/mL after hepatitis B vaccination were deemed to be in a seronegative state and were incorporated into the non-response group. Individuals with anti-HBs levels ≥ 10 mIU/mL are associated with complete clinical protection, yet they may experience reduced durability and are thus included in the low-response group. Conversely, individuals with anti-HBs levels equal to or exceeding 100 mIU/mL were classified as complete vaccine responders and included in the high-response group.
2.2. Inclusion and Exclusion Criteria
Inclusion Criteria: (1) Children who have been clinically diagnosed with ALL. (2) Children aged 0 to 18 years with a history of complete hepatitis B vaccination. (3) Children tested negative for HBV serological markers, defined as HBsAg negative, anti-HBc negative, and anti-HBs titer < 10 mIU/mL, to ensure study subjects were genuine HBV susceptible individuals without prior infection or occult infection. (4) Children who have not used immunosuppressants or received blood transfusions within the past 3 months. (5) Children must be in a state of immune restoration with normal immune function for at least 12 months after chemotherapy, with a blood test indicating a neutrophil count > 1.40 × 10^9^/L and a platelet count > 150 × 10^9^/L. (6) Subjects must undergo a comprehensive evaluation by a clinician regarding clinical treatment, duration of immunosuppression, and the risk of diseases requiring prevention.
Exclusion Criteria: (1) Individuals who are users of monoclonal antibodies, especially anti-tumor necrosis factor agents. (2) Individuals who are users of intermittent or low-dose chemotherapeutic agents or other immunosuppressive drugs.
2.3. Genetic Polymorphism Detection
HLA genotyping was conducted using Sequence-specific primer polymerase chain reaction (PCR-SSP). Specifically, allele-specific primers were designed to amplify the following loci: HLA-A, -B, -Cw, -DRB1, and -DQB1. The amplified products were then compared with standard markers using a scanning imaging analysis system to ascertain the HLA-A, -B, -Cw, -DRB1, and -DQB1 genotypes. Subsequently, the positive products were subjected to gene sequencing.
2.4. Statistical Analysis
Statistical analyses were conducted using SPSS 24.0, and graphical visualizations were generated using GraphPad Prism 7.00 and RStudio (version R-4.5.0). The chi-square test was utilized to compare the disparities among children in different response groups with respect to gender, risk groups, transplant status, radiation therapy history, immigration status, and genotype carrier rates. The Shapiro–Wilk test was used to assess the normality of anti-HBs titers across groups, both before and after hepatitis B vaccination. Data that met the normality criteria were subjected to t-test analysis. Binary logistic regression was used to evaluate the association between HLA alleles and anti-HBs titers. Statistical significance was defined as a two-tailed p-value < 0.05. The p-values for differences in HLA gene polymorphisms between the two groups were corrected using the Bonferroni method.
3. Results
3.1. Analysis of the Immune Response Levels in Children with ALL After Hepatitis B Vaccination
Before vaccination, all participants (n = 101) were seronegative for anti-HBs, with a mean baseline titer of 3.38 ± 2.97 mIU/mL. After vaccination, a 100% seropositivity rate was achieved, accompanied by significantly elevated anti-HBs titers (429.61 ± 303.13 mIU/mL; p < 0.001 compared with baseline) (Figure 1A). We categorized participants into low-responder and high-responder groups based on post-immunization anti-HBs levels exceeding 100 mIU/mL. The results revealed statistically significant differences in antibody levels between the low-response group (56.47 ± 28.38 mIU/mL) and the high-response group (479.93 ± 287.70 mIU/mL; p < 0.001) (Figure 1B).
Twelve children (11.88%) in the low-response group and 89 children (88.12%) in the high-response group demonstrated positive anti-HBs titers. No significant inter-group disparities were detected in demographic factors such as age, gender, and immigration status or clinical characteristics including risk groups, transplantation status and radiation therapy history across response categories (all p > 0.05) (Table 1).
3.2. HLA Genotype Distribution and Immune Response Levels to Hepatitis B Vaccine in Children with ALL
Univariate analysis of HLA allele distribution between vaccine response groups revealed distinct patterns at the HLA-A, -B, -C, -DRB1, and -DQB1 loci. The frequencies of HLA—A alleles were predominantly represented by A1101 (54.46%), A0201 (27.72%), A2402 (17.82%), and A0207 (14.85%). There were no statistically significant differences in carrier frequencies between the response groups. The major HLA—B alleles were B4001 (27.72%), B4601 (18.81%), B5101 (14.85%), B1301 (10.89%), and B4002 (10.89%). Significant inter-group differences in carrier frequency were detected for B3501 (p < 0.01), while no other HLA—B alleles exhibited significant associations. The most prevalent HLA—Cw alleles were Cw0102 (29.70%), Cw0304 (28.71%), Cw0702 (28.71%), Cw0801 (17.82%), Cw0303 (14.85%), and Cw0602 (12.87%). Significant differences in carrier rates were noted for Cw0303 (p < 0.05), whereas other Cw alleles did not differ significantly. The distribution of HLA—DRB1 alleles was characterized by the predominance of DRB10901 (44.55%), DRB10803 (20.79%), DRB11202 (19.80%), DRB11501 (15.84%), DRB11101 (12.87%), and DRB10701 (11.88%). No significant association was observed between any DRB1 allele and vaccine response levels. Likewise, in HLA-DQB1, the most prevalent alleles were DQB10303 (43.56%), DQB10301 (39.60%), DQB10601 (23.76%), DQB10302 (16.83%), and DQB10202 (11.88%). Similarly, no significant association was found between any DQB1 allele and vaccine response. Since this study involved comparing multiple HLA polymorphic loci between two groups, Bonferroni correction revealed that only the B*3501 carrier frequency showed a significant intergroup difference (p < 0.00357 for Bonferroni correction) (Table 2, Figure 2).
3.3. Association Between HLA Genotypes and Antibody Levels
We included alleles B3501, Cw0303, and DQB10602 with p < 0.1 from univariate analyses, along with DQB10301, which may affect hepatitis B antibody levels, into a binary logistic regression model for in-depth analysis. The findings revealed that B3501 exerted a statistically significant influence on antibody response levels (OR = 0.113, 95% CI: 0.018–0.698, p = 0.019), whereas Cw0303, DQB10301, and DQB10602 did not (Table 3).
4. Discussion
Children who have undergone therapy for ALL, particularly those who received allogeneic hematopoietic stem cell transplantation, experience prolonged immunosuppression and ablation of pre-existing immune memory. Consequently, even those previously vaccinated against hepatitis B may become seronegative for anti-HBs and are at risk for HBV infection post-treatment [3,20]. Revaccination is therefore essential to restore protection during long-term survivorship. This study specifically focuses on ALL survivors who were anti-HBs-negative following completion of therapy, representing a population that requires a complete de novo hepatitis B vaccination series. This study aims to evaluate the immunogenicity of the standard three-dose hepatitis B vaccine regimen in this susceptible population and to investigate the associated HLA genes. Our data indicated that children with ALL who achieve clinical remission exhibit significantly stronger immune responses to the hepatitis B vaccine. This phenomenon likely reflects immunological recovery in stable ALL patients, in whom vaccine responsiveness approaches that of immunocompetent individuals [21]. This finding corroborates the report by Viana et al., which stated that post-chemotherapy booster vaccination significantly elevates anti-HBs titers to protective levels (10 mIU/mL) in ALL patients [21].
This study has not revealed statistically significant differences between the high anti-HBs response group and the low anti-HBs response group in terms of demographic characteristics such as age, gender and immigration status. Furthermore, no significant differences were observed between the two groups in clinical characteristics, including risk groups, transplant status, and radiation therapy history. The findings may have resulted from the fact that all patients included in the present study were from the same specialized hospital, had the same disease type, received similar treatment regimens, and were in a state of near-complete recovery at the time of vaccination, resulting in minimal internal variability. Additionally, with the current sample size, it may be challenging to detect the effects of factors with smaller effect sizes, such as age and gender. Collectively, these factors highlight the role of genetic factors.
Our genetic association study uncovers a remarkable and distinct pattern of HLA-mediated immunity following hepatitis B vaccination. Univariate analysis results suggest that both B3501 and Cw0303 may significantly influence antibody levels following hepatitis B vaccination in children with ALL. Notably, compared with the general Han Chinese population (3.0%), the B3501 allele had a higher carrier frequency (6.93%) among pediatric ALL patients in this study, predominantly observed in the poor-response patient group. No association has been identified linking B3501 to vaccine-induced HBV antibody levels. Previous research has indicated that the peptide-deficient B3501 tetramer is capable of directly facilitating the activation of CD8^+^ T cells, which implies that B3501 exhibits substantial potential for T cell activation [22]. Activated CD8^+^ T cells may enhance the production of anti-HBs after hepatitis B vaccination by assisting the B-cell response, suggesting that B*3501 contributes to the elevation of anti-HBs levels [23].
Nevertheless, the antibody levels generated are insufficient to enable all patients to achieve a complete immune state. Similarly, Cw0303 were predominantly distributed in the low-response group, which is in line with the study by D. Wolday et al., suggesting that they bind strongly to their respective peptide fragments but exhibit low or no antibody production [24]. Nevertheless, after applying the Bonferroni correction to p-values and employing a binary logistic regression model, the association with Cw0303 weakened and lost statistical independence, whereas that with B3501 remained robust and significant. These findings indicate that in this study, Cw0303 may be genetically linked to B3501; however, its role in anti-HBs production is not entirely independent of B3501.
Previous studies have demonstrated that specific HLA class II alleles (DRB1 and DQB1) are associated with antibody responses to the hepatitis B vaccine [25]. For example, the alleles DQB101, DQB10301, DQB115, DQB10501, DQB106, and DQB10602 are strongly associated with significantly enhanced antibody responses to hepatitis B vaccines. In contrast, alleles such as DRB10301, DRB104, DRB107, DRB11302, and DQB1*02 exhibit the opposite effect. It is noteworthy that in this study, DRB1 and DQB1 did not significantly influence the antibody response levels in children with ALL following hepatitis B vaccine booster administration. This finding suggests that in the pediatric ALL population, the HLA class I pathway, particularly the HLA-B locus, may play an unexpectedly dominant and irreplaceable role in humoral immunity against hepatitis B vaccination.
The study has several limitations. Firstly, the sample size was relatively limited. Secondly, antibody titers were measured over a short-term period, and it was not feasible to evaluate the durability of the vaccine due to the absence of long-term antibody monitoring data in children with ALL. Durability evaluation will be gradually considered in subsequent studies. Finally, this study strictly defined the “no history of hepatitis B infection” group by excluding individuals who were anti-HBc-positive. This implies that our findings cannot be directly generalized to all HBsAg-negative patients, as this population includes individuals with prior infection who retain immune memory and may exhibit stronger vaccine responses. Future studies could compare post-vaccination responses between anti-HBc-negative and anti-HBc-positive patients, and long-term continuous cohort monitoring data may continue to provide supplementary information.
5. Conclusions
This study demonstrates that administering three doses of hepatitis B vaccine to children with ALL following recovery is effective, with all recipients exhibiting protective immunity against HBV. HLA-B*3501 may be negatively correlated with hepatitis B vaccine-induced immune response in children with ALL. This finding requires further validation in other cohorts.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wang L. Yao X. Yang L. Global, regional, and national burden of children and adolescents with acute lymphoblastic leukemia from 1990 to 2021: A systematic analysis for the global burden of disease study 2021 Front. Public Health 202513152575110.3389/fpubh.2025.152575139944070 PMC 11813771 · doi ↗ · pubmed ↗
- 2Rajendran P.V. Thankamony P. Rajeswari B. Sojamani G.C. Nair M. Parukuttyamma K. Krishna Km J. Loss of protective anti-H Bs titers and seroconversion to hepatitis B vaccination in children during chemotherapy for acute lymphoblastic leukemia Pediatr. Blood Cancer 202370 e 3015410.1002/pbc.3015436545908 · doi ↗ · pubmed ↗
- 3Xiong D. Cai W. Zhao W. Risk factors of HBV reactivation in leukemia patients with resolved HBV infection after allogeneic hematopoietic stem cell transplantation Clin. Res. Hepatol. Gastroenterol.20244810244710.1016/j.clinre.2024.10244739181184 · doi ↗ · pubmed ↗
- 4Pattyn J. Hendrickx G. Vorsters A. Van Damme P. Hepatitis B Vaccines J. Infect. Dis.2021224 S 343S 35110.1093/infdis/jiaa 66834590138 PMC 8482019 · doi ↗ · pubmed ↗
- 5Sarfaraz N. Somarowthu S. Bouchard M.J. The interplay of long noncoding RN As and hepatitis B virus J. Med. Virol.202395 e 2805810.1002/jmv.2805835946066 · doi ↗ · pubmed ↗
- 6Yan R. Sun M. Yang H. Du S. Sun L. Mao Y. 2024 latest report on hepatitis B virus epidemiology in China: Current status, changing trajectory, and challenges Hepatobiliary Surg. Nutr.202514667710.21037/hbsn-2024-75439925891 PMC 11806133 · doi ↗ · pubmed ↗
- 7Han J.W. Yang H. Lee H.L. Bae S.H. Choi J.Y. Lee J.W. Kim H.J. Lee S. Cho S.G. Min C.K. Risk factors and outcomes of hepatitis B virus reactivation in hepatitis B surface antigen negative patients with hematological malignancies Hepatol. Res.20164665766810.1111/hepr.1260326445232 · doi ↗ · pubmed ↗
- 8Guruprasad B. Kavitha S. Aruna Kumari B.S. Vijaykumar B.R. Sumati B.G. Mahua S. Appaji L. Jayshree R.S. Risk of hepatitis B infection in pediatric acute lymphoblastic leukemia in a tertiary care center from South India Pediatr. Blood Cancer 2014611616161910.1002/pbc.2506524798418 · doi ↗ · pubmed ↗