Coexistence of Calliergonella cuspidata and Hamatocaulis vernicosus Under Different Fen Topography Types and Microhabitat Conditions
Monika Kalvaitienė, Ilona Jukonienė

TL;DR
This study explores how two moss species coexist in wetlands under different environmental conditions.
Contribution
The study reveals how microhabitat and hydrological conditions influence competitive interactions between two moss species.
Findings
Under wet conditions, Hamatocaulis vernicosus expands and competes successfully with Calliergonella cuspidata.
Calliergonella cuspidata only shows competitive advantage in hummocky microtopographic settings.
Stable hydrological conditions support coexistence, while altered water regimes threaten H. vernicosus.
Abstract
Hamatocaulis vernicosus and Calliergonella cuspidata commonly co-occur in base-rich fens, reflecting overlapping ecological niches. While C. cuspidata is a widespread and ecologically plastic fen species often associated with eutrophicated wetlands, H. vernicosus is a habitat-specialist species of conservation concern. This study investigated the competitive interactions between these two moss species and the role of microhabitat conditions in their coexistence. A reciprocal transplant experiment was conducted in a natural, rich fen in southeastern Lithuania using replicated experimental plots across different microtopographic and hydrological conditions. Species cover and spread were monitored to assess competitive performance following transplantation. The results showed that under wet conditions, H. vernicosus was able to expand into surrounding areas and successfully compete with C.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
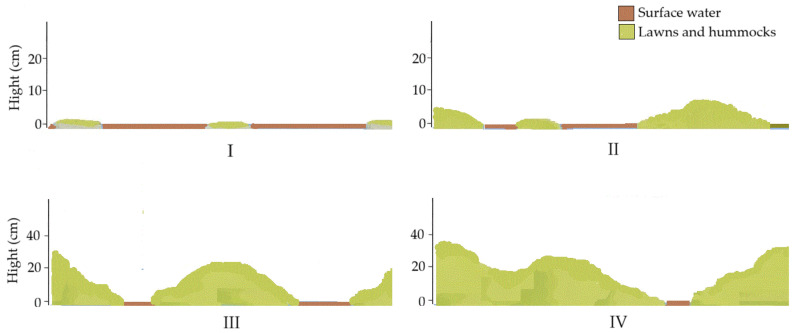 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPeatlands and Wetlands Ecology · Ecology and Vegetation Dynamics Studies · Bryophyte Studies and Records
1. Introduction
Fens, which are characterised by high mineral concentrations arising from ground or surface water [1], are among the most species-rich and ecologically complex wetland habitats. Their topographic heterogeneity, expressed through microforms such as hummocks and hollows, creates spatial variation in hydrological, chemical, and microclimatic conditions [2], supporting a wide variety of bryophytes, including species of contrasting ecological requirements [3,4,5,6,7,8]. Hamatocaulis vernicosus (Mitt.) Hedenäs and Calliergonella cuspidata (Hedw.) Loeske are two moss species commonly found together in rich fens. However, they differ markedly in their ecological strategies, rarity, and sensitivity to environmental change.
Hamatocaulis vernicosus, a rare species of conservation priority, is protected under the Bern Convention [9] and listed in Annex II of the EU Habitats Directive [10], reflecting its vulnerability to environmental degradation, particularly eutrophication and hydrological fluctuations [11]. It typically occurs in mesotrophic and minerotrophic fens, which are characterised by stable hydrological regimes, high mineral availability, and minimal disturbance [11,12,13,14].
In contrast, C. cuspidata exhibits broad ecological tolerance and occurs across a wide range of habitats, including nutrient-rich or transitional fens [5,15]. Its ability to adapt morphologically in response to changes in light, moisture, and nutrient availability allows it to effectively colonise open or disturbed microhabitats [16]. The species is identified as a moss species that increases in abundance in fens undergoing early successional change triggered by nutrient enrichment [17,18,19,20]. Its expansion has been associated with the decline of calcicolous brown mosses, including H. vernicosus, particularly under elevated potassium or nitrogen levels [12,15,18,21].
The frequent occurrence of the competitive species C. cuspidata alongside the endangered European species H. vernicosus is believed to pose a significant threat to the latter [12,22]. Monitoring schemes were proposed that focus on interactions between H. vernicosus and C. cuspidata, as these may signal the initial phase of nutrient enrichment in seemingly intact fens within agricultural landscapes [15,22,23]. However, the experimental results show that increased N and P concentrations had a positive effect on the growth and biomass accumulation of the rare H. vernicosus, suggesting that nutrient availability alone does not fully explain competitive outcomes [24]. In this context, competitive outcomes between these species may also be shaped by fine-scale environmental heterogeneity, such as microtopography, which can modify local growing conditions independently of nutrient status. Despite its potential importance, the role of microrelief in mediating competition between C. cuspidata and H. vernicosus remains poorly understood. Field experiments with H. vernicosus demonstrated the species survival in the presence of C. cuspidata, depending on habitat conditions [25,26].
Since the habitats of H. vernicosus vary across Europe, it is essential to understand the role of competition in the local coexistence of these two species. Accurately interpreting monitoring signals and informing effective fen conservation and restoration strategies requires clarifying this relationship. Compared to many other regions of Europe, H. vernicosus habitats in Lithuania are diverse, occurring under a broader range of water chemistry conditions and with higher mineral content [14]. A key factor is the specific topography, which is characterised by shallow hollows and lawns fed by surface water that maintain a stable moisture regime. Lithuania stands out in Europe, having 37 Natura 2000 sites supporting H. vernicosus populations, as well as for the diversity of its habitats [27]. This highlights Lithuania’s significant responsibility for conserving this species on a European scale, and emphasises the importance of understanding the ecological requirements and potential threats facing H. vernicosus in this region.
Here we present the results of a transplant experiment conducted in the natural habitats of H. vernicosus, where the two species co-occur at equal abundance, forming distinct or mixed patches. During this study, we aimed: (1) to determine how species can survive when surrounded by patches of other species in different microhabitats; (2) to establish whether, once introduced, one species can successfully expand by replacing another species in its patches; and (3) to investigate the role of topographically driven microhabitat conditions in species competition.
This is important for predicting whether the growth of C. cuspidata alongside H. vernicosus could negatively impact H. vernicosus’s growth and survival across different fen topographies, and whether special management measures are always required to protect H. vernicosus.
2. Results
2.1. Changes in the Transplanted Subplots
2.1.1. Hamatocalis vernicosus
In the transplanted subplots of topography type I, the mean cover of H. vernicosus decreased from approximately 90% to 35.5% (Table 1). The PCoA ordination map reveals clearer separation of samples between 2019 and 2020–2021, indicating that population changes were more apparent over time rather than at early stages (Figure 1). The transplanted subplots of topography type II remained the most stable, with 41% of H. vernicosus coverage persisting. Although some fluctuations were observed in 2019, they did not persist in later years, reflecting a consistent, and relatively minor, level of change across the monitoring period (Figure 1). Hamatocaulis vernicosus in topography type III retained approximately 36% of mean cover, and subplots showed changes over the course of the study, with clearer differences becoming apparent at later stages. Topography type IV experienced the sharpest decline of H. vernicosus (Table 1). Changes in population structure are evident both in the early stages and in later years.
The changes in transplanted subplot structure varied greatly depending on the topography type, with the strongest structural divergence observed in types III and IV (Table 2, Figure 1). The cover of Calliergonella cuspidata increased in all transplanted H. vernicosus subplots (Table 1), although its contribution to structural changes—reflected in shifts in the proportional composition of subplot components such as H. vernicosus and herb covers —was most pronounced in topography type IV. In type I, it is likely that the dominance of H. vernicosus was mainly reduced by the amount of surface water. In type II, changes were minor, and H. vernicosus retained most of its coverage, showing structural stability. In type III, herb cover took over part of the former H. vernicosus location (Table 1).
2.1.2. Calliergonella cuspidata
As in the case of H. vernicosus transplantation, the C. cuspidata transplanted subplot exhibited temporal changes (Figure 1). The most significant decline in C. cuspidata coverage was recorded in topography type I, where mean cover decreased to 17% (Table 3). There, changes were most apparent in the later stages of the study, with an increase in surface water area largely associated with the observed decline (Figure 1). In type II, coverage decreased from 97% to 36%, with significant differences observed between the early (2019) and later (2020–2021) periods, corresponding to increases in H. vernicosus and surface water (Figure 1, Table 3). In type III, coverage declined to 23%, reflecting strong sensitivity to the expansion of H. vernicosus and herbs. For topography type IV, coverage remained the highest, at about 50%, with the most noticeable changes occurring between early and later periods, mainly driven by an increase in herb cover. Overall, the dynamics of the transplanted C. cuspidata subplots were statistically significant, except for types III and IV, for which no consistent time effects were detected; however, multivariate dispersion analysis still revealed increased subplot heterogeneity after transplanting (Table 2).
2.2. Changes in the Surrounding Subplots after Transplantation
After H. vernicosus transplantation, a statistically significant positive trend was recorded in surrounding subplots of all topography types (Mann–Kendall test p ≤ 0.05) (Table 4). The most significant absolute increase (ΔP) was recorded in I (36.6%), and the smallest in IV (1.12%) topography type. Meanwhile, no significant changes were observed in the control subplots (p ≥ 0.5), and ΔP ranged from −5.66 to 4.18, showing only minor or negative changes (Figure 2).
Calliergonella cuspidata also showed a consistent, statistically significant positive trend in the surrounding fields after transplantation across all I–IV topography types (Table 4). The absolute change (ΔP) was highest in the surrounding subplots of IV (22.66%) and lowest in II (2.93%) topography type. A significant positive trend was also observed in the surrounding subplots for topography types II–IV, with ΔP ranging from 5.20 to 66.17% in type IV. Only in the I topography type control subplots change was negative (ΔP = −10.54), and the trend was statistically insignificant (p = 0.33). This shows that, except for topography type I, this species exhibited strong growth under both transplantation and control conditions, and this was especially evident in topography type IV, where the change was most significant.
3. Discussion
Our research focuses on the relationship between C. cuspidata and H. vernicosus within the context of related studies. Calliergonella cuspidata is often considered a competitively superior species when it co-occurs with H. vernicosus, which may contribute to the rarity and threatened status of the latter [22]. This interpretation is particularly applicable to eutrophicated habitats as C. cuspidata seemed to have an especially advantageous position as a nutrient-demanding species [12,15,18,21].
Our research differs from previous experimental studies primarily because it was conducted under natural field conditions in a relatively undisturbed rich fen. We focused on changes in local topography and microhabitat conditions, which ranged from flooded lawns and low hummocks to more elevated hummocky surfaces. The hollow-hummock gradient seemed to be important not only for the distribution of H. vernicosus, as shown in our previous study [14], but also for its competitive traits. Our recent results show that flooded lawns and low hummocks surrounded by spring flushes represent the most suitable microhabitats for H. vernicosus. These conditions not only support the persistence of the species but also facilitate its survival and dispersal once established within C. cuspidata patches. Although the abundance of H. vernicosus declined in transplanted subplots under highly flooded conditions, this reduction is more likely attributable to excessive hydrological stress than to competitive suppression. On the other hand, its expansion in the surrounding subplots was the highest among all topographical types. This interpretation is consistent with its known flood tolerance and previous observations of successful establishment and spread under inundated conditions [12,28]. In contrast, C. cuspidata showed limited performance under similar conditions, indicating lower tolerance to prolonged inundation. Our research further supports the conclusion of experimental laboratory studies that H. vernicosus is not a weaker competitor and may even outperform C. cuspidata in flooded conditions [29], suggesting that optimal water level, rather than interspecific competition, is the primary driver shaping H. vernicosus growth. Firstly, it keeps the vascular plant cover low [12], without reducing the solar radiation available to bryophytes. Secondly, spring waters create cool conditions, which also enhance the development of H. vernicosus as demonstrated in laboratory experiments [29]. The species also appears adapted to elevated concentrations of calcium (Ca^2+^), iron (Fe^2+^), and manganese (Mn^2+^), enabling it to form dense lawns just above the groundwater level rather than on higher hummocks [24,30]. C. cuspidata—like several morphologically and phylogenetically distinct species, such as Aulacomnium palustre, Helodium blandowii and Marchantia polymorpha—shares intermediate hummock-forming frequency and desiccation avoidance. The importance of suitable fen conditions. These ecological requirements are consistent with observations from Hamatocaulis vernicosus reintroductions, where successful establishment occurred only under suitable fen conditions [25,26].
It is important to note that, despite being in control plots without transplantation, expansion of both H. vernicosus and C. cuspidata was observed, and the absolute numbers, with the exception of those for C. cuspidata expansion in hummocky areas, were not significant. Apparently, over a more extended period as they grow together, these species reach equilibrium, and changes in their abundance are not significant; however, in the case of disturbances, H. vernicosus has the competitive advantage.
It is important to note that, although expansion of both H. vernicosus and C. cuspidata was observed during the study period, their responses differed substantially between microhabitats. The expansion of H. vernicosus was most pronounced in the lawn and other non-hummock areas associated with higher groundwater levels and was largely restricted to transplanted subplots, with minimal change in hummocky conditions. This indicates that the observed increase in H. vernicosus was driven by transplantation and constrained by drier, elevated microhabitats. In contrast, C. cuspidata showed robust expansion in hummocky areas, including substantial increases in control plots. Overall, these results demonstrate species-specific and topography-dependent expansion patterns, with H. vernicosus responding primarily to transplantation and C. cuspidata exhibiting high ecological plasticity.
Our experiments show that C. cuspidata has a competitive advantage over H. vernicosus only in hummocky areas. Hummock microtopography establishes specific microclimatic conditions, with small-scale variations in soil thermal properties and water regimes [31]. Due to hummocks, bryophytes are distant or isolated from mineral fen water [32,33]. In the absence of cold spring water, bryophytes experience warmer microclimatic conditions. So, our results are in accordance with laboratory experiments showing that warmer environments are favourable for the growth of C. cuspidata [29]. Although H. vernicosus decreased significantly in drier conditions in recent years of the experiment compared to the first years, this decrease was followed not only by an increase in C. cuspidata, but also by the growth of herbs. It is unlikely that H. vernicosus and C. cuspidata can also benefit from the increased shading that results from the enhanced growth of vascular plants [34]. We did not analyse habitat eutrophication. Based on research in hummocky areas, we cannot confirm or deny whether C. cuspidata is competitively stronger than fen moss specialists when nutrient supply is increased [18] because nutrient cycling in hollow-hummock microtopography is different. Some studies show that drier environments and hummocks in fens typically have faster nutrient cycling [35] and nitrogen release [36]. In fen systems, hydrological gradients associated with microtopography strongly influence nutrient availability and plant performance, often overriding nutrient enrichment effects observed at the habitat scale [15,28].
Although our studies covered natural fen with changing topographic types, it is important to note that the expansion of drier areas can be accelerated by human activity [1]. H. vernicosus displays fragmented and human-sensitive populations throughout Europe, with its local distribution being strongly influenced by wetland hydrology [37]. Drainage of wetlands enhances the processes when, similar to the formation of hummocks, the top peat layer loses contact with mineral-rich groundwater. Drained fens develop more homogenised areas dominated by dry-adapted mosses, instead of heterogenic fen areas with changing hummock-hollow microtopography [30]. In such conditions, C. cuspidata will have an advantage over H. vernicosus, as stated in hummocky areas by us.
4. Materials and Methods
4.1. Study Site and Experimental Design
The experimental study was conducted at the Bražuolė fen, in the Trakai district, Lithuania (Figure 3). This site is situated along the right bank of the Bražuolė River and represents a sparsely populated area characterised by low-intensity land use. The fen supports naturally occurring populations of H. vernicosus, making it a suitable location for transplantation and monitoring studies. The habitat is predominantly wet, with high groundwater levels and a mosaic of hummock, hollow and lawn microtopographies, providing a range of microhabitats relevant for assessing species-specific expansion patterns. Its relatively undisturbed condition allows for evaluation of natural responses to experimental interventions without substantial confounding anthropogenic impacts.
The research was carried out from 2019 to 2021. Four study areas were selected within the fen to represent different topographical and hydrological conditions across the site (Figure 4). Two study areas were dominated by bryophyte lawns associated with a consistently high groundwater table; one study area exhibited flat, continuous bryophyte lawns up to two-thirds submerged in water (topography type I), whereas the other—displayed abundant surface, interspersed with bryophyte lawns just above the surface water or low hummocks (5–10 cm high) (topography type II). The remaining two sites were characterised by hummock–hollow microtopography, with more elevated, densely vegetated hummocks. One of these hummocky sites exhibited water-filled interhummock hollows (topography type III), whereas the other displayed comparatively drier interhummock areas with limited surface water presence (topography type IV). The selected study areas reflected a range of typical fen microhabitats, from wetter, more open lawns to drier, more hummock-dominated zones, capturing the natural heterogeneity of fen ecosystems. The characteristics of each study area are summarised in Appendix A. In each study area, 12 long-term study plots were established, with stakes placed at the corners to mark plot boundaries for future identification. The plots were categorised as follows: (a) four plots dominated by H. vernicosus, (b) four plots dominated by C. cuspidata, (c) four control plots: two of which were dominated by H. vernicosus, and two by C. cuspidata. The study plots covered different microtopography types, where both species were present: submerged lawns (topography type I), lawns or low hummocks above water level (topography type II), hummocks surrounded by spring waters (topography type II) and comparatively dry interhummock areas (topography type IV). In total, 48 study plots were created across all areas. Each study plot measured 30 × 30 cm and was divided into nine equal subplots of 10 × 10 cm. Within each subplot, the following parameters were recorded:
- (a)Total herb cover (%);
- (b)Surface water cover (%);
- (c)Total bryophyte cover (%);
- (d)Species composition of all bryophytes and their individual cover (%).
Schematic representation of topography types I–IV.
A total of 432 subplots were assessed across all study plots. The experimental design involved transplanting vegetation from the central subplot of each study plot to the central subplot of a plot dominated by the opposite species, creating a reciprocal transplant between H. vernicosus and C. cuspidata (Figure 5). Specifically, a central subplot from a study plot dominated by H. vernicosus was moved to a study plot dominated by C. cuspidata, and vice versa. The study plots were monitored from 2019 to 2021. The experimental plots were initially planted in April, with subsequent observations and recordings carried out in June and September of the same year. In 2020, additional monitoring took place during spring and the second half of summer, and in spring 2021. In order to compare temporal changes in the abundance of C. cuspidata and H. vernicosus under natural conditions, transplantation was not carried out in the control plots.
4.2. Data Analysis
Changes were assessed in transplanted subplots and in surrounding subplots, and these were compared with subplots from control study plots. Separate analyses were conducted to evaluate changes within the central transplanted subplots. Additional analyses assessed whether the introduced species altered adjacent plots relative to the controls.
The PERMDISP test based on Bray–Curtis dissimilarities was used to assess whether the dispersion of transplanted H. vernicosus and C. cuspidata subplots across different topographic types was homogeneous. Distances to group centroids were calculated for each topography type, and significance was tested using 999 permutations. To evaluate differences in subplot composition (covering all components of the transplanted subplot) in the topographic types studied, a one-way PERMANOVA analysis was performed. The analyses were performed using the Bray–Curtis dissimilarity measure. A SIMPER (Similarity Percentage) analysis was performed to determine which components most influenced the changes in the transplanted subplot during the study period. Community compositional changes were quantified using the Bray–Curtis dissimilarity index based on percentage cover data of herbs, surface water, and individual moss species. Only those components whose cumulative contribution (cumulative %) accounted for up to 80% of the total dissimilarity were used for the analysis.
In addition, to illustrate differences in community structure, principal coordinate analysis (PCoA) plots were generated to depict the distribution of subplots across different types of topography during the study period.
The Mann–Kendall test was applied to test for temporal trends in the abundance of the transplanted species increased in the surrounding subplots following transplantation to the central subplot. To quantify the magnitude of change in H. vernicosus and C. cuspidata, absolute cover change was calculated as Δcover (ΔP) = final cover − initial cover, and the same procedure was applied to control subplots for comparison. A regression curve was used to visually display temporal changes in species cover in both the surrounding and control subplots, enabling direct comparison of trends between the experimental and control conditions.
All statistical analyses and graphical visualisations were conducted using PAST statistical software, version 5.3 [38], and additional figures were created in Microsoft Excel. Any p-values less than 0.05 were regarded as statistically significant.
Taxonomic nomenclature followed the checklist of bryophytes of Europe [39] and the World Checklist of Vascular Plants [40].
5. Conclusions
Our studies, conducted in topographically and micro-relief-wise different areas of the same fen that are not affected by external nutrient enrichment from the surrounding area, demonstrate the significant role of environmental condition alterations within fens in preserving and maintaining H. vernicosus populations. If the water regime maintains heterogeneity in fen microhabitats, C. cuspidata and H. vernicosus can coexist successfully without additional management to increase the abundance of the latter. This is especially true in flooded areas dominated by lawns or low hummocks compared to hummocky areas. However, the abundance and competitive characteristics of H. vernicosus may decline with changes in the water regime. Implementing supplementary management measures is solely necessary in fens where drier, hummocky areas are naturally or human-impact-dominated.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wochal D. Marcsz K. Barabach J. Bak M. Lementowicz M. The hen that vanished: The untold story of drainage and peat extraction in Bagno Chlebowo peatland with implications for nature conservation Glob. Ecol. Conserv.202561 e 0364710.1016/j.gecco.2025.e 03647 · doi ↗
- 2Ahmad S. Wang M. Bates A. Martini F. Regan S. Saunders M. Liu H. Mc Elwain J. Gill L. Flatlining fens? Small-scale variations in peat properties and microtopography as indicators of ecosystem homogenization Ecol. Indic.202517211331710.1016/j.ecolind.2025.113317 · doi ↗
- 3van Diggelen R. Middleton B. Bakker J. Grootjans A. Wassen M. Fens and floodplains of the temperate zone: Present status, threats, conservation and restoration Appl. Veg. Sci.2006915716210.1111/j.1654-109X.2006.tb 00664.x · doi ↗
- 4Hájek M. Horsák M. HájkováP. DítěD. Habitat diversity of central European fens in relation to environmental gradients and an effort to standardise fen terminology in ecological studies Perspect. Plant Ecol. Evol. Syst.200689711410.1016/j.ppees.2006.08.002 · doi ↗
- 5Andersen D.K. Nygaard B. Fredshavn J.R. Ejrnǽs R. Cost-effective assessment of conservation status of fens Appl. Veg. Sci.20131649150110.1111/avsc.12020 · doi ↗
- 6Lamers L.P.M. Vile M.A. Grootjans A.P. Acreman M.C. van Diggelen R. Evans M.G. Richardson C.J. Rochefort L. Kooijman A.M. Roelofs J.G.M. Ecological restoration of rich fens in Europe and North America: From trial and error to an evidence-based approach Biol. Rev.20159018220310.1111/brv.1210224698312 · doi ↗ · pubmed ↗
- 7Jiménez-Alfaro B. Aunina L. Carbognani M. DítěD. Fernández-Pascual E. Garbolino E. Hájek O. HájkováP. Ivchenko T. Jandt U. Habitat-based biodiversity responses to macroclimate and edaphic factors in European fen ecosystems Glob. Change Biol.2023296756677110.1111/gcb.1696537818677 · doi ↗ · pubmed ↗
- 8KalvaitienėM. JukonienėI. The ecological drivers of the mosaic structure of bryophyte and vascular plant cover in the rich fens of Lithuania Plants 202514266210.3390/plants 1417266240941825 PMC 12429954 · doi ↗ · pubmed ↗