Genetic Diversity and Excretion Kinetics of Enteroviruses Excreted by Patients with Primary Immunodeficiency in Tunisia over a Five-Year Period (2020–2024)
Imene Ben Salem, Haifa Khemiri, Marwa Khedhiri, Najla Mekki, Marie-Line Joffret, Nadia Driss, Ilhem Ben Fraj, Monia Ben Khaled, Ines Ben Mrad, Mohamed-Ridha Barbouche, Henda Touzi, Zina Meddeb, Monia Ouederni, Maël Bessaud, Imen Ben Mustapha, Henda Triki, Sondes Haddad-Boubaker

TL;DR
This study analyzed enterovirus excretion in Tunisian patients with primary immunodeficiency, revealing diverse serotypes and long excretion periods.
Contribution
The study reports two previously unreported enterovirus serotypes in Tunisia and identifies a potential recombination event among coxsackieviruses.
Findings
EV excretion kinetics ranged from 30 to 946 days in Tunisian PID patients.
Two previously unreported serotypes, Coxsackievirus A5 and Echovirus type 19, were detected in Tunisia.
A possible recombination event was identified involving sequences from Coxsackievirus B5, A9, and B1.
Abstract
Enteroviruses (EVs) are small, non-enveloped RNA viruses that can cause diverse clinical outcomes, particularly severe in patients with primary immunodeficiency (PID) due to their impaired ability to clear infections. This study aimed to characterize EV excretion among 138 Tunisian PID patients over a five-year period, to identify circulating EV serotypes and assess their genetic diversity. A total of 558 stool samples were collected and analyzed by virus isolation and intratypic differentiation using RT-qPCR. Molecular typing was performed through Sanger sequencing of the VP1 region and whole genome sequencing using Next-Generation Sequencing (NGS) technologies. Phylogenetic analysis was conducted using the Maximum Likelihood (ML) method. EVs were detected in 55 stool samples from 23 patients. The excretion kinetics of EVs ranged between 30 and 946 days. Thirteen serotypes were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
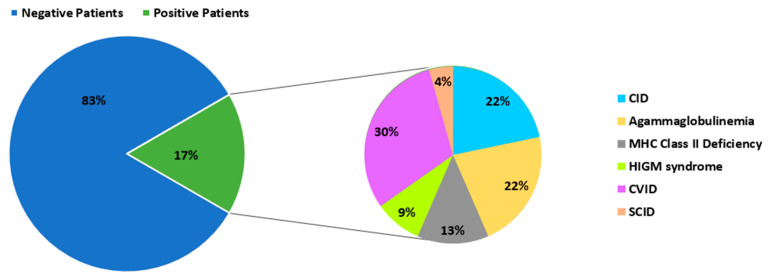 Figure 1
Figure 1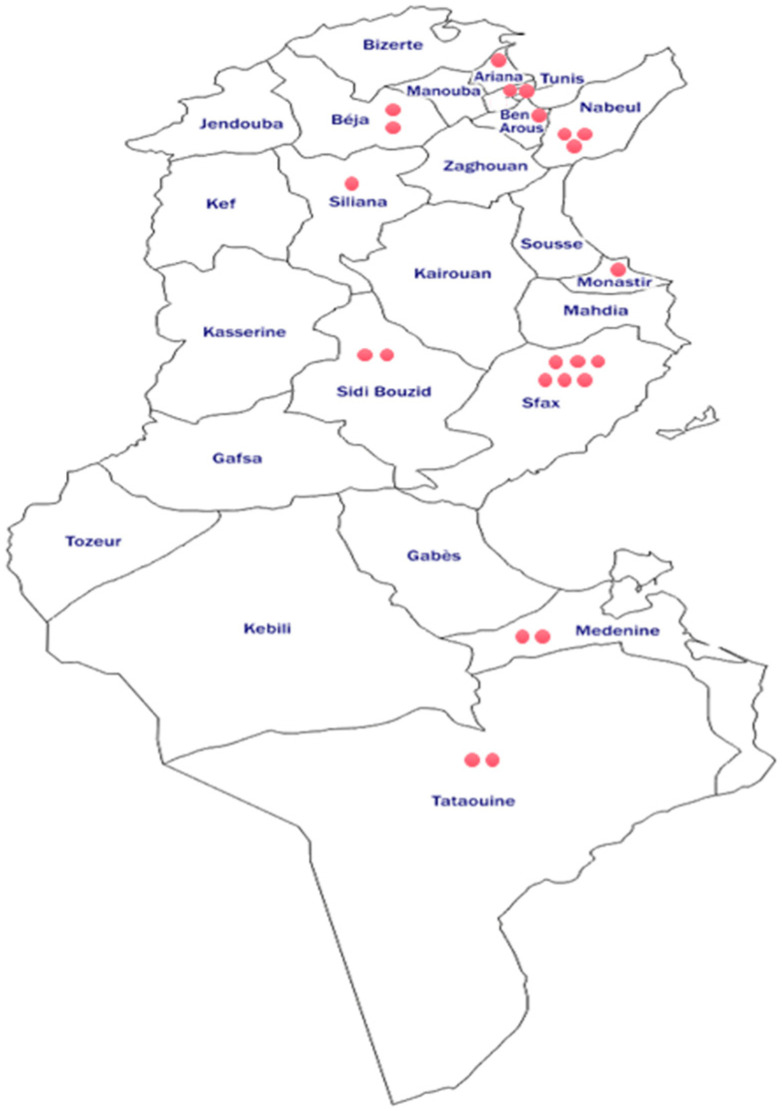 Figure 2
Figure 2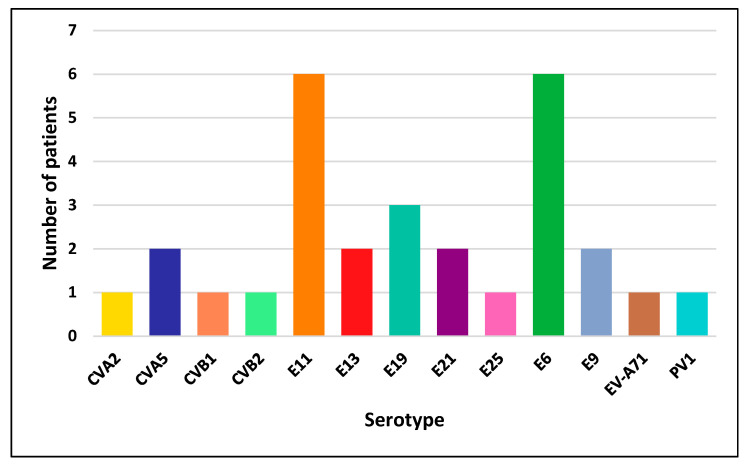 Figure 3
Figure 3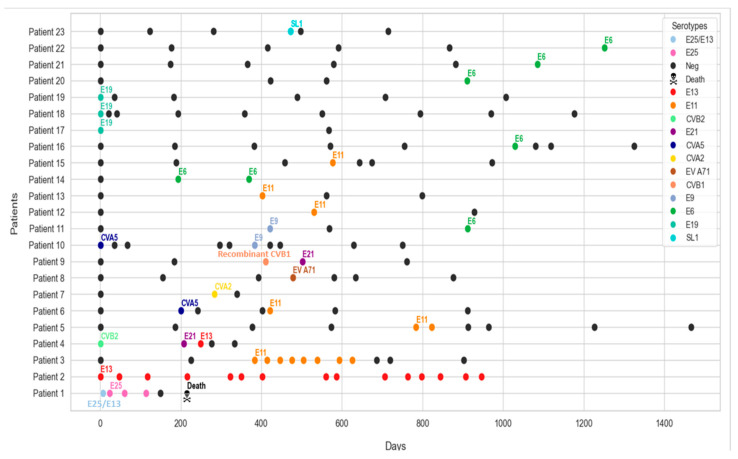 Figure 4
Figure 4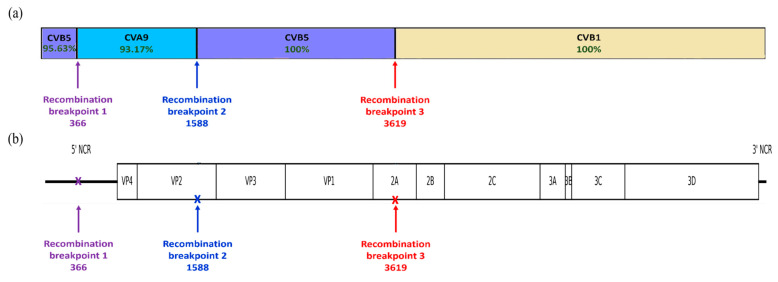 Figure 5
Figure 5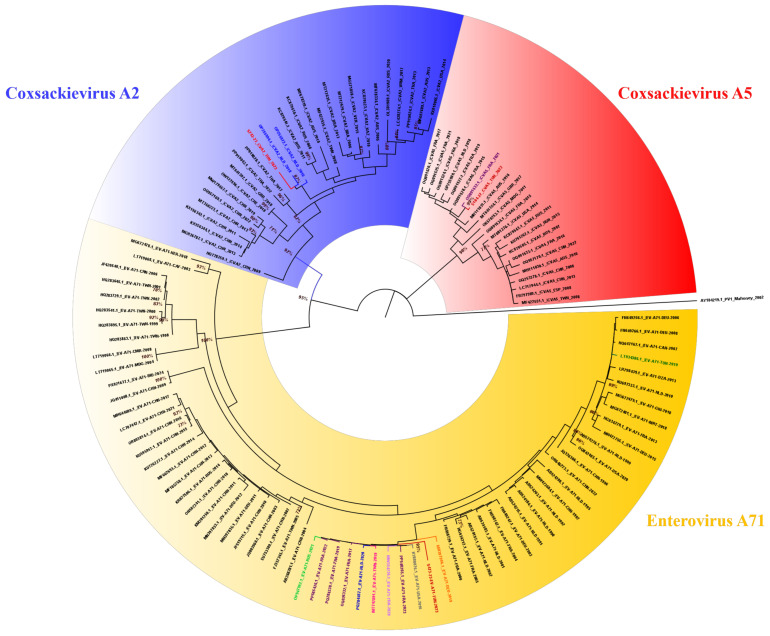 Figure 6
Figure 6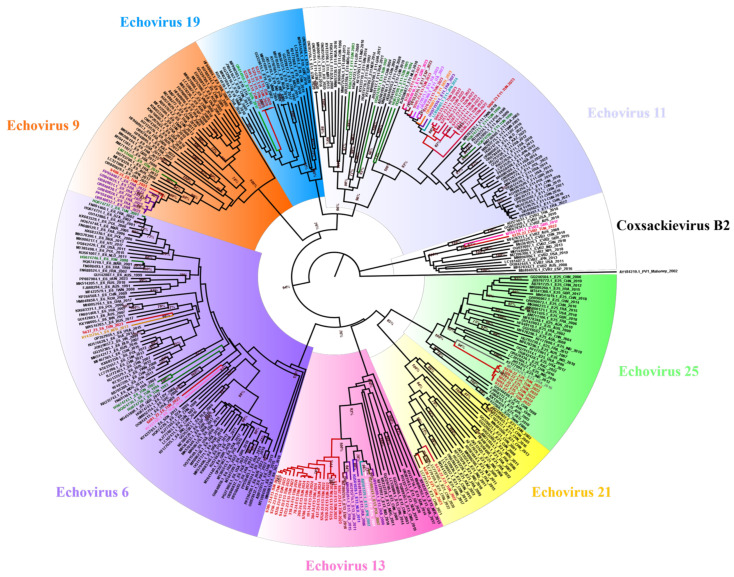 Figure 7
Figure 7- —Tunisian Ministry of Health
- —Tunisian Ministry of Higher Education and Research
- —World Health Organization, Tunisia office
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral Infections and Immunology Research · Respiratory viral infections research · Viral gastroenteritis research and epidemiology
1. Introduction
In the framework of the Global Polio Eradication Initiative (GPEI), surveillance activity of Poliovirus (PV) is based on the investigation of Polio suspected cases presenting with Acute Flaccid Paralysis (AFP), environmental surveillance in wastewater and surveillance of PV and Non-Polio Enterovirus (NPEV) excretion in patients with primary immunodeficiencies (PID) [1,2].
The genus Enterovirus (EV) is classified within the Picornaviridae family [3], which comprises small, non-enveloped viruses [4]. They are characterized by icosahedral symmetry and a single-stranded positive-sense RNA genome of approximately 7500 nucleotides in length [5]. The genus EV consists of 15 species that share common virion structure, genomic organization and replication mechanisms. Typically, enteroviral infections result in mild symptoms, but more severe clinical manifestations can arise, including myocarditis, AFP and various clinical disorders, particularly with PID patients who are at increased risk of developing recurrent, severe and life-threatening infections [6].
PIDs represent a genetically wide spectrum of disorders that affect specific components of the innate and adaptive immune system [7,8]. The overview of PID cases in Tunisia reveals an important prevalence influenced by the high consanguinity rate observed both in the country and across the Middle East and North Africa (MENA) region [9,10].
EV excretion among PID patients poses a significant challenge, as prolonged viral shedding in these individuals was documented. For instance, a study in India revealed that while the general prevalence of EV shedding in pediatric patients with PID was relatively low, one patient diagnosed with leaky Severe Combined Immunodeficiency (SCID) exhibited a long-term shedding of type 3 Immunodeficiency-Associated Vaccine-Derived PV (iVDPV) over a two-year period [11]. Notably, a patient from the UK with Common Variable Immunodeficiency (CVID) excreted type 2 VDPV for over 28 years [12]. These long-term excretors can harbor divergent PV strains and serve as reservoirs for neurovirulent PV strains that could compromise the efforts of the GPEI [13].
In Tunisia, despite decades of being a Polio-free country, PID individuals remain at a serious risk of persistent EV excretion, especially linked to the use of an oral PV vaccine (OPV). Research published in 2024 revealed the excretion of iVDPVs type 1 and 3 for periods of 134 and 271 days in 2009 and 2019, respectively, in patients with MHC class II deficiency. Rigorous genomic characterization of excreted strains revealed important intra-host diversity and recombinant strains excreted by patients with fatal outcomes [14]. In addition, a previous study carried out in 2012 showed that PID patients present an increased susceptibility to infections with NPEVs. Most cases resolved the excretion within weeks. The longest excretion period was detected in an MHC class II-deficient patient who excreted Echovirus type 11 (E11) over 528 days [15].
The epidemiology of NPEVs in Tunisia is characterized by fluctuation throughout the year, with periods of low and high transmission. Studies conducted between 1992 and 2017 reported frequent isolation of Echoviruses types 6, 11 and 30 [15,16,17]. Other serotypes showed cyclic occurrence, such as Coxsackieviruses type B (CVB), and others have been isolated episodically, mainly Coxsackieviruses type A (CVA) [16]. For PVs, the last cases of paralysis due to wild PV occurred in 1992, and the last wild isolate was detected in 1994. Since then, all PV isolates detected have been Sabin-like (SL) [18] or vaccine-derived [14,15].
In Tunisia, surveillance of EV excretion in PID patients was introduced in November 2023 as part of the National PV surveillance program, complementing the existing AFP surveillance. This study was conducted within the framework of the PID surveillance implementation. The studies carried out between 2005 and 2012 [15,16] provided partial insights into EV circulation among this population, and the present one expands the scope by providing a thorough follow-up of all Tunisian patients with PID, collected from the 24 Tunisian governorates, aiming to identify different EV serotypes, study their excretion kinetics and assess the genetic diversity of circulating strains.
2. Materials and Methods
2.1. Ethical Statement
This study was ethically approved by the Biomedical Ethics Committee of Pasteur Institute of Tunis (2019/20/I/LR161IPT/V1). Written informed consent was obtained from all patients or their legal guardians. Patient confidentiality was strictly maintained throughout the research process in accordance with the Tunisian and International ethical guidelines and legal requirements.
2.2. Studied Population and Sampling
A total of 138 Tunisian patients diagnosed with various types of PID received clinical follow-up over a 5-year period, from September 2020 to December 2024. These patients originated from all Tunisian governorates and presented Predominantly Antibody Deficiencies (PADs) such as Agammaglobulinemia, Common Variable Immunodeficiency (CVID) or Hyper-IgM syndrome (HIGM), and Combined Immunodeficiencies (CIDs) including Severe Combined Immunodeficiency (SCID) and MHC class II deficiency.
A total of 558 fecal specimens were collected in accordance with a meticulously established algorithm in collaboration with the World Health Organization (WHO). The study protocol includes a six-month follow-up for patients who do not excrete EVs, and a monthly follow-up for excretors, continuing until two consecutive negative results are obtained. The samples are transported to the Laboratory of Clinical Virology at Pasteur Institute of Tunis within a timeframe of 24 to 48 h at a temperature of +4 °C. Upon arrival, they are processed within 24 to 48 h and, thereafter, conserved at −20 °C for long-term storage. The integrity of the results is ensured through rigorous adherence to WHO standard protocols [19].
2.3. Virus Isolation in Cell Culture
Virus detection was carried out through specimen inoculation on cell culture, following the WHO standard protocols [19]. Stool suspensions were prepared and inoculated onto two distinct cell lines: RD cells (Human Rhabdomyosarcoma) and L20B cells (transgenic mouse cell line expressing the gene encoding the human cellular receptor for PV). The inoculated cells were incubated at 37 °C with daily monitoring for 10 days. The presence of a PV is suspected when a cytopathic effect (CPE) appears on L20B and RD cell lines, while NPEVs generally induce a positive culture result only in RD cell lines.
2.4. Intratypic Differentiation of Polioviruses
PVs, showing a CPE on L20B and RD cell lines, were further identified using intratypic differentiation (ITD), a multiplex real-time PCR enabling the identification of the serotypes of PV strains as well as their vaccine or wild origin, using the rRT-PCR 5.2 ITD kit (CDC, Atlanta, GA, USA) according to the manufacturer’s instructions.
2.5. Typing of NPEVs
NPEVs were typed using whole genome sequencing (WGS) and partial sequencing of the VP1 region when a WGS could not be obtained. Viral RNA extraction was performed using the QIAamp viral RNA mini kit (Qiagen, Germany) in adherence to the manufacturer’s instructions. All RNA extracts were either immediately used or stored at −20 °C for subsequent use.
Out of the 558 stool samples analyzed, 55 tested positive for EV and underwent WGS using Next-Generation Sequencing (NGS) technologies, notably Oxford Nanopore MinIon (Oxford Nanopore Technologies, Oxford, UK) and Illumina (Illumina inc., San Diego, CA, USA), or partially sequenced targeting the VP1 region with Sanger technology (Applied Biosystems, Foster City, CA, USA).
2.5.1. WGS Using Oxford Nanopore MinIon Technology
The extracted RNAs were reverse transcribed to cDNA using the LunaScript^TM^ RT SuperMix kit (New England Biolabs, Ipswich, MA, USA). The obtained cDNA was then amplified using 2 pools of “in-house” primers (Supplementary Table S1) targeting multiple regions of the EV genome. The resulting gene pools were purified using AMPure XP magnetic beads (Beckman Coulter, Brea, CA, USA) prior to quantification with the Qubit dsDNA High Sensitivity (HS) Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) to ensure optimal quality for DNA end repair and barcoding. Barcodes were ligated to the samples using Native Barcoding Expansion (Oxford Nanopore Technologies, Oxford, UK ). Following this step, the barcoded samples were pooled and purified using AMPure XP magnetic beads (Beckman Coulter, CA, USA). Adapters ligation was subsequently carried out, using Adapter Mix II (AMII) (New England Biolabs, MA, USA), followed by an additional purification step to ensure the removal of any unbound adapters and quantification. Finally, the prepared DNA was injected into the flow cell after checking on the number of pores available, to generate EV genome sequences on the MinIon MK1C sequencer (Oxford Nanopore Technologies, Oxford, UK ). Sequencing data were analyzed using Geneious Prime v2025.0.2 following a defined workflow. The analysis included read trimming to remove low-quality bases and adapters, mapping of filtered reads to EV reference genomes for consensus sequence reconstruction and genome coverage assessment, and annotation to identify functional regions.
2.5.2. WGS Using Illumina Technology
Illumina technology (Illumina inc., San Diego, CA, USA) was used to perform EV-positive genomes sequencing, following a previously described protocol. Briefly, for each specimen, the two PCR products were pooled and subsequently purified employing a vacuum-based approach, after which they were transferred to the sequencing platform PIBNet (Pasteur International Bioresources Network, Institut Pasteur Paris). The libraries were generated utilizing 1 ng of DNA with the Nextera XT DNA Library Preparation kit in a SureCycler 8800 thermocycler (Agilent Technologies, Santa Clara, CA, USA). Subsequent to purification on AMPure beads (Beckman Coulter, CA, USA), library quality was assessed using the High Sensitivity D1000 assay (Agilent Technologies) on a TapeStation 2200. Sequencing was carried out on an Illumina NextSeq HiSeq adopting a read depth of 50× (Illumina Inc., San Diego, CA, USA). All procedures followed the manufacturer’s instructions. Data analysis was carried out using CLC Genomics Workbench 8.5 to pair reads and assemble contigs from the raw sequencing data. Subsequently, de novo assembly was performed using CLC Main Workbench [20].
2.5.3. Partial Sequencing in the VP1 Region and Sequence Analysis
RNA extracts were processed by RT-PCR according to a previously described “in-house” protocol, using primers designed by Bailly et al. targeting the VP1-coding region of the genome [21], which is the major antigenic protein and the gold standard for EV typing [22].
The PCR products are purified using the QIAquick PCR kit (Qiagen, Dusseldorf, Germany) and subsequently sequenced bidirectionally on an ABI 3130 automated DNA sequencer (Applied Biosystem, ABI 3130, CA, USA) using the ABI PRISM BigDye Terminator Cycle Sequencing Ready Process kit (Invitrogen, CA, USA). The sequence data were subjected to analysis with Molecular Evolutionary Genetic Analysis (MEGA) software v7.0.26. The serotype was determined by comparing the acquired sequences to sequences available in the GenBank database. The Tunisian sequences were submitted to the GenBank database, and accession numbers are provided in the Supplementary Table S2.
2.6. Recombination Analysis
Full genome sequences were analyzed for potential recombination breakpoints using SimPlot Software v3.5.1, with default parameters (step size: 20 bp; GapStrip: on; 100 replicates; Kimura 2-parameter model; T/t ratio: 2.0; Neighbor-joining method).
2.7. Phylogeny
Due to the limited availability of complete EV genome sequences in the GenBank database and to ensure broad global representation of EV circulation, only the complete VP1 nucleotide sequences were used for the phylogenetic analysis. Sequences of interest were retrieved from the GenBank database (Date of access: January 2025) and covered a temporal range from 1991 to 2024. The duplicated ones were eliminated using the BLASTclust command “-i infile.fasta -o outfile.txt -p F -L 1 -S 99 -b T” to generate a non-redundant set of sequences. Thereafter, all sequences were aligned using the Mafft program “Multiple Alignment using Fast Fourier Transform” (https://mafft.cbrc.jp/alignment/server/index.html). Phylogenetic trees were constructed using MEGA Software v7.0.26. An appropriate outgroup sequence (Accession number: AY184219.1) was included to root the tree. The evolutionary history was inferred using the Maximum Likelihood (ML) approach, with a repetitive bootstrap number of 1000. Following tree reconstruction, sequences that were identical and originated from the same geographic location or were collected in the same or closely related years were further excluded to minimize sampling bias and temporal overrepresentation. The final number of VP1 sequences retained for each serotype is detailed in Table 1, ensuring a balanced and representative dataset for phylogenetic inference.
3. Results
3.1. Demographic and Clinical Data of EV-Positive Patients
The studied cohort comprised children (<15 years) (n = 106) as well as teenagers and adults (n = 32) with a median age of 11 years and a male predominance: 75 males (54%) vs. 63 females (46%). The investigated patients were diagnosed with various forms of PID: 95 patients had Predominantly Antibody Deficiencies (PAD), and 43 had Combined Immunodeficiencies (CIDs). PAD included Agammaglobulinemia (n = 33), Common Variable Immunodeficiency (CVID) (n = 43) and Hyper-IgM syndrome (HIGM) (n = 19). CID were classified into Severe Combined Immunodeficiency (SCID) (n = 13), MHC class II deficiency (n = 18) and other types of CID (n = 12). The distribution of PID patients is illustrated in Table 2.
Among the 138 PID patients, 23 tested positive for EV infection (17%), including one PV excreter and 22 patients shedding NPEVs. The EV-positive group showed a male-to-female ratio of 1.5 (14/9) and a median age of 6 years. Notably, five adult patients were identified as NPEV excreters, with ages spanning from 21 to 58 years. These EV-positive patients provided a total of 55 fecal specimens, corresponding to an overall positivity rate of 9.85%.
Within the various categories of PIDs, patients with CVID, including the single PV-positive case, accounted for 30% of EV-positive patients (n = 23). Agammaglobulinemia and CID each represented 22% of EV-positive cases, followed by MHC class II deficiency (13%). In contrast, HIGM (9%) and SCID (4%) were the least represented groups (Figure 1). Regarding EV infection rates within each PID category, CID patients exhibited the highest positivity rate (41.7%) (5/12), followed by patients with MHC class II deficiency (16.7%) (3/18), CVID (16.3%) (7/43), and Agammaglobulinemia (15.2%) (5/33). Lower EV positivity rates were observed among patients with HIGM (10.5%) (2/19) and SCID (1/13) (7.7.%). However, Fisher’s exact test showed no statistically significant association between PID subtype and EV positivity (p > 0.05). Geographically, the majority of excreters originated from coastal governorates, with the highest rates reported in Sfax (26%) and Nabeul (13%) (Figure 2).
3.2. Serotype Diversity
A total of 13 distinct serotypes were isolated among the EV-positive patients. The group EV-B was the most prevalent, including Echoviruses type 13, 11, 19, 21, 25, 6, 9 and Coxsackievirus B2. The group EV-A, which includes Coxsackieviruses A2 and A5 and EV-A71, was less common, similarly to EV-C, represented only by PV1. Overall, Echoviruses type 11 and 6 were the most frequently observed, involving six patients for each serotype, followed by Echovirus type 19 (three patients) and Echoviruses type 13, 21 and E9 and Coxsackievirus A5 (two patients) for each serotype. Other serotypes, including Coxsackievirus A2, Coxsackieviruses B1-2, Echovirus type 25, EV-A71 and PV1, were detected less commonly in only one patient (Figure 3). Importantly, some patients excreted more than one serotype during the study period; therefore, they were counted in more than one category. Consequently, the numbers shown reflect the total number of patients excreting each serotype, and not the number of unique patients.
3.3. Excretion Kinetics of EVs
Among the 23 EV excreters identified herein, the majority (n = 20) exhibited short-term viral excretion (defined as an excretion period shorter than 6 months), with shedding periods ranging from 30 days (one month) to 114 days (approximately 4 months). Among them, one patient excreted PV for a period of one month.
Two patients showed prolonged excretion of EVs for periods exceeding 6 months. Patient 3 excreted Echovirus type 11 for 242 days. Patient 2 represented an exceptional case with prolonged excretion of Echovirus type 13 for up to 946 days (more than 31 months), representing the longest shedding period recorded in this cohort.
Five patients presented enhanced susceptibility to the excretion of multiple EV serotypes. Patient 4, with CID, excreted several serotypes, of which Echoviruses types 13 and 21, and Coxsackievirus B2 (CVB2) over a period of 249 days (approximately 8 months). Additionally, patients 6, 9, 10 and 11 were infected with multiple EV serotypes during the study period at different time points. Notably, co-infections were reported. Indeed, patient 1 experienced a co-infection with two different serotypes, Echoviruses type 25 and 13, on day 7. Thereafter, he continued to excrete only Echovirus type 25 until virus clearance and death after a shedding period of 114 days (Figure 4).
3.4. Genetic Characterization of PV Strain
The comparison of the obtained Tunisian strain with the reference strain under accession number AY184219.1 revealed the accumulation of eight mutations, as illustrated in Supplementary Figure S1. This corresponds to a mutation rate of 0.9%, which is below the threshold defining VDPV, consequently confirming that the Tunisian isolate is classified as Sabin-like and not a Vaccine-Derived Poliovirus (VDPV). This divergence rate corresponds to a possible excretion period of approximately 11 months or a potentially important mutation rate attributable to the immunodeficiency context. Remarkably, the identified mutations were not located within any of the known antigenic sites.
3.5. Recombination Event
Only one sample obtained on day 411 from patient 9 exhibited evidence of possible recombination events. Simplot analysis suggested the possible occurrence of recombination involving CVB5, CVB1 and CVA9 sequences (Figure 5a). The best match obtained with Basic Local Alignment Search Tool (BLAST) analysis indicated that the 5′ region (nt 1–366) showed 95.63% nucleotide identity with a CVB5 strain (accession number: MG845891.1). The first recombination breakpoint was identified at nt 366, marking the transition from CVB5 to CVA9. The intermediate fragment spanning nt 367–1588 showed 93.17% nucleotide identity with CVA9 (accession number: OR678417.1). A second recombination breakpoint was detected at nt 1588, within the VP2 region, followed by a fragment from 1589 to 3619 showing 100% nucleotide identity with the CVB5 strain (accession number: MG845891.1). The third recombination breakpoint was located at nt 3619 within the 2A region (Figure 5b), after which the region extending from nt 3619 onward showed 100% nucleotide identity with a CVB1 strain (accession number: NC_001472.1). This recombination pattern was consistent with the Bootscan analysis, which revealed a clear shift in similarity profiles corresponding to the identified breakpoints (Supplementary Figure S2).
3.6. Genetic Diversity of NPEV Isolates
To investigate the genetic diversity of NPEVs, the complete VP1 sequences were considered.
Phylogenetic analysis of EV-A sequences (Figure 6, Supplementary Figure S4) was performed using 22 CVA5, 27 CVA2 and 66 EV-A71 sequences retrieved from the GenBank database (Supplementary Table S3). All Tunisian sequences from CVA5 and CVA2 clustered into groups of European sequences from different countries, supported by a robust bootstrap value of 99% and 87%, respectively. On the other hand, the Tunisian EV-A71 sequence isolated in 2023 belongs to genogroup E (Supplementary Figure S3) and is grouped with European, Asian and American (USA) strains isolated between 2016 and 2023. This cluster is strongly supported by a robust bootstrap value of 95%. In contrast, it was genetically distant from a previously reported Tunisian strain isolated in 2019. This phylogenetic pattern underscores the genetic heterogeneity of EV-A71 circulating strains and suggests different independent introduction events to Tunisia rather than autochthonous virus circulation.
Phylogenetic analysis of EV-B sequences (Figure 7, Supplementary Figure S5) was performed using a dataset comprising 75 E11 sequences, 16 CVB2 sequences, 39 E25 sequences, 26 E21 sequences, 37 E13 sequences, 89 E6 sequences, 40 E9 sequences and 22 E19 sequences (Supplementary Table S4). Tunisian E25 sequences grouped closely with a U.S strain detected in 2016, supported with a robust bootstrap value of 94%. Tunisian CVB2, E9 and E19 sequences clustered with European sequences. E6 clustered with European and African sequences. Similarly, Tunisian E21 and E11 sequences clustered with both European and Asian sequences, with a robust bootstrap value of 96% and 83%, respectively. Notably, the Tunisian strains showed genetic divergence from previously reported Tunisian sequences, suggesting the introduction of new viral lineages or evolutionary divergence over time.
4. Discussion
This study provides a comprehensive overview of PV and NPEV excretion among all patients with PID recorded across the 24 Tunisian governorates over a five-year period from 2020 to 2024. Our objectives are to shed light on viral excretion dynamics, assess serotype diversity and describe epidemiological patterns of excreted PVs and NPEVs.
A total of 138 Tunisian PID patients were enrolled in this study, including 95 patients with PAD (Predominantly Antibody Deficiency) and 43 with CID (Combined Immunodeficiency), as both conditions result in immunoglobulin deficiencies and can be associated with prolonged viral excretion. In PAD, the absence or dysfunction of B cells leads to impaired production of neutralizing antibodies, which are crucial for viral clearance. In contrast, CID affects both T- and B-cell compartments, compromising not only antibody production but also cytotoxic T cell-mediated viral elimination.
Among patients, 23 (17%) tested positive for EV infections, with a median age of 6 years, including five adults (22%) aged between 21 and 58 years. In a previous study conducted in 2012 and targeting 82 PID patients, 11 EV excreters were reported, including 2 adults aged 24 and 26 [15]. Indeed, the persistent immune system dysfunction can predispose adults to chronic or recurrent EV infections. Thus, it is relevant, in the framework of PID surveillance, to investigate all age groups in contrast to AFP, which may concern only children younger than 15 years. Interestingly, the geographic distribution of EV-positive patients revealed a clustering in the governorates of Sfax (26%) and Nabeul (13%). This finding aligns with previous national surveillance data reporting higher EV circulation in urban and coastal regions, likely reflecting increased population density and improved access to healthcare facilities. It may also be linked to higher rates of PID in these areas in association with the important reported consanguinity rates [16].
Among the EV-positive patients, CVID accounted for the largest proportion of cases (30%), followed by Agammaglobulinemia and CID (22% each). However, the proportion of EV-positive cases alone does not reflect true differences in susceptibility to infection. When infection rates were analyzed within each PID category, patients with CID exhibited the highest EV-positivity rate (41.7%), suggesting a potentially increased susceptibility to EV infections in this group. Nevertheless, given the limited number of patients in PID categories, these findings should be interpreted with caution, as the statistical power to detect significant intergroup differences remains limited. These observations are at odds with earlier reports, such as the study elaborated by Mohanty et al. in India, where CVID stands out as the most common type of PID among EV-positive patients (6 patients out of 29) [23]. This disparity can be the result of variations in cohort composition, diagnostic procedures, or surveillance strategies across populations. The relatively low rates of EVs infection observed in patients with SCID (4%) and HIGM (9%) may be linked to reduced life expectancy, as these patients rarely survive beyond one year of age. Earlier diagnosis and hematopoietic stem cell transplantation can improve the prognosis and survival of these patients [15,24].
The detection of only a single case of SL1 PV, for a possible period of 11 months, aligns with the Tunisian patterns. The circulation of VDPVs remains limited, and only SL strains have become predominant as a result of sustained high levels of vaccination coverage [25]. The latest cases of iVDPVs excreters identified in Tunisia, through surveillance of AFP, were received in 2009, 2016 and 2019 from three PID patients with MHC class II deficiency, with an excretion duration ranging from 44 to 271 days [14]. The infection rate of NPEV (17%) in our study was higher than in previous national studies. In 2012, the EV infection rate among PID patients was 13.4% [15]. In addition, in an 11-year period study conducted between 2007 and 2017, the prevalence of PV and NPEV in PID patients was 6.8% and 12.4%, respectively [17]. This increase could be linked to the enhanced surveillance programs, the quality of the samples, in addition to the intensified screening of high-risk groups.
The distribution of the 12 NPEV serotypes identified in our study highlights the important EVs diversity in our country, with the detection of two previously unreported serotypes: Coxsackievirus A5 (CVA5) (n = 2) and Echovirus type 19 (E19) (n = 3) [16,17,26]. This may reflect the natural diversity of EV circulation patterns as well as the importance of ongoing molecular surveillance in the detection of uncommon serotypes that may go unnoticed. The predominance of species B among the identified NPEVs in the present study was reported in other countries in Africa, Asia and Europe [27,28,29]. Echoviruses type 11, 6 and 19 were more frequently isolated. In contrast, other serotypes were sporadically isolated during the study course, including Coxsackieviruses A2 and B1-2, as well as Echoviruses type 25 and EV-A71. These trends reflect the inherent instability of EV epidemiology, driven by factors such as herd immunity changes and global population movements. Large-scale outbreaks in Europe and Asia have demonstrated that changes in population-level immunity can result in the replacement of serotypes or the reappearance of previously rare EV types [30,31]. In addition, a possible recombination event involving CVB5, CVB1, and CVA9 was identified with an important nucleotide identity percentage (≥95%). This genomic configuration suggests sequential recombination events between EV-B and EV-A serotypes, reinforcing the well-established role of recombination as a major evolutionary mechanism within the EV-A and B species. These findings are consistent with previous studies reporting inter- and intra-serotypic recombination among circulating CVB and CVA9 strains, which may contribute to genetic diversification, viral fitness, and the emergence of novel variants [32]. These patterns underscore the critical need for strengthened EV surveillance and tailored management approaches in PID populations, in light of evolving immunization policies and regional disparities in healthcare infrastructure. This shift in the epidemiological landscape, especially after the eradication of types 2 and 3 wild PVs (WPV) [33], creates a favorable environment for the transmission and circulation of NPEVs [31,34].
The investigation of EV excretion kinetics in patients with PID revealed short-term excretion among the majority of the excreters (20 out of 23), with viral clearance occurring within a limited time frame. Nevertheless, two patients exhibited prolonged excretion lasting about 8 months. Patient 3, diagnosed with Agammaglobulinemia, excreted Echovirus type 11 for 242. Atypically, patient 2, diagnosed with CVID, excreted Echovirus type 13 for an extended period up to 946 days (approximately 31 months). In fact, there has been an association between CVID and long-term shedding of EVs. Notably, all chronic EV excretors recorded in the WHO registry have been diagnosed with CVID [35]. This finding is anticipated, given that CVID consists of a broad spectrum of associated pathologies and longer life expectancy compared to other types of PIDs, which contribute to the likelihood of prolonged EV excretion [36,37]. In Tunisia, the longest recorded shedding period of EVs was reported in a 4-year-old child with MHC class II deficiency, who excreted Echovirus type 21 for 588 days [15]. There have been reports of prolonged EV shedding in pediatric patients with PID globally, with the duration varying depending on individual immune status. One of the extremely chronic shedding periods involved a child diagnosed with CVID in the United Kingdom, who excreted iVDPV2 for over 28 years, the longest duration ever reported [12]. In Iran, prolonged iVDPV2 shedding has also been observed for a period lasting up to 720 days [13]. Additionally, an Indian patient with SCID was identified to have shed type 3 iVDPV for two years [11].
Several patients in our cohort (n = 6) exhibited shedding of numerous EV serotypes either consecutively or simultaneously. This high EV susceptibility may be attributed to impaired immune responses to clear infections or continuous exposure within a high-transmission environment. Notably, patient 1 showed concomitant infection with Echoviruses type 25 and 13 during the first week of infection, before virus clearance 114 days later, and death. Similarly, patients 4, 6, 9, 10, and 11 experienced multiple shedding episodes. Particularly, patients 6 and 11 excreted various EV serotypes over time, suggesting possible reinfection, viral reactivation, or prolonged subclinical persistence. This pattern of recurrent or overlapping EV infections has also been reported in previous studies involving PID patients [38,39]. In addition, patients were more prone to be vulnerable to the excretion of EV-B group serotypes, including predominantly Echoviruses type 6, 11 and 19. Several factors may account for the higher frequency of these serotypes. In fact, the EV-B group is known to circulate widely in many countries and is frequently associated with both gastrointestinal and systemic infections. As a result, patients are likely to experience repeated exposure. Furthermore, EV-B viruses have been reported to display efficient replication in intestinal epithelial cells and may possess specific determinants that facilitate persistence and prolonged shedding. Immunodeficiency also plays a crucial role. In patients with PID, viral clearance may be delayed, allowing these serotypes to be detected more often than others. Overall, the important detection of E6, E11 and E19 probably reflects an interplay of virological, immunological and epidemiological factors rather than a single mechanism [40,41]
In order to further characterize the isolated NPEV serotypes and determine their indigenous or imported origin, a phylogenetic analysis was carried out for all serotypes. The phylogenetic pattern indicates multiple and diverse introductions of NPEV strains from different geographic origins over time, especially of European, African, American and Asian origin [42,43]. This could be explained by the important commercial, cultural and tourist exchanges between Tunisia, the European countries and certain Asian nations. The EV-A71 strain identified in Tunisia was found to belong to genogroup E. This genogroup has previously been reported only in West and Central Africa, including Niger, Cameroon, the Central African Republic, Senegal, Guinea, and Mauritania, suggesting that genogroup E may be indigenous to the African continent. The detection of this lineage in Tunisia expands its known geographic range and highlights the potential for wider circulation of this genogroup across Africa [44]. Moreover, the absence of a distinct clade including Tunisian sequences suggests the lack of continuous endemic circulation within the country and reflects sporadic importation events rather than persistent local transmission. This highlights the dynamic nature of EV circulation, likely influenced by human mobility and regional connectivity.
5. Conclusions
In conclusion, the present study provides valuable insights into the excretion kinetics and the molecular diversity of EVs in Tunisia, thereby advancing our understanding of their genetic epidemiology and phylogenetic relationships. Through the molecular characterization and comparative analysis of circulating strains, this study provides an additional perspective on the dynamics of EV circulation across the country. These results enhance the current national and international database of EVs and provide a valuable foundation for upcoming epidemiological surveillance, molecular investigations, and public health strategies aimed at monitoring and controlling EV infections in Tunisia.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Global Polio Eradication Initiative. (n.d.). Global Polio Eradication—Eradicate Polio Now Available online: https://www.polioeradication.org/(accessed on 5 September 2025)
- 2Chumakov K. Kew O. The Poliovirus Eradication Initiative ASM Press e Books Washington, DC, USA 201444945910.1128/9781555816698.ch 28 · doi ↗
- 3Oberste M.S. Pallansch M.A. Enteroviruses and Parechoviruses Clinical Virology Manual 5th ed.John Wiley & Sons Hoboken, NJ, USA 201627729210.1128/9781555819156.ch 20 · doi ↗
- 4Wang S. Wang K. Zhao K. Hua S. Du J. The structure, function, and mechanisms of action of enterovirus non-structural protein 2C Front. Microbiol.20201161596510.3389/fmicb.2020.61596533381104 PMC 7767853 · doi ↗ · pubmed ↗
- 5Cifuente J.O. Moratorio G. Evolutionary and Structural overview of human picornavirus Ca PSID antibody evasion Front. Cell. Infect. Microbiol.2019928310.3389/fcimb.2019.0028331482072 PMC 6710328 · doi ↗ · pubmed ↗
- 6Modrow S. Falke D. Truyen U. Schätzl H. Viruses with Single-Stranded, Positive-Sense RNA Genomes Springer e Books Heidelberg, Germany 201318534910.1007/978-3-642-20718-1_14 · doi ↗
- 7Li L. Ivanova O. Driss N. Tiongco-Recto M. Da Silva R. Shahmahmoodi S. Sazzad H.M.S. Mach O. Kahn A. Sutter R.W. Poliovirus excretion among persons with primary immune deficiency disorders: Summary of a Seven-Country Study series J. Infect. Dis.2014210 S 368S 37210.1093/infdis/jiu 06525316857 · doi ↗ · pubmed ↗
- 8Geha R.S. Notarangelo L.D. Casanova J. Chapel H. Conley M.E. Fischer A. Hammarström L. Nonoyama S. Ochs H.D. Puck J.M. Primary immunodeficiency diseases: An update from the International Union of Immunological Societies Primary Immunodeficiency Diseases Classification Committee J. Allergy Clin. Immunol.200712077679410.1016/j.jaci.2007.08.05317952897 PMC 2601718 · doi ↗ · pubmed ↗