Genomic Characterization, Safety Assessment, and Probiotic Potential of a Novel Human-Derived Enterococcus lactis IOBRA9746
Dongxiong Hu, Jiao Yan, Yuxin Li, Jun Wu, Yifan Xie, Lirong Peng, Erhong Zhang, Hao Jiang, Qinmiao Sun, Xiao Chu

TL;DR
A new human-derived probiotic strain, Enterococcus lactis IOBRA9746, was found to safely reduce liver fat in mice, offering potential for treating diet-related metabolic diseases.
Contribution
The study introduces a novel BSH-active Enterococcus lactis strain with a demonstrated safety profile and liver-targeted lipid-lowering effect.
Findings
The strain showed potent bile salt hydrolase activity and no toxicity in mice.
It reduced hepatic lipid droplet accumulation without affecting serum cholesterol levels.
The probiotic modulates bile acid metabolism to inhibit liver fat deposition.
Abstract
Probiotic strains exhibiting bile salt hydrolase (BSH) activity represent a promising therapeutic strategy for ameliorating metabolic disorders via targeting the gut–liver axis. Herein, we characterized a newly isolated human-derived Enterococcus strain and investigated its therapeutic potential. Genomic analysis confirmed its safety profile, while in vitro assays demonstrated potent BSH activity. Subsequently, C57BL/6J mice fed a high-cholesterol diet were orally administered this strain over an eight-week intervention period. Although the treatment did not significantly reduce serum cholesterol levels, a marked reduction in hepatic lipid droplet accumulation was confirmed by H&E and Oil Red O staining. Mechanistically, the strain’s potent BSH activity likely modulates bile acid metabolism within the gut–liver axis, thereby specifically inhibiting hepatic lipid deposition.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
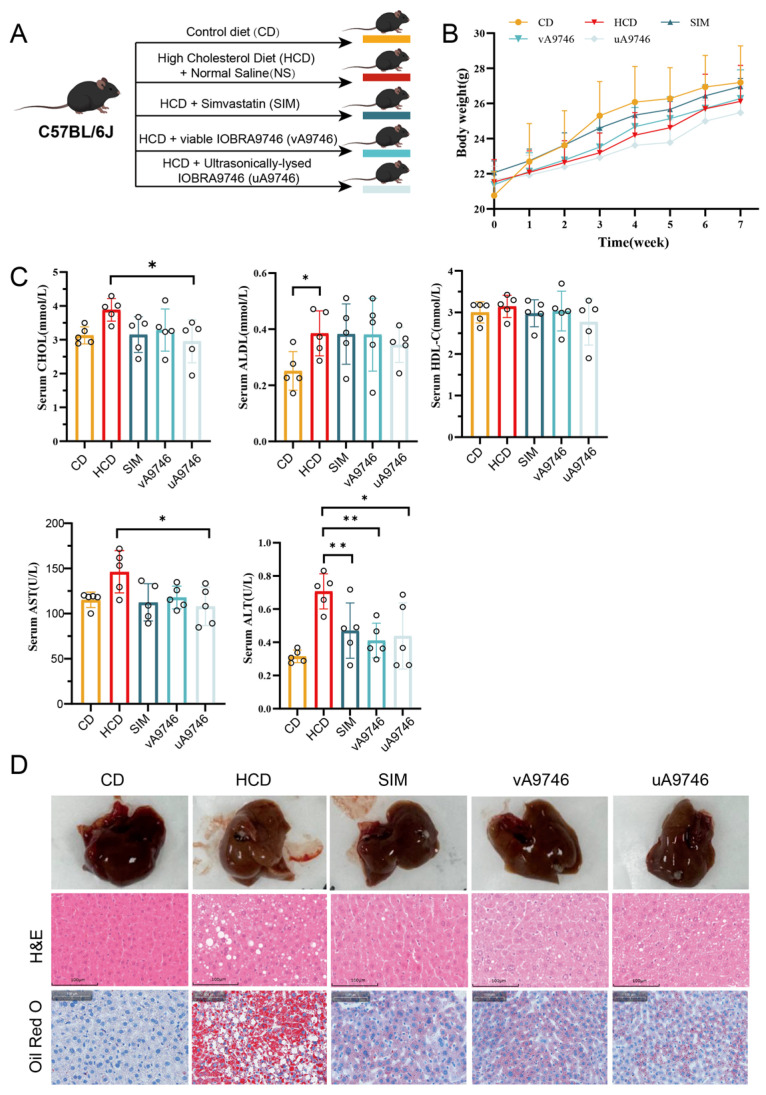 Figure 4
Figure 4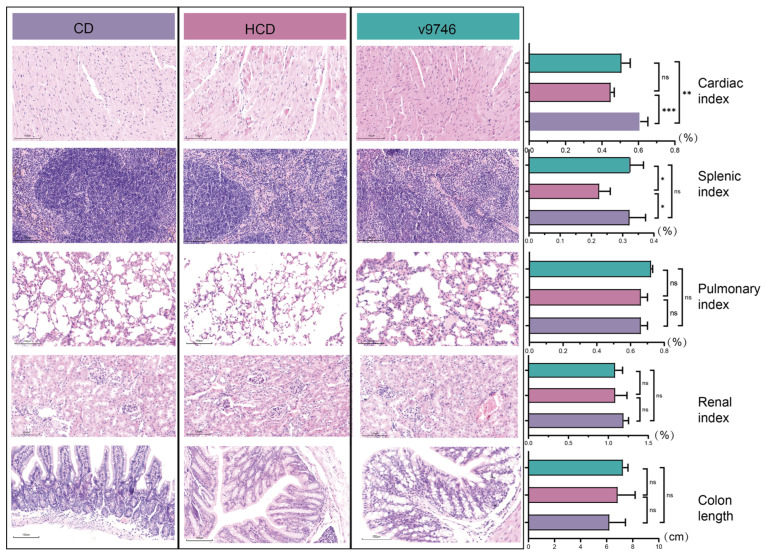 Figure 5
Figure 5- —Natural Science Foundation of Yunnan Province
- —China Postdoctoral Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Gut microbiota and health · Liver Disease Diagnosis and Treatment
1. Introduction
Suboptimal dietary patterns, particularly those rich in fats and cholesterol, significantly contribute to the pathogenesis of metabolic disorders including hyperlipidemia, cardiovascular disease (CVD), and non-alcoholic fatty liver disease (NAFLD) [1]. The liver serves as a central regulator in these processes, orchestrating cholesterol homeostasis, de novo lipogenesis, and the assembly/secretion of lipoproteins [2]. Notably, the gut microbiota has recently emerged as a critical environmental modulator of host metabolism and immunity via a complex bidirectional communication network known as the gut–liver axis. This axis operates primarily through the portal vein, allowing microbial metabolites and components to directly influence hepatic physiology [3]. Dysbiosis of the gut microbiota is intimately linked to the pathogenesis of various human diseases by promoting increased intestinal permeability, chronic low-grade inflammation, and disrupted energy metabolism [4]. Consequently, interventions aimed at restoring microbial balance, such as probiotic supplementation, have garnered significant attention as non-pharmacological strategies to enhance hepatic health and ameliorate metabolic dysfunction [5].
A pivotal mechanism by which certain probiotics exert cholesterol-lowering and lipid-reducing effects involves the enzymatic activity of bile salt hydrolase (BSH) [6]. Within the intestinal lumen, BSH catalyzes the deconjugation of conjugated bile acids (BAs) into free primary or secondary forms. These deconjugated BAs exhibit reduced solubility and are consequently excreted more readily in feces, thereby disrupting the enterohepatic circulation of bile acids. This interruption triggers hepatic upregulation of de novo bile acid synthesis from endogenous cholesterol pools [7]. Collectively, this cascade establishes a direct mechanistic link between gut microbial activity and hepatic cholesterol catabolism, positioning BSH-active probiotics as promising therapeutic agents for targeted amelioration of hepatic lipid accumulation.
The genus Enterococcus, comprising Gram-positive lactic acid bacteria, represents a prevalent commensal population within the human gastrointestinal tract and serves as a significant reservoir of BSH-producing strains. Notably, human-derived isolates warrant particular attention due to their intrinsic adaptation to the host milieu [8]. However, select members of this genus function as opportunistic pathogens harboring virulence determinants and antibiotic resistance genes, thereby presenting significant biosafety concerns [9]. E. lactis, a well-characterized and safety-validated species within the Enterococcus genus, was first isolated from Italian raw milk cheeses by Morandi et al. (2012) [10]. Subsequently, Ahmed et al. (2023) [11] further assessed the probiotic properties and safety profile of human-derived E. lactis strains 10NA and 50NA. Comprehensive whole-genome analysis confirmed the absence of transferable antibiotic resistance genes in these isolates. Additionally, cell culture assays indicated no significant cytotoxicity of their metabolites toward Caco-2 cells.
Previous studies have established significant BSH activity in Enterococcus species [11]. Nonetheless, existing research on E. lactis remains predominantly focused on in vitro safety assessments and genomic analyses, with limited exploration of its in vivo lipid-modulating efficacy and systemic safety profiles [11,12]. In this study, we isolated an E. lactis strain IOBRA9746 from the intestinal tract of a healthy adult donor and performed taxonomic identification and genomic safety evaluation, including screening for putative virulence and mobile genetic elements. The probiotic potential of this human-derived strain is primarily attributed to its functional BSH activity, which plays a pivotal role in bile acid metabolism and lipid homeostasis. To comprehensively evaluate its impact on lipid metabolism, we implemented an 8-week dietary intervention model in which mice were fed a high-cholesterol diet. Given the central role of the liver in cholesterol metabolism and lipid regulation, a comprehensive set of assessments was employed, including serum lipid profiling to monitor systemic alterations, liver function tests to assess hepatic health status, and histological analyses via H&E and Oil Red O staining to directly visualize hepatic lipid accumulation. Furthermore, a thorough safety assessment was conducted through histopathological examination of multiple organs to ensure the strain’s suitability for potential probiotic applications.
2. Material and Methods
2.1. Sample Collection and Strain Isolation
The feces sample was collected from a healthy adult in Kunming city, Yunnan Province, China, who had not been administered probiotics or antibiotics for at least three months. The study protocol was reviewed and approved by the Human Research Materials Ethics Committee of Yunnan University (Approval No. CHSRE2023020; Approval Date: 10 March 2023). Written informed consent was obtained from the donor prior to sample collection. Samples were collected in 50 mL sterile centrifuge tubes and immediately transported to the laboratory. One gram of fecal sample was homogenized in sterile saline and serially diluted to 10^−4^. To maximize the recovery of potential probiotic candidates, aliquots (100 μL) of the diluted samples were plated onto a diverse set of selective and non-selective media, including MRS Medium (Solarbio, Beijing, China), Briggs Medium Supplemented (Hopebio, Qingdao, China), Bifidobacterium Medium (Hopebio, China), and BS Medium (Hopebio, China). Notably, Enterococcus species were successfully isolated from various media types, including those targeting other genera, which reflects their inherent environmental robustness. Specifically, strain IOBRA9746 was isolated from Bifidobacterium medium and purified by subculturing on MRS agar for 2–3 passages to ensure consistent colony morphology. Purified isolates were preserved in 25% (v/v) glycerol (Sigma-Aldrich, St. Louis, MO, USA) at −80 °C. The 16S rRNA gene fragment was amplified using the purified single colony as a template with primers 27F and 1492R. The PCR product was sent to Sangon Biotech for sequencing, and subsequent sequence alignment was performed using the EzBioCloud database (11 September 2024) [13].
2.2. Whole-Genome Sequencing and Annotation
Genomic DNA of the strains was extracted using the FastPure BacteriaDNA lsolation Mini Kit-BOX2 Kit (Vazyme, Nanjing, China), following the manufacturer’s protocol to ensure high-quality DNA suitable for sequencing. The extracted DNA was subjected to quality control: purity was assessed by 1% agarose gel electrophoresis, and concentration was measured using a Qubit 4.0 fluorometer with the dsDNA HS Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA), following the manufacturer’s instructions. Whole-genome sequencing was conducted using a hybrid approach combining long-read sequencing via the PacBio Sequel II platform and short-read sequencing on the MGIseq-2000 platform to ensure high-contiguity assembly. Raw sequencing reads from the short-read platform were first processed using fastp (v0.12.4) to trim adapters and filter low-quality sequences. A hybrid de novo assembly was then performed using Unicycler (v0.5.1) to generate a high-quality, circularized consensus sequence. The integrity and quality of the final genome assembly were comprehensively evaluated. CheckM (v1.0.12) was employed to estimate the completeness, contamination, and strain heterogeneity [14]. Key assembly metrics, such as N50, L50, and the total genome size, were calculated using QUAST (v5.1.0rc1) [15]. The assembled genome was subsequently annotated. Initial structural annotation, including the prediction of coding sequences (CDSs), tRNAs, and rRNAs, was carried out with Prokka (v1.14.6) [16]. For a comprehensive functional characterization, the predicted protein sequences were analyzed using eggNOG-mapper (v2.1.12) [17] against the eggNOG database (v5.0.2) to assign functions based on the Gene Ontology (GO) [18], KEGG [19], and Clusters of Orthologous Groups (COG) databases [20]. To assess the genomic safety profile of the strain, putative virulence factors were identified by searching against the Virulence Factor Database (VFDB; core dataset, 2025) [21] and the Pathogen-Host Interactions database (PHI-base, v4.18) [22]. Antimicrobial resistance (AMR) genes were identified using the Resistance Gene Identifier (RGI, v6.0.3) with the CARD database (v3.2.8) [23] and confirmed with ResFinder (v4.7.2) [24]. Finally, the genome was scanned for mobile genetic elements, such as insertion sequences (ISs) using Isfinder (29 September 2024) [25], and potential genomic islands using IslandViewer 4 [26].
2.3. Phylogenomic and Genetic Analysis
The phylogenetic tree based on the 16S rRNA gene sequences of the top 50 closely related strains was constructed using MEGA 10.2.6 software with the neighbor-joining, maximum likelihood, and maximum parsimony methods [27]. To determine the phylogenomic position and taxonomic classification of strain IOBRA9746 with related taxa, a phylogenomic tree was constructed based on a set of conserved single-copy genes. A total of 102 type strains of closely related Enterococcus species were obtained from the GTDB-Tk database (v2.4.0) (accession numbers are listed in Supplementary Table S2) [28]. The set of 120 bacterial single-copy marker proteins was extracted from each genome using the GTDB-Tk identify module. Each orthologous protein set was then individually aligned using MAFFT (v7.526) [29], and the resulting alignments were trimmed with trimAl (v1.4) [30] to remove poorly aligned regions. A maximum likelihood (ML) phylogenomic tree was inferred with IQ-TREE (v2.2.0) [31]. The best-fit substitution model was determined by ModelFinder, integrated within IQ-TREE. The robustness of the tree topology was assessed by performing 1000 ultrafast bootstrap (UFBoot) replicates. Finally, the resulting tree was visualized and annotated using the iTOL (v6) online tool [32].
2.4. Probiotic Property and Safety Assessment
The ability of strain IOBRA9746 to tolerate acidic stress and its growth profile across a range of temperatures were characterized by monitoring its growth dynamics [33]. To prepare the inoculum, an overnight culture grown in MRS broth at 37 °C was used. The inoculum was added (1%, v/v) into a series of fresh MRS broths, with the pH pre-adjusted to 3.5, 4.0, 5.0, and 6.0 using 1 M HCl. For the temperature profile assay, the cultures were incubated at 30 °C, 37 °C, 45 °C, and 55 °C. Bacterial proliferation was monitored by measuring the optical density at 600 nm (OD_600_) using a microplate reader (BioTek Instruments, Winooski, VT, USA). All experiments were performed in triplicate, and the average growth curves were plotted.
The capacity of the strain to auto-aggregate was determined by monitoring the sedimentation of a bacterial suspension [34]. Briefly, cells from an overnight culture were collected by centrifugation (4000× g, 10 min, 4 °C), washed twice with sterile PBS (pH 7.2), and finally resuspended in PBS. The cell density was standardized to an optical density (OD_600_) of 0.60 (A_0_). The suspension was then homogenized by vortexing for 10 s and left undisturbed at 37 °C. At specified time points (1, 3, and 5 h), a sample from the supernatant was carefully removed to measure its OD_600_ (A_t_). Aggregation was expressed as the percentage reduction in the supernatant’s optical density over time: Aggregation (%) = (1 − A_t_/A_0_) × 100%.
The cell surface hydrophobicity was quantified using the microbial adhesion to hydrocarbons (MATH) method, with xylene as the hydrocarbon phase. An overnight culture was harvested and resuspended in PBS to an initial optical density at 600 nm (OD_600_) of approximately 0.80 (A_0_). In total, 2 mL of the suspension was thoroughly mixed with an equal volume of n-xylene by vortexing at maximum speed for 2 min. The mixture was then incubated at room temperature for 30 min to allow for phase separation. Following incubation, 1 mL of the lower aqueous phase was carefully withdrawn, and its OD_600_ was measured (A_1_). The percentage of hydrophobicity was calculated as the partitioning of cells to the hydrocarbon phase, using the formula: Hydrophobicity (%) = [(A_0_ − A_1_)/A_0_] × 100%.
The co-aggregation ability of strain IOBRA9746 with the pathogenic bacteria was assessed using the following procedure [35]. The overnight culture was harvested and resuspended in PBS to an initial optical density at 600 nm (OD_600_) of approximately 0.80. For the assay, equal volumes (1 mL) of the probiotic and pathogen suspensions were mixed. As controls, monoculture suspensions were prepared by mixing 1 mL of each bacterial suspension with 1 mL of PBS. The initial OD_600_ of the individual probiotic (A_pro_) and pathogen (A_path_) suspensions were recorded. All tubes were vortexed for 10 s and incubated statically at 37 °C for 5 h. After incubation, the final OD_600_ of the mixture’s supernatant (A_mix_) was measured. The co-aggregation percentage was calculated using the formula: Co-aggregation (%) = [(A_pro_ + A_path_)/2 − A_mix_]/[(A_pro_ + A_path_)/2] × 100%. The pathogens used in this study include Salmonella enterica ATCC13076 (S1), Listeria monocytogenes ATCC19115 (S2), Pathogenic Escherichia coli ATCC 25922 (S3), Proteus vulgaris ATCC29905 (S4), Yersinia enterocolitica ATCC23715 (S5), Pseudomonas aeruginosa PAO1 (S6), Shigella dysenteriae ATCC13313 (S7) and Shigella boydii ATCC9207 (S8).
The survival of the strain under simulated gastrointestinal (GI) tract conditions was evaluated using a sequential in vitro digestion model, encompassing exposure to simulated gastric fluid (SGF) followed by simulated intestinal fluid (SIF) [36]. The SGF was prepared by dissolving pepsin (Yuanye Bio-Tech, Shanghai, China, 9001-75-6) at 0.1% (w/v) in sterile 0.85% saline solution, with the pH adjusted to 2.0 and 3.0 using 1 M HCl. The SIF was prepared by dissolving pancreatin (Yuanye Bio-Tech, China, 8049-47-6) at 0.1% (w/v) and bile salts (Yuanye Bio-Tech, China, S30875) at 0.15% (w/v) in sterile 0.85% saline, with the pH adjusted to 8.0 using 1 M NaOH. Both solutions were sterilized by filtration through a 0.22 μm membrane filter before use. A total of 10 mL overnight bacterial culture was centrifuged (4000× g, 10 min, 4 °C) and washed twice with sterile phosphate-buffered saline (PBS, pH7.2). Cells were harvested and resuspended in 1 mL PBS. The prepared cell suspension was inoculated (2%, v/v) into 20 mL of SGF and incubated at 37 °C with gentle agitation. After 3 h of incubation in SGF, an aliquot of 3 μL was taken for enumeration of viable cells by the plate counting method. Subsequently, the remaining culture (15 mL) from the SGF treatment was collected by centrifugation (4000× g, 10 min), and the cell pellet was resuspended in 15 mL of pre-warmed SIF. The suspension was then incubated for an additional 3 h at 37 °C. Samples were collected at 3 h post-incubation to determine the viable cell counts.
To assess the hemolytic activity of the strain, the bacteria was cultivated using Columbia blood agar (HopeBio, China, HB8511) at 37 °C for 2 days. Then, the culture was examined for the presence of α- or β-hemolysis around the colonies.
To test the antibiotic susceptibility of the culture, eight antibiotics were selected: amikacin (30 µg, AK-30), chloramphenicol (30 µg, C-30), ciprofloxacin (2 µg, CIP-5), ampicillin (10 µg, AMP-10), erythromycin (15 µg, E-15), vancomycin (30 µg, VA-30), tetracycline (30 µg, TE-30) and gentamicin (10 µg, CN-10). An aliquot of 50 µL fresh bacterial culture was spread on an MRS agar, followed by the placement of paper disks saturated with antibiotics. The plates were placed at 37 °C for 2 days, and the inhibition zones were measured and recorded.
2.5. Bile Salt Hydrolase (BSH) Activity Assessment In Vitro
The bile salt hydrolase (BSH) activity of strain IOBRA9746 was determined by quantifying the amount of amino acid released from conjugated bile salts, following the ninhydrin-based colorimetric method described by Shehata et al. [37]. Briefly, the strain was cultured in MRS broth at 37 °C for 18 h. The cells were then harvested by centrifugation (15,000× g, 30 min, 4 °C), washed twice with sterile saline (0.85%), and resuspended in the same saline solution to a high density (OD_600_ = 10.0). The cell suspension was mechanically lysed on ice using an ultrasonic processor (Scientz, Ningbo, China) equipped with a Φ2 mm microtip. The sonication program consisted of a total of 3 min of processing time, with cycles of 3 s ‘on’ at 40% amplitude followed by 5 s ‘off’ to prevent overheating. The resulting cell lysate was clarified by centrifugation (15,000× g, 10 min, 4 °C), and the supernatant, designated as the cell-free extract (CFE), was collected for the subsequent enzyme assay. The enzymatic reaction was initiated by mixing 100 μL of the CFE with 100 μL of a 20 mM substrate solution in a microcentrifuge tube. The substrates tested individually were sodium glycocholate (GCA), sodium glycodeoxycholate (GDCA), and sodium glycochenodeoxycholate (GCDCA). The reaction mixture was incubated at 37 °C for 30 min. The reaction was then terminated by adding 200 μL of 15% (w/v) trichloroacetic acid (TCA). Precipitated proteins were removed by centrifugation (15,000× g, 15 min, 4 °C). A control reaction was prepared similarly but with TCA added prior to the incubation to account for any non-enzymatic hydrolysis. The amount of glycine released was quantified using the ninhydrin colorimetric method. Briefly, 300 μL of the supernatant from the previous step was mixed with 1.0 mL of ninhydrin reagent. The mixture was heated in a boiling water bath for 15 min, immediately cooled on ice for 2 min, and then 1.0 mL of 95% ethanol was added. The absorbance of the final solution was measured at 570 nm (OD_570_). A standard curve was generated using known concentrations of glycine to correlate absorbance with amino acid concentration. One unit (U) of BSH activity was defined as the amount of enzyme required to release 1 μmol of amino acid from the conjugated bile salt per minute. The specific activity was expressed as U per milligram of protein in the CFE. BSH activity was measured in three independent experiments using separately cultured bacterial preparations (n = 3 biological replicates). Data are presented as mean ± SD.
2.6. Animal Treatment
All animal procedures were approved by the Animal Ethics Committee of Yunnan University (Permit number: YNU20241036, approved on 8 August 2024). Male C57BL/6J mice (5 weeks of age) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). Animals were housed in the specific pathogen-free (SPF) barrier facility of the Experimental Animal Center of Yunnan University in individually ventilated cages (IVC) under controlled conditions (temperature 25 ± 2 °C, relative humidity 50 ± 5%, 12 h light/12 h dark cycle) with free access to food and water. After 1 week of acclimatization, the general health status of all mice was examined and no abnormalities affecting animal welfare were observed. Mice were then fasted for 24 h, weighed, and subsequently randomly allocated according to body weight into five groups (n = 5 per group; total n = 25): (1) control diet group (CD); (2) high-cholesterol diet with saline (HCD); (3) high-cholesterol diet with simvastatin (SIM); (4) high-cholesterol diet with live strain IOBRA9746 (vA9746); and (5) high-cholesterol diet with ultrasonically lysed strain IOBRA9746 (uA9746). The CD group received a standard chow diet (XT102C), whereas the other four groups were fed a high-cholesterol diet (XT108C). Both diets were obtained from Jiangsu Xietong Pharmaceutical Bioengineering Co., Ltd. (Nanjing, China). Simvastatin was prepared as a suspension adjusted to the mice’s weight with sterile saline. Bacteria were cultivated in MRS liquid medium 12 h prior to use, harvested by centrifugation, washed twice, and resuspended in sterile saline at a concentration of 1 × 10^8^ CFU/mL.
2.7. Sample Collection and Examination
The mice feeding experiment spanned 8 weeks, after which isoflurane was administered to each mouse to induce anesthesia, followed by anatomical examination. The spleen, liver, kidneys, lungs, heart, and colon were carefully excised, and all were weighed individually for each animal. Blood samples were allowed to stand for 30 min at room temperature and then centrifuged at 1000 rpm for 20 min in a refrigerated centrifuge (Eppendorf Centrifuge 5910 R, Eppendorf SE, Enfield, CT, USA). Plasma was collected, aliquoted, and stored at −80 °C until analysis. Plasma total cholesterol (CHOL), high-density lipoprotein cholesterol (HDL), low-density lipoprotein cholesterol (LDL), aspartate aminotransferase (AST), and alanine aminotransferase (ALT) levels were determined using an automated biochemical analyzer (Dimension^®^ EXL™ 200, Siemens Healthcare Diagnostics Inc., Newark, DE, USA). Liver samples were fixed in 4% paraformaldehyde (Biochem, Shanghai, China) at 4 °C overnight. They were then sectioned into 5-micrometer slices using a Leica RM2235 microtome (Leica Biosystems Microtomes, Heidelberg, Germany). Portions of the liver, spleen, kidney, lung, heart and colon were collected immediately after euthanasia and fixed in 4% paraformaldehyde at 4 °C for at least 24 h. Fixed tissues were dehydrated through a graded ethanol series, cleared in xylene, embedded in paraffin, and sectioned at a thickness of 4 µm. For routine histopathological evaluation, paraffin sections of all organs were stained with hematoxylin and eosin (H&E) according to standard protocols. To assess hepatic lipid accumulation, additional liver samples were embedded in optimal cutting temperature (OCT) compound, snap-frozen, and cut into 8 µm cryosections, which were subjected to Oil Red O staining and lightly counterstained with hematoxylin. After mounting, all stained sections were examined and imaged using a light microscope (Olympus, Tokyo, Japan) equipped with a digital camera. For the statistical comparison of serum indicators and organ indices, differences between groups were analyzed by one-way ANOVA followed by Tukey’s post hoc test to correct for multiple comparisons. A p-value < 0.05 was considered statistically significant. All analyses were performed using GraphPad Prism 9.5 software [38].
3. Results
3.1. Isolation and Phylogenetic Analysis of Strain IOBRA9746 from Human Gut Microbiota
Strain IOBRA9746 was isolated from a fecal sample of a healthy human volunteer with no recent history of probiotics or antibiotics, using MRS agar. Whole-genome sequencing revealed that the strain IOBRA9746 harbors six copies of 16S rRNA gene. Pairwise BLAST 2.17.0+ analysis indicated that these copies are nearly identical, with similarities ranging from 99.936% to 100% (Supplementary Table S1). Subsequently, one full-length 16S rRNA gene copy (IOBRA9746_01735; 1557 bp) was selected as the representative sequence for taxonomic analysis. Pairwise BLAST analysis identified strain IOBRA9746 as a member of the genus Enterococcus, showing 99.8% sequence identity to E. durans. However, phylogenetic trees based on 16S rRNA gene sequences consistently clustered IOBRA9746 with E. lactis and E. faecium, rather than with E. durans (Figure 1A; Supplementary Figures S1–S3). To further delineate the phylogenetic affiliation of IOBRA9746, a comparative genomic approach was employed, constructing phylogenomic trees from conserved gene sequences. These analyses were based on a selection of reference strains from the GTDB database, as detailed in Supplementary Table S2. In contrast to the 16S rRNA gene-based phylogeny, IOBRA9746 formed a monophyletic group with E. lactis, with the highest ANI value of 98.0%, exceeding that recorded for E. faecium (94.2%) and E. durans (81.9%) (Figure 1B,C, Supplementary Figure S4). According to the species demarcation thresholds proposed by OGRI [39], which include a 16S rRNA gene sequence similarity threshold of 98.65% and an ANI threshold of 96%, IOBRA9746 exhibited a closer genetic affinity to the E.lactis species. Given the established safety and probiotic potential of E. lactis strains through comprehensive genomic and phenotypic analyses, the close phylogenetic proximity of IOBRA9746 to E. lactis suggests a similar potential for probiotic and safe attributes [11,40].
3.2. Genomic Analysis and Virulence Factor Prediction
To further ascertain the safety of strain IOBRA9746, a comprehensive genomic analysis was conducted. The assembled genome of IOBRA9746 comprises eight contigs, all of which were predicted to be circular (Supplementary Table S3, Supplementary Figure S5). The primary chromosome (IOBRA9746_01) spans 2,676,302 bp with a G+C content of 38.36% and harbors a total of 2631 genes (Figure 2A). Of the identified genes, 1201 genes were annotated with KEGG pathways, predominantly involved in carbohydrate metabolism (Figure 2B). Consistent with this, the majority of genes were found to be enriched in carbohydrate metabolism or catalytic activity within COG and GO systems (Supplementary Figures S6 and S7). Nevertheless, 23 genes implicated in drug and antimicrobial resistance were identified via the KEGG database. All genes in the chromosome were further confirmed for pathogenicity potential using additional databases VFDB, CARD, and ResFinder. Ultimately, 35 genes were identified as potentially pathogenic (Supplementary Table S4). But, genes such as tufA, bopD, sgrA, acm, and clpP, associated with biofilm formation and host cell adherence, were previously identified in non-pathogenic E. lactis species. Other identified genes, including vanY, efrA, AAC(6′)-Ii, were related to antibiotic resistance. Consequently, the antibiotic susceptibility of IOBRA9746 was tested against eight antibiotics, demonstrating sensitivity to all (Figure 2C). Subsequently, the potential for horizontal gene transfer of these pathogenicity-related genes was investigated by examining flanking mobile genetic elements. As shown in Figure 2D, only the tufA was identified within a genomic island, bordered by an IS gene. The absence of cytolysin genes was further corroborated by phenotypic analysis. After 48 h on Columbia Blood Agar, IOBRA9746 did not exhibit haemolysis activity, in contrast to the Escherichia isolates IOBRA9596, which produced strong β-haemolysis (Figure 2C).
For the remaining assembled contigs presumed to be plasmids, 351 genes were annotated across a total length of 305,169 bp, with the longest contig, IOBRA9746_02, encompassing 221 genes (Supplementary Table S3). While these contigs are predicted to be circular, IOBRA9746_02 exclusively contains a T4CP (Type IV Coupling Protein) gene. In contrast, contig IOBRA9746_03, containing 56 genes, features both an oriT site and a relaxase gene (Figure 2A). Thus, IOBRA9746_02 may represent an incomplete plasmid or an artifact of misassembly. Despite the potential presence of a plasmid in IOBRA9746, no pathogenic genes were identified within these contigs (Figure 2A, Supplementary Figure S8).
3.3. Probiotic Characteristics and Bile Salt Hydrolase (BSH) Assessments In Vitro
Next, the probiotic potential of the IOBRA9746 was tested through a series of in vitro assessments. The cell surface hydrophobicity and auto-aggregation capacity of the strain, which indicate its potential for mucosal adhesion and biofilm formation, were quantitatively assessed over a temporal gradient. Hydrophobicity analysis revealed a time-dependent enhancement of microbial adhesion to nonpolar solvents, with values increasing from 5.78 ± 1.08% at 1 h to 14.32 ± 8.41% (mean ± SD, n = 3) by 5 h incubation (Figure 3A). Measurements of auto-aggregation also demonstrated progressive microbial self-association, exhibiting 7.35 ± 0.84%, 26.76 ± 3.03%, and 33.062 ± 6.67% (mean ± SD, n = 3) aggregation efficiencies at 1 h, 3 h, and 5 h respectively (Figure 3A). The co-aggregation efficiency between strain IOBRA9746 and common pathogenic bacteria was quantitatively evaluated to assess their interspecies interaction potential, which may contribute to competitive exclusion of pathogens. As shown in Figure 3B, strain IOBRA9746 exhibited the highest co-aggregation efficiency with Yersinia enterocolitica. The survival rates of strain IBORA9746 were measured under varying temperature conditions and pH levels. The optimal growth conditions were 37 °C and pH 7.0, while the strain retained viability under extreme conditions of 45 °C and pH 4.1 (Figure 3C). To further assess gastrointestinal tolerance, IOBRA9746 was sequentially exposed to simulated gastric juice and intestinal fluid. As assessed by serial-dilution spot plating, IOBRA9746 showed no apparent reduction in viable counts following SGF and subsequent SIF exposure (Figure 3D), suggesting strong tolerance to simulated gastrointestinal conditions. Subsequently, the BSH activity of strain IOBRA9746 was quantified via an in vitro enzymatic assay. This analysis confirmed extracellular BSH functionality and specifically determined catalytic efficiency against three glycine-conjugated bile salts: GCA, GDCA, and GCDCA (Figure 3E). The specific activities for these substrates were measured as 51.15 ± 2.997, 43.02 ± 3.967, and 17.97 ± 2.391 μmol·min^−1^·mg^−1^ protein (mean ± SD, n = 3), respectively.
3.4. Systemic Metabolic Modulation by Strain IOBRA9746 In Vivo
To evaluate the in vivo cholesterol-lowering efficacy and safety profile of strain IOBRA9746, an 8-week animal intervention study was conducted (Figure 4A). C57BL/6J mice were randomly divided into five groups: normal chow diet (CD), high-cholesterol diet (HCD), HCD treated with simvastatin (SIM), HCD supplemented with viable strain IOBRA9746 (v9746) and HCD supplemented with ultrasonically lysed strain IOBRA9746 (u9746). Body weight was monitored weekly throughout the experimental period, and no significant differences were observed among these three groups (Figure 4B), suggesting that the bacterial administration did not adversely affect normal growth or cause apparent toxicity. Serum lipid profiles were analyzed at the endpoint. Compared with the HCD group, total cholesterol (CHOL) was numerically lower in both the SIM and v9746 groups; however, no statistically significant differences were observed (p > 0.05). This may be due to inter-individual variability, the preventive design with concurrent HCD feeding, and the moderate cholesterol-loading model used. Nevertheless, high-density lipoprotein cholesterol (HDL-C) levels remained stable across all groups (Figure 4C), indicating no adverse effect on beneficial lipid fractions. Despite the modest changes in serum parameters, histopathological examination of liver sections revealed marked improvements. H&E staining demonstrated that the HCD group developed prominent hepatic steatosis characterized by extensive lipid droplet accumulation (Figure 4D). In contrast, SIM, u9746 and v9746 groups showed a substantial reduction in intrahepatic lipid deposition, with fewer and smaller lipid vacuoles, approaching the morphology observed in the CD group, and Oil Red O staining further confirmed this protective effect (Figure 4D). Corroborating the histopathological findings, serum biomarkers of liver function showed a pattern consistent with diet-induced hepatic injury. While AST levels were only marginally elevated, ALT was significantly higher in the HCD group, indicating hepatocellular damage (Figure 4C). The attenuation of serum ALT levels in mice supplemented with strain IOBRA9746, coupled with the marked reduction in hepatic lipid accumulation, strongly suggests that the strain’s primary mechanism of action is the modulation of hepatic lipid metabolism to protect against steatosis.
3.5. The Effect of Strain IOBRA9746 on Histological Changes in High-Cholesterol Fed Mice
To comprehensively assess the biosafety of prolonged bacterial administration, major organs including heart, lung, spleen, and ileum were examined histologically. No pathological abnormalities, inflammatory infiltration, or structural damage were detected in any of these tissues. Cardiac tissue exhibited well-organized myocardial bundles with distinct cross-striations and branching patterns, interspersed with minimal connective tissue and devoid of degeneration, necrosis, or inflammatory infiltrates. Splenic architecture maintained clear demarcation between red and white pulp, displaying defined lymphoid follicles and intact periarteriolar lymphoid sheaths. Pulmonary sections demonstrated characteristic honeycomb alveolar structures with thin, continuous walls composed of a single-layered alveolar epithelium and abundant capillaries; alveolar lumina showed no fluid accumulation or foreign material, with no interstitial thickening. Renal parenchyma presented well-delineated glomerular and tubular structures, featuring morphologically normal glomeruli and intact renal tubular epithelium devoid of luminal casts or debris. Colonic mucosa exhibited distinct histological layers with an uninterrupted epithelial surface, abundant regularly arranged glands in the lamina propria opening onto the mucosa, and no evidence of epithelial damage or inflammatory responses. Collectively, all organs displayed normative histology without pathological alterations (Figure 5).
Experimental design, body weight, serum biochemical parameters and liver histology in mice fed a high-cholesterol diet and treated with probiotic strains. (A) Schematic overview of the in vivo experimental design and group allocation. Five-week-old male C57BL/6J mice were randomly assigned to five groups. All groups were fed their respective diets and received the indicated treatments for 8 weeks. (B) Time course of body weight changes during the 8-week intervention in each experimental group. (C) Serum biochemical parameters measured at the end of the experiment, including total cholesterol (CHOL), low-density lipoprotein cholesterol (LDL-C), high-density lipoprotein cholesterol (HDL-C), aspartate aminotransferase (AST) and alanine aminotransferase (ALT). Data are presented as mean ± SD (n = 5 per group). Statistical comparisons were performed using one-way ANOVA followed by Tukey’s post hoc test. * p < 0.05, ** p < 0.01. (D) Representative liver sections from each group stained with hematoxylin and eosin (H&E) and Oil Red O.
To evaluate potential systemic effects, we measured organ indices and colon length. No significant differences were observed in pulmonary index, renal index, or colon length among groups. However, variations were noted in cardiac and splenic indices: Group v9746 exhibited a significantly higher cardiac index compared to Groups CD and HCD, while Group HCD showed a marked decrease in splenic index relative to Group CD, with significant divergence from Group v9746 (Figure 5). Crucially, these organ index variations were not accompanied by histopathological abnormalities. As described above, comprehensive microscopic examination of cardiac and splenic tissues revealed preserved tissue architecture without evidence of inflammation, cellular hypertrophy, or necrosis. This disparity suggests that observed organ weight variations likely reflect subtle physiological adaptations, such as transient edema or hemodynamic fluctuations, rather than overt pathological damage, although the biological relevance of these findings needs further investigation.
Effects of probiotics on organ histology and organ indices in mice fed a high-cholesterol diet. Representative hematoxylin and eosin (H&E)-stained sections and corresponding organ indices from mice in the control diet (CD), high-cholesterol diet (HCD) and HCD + live 9746 (V9746) groups after 8 weeks of intervention. Shown are histological images of the heart, spleen, lung, kidney and colon, together with the calculated cardiac index, splenic index, pulmonary index, renal index and colon length. Data are presented as mean ± SD (n = 5 per group). Statistical comparisons were performed using one-way ANOVA followed by Tukey’s post hoc test. * p < 0.05, ** p < 0.01, *** p < 0.001.
4. Discussion
The human gut harbors a substantial microbial reservoir, serving as a valuable source for probiotic candidate screening [41]. Previous studies have established a significant consensus on the safety of human gut-derived E. lactis through comprehensive genomic analyses and in vitro assessments [11]. In this study, a comprehensive safety assessment of the strain was conducted through rigorous genomic characterization coupled with in vitro and in vivo experimental validations. We first performed a phylogenetic analysis to resolve the strain’s taxonomic positioning within the Enterococcus phylogeny, as the genus Enterococcus exhibits considerable diversity across disparate ecological niches, which can lead to potential misclassification based solely on 16S rRNA gene sequencing. For instance, 16S rRNA gene analysis indicated high similarity (98.8–99.4%) between E. lactis BT159^T^ and type strains E. hirae ATCC 8043^T^, E. durans CECT 411^T^, and E. faecium ATCC 19434^T^ [10]. This exemplifies the potential unreliability of single-gene approaches for accurate taxonomic delineation within Enterococci. Moreover, recent genomic studies have demonstrated that strains traditionally classified as E. faecium clade B (community-associated isolates) are conspecific with E. lactis. Core genome phylogeny, ANI, and digital DNA–DNA hybridization analyses also support their taxonomic reclassification as E. lactis [42]. Thus, precise taxonomic delineation of this group is essential for the identification of safe probiotic candidates. In this study, we sequenced the genomic DNA of strain IOBRA9746 using both short-read and long-read platforms, generating a near-complete genome assembly. Initial phylogenetic analysis of the 16S rRNA gene revealed ambiguous taxonomic placement as well. However, the reconstructed genomic tree positioned IOBRA9746 within a monophyletic clade with E. lactis. This relationship was further corroborated by ANI analysis. Based on the analysis above, the taxonomic status of IOBRA9746 has been unequivocally elucidated, confirming its classification within the E. lactis group and demonstrating its safety potential.
In addition to phylogenetic validation, we further characterized the genetic repertoire of this strain. Genomic safety assessment revealed the presence of virulence-associated genes including vanY, AAC(6′)-Ii, and msrC in strain IOBRA9746, but most of them were previously documented in E. lactis isolates (see Table S4, see isolates no. in Table S5). Crucially, all identified genes resided outside mobile genetic elements, with the sole exception of tufA—a gene encoding the elongation factor Tu implicated in bacterial adhesion capabilities. Subsequently, we experimentally validated the safety of the strain. Hemolytic activity and antibiotic susceptibility analyses further corroborated its safety profile. Furthermore, long-term administration of the strain was conducted in mice fed a specific dietary regimen. Histopathological examination of major organs was performed to assess its potential impact. The results revealed no significant adverse effects on organ integrity or morphology following prolonged oral gavage.
We next centered on the BSH activity of the strain IOBRA9746. BSH mediates the hydrolysis of glycine/taurine-conjugated bile salts into deconjugated bile acids. One of the hydrolysis products, taurine, exerts direct and multifaceted effects on host metabolism, immune response, and neurological function [43]. For instance, taurine modulates the NLRP6 inflammasome to enhance IL-18 secretion, thereby ameliorating intestinal inflammation [44,45]. It also promotes the expansion of anti-inflammatory microbiota while inhibiting the colonization of pathogenic bacteria [45]. Furthermore, taurine acts as a mitochondrial matrix buffer, facilitating fatty acid oxidation and enhancing energy metabolism. However, taurine may also elicit detrimental effects. It can stimulate the production of hydrogen sulfide (H_2_S) and enrich H_2_S-producing bacteria such as Bilophila wadsworthia [46,47]. Excessive H_2_S induces DNA damage in intestinal epithelial cells [48,49,50] and may have been associated with ulcerative colitis [51]. In this study, we evaluated the BSH activity of the strain IOBRA9746 in vitro and observed distinct substrate specificity, with preferential hydrolysis of glycine-conjugated bile salts over taurine-conjugated counterparts. This specificity suggests that IOBRA9746 is unlikely to contribute to excessive H_2_S generation in vivo, thereby reinforcing its safety profile and supporting its potential for further development as a probiotic candidate. While prior genomic analyses indicated the presence of bsh genes in E. lactis isolates [12,52], quantitative assessment of its enzymatic activity and in vivo validation remain less explored. BSH facilitates cholesterol excretion and modulates lipid metabolism, thereby reducing cholesterol levels [53]. To address the knowledge gap regarding E. lactis BSH functionality, particularly in contrast to well-studied Enterococcus strains like E. faecium WEFA23 [54], we conducted an 8-week oral gavage experiment in mice fed a high-cholesterol diet (HCD). In the HCD-fed mouse model, IOBRA9746 administration attenuated hepatic lipid droplet accumulation, whereas no statistically significant differences were detected in serum lipid parameters.
Although direct mechanistic datasets are not yet available, a plausible explanation for the liver-targeted benefit is related to BSH-driven modulation of bile acid metabolism along the gut–liver axis. BSH-mediated deconjugation can reshape the intestinal bile acid pool and enterohepatic circulation, potentially influencing bile-acid-dependent signaling pathways (e.g., FXR and TGR5) that regulate hepatic lipid homeostasis. Therefore, we speculate that IOBRA9746 may reduce hepatic lipid deposition primarily through bile acid remodeling and downstream signaling rather than by inducing a robust systemic cholesterol-lowering effect. This proposed mode of action requires direct validation in future studies, including serum/fecal bile acid profiling, hepatic gene-expression analyses (e.g., CYP7A1, FXR and downstream targets), and microbiota functional characterization.
Several limitations should be acknowledged. First, the present study employed a single bacterial dosage (1 × 10^8^ CFU/mL) without evaluating a broader concentration gradient. This dosage was selected to align with the established therapeutic window (10^8^–10^9^ CFU/mL) commonly reported in probiotic interventions [55,56,57]. For instance, recent genomic and in vivo profiling of Pediococcus pentosaceus SPARC2 demonstrated that dosages as high as 10^9^ CFU are safe and capable of significantly enhancing health parameters and modulating gut microbiota [58]. While our selected dose successfully elicited hepatic lipid reduction, we acknowledge that a dose–response study is necessary to determine the optimal intake level and to explore whether higher doses could yield superior therapeutic outcomes. Second, the experimental design aimed to evaluate the preventive rather than therapeutic potential of the strain. Therefore, mice were subjected to bacterial intervention concurrently with HCD feeding. Notably, some mice in the HCD group did not develop pronounced hypercholesterolemia, further complicating the interpretation of serum-based outcomes. However, significant hepatic lipid accumulation was observed in the vehicle group, confirming that the model was sufficient to evaluate the strain’s efficacy in preventing diet-induced hepatic steatosis. Third, the sample size was limited to five mice per group in adherence to ethical principles of reduction. We acknowledge that without a prior power calculation, this sample size might be insufficient to overcome the inter-individual variability observed in serum lipid profiles, which explains why significant differences were detected in histopathological outcomes but not in serum parameters.
Finally, the strain showed modest in vitro surface-associated traits (hydrophobicity and aggregation-related phenotypes) compared with certain benchmark probiotics reported in the literature [59,60,61,62,63,64]. As these assays are simplified proxies for adhesion/competitive exclusion, they may not directly predict in vivo metabolic efficacy. Notably, the biological activity observed with the ultrasonically lysed preparation suggests that non-viable components or released intracellular factors (potentially including BSH and cell wall constituents) may contribute to the observed effects, supporting a potential postbiotic application. Future work should integrate dose gradients, established hypercholesterolemic models, and colonization/persistence analyses to clarify the relative contributions of adhesion-dependent versus metabolite/enzyme-mediated mechanisms.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Afshin A. Sur P.J. Fay K.A. Cornaby L. Ferrara G. Salama J.S. Mullany E.C. Abate K.H. Abbafati C. Abebe Z. Health Effects of Dietary Risks in 195 Countries, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017 Lancet 201939319581972 Correction in Lancet 2021, 397, 2466. https://doi.org/10.1016/S 0140-6736(21)01342-810.1016/S 0140-6736(19)30041-830954305 PMC 6899507 · doi ↗ · pubmed ↗
- 2Postic C. Girard J. Contribution of de Novo Fatty Acid Synthesis to Hepatic Steatosis and Insulin Resistance: Lessons from Genetically Engineered Mice J. Clin. Investig.200811882983810.1172/JCI 3427518317565 PMC 2254980 · doi ↗ · pubmed ↗
- 3Tripathi A. Debelius J. Brenner D.A. Karin M. Loomba R. Schnabl B. Knight R. The Gut–Liver Axis and the Intersection with the Microbiome Nat. Rev. Gastroenterol. Hepatol.201815397411 Correction in Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 785. https://doi.org/10.1038/s 41575-018-0031-810.1038/s 41575-018-0011-z 29748586 PMC 6319369 · doi ↗ · pubmed ↗
- 4Turnbaugh P.J. Ley R.E. Mahowald M.A. Magrini V. Mardis E.R. Gordon J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest Nature 20064441027103110.1038/nature 0541417183312 · doi ↗ · pubmed ↗
- 5Yoo J. Kim S. Probiotics and Prebiotics: Present Status and Future Perspectives on Metabolic Disorders Nutrients 2016817310.3390/nu 803017326999199 PMC 4808900 · doi ↗ · pubmed ↗
- 6Jones M.L. Martoni C.J. Parent M. Prakash S. Cholesterol-Lowering Efficacy of a Microencapsulated Bile Salt Hydrolase-Active Lactobacillus reuteri NCIMB 30242 Yoghurt Formulation in Hypercholesterolaemic Adults Br. J. Nutr.20121071505151310.1017/S 000711451100470322067612 · doi ↗ · pubmed ↗
- 7Begley M. Gahan C.G.M. Hill C. The Interaction between Bacteria and Bile FEMS Microbiol. Rev.20052962565110.1016/j.femsre.2004.09.00316102595 · doi ↗ · pubmed ↗
- 8Franz C.M.A.P. Huch M. Abriouel H. Holzapfel W. Gálvez A. Enterococci as Probiotics and Their Implications in Food Safety Int. J. Food Microbiol.201115112514010.1016/j.ijfoodmicro.2011.08.01421962867 · doi ↗ · pubmed ↗