A One Health Perspective on Proteus mirabilis: The Interaction of Virulence and Antimicrobial Resistance Across Human and Animal Reservoirs
Ibtisam Faeq Hasona, Amal Awad, Gamal Younis, Wafaa Farouk Mohamed

TL;DR
Proteus mirabilis spreads between humans, animals, and the environment, combining strong infection-causing traits with antibiotic resistance, requiring a unified health approach to control.
Contribution
The paper highlights the shared transmission of virulence and AMR in P. mirabilis across human and animal reservoirs, emphasizing the need for One Health strategies.
Findings
Identical multidrug-resistant P. mirabilis clones are found in humans, livestock, food, and the environment.
Virulence traits like urease and biofilm formation aid in AMR dissemination through mobile genetic elements.
Integrated surveillance and antimicrobial stewardship are critical to managing P. mirabilis spread.
Abstract
Proteus mirabilis (P. mirabilis), a common commensal and opportunistic pathogen, circulates freely across interconnected human, animal, and environmental reservoirs, embodying the One Health concept. Its key virulence factors—urease activity, swarming motility, and biofilm formation—drive severe urinary tract infections, particularly catheter-associated ones. These virulence traits concurrently facilitate the acquisition and dissemination of antimicrobial resistance (AMR) via mobile genetic elements, leading to extensively drug-resistant clones. Epidemiological and genomic evidence confirms that identical multidrug-resistant clones and resistance mechanisms (ESBLs, carbapenemases) are shared among human clinical isolates, livestock, food products, and environmental samples. This demonstrates continuous, multi-directional transmission through interconnected zoonotic, foodborne, and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
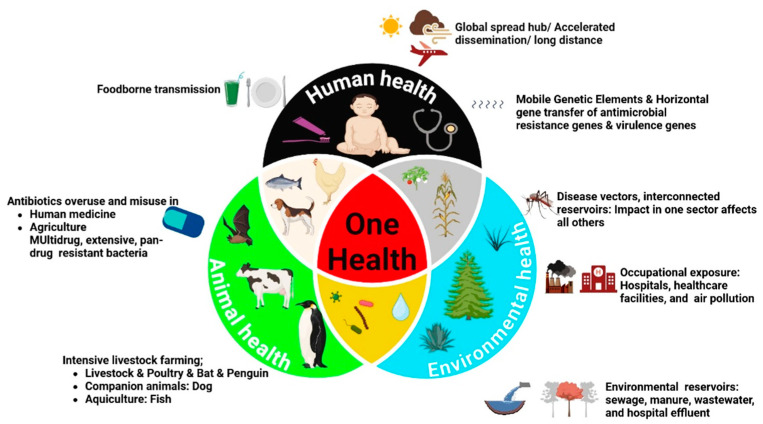 Figure 1
Figure 1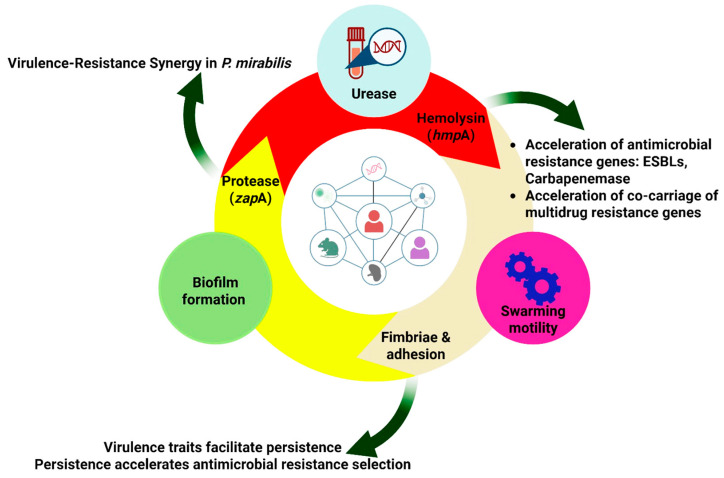 Figure 2
Figure 2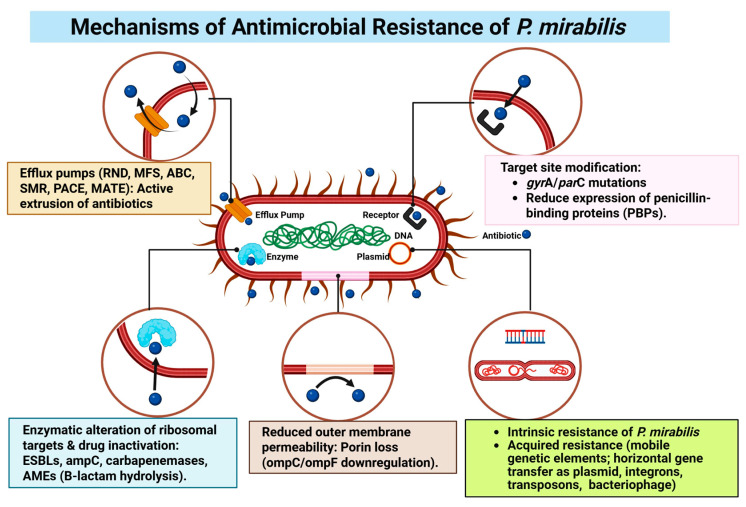 Figure 3
Figure 3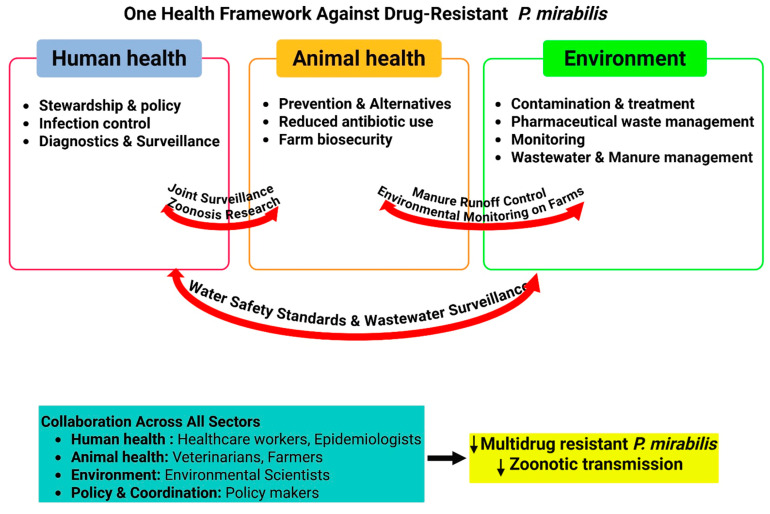 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Escherichia coli research studies · Bacterial biofilms and quorum sensing
1. Introduction
Globally, the incidence of zoonoses is estimated worldwide to be about a billion cases per year [1], causing enormous pressure, especially in low- and middle-income countries (LMICs). One of the decisive factors here is foodborne disease. According to the World Health Organization (WHO), annually, foodborne illnesses amount to 600 million cases with 420,000 deaths [2,3]. This is due to the production of pathogenic bacteria of more than 50%, amounting to over 300 million cases of human infections every year, directly attributed to livestock production [2]. The ramifications extend beyond public health to substantial and global economic losses. This burden is profound across all income levels but disproportionately affects LMICs. The national economic toll is significant, with total annual costs estimated at 391 million in Burkina Faso [4,5]. Collectively, analyses indicate that the total annual economic cost of foodborne diseases across all LMICs reaches approximately 15 billion in direct costs and up to $95.2 billion in indirect costs annually [6,7].
This economic challenge is not confined to LMICs; even in high-income countries with advanced regulatory systems, the toll remains substantial, as illustrated by recent U.S. estimates of $17.6 billion annually for 15 key pathogens [8]. A forward-looking perspective must consider that these costs are projected to rise due to population growth, urbanization, and climate change, factors that may alter pathogen distribution and intensify pressures on global food systems. Therefore, robust estimation of this economic impact is foundational to informed policy-making, evidence-based resource allocation, and the development of effective intervention strategies that address both immediate health needs and long-term economic sustainability [9]. Future research should prioritize generating more localized and comprehensive cost estimates to guide targeted and cost-effective interventions worldwide. Of particular mention here among the opportunistic bacteria causing foodborne illnesses is the Proteus spp., a common bacteria observed in different body parts of humans and animals, like the gut, skin, and mouth. It is also a common bacteria observed in waste matter, soil, and plants [10]. Proteus mirabilis (P. mirabilis) exemplifies a prototype One Health challenge, owing to its capacity for cross-sectoral transmission and exchange of antimicrobial resistance across human, animal, and environmental reservoirs. P. mirabilis, a well-known Gram-negative (GN), rod-shape, facultative anaerobic bacterium, belongs to the genus Proteus within the family Morganellaceae. Traditionally classified in the Enterobacteriaceae family, the taxonomy of P. mirabilis was reassessed with advancements in genomic analysis, leading to its reclassification under Morganellaceae by Adeolu et al. in 2016 based on phylogenetic studies [11]. This organism is a facultative anaerobe, and it has cells that have a dimension of about 0.5 to 1.0 µm and a length of 1.0 to 3.0 µm [12]. The organism is a flexible species that has an optimal growth rate at a temperature of 34–37 °C, and therefore, it thrives very well in the human body because it is a perfect environment for the organism to survive [13]. The organism has a very distinct characteristic of producing a series of concentric rings during swarming motions on solid surfaces, an action that is controlled by a series of at least ten genetic factors for adhesion and peritrichous flagella responsible for the organism’s ability to swim and swarm effectively [14,15]. This multicellular system, whose mode of action involves the production of long swarmer cells by the conversion of the organism’s vegetative cells to elongated cells that help the organism to colonize and spread quickly through the human body, is an important virulence factor [12,15]. This system is very closely associated with the invasion of cells and the production of virulence-associated factors [16]. Furthermore, this unique mode of motion, as well as the ability of the organism to elongate and secrete polysaccharides upon coming into contact with a surface, increases the organism [17]. Given its wide distribution, significant pathogenicity, and escalating role in antibiotic-resistant infections across human and veterinary medicine, a deeper understanding of P. mirabilis is urgently needed. Through a One Health lens, this review examines virulence in human and animal hosts, analyzes AMR dynamics across interconnected reservoirs, and evaluates collaborative strategies for containment. By consolidating data on its key characteristics, pathogenic factors, and evolving resistance profile, this work provides a comprehensive analysis framed within the imperative of a unified One Health approach. Figure 1 below illustrates the interwoven process of spreading resistance to P. mirabilis among human, animal, as well as environmental segments. The important role of joint activities regarding the risk to health from this MDR pathogen, in both areas of health, is highlighted in this work.
2. Clinical Epidemiology and Pathogenic Profile of P. mirabilis in Human Infections
P. mirabilis has the highest isolation rate and geographical range among its kind and has been identified as a normal flora of the intestinal tract of both human beings and animals [12,13]. It has a remarkably versatile range, inhabiting soil, water, and sewage, where the breakdown of organic matter contributes to its metabolic activity [12,18].
The major clinical relevance is due to its existence as one of the leading opportunistic pathogens that cause urinary tract infections (UTIs). It is a leading cause of both community-acquired and healthcare-associated UTIs and is typically recognized as the second most prevalent Enterobacterales species isolated after Escherichia coli (E. coli) [19], account for 1–10% of all human UTI cases [20]. In complex circumstances, infection rates rise dramatically. Following urinary catheterization, the incidence of catheter-associated UTIs (CAUTIs) caused by P. mirabilis can increase to between 20% and 45% or more [12,17,21,22]. Although CAUTIs are typically polymicrobial, P. mirabilis is one of the most commonly detected pathogens [23] and constitutes the predominant causative agent (10–44%) in double-J stent-associated urinary tract Infections (DJUTIs) [24].
These infections are more prevalent in patients with long-term catheter usage, with the highest incidence in older patients and women [12,18,20]. P. mirabilis causes 1–2% of UTI infections in healthy adult women, but accounts for 5% of hospital-acquired UTI cases in hospitalized females [12]. CAUTIs that arise after UTIs have a substantial mortality rate [25]. Cystitis, bacteriuria, acute pyelonephritis, catheter blockage, and fever are some of the symptoms [20]. The underlying reasons for these demographic disparities are multifactorial. The heightened incidence among elderly patients is primarily driven by age-associated factors such as increased comorbidities (e.g., type 2 diabetes, neurological disorders), immunosenescence (the natural decline of immune function with age), and, most critically, a greater prevalence of instrumentation like long-term urinary catheterization, which provides a direct portal for infection and is a major risk factor for P. mirabilis CAUTI [17,18,26,27]. Conversely, the higher prevalence in women, especially during reproductive years (ages 20–50), is largely attributed to anatomical differences—namely, a shorter urethra that facilitates bacterial ascension into the bladder—and possibly hormonal influences [28].
P. mirabilis’ strong urease enzyme is a major contributor to UTI complications. This enzyme hydrolyzes urea into ammonia and carbon dioxide, increasing urine pH and precipitating struvite or apatite crystals [12]. These crystals consolidate into bladder stones, which operate as long-term bacterial reservoirs, making infections difficult to treat and likely to recur. This procedure can also result in catheter encrustation and blockage, which may lead to hydronephrosis and kidney injury [18,29]. Urolithiasis in the bladder and kidneys can cause irreversible kidney damage and frequently requires surgical intervention [18,30].
When P. mirabilis invades the host body, it releases endotoxins that enter the bloodstream, triggering the host’s inflammatory immune response and resulting in conditions such as sepsis or systemic inflammatory response syndrome (SIRS), which have a mortality rate of approximately 20–50% [17]. Infections can lead to more dangerous illnesses including bacteremia and sepsis, either through direct tissue damage or bacterial migration on catheter surfaces [12,17]. P. mirabilis is responsible for 5–18% of all Gram-negative bacteremia cases [12,31]. Bacteremia and sepsis induced by this bacterium are associated with a significant death rate, reaching up to 50% in older people, which is relatively greater than mortality from other infectious sources [18,25,30,32,33]. The progression to severe systemic infections such as septicemia typically occurs in vulnerable patient populations with compromised host defenses. Key predisposing conditions include advanced age, diabetes mellitus, malignancy, and other immunocompromised states, as well as exposure to healthcare settings and invasive devices such as urinary catheters [34,35].
P. mirabilis causes a variety of infections outside of the urinary tract, especially in immunocompromised people. Approximately 90% of clinical isolates arise from UTIs and CAUTIs, alongside the remaining 10% related to infections of the respiratory tract, eye, ear, nose, skin, burns, wounds, meningoencephalitis, and osteomyelitis [32]. Its swarming mobility promotes tissue colonization and spreads [12]. It has also been linked to diarrhea, infective endocarditis, rheumatoid arthritis, and hospital-acquired epidemics [14,22]. Additionally, P. mirabilis has been implicated as a cause of food poisoning incidents, accounting for 3.61% of reported cases in Datong, China between 2016 and 2017, with symptoms such as abdominal pain, diarrhea, nausea, and dizziness [36]. In a 2018 incident in Beijing, contamination of braised meatballs with P. mirabilis led to illness among customers, with the bacterium detected on the hands of the chef and waitstaff [37]. Proliferation is significantly easier in immunocompromised people or those getting antibiotic therapy [18,38]. According to one study, P. mirabilis was isolated from 21.6% of cutaneous abscesses, making it the second most prevalent isolate after methicillin-sensitive Staphylococcus aureus [39]. Potential linkages to Crohn’s disease (CD) and intestinal inflammation aggravated by oral inflammation or proton pump inhibitor use have also been reported [40]. Subsequent studies have strengthened this link; comparison of feces and inflamed colon samples from CD patients and healthy individuals revealed a significant increase in the abundance of P. mirabilis in CD patients. Experimental results showed signs of colon shortening, and liver and spleen enlargement, indicating that P. mirabilis plays a critical role in inducing CD inflammation [39]. Furthermore, it has been suggested that oral inflammation could exacerbate intestinal inflammation, and that the use of proton pump inhibitors may promote the proliferation of P. mirabilis and other microbes in the intestines, thus triggering intestinal inflammation [41]. Regarding diarrhea, another study isolated P. mirabilis from 49 out of 486 pediatric diarrheal samples, yielding a detection rate of 10.1% [42].
3. The Pathogenic Spectrum and Host Range of P. mirabilis in Animals
P. mirabilis has been detected in a variety of hosts, including pets, ruminants, and poultry, demonstrating its remarkable versatility in the field of veterinary medicine. Its host range extends far beyond domesticated animals, encompassing a wide array of wildlife, which underscores its broad ecological niche and zoonotic potential. It has been isolated from wildlife in undisturbed habitats, such as Egyptian vulture chicks in the Canary Islands, migratory bird feces in China, and a fruit bat in Indonesia [43]. In wild mammals, it has been found in wild boars in Tunisia and in gorillas, mandrills, and African buffaloes in a national park in Gabon [44]. Notably, its presence in raptors in Spain and in a juvenile sea lion in Uruguay indicates its reach across diverse species and ecosystems [45]. Furthermore, demonstrating its threat to even rare and protected species, a multidrug-resistant strain of P. mirabilis (PM2022) was identified as the cause of fatal lobar pneumonia and hepatic necrosis in a critically endangered Malayan pangolin. Genomic analysis of this strain revealed a concerning array of antibiotic resistance genes, including those encoding for extended-spectrum beta-lactamase (CTX-M-65), and confirmed the presence of key virulence factors [46]. The detection of P. mirabilis in ticks collected from wild mammals and cattle suggests ectoparasites may play a role in its transmission [47]. In controlled environments such as zoos and farms, it has been identified in species including giant pandas and red pandas in China, and farmed foxes, raccoons, and minks [48]. Notably, in zoo settings, Proteus spp. can cause severe systemic infections; a fatal case of septicemia in a Humboldt penguin was attributed to a concomitant infection by P. mirabilis, P. penneri, P. vulgaris, and P. cibarius, highlighting the potential for severe, multi-species Proteus infections in captive wildlife [49]. It is also present in research animal models like diarrheal rhesus macaques, ferrets [50], and tree shrews [51].
Farm animals constitute a significant reservoir for P. mirabilis, with implications for animal health and food safety. Studies have reported its isolation from pigs in various regions [52,53], and it has been found in boar semen, where it negatively impacts sperm quality [54]. Research indicates a notable presence of P. mirabilis in samples from diseased pigs, highlighting its potential pathogenic role [55]. The bacterium is also commonly isolated from healthy and diseased poultry, cattle, and sheep across different countries, with reports from chicken flocks, duck populations, and livestock manure underscoring its widespread occurrence in agricultural settings [52,56,57]. Its association with clinical disease, including diarrhea in poultry and cattle, further points to its importance as an agent affecting livestock health and productivity [55,56].
Companion animals, due to their close contact with humans, represent another critical reservoir and a potential source of zoonotic transmission. P. mirabilis is frequently detected in pets. It has been co-isolated from humans and dogs in shared households [58] and is commonly found in fecal samples from both household and stray dogs [59]. Research indicates that the frequency of intestinal colonization with P. mirabilis is significantly higher in dogs compared to their human cohabitants. Importantly, molecular studies have confirmed the sharing of genetically related P. mirabilis strains between humans and dogs living in the same household, with some of these shared fecal strains also showing genetic relatedness to clinical uropathogenic strains. This provides direct evidence of cross-species transmission and underscores the dog’s role as a potential reservoir for human infections [58]. A focused study on UTIs found a distinct prevalence pattern: while P. mirabilis was not isolated from dogs with UTIs in that cohort, it was identified in 4% of cats with UTIs, and at a much higher rate (26%) in human UTI cases, highlighting species-specific differences in urinary tract colonization [60]. Exotic pets, such as turtles, have also been shown to carry the bacterium [61]. A primary health concern is UTI. P. mirabilis is a notable cause of UTIs in dogs and, to a lesser extent, in cats, as reported in multiple studies across different continents [62,63,64]. In veterinary medicine, this association is particularly significant as P. mirabilis has been specifically linked to the formation of recurrent urinary stones in dogs with urinary system disorders [65]. Beyond UTIs, the bacterium is also associated with gastrointestinal issues like diarrhea in dogs [66] and has been identified in cases of co-infection with major viral pathogens [67].
In the aquaculture sector, P. mirabilis is emerging as a significant primary pathogen across key farmed fish species, characterized by severe disease. It causes high mortality (up to 68.8%) and systemic infections in African catfish (Clarias gariepinus) with clinical signs like skin hemorrhages and ulceration, leading to hepatic/renal damage and anemia [68,69]. Its prevalence in Nigerian catfish farms is significant (13–31%) [68]. In Nile tilapia (Oreochromis niloticus), infection induces renal–hepatic dysfunction, oxidative stress, and histopathological damage [70], though a separate tilapia strain showed probiotic potential against Vibrio spp. [71]. In Indian major carp (Labeo catla), it is a confirmed lethal pathogen [72]. This, coupled with its detection in freshwater fish sold for consumption [73], underscores a direct food safety threat. The ecological role appears complex, as co-infection with Aeromonas hydrophila (A. hydrophila) in catfish showed antagonistic interaction, resulting in lower mortality than a single A. hydrophila infection [74].
Animal-derived foods have become a major focus of public health concern due to their contamination with P. mirabilis. The prevalence of the bacterium in meat and other products varies significantly across different countries and regions, reflecting differences in hygiene practices and environmental conditions. Significant contamination has been reported in various meat types: in India [52], Iran [75], Brazil [76], Egypt [77,78,79], Belgium [80], and China [81,82,83,84]. Chicken meat often shows high contamination rates in these studies, followed by pork and beef. These findings indicate that poor hygiene at poultry and meat stalls may result in substantial contamination and cross-contamination, particularly affecting poultry products. A recent study in China also reported contamination of P. mirabilis in retail meat and aquatic products [82]. Insects such as flies associated with animal-derived food also appear to serve as potential vectors of transmission [85]. Furthermore, the presence of P. mirabilis in wildlife such as wild boars and African buffalo [86], and in freshwater fish sold in markets [73], which are sources of game meat and aquatic products, suggests a potential route of zoonotic transmission from wildlife to humans, although this possibility requires further investigation.
The pathogenicity of P. mirabilis in animals is well-documented across multiple organ systems. As a gastrointestinal pathogen, it has been responsible for severe outbreaks. Notably, it caused the deaths of 400 bamboo rats on a farm in Guangdong, China, presenting with vomiting and diarrhea [87]. Similarly, infections in rabbit farms in China have led to mass fatalities, with affected animals showing lethargy, yellow watery diarrhea, and multi-organ tissue damage by strain HN001 [88] and strain T2018 [89]. In non-human primates, P. mirabilis infection in rhesus monkeys resulted in diarrhea and bloody stools [50]. Furthermore, a specific strain (17f) was identified as the primary cause of diarrhea in lambs in Xinjiang, China [57]. Beyond the gastrointestinal tract, P. mirabilis can cause respiratory disease in animals; in pigs, specific strains have been associated with respiratory symptoms such as fever and difficulty breathing [90,91]. Alarmingly, studies in pigs also indicate that P. mirabilis can cross the placental barrier, leading to fetal death [90,91].
Aside from gastrointestinal difficulties, Proteus spp. Cause a variety of other disorders in chickens. These include embryonic death, yolk sac infections, and mortality among young chickens, turkeys, and ducks [92]. They also cause granulomatous inflammation in the salt glands of waterfowl, quails, and broilers, as well as disorders like arthritis, salpingitis, airsaculitis, and septicemia, which results in carcass condemnation and economic losses for the poultry sector [92]. The bacteria are also responsible for serious, less common illnesses in other animals; for instance, Abdollahi et al. and Najd et al. were the first to report cases of P. mirabilis-induced pyoderma and purulent pericarditis in sheep [93,94]. More generally, P. mirabilis is involved in different diseases in animals, which include UTIs, wound infections, and gastrointestinal disorders, with special significance in poultry and livestock [95].
P. mirabilis has pathogenicity in many types of tissues and organs, and very serious infections may lead to death. The most important role of P. mirabilis is that it is one of the common pathogens that cause UTIs in humans and in domesticated pets (e.g., cats and dogs) [96]. Herout et al. reported a high prevalence of P. mirabilis in a mouse model of CAUTI, demonstrating that P. mirabilis is perpetually involved in UTIs in broad range of species [97]. The risk of becoming infected with P. mirabilis is equally high for both animal and human populations. The continued rise of antibiotic-resistant strains of P. mirabilis in both animals and humans reinforces the importance of understanding how P. mirabilis causes disease and the mechanisms by which it acquires antibiotic resistance. The information presented in this review supports the conclusion that P. mirabilis is a significant contributor to the development of gastrointestinal disease and UTI in animals and therefore poses a serious threat to food safety and animal welfare and public health. In addition to infecting the gastrointestinal and urinary systems, many other diseases caused by P. mirabilis involve many other organ systems. Furthermore, the broad geographic and host range of P. mirabilis—now conclusively extended to include aquatic ecosystems where it acts as a primary pathogen with concerning multidrug-resistance profiles—along with its pervasive contamination of the food chain, highlights its dynamic ecology and formidable potential for cross-species transmission. This evidence base solidifies the imperative for a comprehensive “One Health” approach to combating the health problems associated with P. mirabilis. Finally, the array of environmental conditions that can sustain P. mirabilis indicates that this bacterium has a wide range of opportunities to infect new hosts and cause disease.
4. Geographic and Host Variations in Virulence-Associated Genes and Factors of P. mirabilis
The virulence arsenal of P. mirabilis is not only central to its pathogenicity in human infections but also plays a critical role in animal diseases, creating a common biological foundation for cross-species transmission and persistence within the One Health framework. The pathogenic potential of P. mirabilis is underpinned by a diverse repertoire of virulence-associated genes (VGs), whose distribution varies by both geography and host species. Epidemiological studies indicate notable variation in VGs profiles. For instance, isolates from North America (United States and Canada) and from poultry sources have been reported to harbor higher numbers of VGs compared to other regions and hosts, highlighting the role of specific reservoirs in shaping virulence [95].
A complex collection of factors is critical for the pathogenicity, persistence and colonization of P. mirabilis (Figure 2), particularly for the urinary tract. Some of these virulence factors include the ability to adhere to tissue, form a biofilm, produce hemolysin and urease, exhibit flagellar motility, swarm and utilize fimbriae to attach themselves [13,18,27,33].
Initial attachment occurs through attachment pili, flagella, and fimbriae, while outer membrane proteins, such as adhesins and lectins, attach to surfaces of the host. There are specific mechanisms necessary to attach to uroepithelial cells that result in the production of UTIs where bacteria are able to grow within the bladder, overcome the effects of urine flow, and establish chronic infection [98]. In addition to adhesive factors, some of the virulent strains develop protective capsules that protect themselves from host defense mechanisms, such as phagocytes [99], while others utilize antigenic variation to evade immune response [100]. This coordinated interplay of virulence factors enables P. mirabilis to overcome host defenses, establish infection, and often lead to chronic, difficult-to-treat conditions. A concise overview of the core virulence machinery of P. mirabilis is presented in Table 1.
As summarized in Table 1, the pathogenicity and clinical severity of a P. mirabilis infection are not strictly dependent on the simultaneous presence of all virulence factors but are rather determined by a dynamic and often synergistic interplay among them. This sophisticated molecular interplay efficiently enhances its virulence and underscores its significance in disease development [18]. While basic colonization may be achieved through key adhesins (e.g., MR/P fimbriae) and motility, the progression to severe, complicated disease—such as catheter-associated pyelonephritis, struvite urolithiasis, or systemic sepsis—typically requires a coordinated expression of multiple factors [21]. For instance, urease activity is indispensable for stone formation and catheter encrustation, hemolysin and protease are critical for tissue invasion and immune evasion during systemic spread, and robust biofilm formation underpins chronicity and treatment failure. The swarming phenotype acts as a master regulator, often upregulating the expression of other virulence determinants (urease, protease, hemolysin), thereby linking motility directly to enhanced pathogenic potential.
From a practical and diagnostic perspective, the pathogenic risk posed by a clinical P. mirabilis isolate can be inferred by profiling its virulence gene repertoire. Molecular screening for key genes—such as those encoding urease (ureC), hemolysin (hmpA), MR/P fimbriae (mrpA), protease (zapA), and biofilm-associated functions—allows for stratification of isolates into risk categories [18]. Isolates harboring a full complement of these genes are classified as high-risk, particularly in settings involving indwelling devices or immunocompromised hosts, as they possess the genetic arsenal for persistent colonization, tissue damage, and antibiotic tolerance. Conversely, isolates lacking critical genes (e.g., ureC) may be considered lower-risk for causing complicated infections, though they retain the capacity for acute cystitis. This genotype-to-phenotype correlation underscores the potential of virulence factor profiling as a tool for prognostic assessment, guiding infection control measures, and personalizing therapeutic strategies in both human and veterinary medicine within the One Health framework. The subsequent Section 4.1, Section 4.2, Section 4.3, Section 4.4, Section 4.5 and Section 4.6 provide a detailed examination of each major factor, elucidating its specific role in pathogenesis and its contribution to the bacterium’s success across different hosts within the One Health paradigm.
4.1. Urease as a Central Virulence Factor in P. mirabilis Pathogenesis
P. mirabilis exploits a range of virulence agents, with the cytoplasmic nickel metalloenzyme urease being particularly important for pathogenesis. This enzyme, situated in the cytoplasm or outer membrane, catalyzes the hydrolysis of urea into ammonia and CO_2_, resulting in a large local pH increase [18,101]. The ammonia produced is extremely alkaline and immediately cytotoxic to mammalian cells, producing significant tissue damage and compromising the uroepithelium, facilitating bacterial invasion [102].
The relationship between urease activity and pathogenicity is well explained in the development of urinary stones [103]. These stones create a niche in which bacteria are protected from human immune response and drugs, thereby leading to persistent and recurrent infections [12].
Urease activity in medical devices precipitates minerals, which interact with bacteria on urinary catheters and produce a crystalline biofilm that obstructs urine flow [104], also to the development of acute pyelonephritis [12,27]. The crystalline biofilm is responsible for the encrustation and blockage of catheters, a common result from CAUTIs. The bacteria embedded in such crystalline formations are resistant to drugs as well as to the human immune response. In a polymicrobial environment, interactions may become synergistic, enhancing overall pathogenicity even when bacterial loads are equivalent to that of mono-species infections [104]. The understanding of these detailed virulence pathways offers promising options for the design of future therapies against UTIs [105]. This stone-forming capability, critical in human CAUTIs, poses similar risks in animals with urinary catheters or anatomical abnormalities, underscoring a shared pathological mechanism across species. The P. mirabilis urease gene cluster consists of structural genes such as ureABC and the urea-induced regulator ureR, which can be detected by PCR techniques [106].
4.2. Adhesive Structures and Biofilm Formation in P. mirabilis: Key Fimbriae and Their Roles
Fimbriae are hair-like structures on the bacterial cell surface; they are essential for the adhesion of P. mirabilis bacteria to the bladder and renal epithelium and to other inert substances such as catheters [18,21]. Chaperone-usher fimbriae are particularly important for the molecularly precise adhesion and colonization of the urinary tract epithelium by bacteria [18]. These organelles are critical for biofilm production, assisting in immune evasion and persistent infection, and contribute to crystalline biofilm and catheter encrustation characteristic of P. mirabilis UTIs and infectious stone formation, highlighting their pathogenic relevance [18,107].
Important fimbrial adhesins are necessary for initial colonization and biofilm formation. These include P. mirabilis fimbriae (PMF, pmfA), Ambient-Temperature Fimbriae (ATF, atfA), Uroepithelial Cell Adhesin (UCA, ucaA), Mannose-Resistant/Proteus-like (MR/P) fimbriae (encoded by mrpA), and Non-Agglutinating Fimbriae (NAF) [108]. The genes mrpA, pmfA, ucaA, and atfA are key molecular targets detectable using PCR [109]. MR/P fimbriae are particularly important for bladder and kidney biofilm production and colonization and play a critical role in catheter biofilm establishment [21].
PMF fimbriae, first discovered in strain HI4320 [110], are made easier to assemble and locate in the urinary tract by a five-gene operon (pmfA, pmfC, pmfD, pmfE, and pmfF) [108,111]. Other fimbriae allow for niche-specific colonization: UCA improves adherence to uroepithelial cells, NAF boosts in vitro adhesion, and ATF promotes environmental survival through optimum expression at lower temperatures [108]. P-like pili (PMP) are another adherent agent that may help with urinary tract adhesion [112], and MR/K hemagglutinin, enabling catheter attachment [102].
Aside from chaperone-usher fimbriae, P. mirabilis has a possible type IV pilus system—a dynamic structure implicated in virulence, twitching motility, and biofilm generation, with functional similarities to MR/P fimbriae [18,113]. Genomic investigations reveal one or two probable type IV pilus loci [114]. This system acts by pilin polymerization/depolymerization and includes components including major/minor pilins, a pre-pilin peptidase, assembly/retraction ATPases, a secretin and auxiliary proteins [115]. The adhesive and biofilm-forming capabilities of P. mirabilis are equally critical in veterinary settings, facilitating persistent urinary tract and wound infections in companion and livestock animals, thereby contributing to the spread of resistant strains across species.
4.3. Swarming Motility: An Overview
Proteus spp. have a dimorphic life cycle, which is essential for their pathogenicity. They appear in liquid settings as short, motile swimmer cells with peritrichous flagella. When cells come into contact with a solid surface, they undergo complicated differentiation and become elongated (20–80 µm), hyperflagellated, multinucleated swarmer cells. This differentiation entails continuous DNA replication in the absence of cell division, resulting in polyploid filaments. These cells move together in coordinated rafts and form characteristic concentric rings on agar. The migration cycle of swarming cells is divided between periods of migration and consolidation, wherein cells temporarily readopt a shorter cell form prior to further differentiation or not. This process, important for the movement into new areas and host colonization [108,110,116]. Elaborating on this key characteristic, the regulatory dynamics of swarming involve the differentiation of P. mirabilis controlled by a network activated by specific signals. The primary trigger is surface contact, sensed via restricted flagellar rotation, which upregulates regulators like umoB and lrp [117]. Without this signal, cells remain as short “swimmers” [33]. These regulators drive genes for elongation, hyperflagellation, and polysaccharide production [117]. Chemical inducers include L-glutamine, other amino acids, and putrescine [33]. Zinc homeostasis is also crucial [118]. Conversely, high osmolarity represses swarming, and high urea may inhibit swarming in vivo [36]. Swarming upregulates virulence factors: urease, zapA protease, and hemolysin [117], promoting stone formation [119], host cell lysis [117], and protein degradation. Additionally, swarming initiates biofilm formation on catheters [120], leading to persistent infections.
Swarming has a closely regulated mechanism associated with higher pathogenicity. In the transition from swimming to swarming mode, there is a higher expression of flagella and a regulated expression controlled by a tier of genes, with the primary role associated with the master regulatory genes flhDC. Another key gene associated with the unique feature of cell elongation is ccmA. Beyond the swarming motility is the cell’s metabolic and pathogenic shift for differentiated swarmer cells. Compared with swimmers, there is a higher production of key virulence factors urease, protease, hemolysin (hmpA), and IgA-degrading metalloprotease (zapA) associated with swarmer cells. The association between motility and the production of virulence factors underlines the vital role of swarming in the infection process [33,117].
The swarming of P. mirabilis is extremely responsive to environmental stimuli, especially those presented by its host. Certain chemicals in human urine have been identified as effective stimulators of swarm motility, including arginine, glutamine, histidine, malate, and ornithine. Another significant stimulator is the polyamine putrescine, whose biochemical pathway is unusually connected with urea metabolism-one of the key processes underlying pathogenesis in the urinary tract. This indicates that the urinary system’s chemical environment imposes a direct influence on the invasive behavior of the bacterium. Additionally, physical mobility across surfaces by swarm rafts is aided by chemicals such as capsular polysaccharides and slime [12,108,113].
The swarming phenotype has significant clinical implications, especially in CAUTIs. P. mirabilis can quickly spread across the surfaces of silicone or latex urinary catheters, allowing for fast colonization and ascent into the bladder and kidneys. This surface movement is thought to be associated with increased expression of virulence genes, potentially beginning infection [121]. Swarmer cells are significantly more invasive to urothelial cells than vegetative swimmer cells. Flagella-driven motility has an important role in virulence, as evidenced by research demonstrating that flagella-negative mutants are much less harmful. As a result, the ability to swarm is an important contributor to the creation of strong biofilms, the spread of infection, and improved antibiotic resistance in chronic infections [12,18,120]. This swarming capability, while extensively studied in human CAUTIs, likely facilitates similar rapid colonization and biofilm formation in animal hosts with urinary catheters or anatomical susceptibilities, highlighting a conserved invasion strategy.
4.4. Hemolysin as a Critical Virulence Factor in P. mirabilis: Roles in Cytolysis, Iron Acquisition, and Pathogenesis
Hemolysin is an important multifunctional virulence factor in P. mirabilis, principally through the action of a calcium-independent, Serratia-like toxin produced by the hmpA gene. The production of this hemolysin is promoted by its specialized transporter, encoded by the highly conserved hmpB gene, indicating the importance of this system in pathogenesis [108,117]. These genetic markers (hmpA and hmpB) are successfully discovered using polymerase chain reaction [109]. Hemolysins, which have a wide range of cytotoxic effects, possess the capability to lyse not only nucleated host cells but also erythrocytes [122]. This membrane-disrupting capability serves two functions: it liberates important nutrients such as iron from red blood cells and thus supports bacterial survival; and it promotes the invasion of tissues and the evasion of immunity through the destruction of host defense cells and the creation of pathways for disseminating bacteria within the host [12,117]. The significant reduction in virulence observed with hmpA knockout mutants illustrates that the hemolysin contributes significantly to the process of infecting a host [18,123]. Thus, it can be concluded that hemolysins are a major factor in determining both the severity and persistence of infections caused by P. mirabilis, making the mechanisms by which they function an appropriate target for the development of potential therapeutic interventions [31,108]. The Iron-acquisition function of hemolysin is particularly vital for bacterial survival in the iron-limited environments of both human and animal hosts, highlighting a conserved virulence strategy.
4.5. Protease-Mediated Immune Evasion: A Key Virulence Strategy of P. mirabilis
P. mirabilis employs proteases as virulence factors to overcome the highly unfavorable urinary tract environment successfully [124]. Of great significance among these proteases is zapA (mirabilysin), a potent metalloprotease that directly damages the host’s immunity by selectively degrading vital host defense proteins like immunoglobulins—IgA and IgG—as well as complement factors [124]. When this zapA is overexpressed during swarming differentiation processes, its production is consistently high at the edge of a motility bacterial swarm [125]. These targeted actions of proteases and especially zapA’s degradation of vital host factors have benefited bacterial escape and persistence and are consequently important in the host’s defense against this pathogen and have been identified as a biotarget/drug target for potential treatment [12], whose development as a drug target is further aided by the discovery of targeted chemicals against this enzyme [124]. This immunoglobulin-degrading strategy, crucial for evading human immune defenses, likely supports bacterial persistence in animal hosts as well, underscoring a conserved immune evasion mechanism within the One Health context.
4.6. Biofilm Formation and Quorum-Sensing Regulation in P. mirabilis: A Foundation for Persistent Infection
The process begins when free-floating (planktonic) bacteria encounter a surface. This initial attachment is reversible and weak, driven by physical forces like van der Waals interactions, hydrophobic effects, or electrostatic charges. Bacteria use appendages such as flagella or pili to approach the surface [11]. Once in contact, specific adhesins (proteins on the bacterial surface) may bind to the substrate, making the attachment more secure and irreversible. Environmental factors, like nutrient availability or surface conditioning (e.g., a layer of organic molecules), can influence this step. After attaching, bacteria begin to proliferate and form microcolonies. They divide and produce extracellular polymeric substances (EPSs), a sticky matrix of polysaccharides, proteins, and DNA that anchors them to the surface and to each other. This EPS layer not only provides structural support but also traps nutrients and protects the growing community from external threats like antibiotics or immune responses.
As the biofilm grows, it develops into a complex, three-dimensional structure. This stage involves the formation of water channels within the EPS matrix, which facilitate nutrient and oxygen distribution while removing waste. Genetic regulation fine-tunes the community, with some cells differentiating into distinct roles (e.g., persister cells that resist stress). At this point, the biofilm becomes highly resistant to antibiotics, up to 1000 times more so than planktonic cells, due to the protective EPS and slower metabolic rates of deeper layers [12]. EPS plays a central role in this biological process, and it contributes significantly to biofilm formation as it plays a role in cell and surface attachment and protects against antimicrobials, host immunological defense, and oxidative stress [126].
EPSs mask pathogen-associated molecular patterns (PAMPs), reduce phagocytosis, and neutralize antimicrobial peptides [127]. However, biofilms are most feared in indwelling medical devices such as urinary catheters and implants, which act as a protective barrier referred to as CAUTIs [12,18,127]. Act as reservoirs for chronic infections, necessitating device removal due to resistance to Treatment [128]. Biofilm-mediated UTIs demonstrate high recurrence rates and elevate risks of ureteral injury, drainage dysfunction, and renal failure [129]. Polymicrobial Interactions and functional redundancy further stabilize biofilm signaling under environmental stressors, though disruptions in microbial diversity can impair QS-mediated Cross-kingdom communication [36,113].
Biofilm Antibiotic Tolerance (BAT). As a thumb rule, bacteria in biofilms are really more resistant to antimicrobial intervention than their counterparts in planktonic habitats [22]. Therefore, BAT is expected to incorporate alternate paths to bacterial antimicrobial resistance [25]. Additionally, reduced metabolic activity in deeper biofilm layers diminishes antibiotic Efficacy, while efflux pumps and persister cells enhance survival [122,123,124]. Persister cells further reinforce biofilm resilience, which enters dormancy via a stringent response (ppGpp) and toxin–antitoxin systems (e.g., HipBA). These cells evade antibiotics and repopulate biofilms post-treatment [110,111]. Agr-Mediated dispersal via phenol-soluble modulins (PSMs) contrasts with SarA’s promotion of polysaccharide intercellular adhesin (PIA) [112].
Biofilm regulation is a multifaceted process governed by genetic, biochemical, and Environmental mechanisms, enabling microbial communities to adapt to diverse conditions [92]. Central to this regulation are QS systems and second-messenger molecules, which coordinate the development of biofilms across bacterial species [93]. Cyclic di-GMP (c-di-GMP) is a pivotal second messenger that promotes biofilm stability by upregulating adhesins, pili, and EPS production while suppressing motility [98].
In P. mirabilis, the QS system is not fully characterized compared to other GNB. While classic acyl-homoserine lactone (AHL)-based LuxI/LuxR systems are absent [13], the bacterium possesses the luxS gene responsible for producing autoinducer-2 (AI-2) [130]. However, disruption of luxS does not significantly affect swarming motility or virulence in murine models, suggesting functional redundancy or the presence of alternative signaling pathways [33,130]. Notably, P. mirabilis can respond to exogenous AHLs (e.g., N-butanoyl homoserine lactone), which influence biofilm architecture, indicating cross-species communication capabilities [128]. In P. mirabilis, QS governs essential characteristics such as the production of virulence factors (e.g., urease, protease, hemolysins) and the growth of the biofilm itself [12,131]. Beyond these roles, QS is implicated in broader physiological processes such as metabolic adaptation and stress responses. Critically for clinical outcomes, QS also contributes to the challenge of antimicrobial resistance through QS-mediated biofilm formation, which promotes antibiotic tolerance by limiting drug penetration and fostering resistant subpopulations. Furthermore, the dispersal phase triggered by QS can facilitate the spread of resistance genes via horizontal gene transfer, as observed in other biofilm-forming pathogens [132,133]. Reverse correlation of biofilm formation ability and swarming motility was estimated. Based on the study findings It is hypothesized that P. mirabilis benefited from adhesins such as MR/P fimbriae for production of biofilm and successful colonization and then they shift from biofilm formers to strong swarmers in order to reach deeper urinary organs and hlyA toxin is used to overcome the immune system cells [134].
In response to these challenges, the scientific community has shifted its focus toward exploring alternative antimicrobial agents that can target biofilms through multifaceted mechanisms [135,136]. Plant-derived essential oils (Eos) have emerged as a promising frontier. These complex phytochemical mixtures exhibit broad-spectrum antimicrobial activity, disrupting biofilms through multiple pathways: destabilizing microbial membranes; inhibiting QS—a critical communication system that regulates biofilm development; and degrading Extracellular matrix components [137,138]. Furthermore, Eos often exhibit synergistic effects When combined with conventional antimicrobials, thereby enhancing drug efficacy while minimizing the development of resistance [139]. Curcumin, which is also derived from Curcuma longa (turmeric), is an anti-quorum Sensing agent that inhibits P. mirabilis [140]. Other phytochemicals, such as linalool, derived from floral plants, also inhibit Motility and reduce biofilm-associated extracellular polysaccharides. These compounds could potentially enhance the effectiveness of conventional antibiotics and offer a promising alternative or complementary strategy for controlling P. mirabilis infections and biofilm-related Complications [141].
5. Antimicrobial Resistance in P. mirabilis: Mechanisms, Drivers, Epidemiology, and Transmission from a One Health Perspective
5.1. Introduction: The Global and One Health Burden of AMR
The global spread of antimicrobial resistance (AMR) in P. mirabilis is a quintessential One Health issue, driven by interconnected human, animal, and environmental reservoirs. AMR poses a serious global challenge, with approximately 60% of all human pathogens and 75% of emerging human infectious diseases having zoonotic origins, and it is noted that most life-threatening human diseases originate from animals [142,143]. The global burden of AMR is staggering, causing an estimated 4.95 million deaths in 2019, of which 1.27 million were directly attributable to resistant infections [144]. Projections suggest AMR could lead to 10 million deaths annually by 2050 if unchecked, with associated economic losses of up to 100 trillion annually [[145](#B145-microorganisms-14-00444)]. The implications involve the loss of 60–100 trillion in global output and threaten the achievement of the Sustainable Development Goals [[146](#B146-microorganisms-14-00444)]. The World Health Organization ranks AMR among the top ten global public health threats [[3](#B3-microorganisms-14-00444),[147](#B147-microorganisms-14-00444)]. In the United States alone, about 2.8 million antibiotic-resistant infections occur each year, resulting in 35,000 deaths and 20 billion in direct healthcare costs [148]. Emerging hotspots for AMR include China, India, Brazil, and Kenya [149,150]. Furthermore, high rates of resistant bacteria in humans are compounded by challenges related to antibiotic use in both animal and human healthcare [150].
The development of AMR represents a significant worldwide public health concern that transcends healthcare and agricultural boundaries [151]. The One Health approach—an integrated strategy considering the interconnected health of humans, animals, and ecosystems—is crucial for tackling infectious diseases like those caused by multi-resistant P. mirabilis, which proliferates across hospitals, farms, and the environment [12].
The factors contributing to AMR are diverse and rising exponentially. A primary driver of this crisis is the inappropriate or excessive use of antimicrobials in human healthcare, including over-prescription and self-medication. Inappropriate antibiotic use in primary care settings is highly prevalent, ranging from 15.4% in Canada to 88% in Pakistan [149]. A multi-hospital cohort study found that nearly two-thirds of COVID-19 patients received empirical antibiotics, despite only 3.5% having a confirmed bacterial co-infection [152].
Concurrently, the intensive use of antimicrobials in animal husbandry for therapeutic, prophylactic, and growth-promotion purposes is a critical driver [146]. Globally, animals consume more antibiotics than humans, with livestock accounting for over 73% of total antimicrobial use [149,153]. Notably, major antibiotic classes like tetracyclines, sulfonamides, and fluoroquinolones are excreted in significant proportions unchanged in manure and urine [154,155]. Due to their environmental persistence and low bioavailability in animal guts [156], they maintain biologically active concentrations in soil and water. These pharmacological properties allow them to exert sustained selective pressure for extended periods, thereby promoting the development and maintenance of environmental reservoirs of resistance genes [157]. A major concern is that a considerable portion of administered antibiotics is excreted and can persist in the environment, continuously exerting this selection pressure [155]. While some antimicrobial classes, such as carbapenems, are used exclusively in humans, and others like flavophospholipol and ionophores are exclusively for animals [158,159], most drug classes used in humans, including critical ones like quinolones and broad-spectrum beta-lactams, are also administered to animals [153].
Global antimicrobial consumption is soaring, largely in the human health sector and animal farming. Between 2000 and 2015, global antibiotic usage climbed by 65%, with a 39% rise in consumption rates [160]. The most significant increases were observed in low-income nations, which showed a 56% growth in use, with massive jumps in the consumption of cephalosporins (399%), quinolones (125%), and macrolides (119%) [161]. Estimates also reveal that antimicrobial use in BRICS countries will go up by an astounding 99%, driven by rapid population growth [158]. Current total use stands at about 131,000 tons annually and is set to rise by over 67% to nearly 200,000 tons by 2030 [162]. In 2013, an estimated 131,109 tons of antimicrobials were used in food animals, with the figure expected to climb to 200,235 tons by 2030 [163]. By 2030, antimicrobial use in livestock is anticipated to increase by more than 67% from 2010 levels (around 63,000 tons), exceeding 105,000 tons [158]. The average global annual consumption per kilogram of animal produced is estimated to reach 45 mg/kg for cattle, 148 mg/kg for chickens, and 172 mg/kg for pigs [158]. It is noteworthy that antimicrobial use intensity (mg per kg of animal produced) in cattle was estimated to have decreased by 34% in 2021 [164], although rising overall consumption continues to drive resistance rates.
This widespread antimicrobial selection pressure has a direct and measurable consequence: the proliferation of multidrug-resistant bacteria across the food chain. In aquaculture, a critical unifying concern is multidrug resistance. Isolates from African catfish (Clarias gariepinus) have shown high phenotypic resistance indices, with a MAR index of 0.60 [69]. Genomic analysis has confirmed this threat at the molecular level, with whole-genome sequencing of a pathogenic strain from Indian major carp (Labeo catla) revealing it to harbor multiple antimicrobial resistance genes [72]. The contamination of the food supply is equally alarming. A genomic characterization of P. mirabilis strains from retail meat and aquatic products in China revealed high genetic diversity. Alarmingly, 91% of the strains exhibited multidrug resistance profiles, carrying a wide array of clinically important resistance genes, including blaCTX-M, cfr, and even genes conferring resistance to tigecycline (tmexCD3-toprJ1) and carbapenems (blaNDM-1) [82].
AMR poses a severe threat to global economic growth. Projections indicate it could reduce global Gross Domestic Product (GDP) by up to 3% by 2030 and by 1.1% to 3.8% by 2050 [82,146]. Another study suggests the global GDP decline by 2050 could range between 2% and 3.5% [165]. This near-term impact would coincide with an additional 700 billion in global healthcare expenditures that year, a burden disproportionately borne by low-income countries [[82](#B82-microorganisms-14-00444),[165](#B165-microorganisms-14-00444),[166](#B166-microorganisms-14-00444)]. Long-term cumulative costs could reach up to 100 trillion [165]. The financial repercussions are significant at regional levels as well, as highlighted by a European Centre for Disease Prevention and Control (ECDC) report citing approximately €1.5 billion in annual additional patient care expenses due to AMR-related infections [167]. The financial strain arises from the complexities of managing drug-resistant infections, which necessitate longer hospital stays, more expensive alternative drugs, increased morbidity, and reduced productivity [168]. Concerns are mounting regarding the escalating financial burden of treating MDR infections. This escalating burden risks exacerbating poverty and hindering progress toward the Sustainable Development Goals. Addressing this complex threat requires an urgent and effective implementation of the One Health approach, recognizing the inextricable links between human, animal, and ecosystem health [169].
5.2. The Resistance Profile of P. mirabilis: An Overview
The global state of antimicrobial resistance (AMR) in P. mirabilis is both dynamic and alarming, with nearly 48% of strains exhibiting antibiotic resistance [170]. This pathogen accumulates additional antimicrobial resistance genes (ARGs) via horizontal gene transfer, driving the development of multidrug-resistant (MDR) and extensively drug-resistant (XDR) strains [95]. Key ARGs facilitating this propagation include blaTEM, blaCTX-M, blaKPC, blaNDM, blaVIM, and mcr-1 [171]. P. mirabilis possesses a formidable resistance profile encompassing both innate and acquired mechanisms. It is intrinsically resistant to nitrofurans, polymyxins (including colistin), tigecycline, and tetracycline [172], and has developed acquired resistance to numerous other classes including trimethoprim/sulfamethoxazole, aminoglycosides, carbapenems, fluoroquinolones, β-lactams, imipenem, cephalosporins, penicillins, and aztreonam [30,170]. This adaptability is bolstered by its role as an effective ARG reservoir [173].
Crucially, β-lactam resistance is mediated by the synthesis of ESBLs and ampC β-lactamases, which hydrolyze penicillins, cephalosporins, and aztreonam [174]. ESBL-producing isolates frequently exhibit co-resistance to other classes, such as quinolones and aminoglycosides [175]. Furthermore, chromosomal mutations can lead to phenotypic resistance to all β-lactam drugs [170]. Genomic studies confirm a wide array of resistance genes [176], with increasing reports of strains harboring multiple ARGs, including those for quinolones (e.g., qnr, aac(6′)-Ib) and aminoglycosides (e.g., APH, AAC, AAD) [170].
Surveillance data reveal a severe and complex resistance landscape with significant geographical and host-based variation. Isolates from China harbor the highest ARG counts, while those from the United States contain the fewest [95]. Animal-derived isolates consistently possess more ARGs than human clinical isolates, with urine isolates showing the greatest ARG diversity among human samples [95]. This interconnectivity is highlighted by the presence of similar ARGs in raw meat, migratory birds, and human clinical samples [177]. The rate of amoxicillin-resistant P. mirabilis (38–48.5%) in medical institutions parallels that of E. coli [170,178], and it ranks as the second most prevalent ESBL-producing Enterobacteriaceae in poultry [179]. This widespread resistance stems from the organism’s high genomic diversity, driven by mobile genetic elements, mutations, and genomic rearrangements, which yield highly diverse virulence and resistance factors even among isolates from the same source [180,181,182]. Such diversity poses a significant challenge to diagnosis, infection control, and treatment [183,184]. The genetic basis of this resistance is further elucidated by the wide array of antibiotic resistance genes (ARGs) identified in P. mirabilis isolates from diverse animal reservoirs. Surveillance studies have mapped specific ARGs to their animal sources, revealing a complex transmission network [86]. More concerning is the identification of genes conferring resistance to last-resort or critically important antibiotics. The polymyxin resistance gene mcr-1 has been reported specifically in poultry isolates [57]. The carbapenemase gene blaKPC has been found in farm animals [52,55]. The fosfomycin resistance gene fosA3 has been identified in livestock and food products [53,61,185], though its distribution shows geographical variation; for instance, recent surveillance in Egyptian livestock (buffaloes and broiler chickens) reported an absence (fosA3-negative) of this gene in these populations [79]. This detailed genotypic landscape underscores how animal reservoirs serve as melting pots for the accumulation and dissemination of ARGs, with clear implications for zoonotic transmission and food safety.
The emergence and circulation of resistant P. mirabilis in animals and the food chain pose a major zoonotic and foodborne threat, epitomizing a One Health challenge. Zoonotic transfer is facilitated by the inseparable interconnectivity of humans, animals, and shared environments, enabling constant exchange of resistant organisms and their genetic elements [186]. While zoonotic pathogens are linked to nearly two-thirds of major recent infectious disease outbreaks, successful cross-species transmission depends on a complex interplay of ecological and genetic factors [186].
Transmission occurs via interconnected environmental and direct routes. Resistant strains from livestock operations enter the environment through wastewater effluent and agricultural runoff, contaminating water and crops and creating secondary exposure routes that hinder containment [187]. Supporting this, a scoping review that analyzed 70 out of 3367 identified studies on transmission pathways concluded that antibiotic residues, heavy metals, and microbial interactions in wastewater are key drivers of AMR. It also noted that wastewater treatment plants (WWTPs), while designed to reduce contaminants, can create conditions favoring horizontal gene transfer, thereby amplifying resistance genes [188]. This environmental pathway, often involving untreated waste, plays a significant role in accelerating the AMR pandemic and underscores the critical need for enhanced vigilance regarding the environmental dimension of infectious diseases, a lesson reinforced by the COVID-19 pandemic [189].
Human infection occurs through multiple pathways, including direct contact with infected animals (a major occupational risk for farm workers), contact with contaminated surfaces, water, or soil, and—importantly—ingestion of contaminated animal products [80]. The contamination of poultry and meat products is a particularly effective route for introducing virulent, resistant strains into the human food chain [80]. An alarming scenario is the potential for meat from broiler chickens or buffalo carrying PDR P. mirabilis—non-susceptible to all antimicrobial categories—to act as a direct reservoir for transmitting untreatable strains to humans, causing severe, therapy-limited infections like complicated urinary tract infections [79]. The emergence of PDR strains represents the final, most critical stage in the resistance continuum (MDR → XDR → PDR). Genomic evidence showing strong relatedness between resistant animal and human clinical isolates confirms a common origin and transmission mechanism [79].
Surveillance has identified major livestock as key reservoirs of resistant P. mirabilis, including cattle [190,191,192,193], buffalo [79,194,195,196,197], pigs [198,199], poultry [78,79,81,200,201,202], ducks [78,202], and dogs [58,59,203,204,205]. The risk to human populations is disproportionately higher in low- and middle-income countries due to inequities in food safety standards, regulatory enforcement, and healthcare access. The persistent detection of clinically relevant resistant P. mirabilis in food animals and retail meat underscores its potent zoonotic threat and the non-negotiable need for a unified, cross-sectoral One Health strategy to manage its spread [95].
The rising tide of MDR strains necessitates a coordinated One Health response. Despite advances, significant gaps remain in data on AMR from animal and environmental sources and their transmission mechanisms [206]. The predominance of MDR strains, which outcompete first-line therapies, highlights the urgent need for novel antibiotic targets and illustrates the profound clinical and public health consequences of AMR in P. mirabilis. Modelling suggests that reducing human antibiotic consumption could decrease resistant colonization by 65.7–99.7% over two decades [207], underscoring the need for improved infection control in both healthcare and agriculture [208]. Effective AMR management requires interdisciplinary collaboration, integrated surveillance, and antimicrobial stewardship across all sectors.
Recent large-scale genomic surveillance of 3403 high-quality genomes from 58 countries confirms that human clinical isolates (especially from UTIs) are the primary resistance reservoir [95]. This study identified a vast repertoire of 239 distinct ARGs, with β-lactamase and carbapenemase genes being exceptionally widespread, and reaffirmed the highest ARG burden in Chinese isolates [95]. Phylogenetic analysis grouped global isolates into 17 clusters, with U.S. isolates showing the widest spread. The minimal genetic variation between isolates from different countries and hosts strongly suggests transnational and cross-host clonal spread, reinforcing the interconnected nature of AMR within the One Health framework [95]. A comprehensive understanding of this escalating threat necessitates a deeper examination of the specific molecular mechanisms empowering P. mirabilis across human, animal, and environmental settings.
5.3. Molecular Mechanisms of Antimicrobial Resistance in P. mirabilis: A Unifying Framework
To overcome antimicrobial compounds, bacteria employ several fundamental molecular strategies. These include modifying the target site to prevent the antimicrobial from binding, inactivating or reducing the compound to ineffective levels through enzymatic degradation, actively expelling the antimicrobial via efflux pumps, and reducing cellular uptake of the compound, typically achieved through an impermeable outer membrane [209,210,211] (Figure 3). P. mirabilis belongs to GNB, has an intricate cell envelope structure composed of a thin layer of peptidoglycan. More importantly, it has an outer membrane rich in lipopolysaccharides. This renders it strong biological armor, making it difficult for antibiotics to penetrate [212].
Horizontal gene transfer (HGT) through mobile genetic elements (MGEs) helps in the dissemination of resistance gene traits among and within bacterial populations after they emerge. These MGEs, which include plasmids, transposons, and integrons, essentially serve as vectors carrying the swift dissemination of ARGs among various species of bacteria in the hospital and community settings [40,170,213,214]. This transfer can occur via mutational alterations or direct interbacterial contact, resulting in treatment failure and resistance growth [215]. Alternatively, the effective resistant strains’ expanding clones propagates these determinants [216]. Interestingly, MGE mobilization and resistance co-selection can be caused by various environmental stresses in addition to antimicrobials. Exposure to heavy metals, oxidative stress, or ultraviolet light might cause the transfer of genetic elements, leading in the selection of ARGs alongside genes giving resistance to these non-antibiotic hazards [217,218]. Antimicrobial resistance essentially occurs when a bacterium develops a physiological adaptation to counteract the effects of a medication, either by structural alterations, enzyme inactivation, or other mechanisms [212]. These skills fall into two main categories that are relevant to P. mirabilis: acquired resistance, which is obtained by genetic mutation or the horizontal acquisition of foreign resistance genes, and intrinsic (inherited) resistance, which is a natural trait of the species or genus.
5.3.1. Intrinsic Resistance in P. mirabilis
Intrinsic resistance is the most fundamental and intrinsic kind of bacterial resistance to antimicrobial agents. In P. mirabilis, non-acquired resistance is a species-specific feature resulting from spontaneous mutations or innate physiological traits rather than horizontal gene transfer. It originates from processes such as the absence of a drug’s target, low-affinity target locations, an impermeable cellular envelope, or the natural generation of inactivating enzymes and can emerge even in the absence of direct antimicrobial pressure [219]. One distinguishing feature of P. mirabilis is its innate resistance to specific antibiotic groups. Most importantly, the species is naturally resistant to polymyxin drugs like colistin [170,220]. The fundamental mechanism of resistance is structural changes to the lipopolysaccharide (LPS) in the outer membrane. These changes diminish the net negative charge on the bacterial cell surface, preventing positively charged polymyxin molecules from initially attaching electrostatically. This intrinsic characteristic substantially limits the treatment choices for infections caused by multidrug-resistant P. mirabilis.
Furthermore, P. mirabilis frequently shows decreased susceptibility to the carbapenem antibiotic imipenem [18,220]. While not completely resistant, more significant resistance can emerge due to the loss of outer membrane porins (e.g., ompC, ompF) or reduced expression of certain penicillin-binding proteins (PBPs), especially PBP1a and PBP2. These modifications restrict medication inflow or lower target binding affinity. P. mirabilis is missing natively manufactured chromosomal β-lactamases; hence, any resistance to β-lactam antibiotics is acquired rather than innate. Aside from these specific examples, intrinsic resistance in P. mirabilis and kindred species can be linked to a variety of constitutive variables, such as efflux pump activity that is always low and the presence of protective proteins. Natural resistance to antimicrobial classes, including β-lactams, aminoglycosides, and fluoroquinolones, is linked to the function of various genes, such as ampC, blaSHV, trxA (thioredoxin), and trxB (thioredoxin reductase), among others [221]. A thorough understanding of these internal mechanisms is essential for establishing effective therapeutic options and countermeasures to combat the growing threat of resistant P. mirabilis strains.
5.3.2. Acquired Resistance and Horizontal Gene Transfer in P. mirabilis
Acquired resistance is the key clinical difficulty in managing P. mirabilis infections, owing to the bacterium’s remarkable ability to get resistance genes against a wide range of essential antibiotics, including beta-lactams, fluoroquinolones, and aminoglycosides. Acquired resistance occurs when previously sensitive bacteria gain the ability to resist an antimicrobial agent.
In prokaryotes, lateral gene transfer, or more recently, lateral genetic transfer (LGT), is a crucial mechanism for transferring and rearranging DNA [222]. Based on reports, up to 25% of some bacterial genomes may have originated from LGT during evolutionary periods [223], demonstrating the scope of LGT. The implications of LGT for human health are significant. In fact, some argue that humans are losing the battle against antibiotics and antibiotic resistance [224]. This happens through the acquisition of ARGs via two major pathways: vertical gene transfer (inheritance from a parent cell to its offspring) and, more importantly, HGT. These processes can occur concurrently, but HGT is particularly important because it allows for the direct exchange of genetic material between modern cells, introducing entirely new resistance genes and mechanisms into a bacterial population and leading to improved collective resistance profiles. HGT is a major driver of antimicrobial resistance spread and operates through three primary mechanisms, each contributing differently to resistance dissemination across the human–animal–environment interface, as summarized in Table 2. HGT is a main driver of AMR spread and can occur through three primary mechanisms: transformation (free DNA uptake), transduction (bacteriophage transfer), and conjugation (direct cell-to-cell transfer via plasmids or other conjugative components).
The ability to transfer genes between species accelerates the spread of resistance. Furthermore, it is worth noting that exposure to various physical or chemical stimuli has the potential to cause selected mutations in bacterial DNA, which contribute to resistance development. Different types of bacteria possess different potential applications of acquired resistance mechanisms due to their intrinsic structural differences. For example, GNB like P. mirabilis can possess all four major types of acquired mechanisms: target-site alterations, enzyme inactivation of drugs, active pumping of out-drugging, and prevention of drug uptake. The complex nature of the outer cell envelope, which includes an outer membrane composed of LPS, and also contributes to the ability of GNB to adapt. On the other hand, Gram-positive bacteria do not have this outer membrane, and therefore, they do not rely on the same mechanisms to restrict drug uptake [212]. To comprehend the spread of both currently known and future discovered resistance genes, it is essential to comprehend the genetic components of each type of resistance mechanism, the nature of their life cycle, the dynamics of LTG, and the ecological context in which these mechanisms develop [225]. The next sections explore acquired resistance mechanisms of P. mirabilis.
5.3.3. Key Vehicles of Horizontal Gene Transfer
Mobile Genetic Elements as Vectors of Antimicrobial Resistance in P. mirabilis
MGEs are critical factors in the acquisition and spread of ARGs in P. mirabilis populations. These mobile DNA segments serve as excellent transporters for the horizontal transmission of resistance determinants across bacterial cells, contributing significantly to the rapid development and extensive dispersion of MDR strains. The primary kinds of MGEs involved in this process include many major sorts.
Plasmids and Integrative Conjugative Elements in AMR Dissemination
An important discovery is that P. mirabilis has integrative and conjugative elements (ICEs), such as ICEPm1, which contains both virulence and antibiotic resistance genes. These elements can replicate independently and self-transfer to other strains and even different bacterial species, making them effective agents of genetic exchange [18]. P. mirabilis has become a serious public health problem due to its strong combination of virulence and high resistance levels. Plasmids are a type of MGEs. They are self-replicating, circular pieces of DNA that exist outside the chromosome and can hold many different types of antibiotic resistance genes. Plasmids allow the rapid transfer of these resistance genes between bacterial cells by conjugation (sending genetic material from one cell to another). Conjugation not only allows for the rapid spread of resistance genes among bacteria of the same species, but also requires very little time to occur; therefore, plasmid-based resistance can spread quickly from one species to another. In the case of P. mirabilis, the plasmid-carrying resistance genes include ESBLs, ampC β-lactamases and carbapenemases, thus allowing for a rapid and widespread dissemination of these resistance types [220].
On plasmids, key resistant genes including blaCTX-M-65, blaCTX-M-2 and blaCMY-2 have been detected in a range of sources, including meat products and clinical samples [95]. These resistant genes frequently co-occur within certain recognized plasmid incompatibility groups (e.g., IncT, IncW, IncFIA, IncFIB and IncK) [170]. Recent field evidence powerfully illustrates this dynamic: a study of pig-derived P. mirabilis in China found that 50% of multidrug-resistant isolates carried the IncQ1α plasmid, identified by the repC gene. Whole-genome sequencing revealed these IncQ1α plasmids carried between 33 and 38 diverse resistance genes. Notably, these IncQ1α-positive isolates also co-harbored structural genes from F-type plasmids (e.g., tra operon genes), exhibiting 48 distinct structural patterns with no apparent regularity, highlighting the complex and adaptive nature of plasmid interactions in animal reservoirs [85].
In addition to the aforementioned plasmid types, some resistance genes have been observed in variations (for example, CTX-M-65) within Tn7-like composite transposons on plasmids associated with highly drug-resistant phenotypes [226]. Plasmids and ICEs are two separate means by which ARGs are generally found in P. mirabilis in comparison with the alternate method of dissemination of AMRs via vertical genetic transmission (i.e., through cell division) [95]. In addition, the original discovery of an ICE containing a significant number of ARGs was made in P. mirabilis [227]. A very large distribution of STX/R391 ICE families contribute primarily to the resistance profile of this pathogen. As the recent spread of the tmexCD-toprJ gene complex encoding the efflux pump via ICEs indicates, these elements should be closely monitored [228].
The predominant form of MGEs found in P. mirabilis is the Insertion Sequence (IS) family—the predominant ISs that are detected most frequently are members of the IS200/IS605 family. In comparison with isolates from humans, animal isolates (e.g., from chickens and dairy cattle) contain a much greater variety and a much more diverse array of MGEs, including ISPpu12, IS26, Tn7, and Tn2 in animal isolates. It seems likely, therefore, that the increased frequency of drug resistance among P. mirabilis is ascribed primarily to the presence of ISs, plasmids, and ICEs, and future research is warranted [229]. P. mirabilis exhibits a clonal propagation distribution for ICEs/ISs/plasmid replicons vs. horizontal transfer of prophages, according to the results of ancestral state reconstruction. This distribution is similar to that observed in Klebsiella pneumoniae (K. pneumoniae) [229]. Therefore, the increased prevalence of multidrug resistance in P. mirabilis is largely attributable to the dynamic interplay of IS elements, plasmids, and ICEs, warranting continued research within a One Health framework.
Integrons as Key Mediators of Antimicrobial Resistance Gene Acquisition in P. mirabilis
Integrons are specialized genetic elements that act as natural systems for collecting, integrating, and expressing gene cassettes, notably those containing antimicrobial resistance genes (ARGs). They constitute a highly efficient mechanism enabling bacteria to rapidly acquire new resistance traits [19]. Functionally, integrons enhance bacterial adaptability by capturing short, mobile gene cassettes and incorporating them at a specific integration site (attL) via a site-specific recombinase (integrase, intI), ensuring the efficient expression and dissemination of acquired genes [230,231,232]. Although integrons themselves are not directly mobile, they are frequently located on plasmids and transposons; this association allows them to assemble multiple resistance genes into a single, easily transferable unit, thereby playing a crucial role in horizontal gene transfer and bacterial evolution [19].
Integrons are commonly found in the Proteobacteria phylum and can be chromosomal or mobile [233,234]. Their importance in clinical settings stems from their ability to associate with other MGEs, such as plasmids and transposons, facilitating the widespread dissemination of resistance determinants [19]. Transposons, for instance, are MGEs that move genes between DNA molecules. They often carry single or multiple ARGs—such as tetracycline (Tn10), ampicillin (Tn3), or chloramphenicol (Tn9) resistance—and can combine with integrons to form complex genetic structures (e.g., Tn21) that further amplify resistance potential [235,236]. In P. mirabilis, the acquisition of ARGs is heavily mediated by these mobile genetic platforms. Integrons specifically assist in transferring cassettes harboring genes responsible for resistance to β-lactams, aminoglycosides, and plasmid-mediated quinolone resistance (PMQR) into recipient cells, significantly contributing to the emergence of MDR phenotypes [170].
Integrons are categorized into several classes based on integrase gene sequences, with Class 1 integrons being most strongly associated with MDR in clinical isolates [181,237]. While other classes (e.g., Class 2 and Class 3) have been identified on mobile elements, their impact appears more restricted [238,239]. These integron systems are key vectors for numerous antibiotic resistance determinants found in Enterobacteriaceae, including P. mirabilis [240]. Notably, complex integrons can serve as genetic platforms for specific plasmid-mediated beta-lactamases and PMQR genes. These structures may incorporate elements like insertion sequence common region 1 (ISCR1) and amplify resistance by duplicating the 3′-conserved segment (3′-CS), thereby expanding the variable repertoire of resistance gene cassettes located between the 5′ and 3′ conserved regions [241]. The presence and activity of integrons, therefore, are strongly correlated with elevated antibiotic resistance rates in P. mirabilis, underscoring their pivotal role in the global AMR crisis [19].
Role of Transposons and Bacteriophages in AMR Gene Mobility
Transposons, sometimes known as “jumping genes,” are mobile DNA sequences that can relocate genetic material, including ARGs, inside a bacterial genome or between various genetic units like plasmids and the chromosome. These elements usually contain integrons or individual resistance genes, which play an important role in mobilizing and distributing resistance determinants [220]. Transposases are considered the most common genes in nature [205].
Another, but more complicated, method of gene transfer is bacteriophage-mediated transduction. According to recent studies, prophage sequences incorporated into bacterial genomes can contain a considerable number of ARGs by phage transduction, a process by which bacterial viruses (bacteriophages) transfer genetic material [242]. P. mirabilis is a host for a variety of bacteriophages as it belongs to the Enterobacterales order. However, the precise function and epidemiological impact of these P. mirabilis-infecting phages in the active spread of ARGs are still unclear; available data indicates that their role may be less significant than that of plasmids or transposons in this particular pathogen [95]. Despite this possible secondary role, the presence of ARGs among prophages shows that P. mirabilis could act as a reservoir, with phages contributing to gene pool mobilization throughout microbial communities [95]. In parallel to their potential role in gene transfer, contemporary research in phage therapy highlights the therapeutic potential of phages to combat the same resistant infections. Despite the wide host range of Proteus for phages, the overall diversity of characterized Proteus-infecting phages remains remarkably low, with only 61 isolates submitted to public databases such as GenBank as of 2025, indicating critical environmental under sampling [243]. This gap limits our full understanding of their epidemiological and therapeutic roles. The newly discovered podovirus Premi, belonging to the Autographiviridae family, represents a valuable addition to this limited repertoire [244]. It is characterized by a lytic cycle and a genome devoid of virulence or toxin genes, which makes it a promising candidate for therapeutic applications, particularly against urinary tract infections [244]. However, its narrow host range—demonstrating activity against only 3 out of 30 tested clinical isolates—underscores the necessity for broader discovery efforts to develop comprehensive therapeutic cocktails [244]. Consequently, there is a pressing need for a larger collection of characterized Proteus phages to improve our capability to develop effective phage-based therapeutics [244]. The wide diversity of MGEs such as bacteriophages and transposons, among others, found in populations of P. mirabilis allows for the rapid movement and sharing of ARGs. The genetic similarity of P. mirabilis from different sources, such as chicken meat and humans with urinary tract infections, also proves that MGEs are important in allowing resistant clones to be spread between hosts and around the world [95].
5.4. Core Resistance Mechanisms
5.4.1. The Role of Efflux Pumps in Multidrug Resistance of P. mirabilis
Within P. mirabilis, the efflux pump is an integral membrane transporter that play a significant mechanism of bacterial pathogenesis, metabolism, and multidrug resistance [182,245,246]. The active removal of many types of antibiotic agents from the bacterial cell, through specialized proteins located across the plasma membrane, decreases the internal concentration of the antibiotic to sub-therapeutic levels [183]. Consequently, the efflux pump prevents the antimicrobial agent from exerting any bactericidal or bacteriostatic action, by removing the agent rapidly and maintaining the intracellular level of the agent below the minimum inhibitory concentration (MIC). As a result, efflux transporters have become promising targets for the development of new inhibitors to combat MDR-associated infectious diseases [247].
Several antibiotic families, including fluoroquinolone, tetracycline, aminoglycosides and some beta-lactams, show a link between efflux pump activity and resistance in P. mirabilis [248]. Overexpression or hyperactivation of the efflux pumps leads to a significant increase in the MDR phenotype of clinical isolates. While efflux pumps in P. mirabilis may not have been studied as extensively as beta-lactamase enzymes [220], they play a critical role in conferring resistance to antibiotics. Interestingly, some antimicrobials can function as specific substrates for these specialized efflux mechanisms, thus confirming their focused role in drug resistance [209].
These efflux systems are categorized into several major superfamilies based on their sequence homology, substrate specificity, structural features, energy sources, and efflux transporters. The most clinically significant families include the Resistance-Nodulation-Division (RND), Major Facilitator Superfamily (MFS), ATP-Binding Cassette (ABC), Small Multidrug Resistance (SMR), Proteobacterial Antimicrobial Compound Efflux (PACE), and Multidrug and Toxic Compound Extrusion (MATE) families [249,250,251].
ABC transporters play a dual role in antibiotic resistance and virulence. Export systems actively pump out antibiotics, as seen in the MacAB-TolC system. A tripartite efflux system consists of MacA (membrane fusion protein), MacB (ATP binding cassette transporter), and TolC (outer membrane channel), which contributes to macrolide resistance and virulence factor secretion in Enterobacteriaceae [252,253]. Current research aims to develop ABC transporter inhibitors as antibiotic adjuvants, though challenges remain due to the redundancy and complex regulation of bacterial efflux systems [254].
The major facilitator superfamily (MFS) is the largest and most extensively characterized group of transmembrane secondary transport proteins in both prokaryotic and eukaryotic systems [255]. This efflux mechanism reduces intracellular antibiotic concentrations, thereby preventing the drug from reaching its biological targets [256].
Functionally, SMR proteins play a critical role in the synthesis and efflux of various lipophilic compounds, including antiseptics, detergents, antibiotics, and other drugs. The SMR family encompasses a diverse group of proteins encoded by genes located on both plasmids and bacterial chromosomes, demonstrating substantial structural and functional heterogeneity. This diversity enables SMR proteins to resist different classes of antibiotics, including β-lactams such as cephalosporins. The genetic basis for this resistance is at tributed to the close association between SMR genes and antimicrobial resistance (AMR) genes within bacterial genomes, thereby enhancing the multidrug resistance capabilities of bacterial cells [257].
MATE transporters are secondary active transporters, primarily involved in the efflux of cationic substrates, and play a critical role in reducing bacterial susceptibility to a range of antimicrobial agents. These include ethidium bromide, berberine, acriflavin, norfloxacin, and tetraphenylphosphonium, all of which are substrates that can accumulate to toxic levels within bacterial cells if not extruded by these pumps [258].
First identified in 2013, PACE family transporters have been found predominantly in Proteobacteria, particularly in clinically relevant pathogens [259], transport proteins that contribute to antimicrobial resistance. The discovery of PACE family transporters has significant implications for public health and clinical practice. Their ability to confer resistance to commonly used biocides like chlorhexidine poses challenges for infection control in healthcare settings [260]. Understanding the structure and function of PACE transporters could lead to the development of novel inhibitors, potentially restoring the efficacy of certain antimicrobials [261].
Resistance Nodulation Cell Division (RND) Superfamily pumps are associated with outer membrane proteins (OMPs) and are facilitated by periplasmic adaptor proteins (PAPs) [250,252].
5.4.2. Target Site Modification as a Mechanism of Antimicrobial Resistance
General Principles of Target Site Alteration
Apart from mechanisms such as enzymatic drug inactivation and efflux pumps, P. mirabilis can develop resistance due to structural changes in the cellular targets that antibiotics are designed to block [262]. Spontaneous chromosomal mutations can affect the genes encoding these essential target proteins, which are typically involved in important processes, including cell wall synthesis and DNA replication. Such genetic changes can greatly reduce the antibacterial agent’s binding affinity and thus its therapeutic effectiveness [262]. Because the mode of action for many antibacterial agents relies heavily on very specific molecular interactions, even small structural alterations to the target site may cause large disruptions to this binding [209]. An example of how mutations can lead to fluoroquinolone resistance would be shown in the bacteria P. mirabilis where mutations commonly occur in the genes for the two main targets of fluoroquinolones which are the enzymes termed DNA gyrase (gyrA and gyrB), and topoisomerase IV (parC and parE). These 2 types of enzymes are the principal sites of action for fluoroquinolones. Therefore, when a mutation occurs in either of these genes, the structure of the enzyme at the point where the fluoroquinolone binds will change which results in an inability of the medication to bind to its target or, in turn, to neutralize the antimicrobial properties of the fluoroquinolone [179].
Strains of P. mirabilis exhibit a lower susceptibility to other antibiotic families, specifically to β-lactams [262], that demonstrates a level of target-based resistance that extends beyond fluoroquinolones. This accumulation of target site mutations results in greater antimicrobial resistance challenges presented by this opportunistic pathogen, complicating potential treatments, and encouraging the emergence of multidrug-resistant strains that are increasingly difficult to manage.
5.4.3. Enzymatic Alteration of Ribosomal Targets and Drug Inactivation
P. mirabilis can develop resistance to aminoglycoside drugs by modifying its bacterial target enzymatically. Modifications to ribosomal RNA (rRNA) or ribosomal proteins can inhibit aminoglycoside molecules from binding effectively, resulting in resistance [262]. This technique is one component of enzymatic drug inactivation, which is carried out by numerous major enzyme families. β-lactamases, aminoglycoside-modifying enzymes (AMEs), and chloramphenicol acetyltransferases are the three main categories of these inactivating enzymes. By hydrolyzing the core β-lactam ring of penicillins, cephalosporins, monobactams, and carbapenems, β-lactamases impart resistance. AMEs, on the other hand, offer broad-spectrum resistance by chemically altering the aminoglycoside drug molecule itself, which lowers the drug’s overall affinity for the target and stops it from binding to the 30S ribosomal subunit [209].
5.4.4. Reduced Outer Membrane Permeability and Porin Loss
The outer membrane of GNB, such as P. mirabilis, serves as a main defense against a variety of antimicrobial drugs. Porins, specialized outer membrane proteins that create channels filled with water, help break through this barrier. Numerous antibiotics and other hydrophilic compounds can passively diffuse into the periplasmic region through these channels. A crucial resistance mechanism is the loss or reduction in expression of these porins. This decrease in porin abundance lowers outer membrane permeability, effectively inhibiting antibiotic entrance and preventing medicines from reaching their intracellular targets [220]. This technique is especially important in cases of β-lactam resistance. For carbapenems, the loss of key porins such as ompC and ompF can result in markedly reduced susceptibility or even clearly apparent clinical resistance [220].
5.5. Epidemiology of Resistance to Key Drug Classes in P. mirabilis
5.5.1. β-Lactam and Cephalosporin Resistance: The Dominant Threat
Cephalosporins are commonly used in clinical settings to treat respiratory, urinary tract, and gastrointestinal infections. This extensive use places significant selection pressure on members of the Enterobacteriaceae family, resulting in resistance [240]. The 2019 surveillance research demonstrated this concern, showing that 8.4% of P. mirabilis isolates were resistant to a variety of medicines, including ciprofloxacin, amoxicillin, gentamicin, amoxicillin/clavulanate, and cefotaxime. Within this resistant subset, genotypic investigation revealed that 28.6% carried the ESBL gene blaCTX-M-2, whereas the majority—71.4%—had a combination of the ampC gene blaCMY-2 and the ESBL gene blaTEM-1 [263]. Despite these resistance trends, several antibiotics remain highly effective. For example, ceftazidime, piperacillin-tazobactam, and meropenem inhibited more than 98.0% of clinical P. mirabilis strains collected from US hospitals between 2011 and 2013 [264]. The detection of identical ESBL and plasmid-mediated ampC (pAmpC) genes in human clinical isolates, livestock, and retail meat underscores the interconnectedness of resistance dissemination, reinforcing the need for integrated surveillance across the One Health spectrum.
Resistance is caused by either overproduction of chromosomal ampC β-lactamases or the acquisition of transferable ESBLs [265]. Other contributing processes include variations in outer membrane porins and proteins that impair antibiotic permeability [266].
Proteus spp. Have acquired virtually the entire arsenal of β-lactamases known in the Enterobacterales order. A principal resistance mechanism in P. mirabilis is the production of these enzymes, especially ESBLs, which hydrolyze the β-lactam ring and inactivate penicillins and cephalosporins [18,180,267]. Although the chromosome of P. mirabilis does not harbor intrinsic β-lactamase genes, the bacterium can produce a wide variety of enzymes, including narrow spectrum penicillinases, ESBLs, acquired ampC cephalosporinases, and carbapenemases [170,268]. ESBLs are plasmid mediated enzymes that confer resistance to penicillins, first through third generation cephalosporins, and aztreonam, although their activity is typically inhibited by clavulanic acid. The location of ESBL genes on mobile genetic elements has facilitated their rapid spread within P. mirabilis populations and to other species, posing a major clinical challenge [19,184]. For instance, there have been two comprehensive reviews on genes for β-lactamases, dealing with their diversity, mobility and epidemiology [269,270].
The range of β-lactamases detected in P. mirabilis includes narrow spectrum enzymes (e.g., TEM, SHV, CARB, and IRT derivatives), acquired cephalosporinases (e.g., DHA, CMY, ACC-1), ESBL types (e.g., TEM/SHV, CTX-M, VEB, PER), and carbapenemases [174]. ESBL production typically results in high level resistance to ceftazidime, cefotaxime, and aztreonam [170]. The first documented case of ESBL mediated resistance in Proteus spp. Dates to 1987 [271]. A 2020 study found that 37% of strains produced ESBLs, all carrying the blaTEM gene, although these isolates remained susceptible to cefotaxime/clavulanic acid, cefoxitin, and imipenem [272]. However, recent data from high-risk reservoirs challenge this notion. A 2025 study revealed a widespread β-lactamase genotype among extensively drug-resistant P. mirabilis isolates, with the blaTEM gene detected in 97.06% of poultry and 81.82% of buffalo isolates. The ESBL gene blaCTX-M was present in 26.47% and 18.18%, and the ampC gene blaCMY-2 in 41.18% and 63.64% of isolates, respectively. Critically, these genotypic patterns were linked to a phenotypic profile of carbapenem non-susceptibility, indicating that the pervasive carriage of such genes in livestock production environments can be associated with the emergence of resistance to last-resort agents, even in the absence of traditional carbapenemase genes [79].
Class A broad spectrum β-lactamases in Proteus species include TEM-1, TEM-2, and SHV-1, which are inhibited by clavulanic acid and confer resistance to ampicillin and amoxicillin [170]. Oxacillinases (OXA) such as OXA-1, OXA-9, OXA-10, and the more recent OXA-320 (associated with integrons and the aminoglycoside resistance gene aadA1) have been reported, primarily from Turkey; these are not effectively inhibited by clavulanic acid, sulbactam, or tazobactam [273].
Early studies identified TEM-52 as the predominant ESBL conferring resistance to extended spectrum cephalosporins in P. mirabilis [274,275]. Most TEM variants are ceftazidimases, while SHV variants are less frequent than TEM and CTX-M types [276]. A shift from TEM/SHV to CTX-M variants became apparent in the early 2010s. Predominant CTX-M enzymes in P. mirabilis include CTX-M-2, -3, -14, and -27 [170,277]. CTX-M-8 is rare, having been found only in Brazil, while CTX-M-65 has been identified in Russia [226]. The blaCTX-M-15 gene is often preceded by ISEcp1, which mobilizes the gene and elevates its expression, leading to high level cephalosporin resistance [278]. CTX-M-14 is the most widely distributed variant in Proteus spp., first reported in South Korea [178] and later in China alongside its reduced activity derivative CTX-M-140 [279]. Notably, the presence of ESBL genes does not always translate to a phenotypic ESBL profile [280]. Initially, blaCTX-M often appears together with blaTEM, but it tends to become the dominant gene in a population over time [240].
5.5.2. Fluoroquinolone Resistance in P. mirabilis
Fluoroquinolones are one of the most common classes of antibiotics that are used to treat infections in many countries throughout Western Europe, North America, and Japan. The spectrum of treatment with fluoroquinolones is extensive and includes infections such as UTIs [281]. Resistance in the bacterium P. mirabilis occurs via multiple mechanisms. The mechanism for high level resistance to fluoroquinolone antibiotics stems primarily from chromosomal alterations in the genes that encode the target enzyme, DNA gyrase (gyrA and gyrB) and topoisomerase IV (parC and parE) [282]. A recent study found that gyrA mutations were not significantly present in their sample groups [79], providing evidence that gyrA mutations may not be an important method for selecting for resistance in both broiler chickens and buffalo bacteria. Specific mutations within the conserved quinolone resistance-determining regions (QRDRs), such as S80 in parC and S83 in gyrA, are strongly linked to resistance, with a mutation at S464 in gyrB leading to an even higher resistance level [283]. Other contributing chromosomal mechanisms include alterations in the outer membrane and the action of efflux pumps, which reduce intracellular drug accumulation [284].
In addition to chromosomal mutations, plasmid-mediated mechanisms play a significant role. These include (i) protection of drug targets by qnr proteins (e.g., qnrA, qnrD, qnrS), (ii) drug modification by enzymes like AAC(6′)-Ib-cr, and (iii) active drug efflux by specific pumps such as qepA or oqxAB [285]. Data from Hasona et al. [79] revealed a variable prevalence of these plasmid-mediated genes in Egyptian isolates: the qnrA gene was detected in 47.06% of broiler chicken isolates but only 9.09% of buffalo isolates, while the qnrS gene was not found (0% in both sample types). These resistance genes are often located near β-lactamase genes (e.g., blaTEM, blaCTX-M) on mobile genetic elements like class 1 and class 2 integrons [286]. While the qnrA6 gene has been found to be chromosomally encoded in P. mirabilis [287], plasmid-borne qnr genes have achieved a global distribution. Proteus spp. Carrying plasmids with the qnrD gene have been reported in numerous countries [284,285,288]. The qnrC gene has been found on a plasmid also carrying resistance genes for ampicillin, sulfamethoxazole, and trimethoprim [289]. The qnrA and qnrS genes are reported less frequently [290,291]. From a One Health perspective, qnrA and qnrD genes have been detected in isolates from companion animals, while only qnrD has been identified in isolates from food products [58,59,82,185,204].
5.5.3. Tetracycline Resistance in P. mirabilis
Tetracycline antibiotics are used to treat several GNB infections; however, high rates of resistance to these drugs have been documented within the Enterobacteriaceae family [292]. This resistance is primarily attributed to an efflux mechanism, where the antibiotic is actively pumped out of the bacterial cell. The genes responsible for this efflux are frequently located on mobile genetic elements; for instance, the class A tetracycline resistance (tet) determinant was the first to be identified from the RP1/Tn1721 transposon system [293]. In Enterobacteriaceae, tetracycline resistance is most commonly associated with the tet(A) to tet(E) gene determinants [294].
P. mirabilis exhibits a natural, intrinsic resistance to classical tetracycline, which is a key factor in its increasing tolerance to this drug class [295]. Tigecycline, a 9-t-butylglycylamido derivative of minocycline, is a newer tetracycline-class antibiotic developed to treat resistant GNB [296]. While the classical tetracycline resistance determinants do not affect tigecycline, the identification of the acrAB efflux pump in P. mirabilis provides an explanation for its reduced susceptibility to this last-resort drug.
A variety of tetracycline resistance genes, including tet(O), catI, tet(J), tet(A48), tet(A), tet(B), tet(C), and tet(M), have been detected in isolates from farm animals, companion animals, and food products, though they have not been found in isolates from wild animals [34,59,78,82,204]. Supporting this, a recent study by Hasona et al. [79] reported a high prevalence of the tet(M) gene in P. mirabilis from Egyptian animals, found in 81.82% of broiler chicken isolates and 70.59% of buffalo isolates. The same study detected the macrolide resistance gene ermB in 23.53% of broiler isolates and 18.18% of buffalo isolates, indicating the co-circulation of resistance genes to multiple antibiotic classes within these populations. Supporting the global prevalence of tetracycline resistance mechanisms, a recent whole-genome sequencing study of 18 Indian P. mirabilis strains reported universal phenotypic resistance to tetracycline. Genomic analysis revealed that all but one isolate carried the intrinsic resistance gene tet(J), alongside other common determinants such as tet(D) and amphenicol resistance genes (cat, catA4). These genes were frequently embedded within MGEs, underscoring the role of horizontal gene transfer in their maintenance and spread within this regional population [297].
5.5.4. Aminoglycoside Resistance in P. mirabilis
Aminoglycosides are classified as broad-spectrum antibiotics; they are primarily produced from Actinomyces species and are indicated for use against both Gram-negative and Gram-positive pathogens [298]. While effective, the general use of aminoglycosides has been severely limited due to bacterial resistance and toxicity to humans and animals [299]. However, they remain one of the most available means of treating infections caused by MDR organisms [298]. The most frequently observed mechanism of aminoglycoside resistance involves enzymatic modification of the drug by AMEs. The second significant mechanism for aminoglycoside resistance is via 16S rRNA methylation, a process that gives rise to a high level of resistance to gentamicin, tobramycin and amikacin [300]. AMEs confer aminoglycoside resistance through the introduction of chemical modifications to the aminoglycoside molecule, and these enzymes are categorized by the type of modification they introduce acetyltransferases (AACs), phosphotransferases (APHs), nucleotidyltransferases, or adenyltransferases (ANTs) [301]. The genes encoding these enzymes are typically located on MGE, e.g., plasmids, integrons, transposons), and most often they co-occur with other genes that confer antimicrobial resistance; this facilitates their co-dissemination [298].
In P. mirabilis, the genetic determinants of aminoglycoside resistance most often occur in the form of gene cassettes located in integrons. Frequently identified genes include aadA1 and aadA2 (aminoglycoside adenyltransferases), aadB (aminoglycoside (2″) adenyltransferase), aac(6′)-Ib (also known as aacA4, an acetyltransferase), and sat2 (streptothricin acetyltransferase) [291]. A recent study by Hasona et al. [79] underscores the high prevalence of one such gene, reporting aadA1 in 97.06% of broiler chicken isolates and 100% of buffalo isolates. Furthermore, several variants of 16S rRNA methyltransferases (such as rmtA, B, C, D, and armA) have been identified in Enterobacterales, including P. mirabilis [302]. These enzymes modify the aminoglycoside binding site on the ribosome, conferring high-level resistance to virtually all clinically available aminoglycosides [303]. Other potential resistance strategies include alteration of membrane proteins or the ribosome and increased drug efflux, but these are not as widespread as AME-mediated resistance [304].
From an epidemiological perspective, the distribution of specific resistance genes varies by source. Isolates from farm animals have been found to carry aac(6′)-Ib-cr, aph(3′)-IIa, rmtB, aacC1, and aacC2 [30]. In companion animals, detected genes include aphAI-IAB, *aac(3′)-*IV, aac(6′)-Ib, and aadA1 [58,59,204]. Meanwhile, foodborne isolates have been shown to harbor aac(6′)-Ib-cr, aph(4)-Ia, aadA1, aadA2, aac(3′)-Ia, *aac(3)-*IV, and aac(3)-Iva [79,82,185,305].
5.6. Co-Carriage of Multidrug Resistance Genes: The Convergence of Threats
Antibiotic resistance in P. mirabilis strains from animals and food is a major public health concern. The threat of this pathogen is exacerbated not only by the commonality of ESBL genes but by their frequent co-existence with a diverse array of other ARGs. These co-carried genes include those conferring resistance to tetracycline (tet(M)), sulfonamide (sul2), macrolide (ermB), quinolone (qnrA), chloramphenicol (catA1), aminoglycoside (aadA1), trimethoprim (dfrA1), and colistin (mcr-1). The descriptions surrounding this co-carried profile have been documented in different reservoirs across many models [79].
Isolated strains from arm-Animal species; swine, avian, and duck species, contained numerous antimicrobial resistance genes, including norA, acrB, blaOXA, blaTEM, blaCTX-M, blaNDM, blaDHA, and blaKPC [32,52,57,78]. Isolated strains from pets; dog and cat, had blaOXA-1, blaTEM, blaCTX-M, and blaDHA [58,59,204] while those from foods had blaCTX, blaOXA, blaDHA, blaCMY-2, blaNDM, blaTEM, blaSHV, blaFOX, blaCIT, blaEBC, and blaMBL [52,82,185]. In addition, farm animal isolates also carry quinolone resistance genes, including qnrS, parC, qnrD, and oqxA [306].
This multidrug-resistant nature is exemplified by strains like XH983, which carries ARGs for aminoglycosides (aph(3′)-Ia, aph(3″)-Ib, aph(6)-Id, aac(3)-IId, aadA5, aadA1), β-lactams (blaKPC-2, blaTEM-1B), phenicols (cat, catA1), sulfonamides/trimethoprim (drfA1, drfA17, sul1, sul2), and tetracycline (*tet(*J)) [307]. The emerging genes include blaCTX-M-65, identified in a Russian isolate with blaVEB, aac6-Ib, and qnrA1 [224] and show an increase in numbers of different resistance mechanisms for aminoglycosides (aacA4, aadB, aphA6), β-lactams with expanded spectrum (blaVEB-6), and carbapenem-resistant strains (blaNDM-1) [308].
ESBL production is strongly associated with fluoroquinolone resistance in clinical settings. This linkage is not merely epidemiological but is mechanistically explained by the frequent co-localization of their respective genetic determinants on the same mobile genetic elements. Specifically, plasmids harboring ESBL genes (especially blaCTX-M variants) often also carry PMQR genes, such as qnr alleles (e.g., qnrA, qnrD, qnrS) and aac(6′)-Ib-cr, alongside determinants for sulfonamides, aminoglycosides, and tetracyclines [170,287]. Alabi This genetic linkage on conjugative plasmids (e.g., IncF, IncI types) facilitates the co-transfer and co-selection of multidrug resistance profiles during horizontal gene transfer events. Consequently, exposure to a single antibiotic class (e.g., β-lactams) can select for and maintain resistance to other unrelated classes (e.g., fluoroquinolones), severely limiting therapeutic options. These include aminoglycoside resistance genes (aad1, aad2, aac3Ia, aac6-Ib, aph(3)-Ia, aph(6)-Id), as well as genes for sulfonamide (sul1, sul2), trimethoprim (dfrA1, drfA32), chloramphenicol (cat, cat1), tetracycline (tet), and macrolide (msr) resistance, particularly alongside CTX-M-type ESBLs [170]. Studies have shown that a very high level of co-production of ESBLs with ampC and carbapenemase enzymes exists in India, as reported by Datta et al. [309]. The increasing trend of co-producing ampC with ESBLs raises the concern of increasing the area of β-lactam resistant. A study in India identified that co-production of an ampC enzyme among 19.4% of the ESBL-producer sample was noted [220].
Current monitoring studies demonstrate the vast prevalence of the co-production of these genes, especially concerning key livestock reservoirs in Egypt. Among sets of broiler chicken isolates, there were high levels of prevalence with int1 (97.06%), dfrA1 (100%), sul2 (97.06%), aadA1 (97.06%), tet(M) (81.82%), qnrA (47.06%), and mcr-1 (11.76%). A similar presence was documented for the β-lactamase gene, with blaTEM being nearly ubiquitous (97.06%) while blaCTX-M, blaOXA-10, and blaCMY-2 had respective detection frequencies of 26.47%, 2.94%, and 41.18%. The buffalo isolates also demonstrated similar patterns of prevalence for int1, dfrA1, and aadA1 (all 100%), sul2 (90.91%), blaTEM (81.82%), and blaCMY-2 (63.64%). In summary, 97.06% of the broiler isolates and 81.82% of the buffalo isolates contained β-lactamase genes [79]. The ability to produce this broad, extensive, and genetically linked resistance profile supports the assertion of P. mirabilis’s complex, co-selected nature as a multidrug resistant organism. The conjunction of β-lactam resistance with determinants for the other antimicrobial classifications creates an extremely limited number of therapeutic options for treating patients with these infections, making it imperative that all sectors of animal, food, and human health begin to develop an integrated surveillance and antimicrobial stewardship infrastructure to combat this situation. Most importantly, we must adopt a wider evolutionary and ecological viewpoint of the issue [310]. When reviewing the molecular epidemiology of AMR in P. mirabilis, it is clear that this pathogen is exceptionally well-adapted. It can continually acquire and disseminate many resistance genes through its extensive mobile genome (mobilome). The presence of strong genetic evidence indicating transmission from animal to human through food certainly depicts a severe One Health crisis (Table 3). The overall impressive transition of this pathogen from a traditional opportunistic pathogen to a means of spreading pan-resistant infections beyond recognized therapeutic limits will represent a healthcare crisis for both humankind and the animal kingdom. The widespread distribution of identical resistance genes across human, animal, and food reservoirs, as detailed above, makes it clear that isolated interventions in any single sector are insufficient. The following section outlines integrated, cross-sectorial strategies essential for containing the spread of resistant P. mirabilis.
6. The One Health Approach to Combat P. mirabilis AMR: Collaborative Strategies
According to WHO [311] and Franklin et al. [312], the One Health approach is a cooperative, multidisciplinary method that aims to achieve optimal health outcomes for people, animals, and ecosystems while acknowledging their interconnection (Figure 4). Given that the same antimicrobial medications are used in human medicine, veterinary care, and agriculture and that both humans and animals can carry and spread the same resistant pathogens, this framework has been widely adopted to address the complex problem of AMR [311]. A One Health approach to AMR recognizes the need for an all-encompassing system that incorporates plant, animal, and human health and takes into account the environment’s crucial role in promoting the spread of bacteria and resistance genes [311]. Understanding the fundamentals of evolutionary biology and how they relate to prokaryotes is essential for long-term management of infectious diseases [313,314]. An understanding of this complex issue is reflected in the global strategy for AMR surveillance, which is in line with the One Health concept [315,316]. The shift to integrated One Health surveillance systems is presently taking place in a number of different parts of the world.
Addressing AMR effectively requires insights from multiple disciplines, framing it squarely within the One Health approach [317,318,319]. Central to this approach is antibiotic stewardship, which is recognized as vital for directly reducing the selection pressure that drives resistance [320,321]. The implementation of One Health principles is therefore pivotal in combating antibiotic resistance effectively [322]. This approach underscores the interdependence of human, animal, and environmental health and emphasizes the importance of collaboration across scientific, social, and political sectors [323] to effectively monitor the emergence and movement of resistance genes and resistant bacteria within and between these different compartments. A major driver of AMR within this context is the overuse and misuse of antimicrobials in animal agriculture, a practice intimately connected to bacterial zoonosis that has significantly accelerated the emergence of resistance [95].
One Health is characterized by an integrated, unifying effort that operates at local, national, and global levels to achieve the best possible health for people, animals, and our environment [324]. Placing AMR on the international political agenda and tackling it through a One Health lens is crucial for driving meaningful change. In a key step, the WHO adopted the Global Action Plan on AMR in 2015, urging countries to establish national surveillance systems for bacteria from both humans and animals and explicitly advocating for a One Health strategy [325]. AMR is a global health issue that perfectly exemplifies the need for a One Health approach. Because bacteria and their mobile genetic elements readily cross species and environmental boundaries, it is critical to understand the connections between the microbiota of humans, animals, and ecosystems—the core of the One Health concept [326,327]. This interconnectedness demands a unified approach that acknowledges the inextricable links between human, animal, and environmental health [322].
National and international bodies have embraced this framework. The U.S. National Action Plan for Combating Antibiotic-Resistant Bacteria (2020–2025) explicitly includes the One Health approach as an effective strategy [328]. A primary objective is to promote antibiotic stewardship in animals, eliminate the use of medically important antibiotics for growth promotion, and bring all other uses of these drugs under veterinary oversight [323]. Globally, the quadripartite organizations—the Food and Agriculture Organization (FAO), the World Organization for Animal Health (OIE), the WHO, and the United Nations Environment Program (UNEP)—have established a joint One Health action plan and a strategic framework for collaborative work on AMR [329]. Assessing the true extent of AMR in LMICs remains challenging due to inadequate surveillance, though initiatives like the WHO’s Global Antimicrobial Resistance and Use Surveillance System (GLASS), launched in 2015, are important steps forward, to fill knowledge gaps and guide strategies at all levels. GLASS was created to progressively integrate surveillance data on antimicrobials used in humans, track antimicrobial use, and understand the role of AMR in the food chain and the environment [330].
Other major strategies highlighting the importance of a multisectoral One Health response include the United States National Action Plan for combating antibiotic-resistant bacteria [331] and the WHO Global Action Plan [325], the 2016 United Nations General Assembly Declaration on AMR [332], and the FAO/OIE/WHO Tripartite Collaboration [333,334]. The threat of AMR is profound, with the potential to hinder infectious disease control, increase healthcare costs, and reverse decades of progress in medicine [335]. Through its comprehensive and collaborative framework, the One Health approach provides a powerful and necessary means to limit the spread of AMR and safeguard modern medicine from the threat of a post-antibiotic era.
6.1. Integrated Surveillance and Monitoring Programs for Antimicrobial Resistance in P. mirabilis
Implementing continuous surveillance programs to monitor antimicrobial resistance patterns in P. mirabilis is crucial for developing effective treatment guidelines and shaping public health policies [33,336]. These surveillance systems collect and analyze data on resistance trends and antimicrobial consumption from the human health, animal health, and environmental sectors, thereby offering a holistic One Health perspective on the AMR situation.
Conducting extensive monitoring of resistance trends in diverse geographical areas and various healthcare environments yields critical intelligence on the dynamically changing resistance profile of this pathogen [18,297]. The use of standardized protocols for data collection, coupled with advanced molecular techniques like whole-genome sequencing, is instrumental for identifying the origins of resistance genes and tracking the dissemination of resistant clones. The critical need for integrated, genomics-driven surveillance is powerfully illustrated by recent studies. Global genomic analyses have mapped the phylogeographic spread of resistant clones and quantified the carriage of ARGs and MGEs across continents [95]. Concurrently, national-level sequencing efforts, such as those analyzing strains from India, provide granular insights into local resistance patterns and virulence gene repertoires, demonstrating the utility of WGS in outbreak investigation and resistance tracking [297]. These studies collectively advocate for the establishment of P. mirabilis as a key sentinel organism for AMR surveillance within the One Health paradigm. Fostering collaboration and sharing data across different sectors enriches the overall understanding of AMR epidemiology and facilitates a swift, coordinated response to emerging threats. Simultaneously, monitoring how antimicrobials are used helps identify areas of misuse and provides a solid foundation for stewardship programs.
The evidence generated by this integrated surveillance is pivotal; it enables healthcare institutions and public health authorities to design and implement focused antimicrobial stewardship initiatives and robust infection control practices to curb the spread of resistant P. mirabilis strains [181,337]. In summary, a unified, cross-sectoral surveillance system produces the essential evidence needed to direct precise interventions and bolster the global effort against AMR.
6.2. Antimicrobial Stewardship: Promoting Responsible Use in Human and Veterinary Medicine
Antimicrobial stewardship is a systematic approach designed to encourage appropriate use of antimicrobials, which is one of the key components in addressing antimicrobial resistance for P. mirabilis. This holistic method includes unique but interrelated actions in both the veterinary and human healthcare settings. In the area of human healthcare, comprehensive stewardship programs emphasize appropriate prescribing based on scientific evidence, use of accurate and prompt diagnostic tools to promote targeted therapy rather than empirical therapy, and educate patients on appropriate use of antibiotics [330]. In the veterinary agrarian sectors, antimicrobial stewardship mandates that all forms of antibiotic applications be under the direction of qualified health professionals. In this case, there is a ban against using antibiotics for the enhancement of growth, instead promoting alternative prophylactic measures such as vaccinations and better husbandry practices; and strictly adhering to through prudent-use guidelines for using antibiotics, including bans on antibiotics that are classified as critically important for treating humans [3].
It is essential that stewardship principles are consistently and coordinately applied in all sectors of the world. Reducing the total volume and inappropriate use of antimicrobials decreases the selective pressure for the development of resistance. Decreasing the overall volume of antimicrobials and limiting inappropriate use reduces the risk for development and transmission of resistant P. mirabilis strains and is critical for ensuring the continued effectiveness of current antimicrobials in the treatment of infections caused by P. mirabilis for decades to come. The partnership plans to develop and implement five new treatments for drug resistant bacteria, identified by WHO as the highest priority for action by 2025 [325].
6.3. Integrated Surveillance and Cross-Sectoral Infection Control Strategies
Employing a robust surveillance system wherein data on P. mirabilis resistance patterns is collected, analyzed systematically and disseminated to provide clinicians with essential information that will aid in administering optimal/effective treatment approaches [263,338]. Additionally, implementing strict biosecurity/infection control measures to contain the spread of antimicrobial resistant P. mirabilis between humans and animals and the environment. In health care settings, this includes adhering to sound hand hygiene practices, ensuring thorough cleaning/disinfection of the environment, isolating infected/colonized patients when indicated, and conducting proper management of urinary catheters to prevent the spread of healthcare acquired infections. In agriculture, it involves ensuring good sanitary conditions on farms, controlling movement of livestock, implementing proper manure/waste management systems and measures to control the spread of resistant bacteria. In addition to the above, it is important to consider safe food handling, improving wastewater treatment to eliminate biological contaminants, and monitoring the quality of water regularly at the community/environmental levels. In order to achieve the goal of interrupting/transmitting resistant P. mirabilis through all sectors by implementing a multi-faceted approach to prevention/control, it would help greatly in the global fight against antimicrobial resistance.
6.4. Innovative Therapeutic Strategies: Development of New Antimicrobials and Alternative Therapies
Antimicrobial stewardship and infection control are essential components in the fight against P. mirabilis AMR, but they will only be successful if new and innovative antimicrobial agents and treatment options that can help manage infections caused by this pathogen are developed [184,339]. The increased prevalence of AMR is largely due to the presence of intrinsic factors, as well as the acquisition of resistance genes from other organisms via horizontal gene transfer. As such, it is critical that an extensive and diverse assortment of drugs is available to effectively deal with the growing incidence of AMR infections [340,341].
Given the history of limited antibiotic development, the emergence of new resistance mechanisms is a major concern. Therefore, there is an urgent need for the identification and clinical testing of new antibiotics containing unique mechanisms of action that can be used to treat multidrug-resistant P. mirabilis [182]. The study of potential antibiotic adjuvants, including β-lactamase inhibitors and efflux pump inhibitors, to assist in restoring the efficacy of current antibiotics against resistant strains of P. mirabilis is an exciting area of research [183,342]. Beyond this line of research, continued exploration of new and alternative sources, as well as new classes of antimicrobial compounds, will provide additional support in overcoming existing resistance. Such innovative approaches not only focus on the destruction of bacteria cells but also on rendering them ineffective, less selective for resistance development, and exploring unconventional mechanisms. Alternative methods to the conventional usage of antibiotics have been explored for their potential in the improvement of treatment outcomes by developing innovative treatment modalities. The major areas of focus for this purpose are as follows:
6.5. Alternative Mechanisms to Treat Resistant Bacteria
The development of alternative mechanisms represents a critical frontier in overcoming antimicrobial resistance. Various alternative therapies, including vaccines, antibodies, pattern recognition receptors, probiotics, bacteriophages, antimicrobial peptides (amps), phytochemicals, metals, and antimicrobial enzymes, all offer approaches to combat microbial Infections [343]. Key strategies include:
6.5.1. Fecal Microbiota Transplantation (FMT)
Antibiotic-mediated disruption of the colonic flora compromises colonization resistance, a key host defense. FMT corrects this dysbiosis by reintroducing a healthy donor microbiota, thereby restoring ecological balance and intestinal function [344].
6.5.2. Fusion Protein Technology
This technology addresses significant challenges in the recombinant expression and stability of Bacteriologically Active Antimicrobial Peptides (BAMPs). The approach is successfully utilized in transgenic expression systems to produce functional peptides. According to Statistical data from the Antimicrobial Peptide Database 3 (APD3, https://aps.unmc.edu/) (accessed on 10 September 2025), there are 3940 known AMPs from the six kingdoms of life. This includes 383 bacteriocins from bacteria; 5 from archaea; 8 from protists, 29 from fungi; 250 from plants; and 2463 from mammals, including some Synthetic peptides [345]. AMPs from a range of species, including amphibians, insects, mammals, and fish, account for 75.65% Of the total number of AMPs. 13.5% of AMPs originate from plants, and 8.53% come from bacteria [346]. AMPs offer a promising avenue for combating infectious diseases and ameliorating the impacts of AMR [347]. A significant number of AMPs have demonstrated antibacterial properties against specific pathogenic microbes [348], through either membrane or non-membrane mediated mechanisms, these peptides, encompassing both Gram-positive and Gram-negative types, have demonstrated robust antibacterial activity against a broad range of both Gram-positive and Gram-negative bacteria [345].
Importantly, the antibacterial property of AMPs is thermostable, and the producer strain Offers a certain degree of self-protection against its own antibacterial peptide [349]. The mechanism of AMPs is to attack bacterial membranes by forming pores in the outer membrane [345]. Additionally, some AMPs kill pathogens by inhibiting intracellular pathways such as cell division [350], DNA migration [351], RNA polymerase [352], cell envelope [353], or cell wall biosynthesis [354,355]. Additionally, certain AMPs are effective against some membrane-enveloped viruses that can disrupt specific membranes in the host. This dual function is thought to arise due to the interdependent relationship between viruses and hosts, as viruses rely on the host replication machinery. Consequently, AMPs may offer the beneficial side effect of protecting against certain viral infections [356]. Additionally, certain AMPs are effective against some membrane-enveloped viruses that can disrupt specific membranes in the host. This dual function is thought to arise due to the interdependent relationship between viruses and hosts, as viruses rely on the host replication machinery. Consequently, AMPs may offer the beneficial side effect of protecting against certain viral infections [356]. Due to their broad-spectrum activity and low likelihood of inducing resistance, ABPs have emerged as promising alternatives to conventional Antibiotics in the fight against drug-resistant pathogens.
6.5.3. CRISPR-Cas Systems
CRISPR-Cas systems, which originated as a bacterial adaptive immune mechanism, have emerged as a powerful and precise tool in addressing the global challenge of AMR. The versatility of this system is further exemplified by the expanding arsenal of Cas proteins [357] including Cas9, Cas12, Cas13, and Cas14 each offering distinct targeting capabilities (e.g., DNA or RNA) sequences, enabling versatile strategies to combat resistant pathogens [358]. These programmable systems are being developed as next-generation antimicrobials due to their ability to target bacterial DNA or antibiotic resistance genes with sequence-specific accuracy. This strategy represents a novel approach that bypasses traditional resistance mechanisms and enhances therapeutic specificity [359,360]. Furthermore, they offer a transformative pathway to combat AMR, with potential applications extending beyond conventional strategies. Beyond this application, the CRISPR-Cas9 platform has facilitated substantial advancements across medicine, enabling innovative therapeutic strategies for a range of intricate ailments including genetic disorders and malignancies, while also serving as a powerful diagnostic and therapeutic tool in microbiology [357]. Integrating these technologies with existing methods could significantly improve the management and potential reversal of the growing AMR crisis. Ongoing research continues to evaluate their effectiveness, address delivery and specificity challenges, and facilitate clinical translation [361,362]. Future progress is contingent upon overcoming technical and ethical barriers to enable safe and effective application. Key specific concerns that necessitate meticulous deliberation include off-target effects, poor delivery efficiency to target tissues, and potential unwanted side effects [357]. Therefore, underscoring the necessity for interdisciplinary collaboration and international cooperation to fully harness their potential against drug-resistant pathogens, more rigorous preclinical and clinical experiments are essential before broad human application [357].
Building on precision genetic tools, micro-/nanomotors (MNMs) are autonomously moving particles that actively penetrate biofilms. This capability enables localized antimicrobial delivery and physical biofilm disruption. A promising forward-looking strategy involves merging this active delivery with CRISPR precision by loading CRISPR payloads onto functionalized MNMs. Such a hybrid system could navigate biofilms to deliver genetic cargo directly to bacterial cells, potentially overcoming the critical delivery barriers that limit standalone CRISPR therapies [363].
6.5.4. Antimicrobial Peptides (AMPs)
Antimicrobial peptides (AMPs), naturally occurring molecules derived from animal and microbial sources, represent a promising new class of antibiotics that primarily exert their effect by targeting the bacterial cell membrane [298]. These peptide-based antimicrobials have garnered significant interest due to their unique amphipathic structure, which is conferred by a combination of positively charged amino acids (such as lysine, arginine, and histidine) and a high proportion (often 50%) of hydrophobic residues [301,302]. Typically comprising fewer than 100 amino acids, this structure enables potent interaction with and disruption of microbial membranes.
A key advantage of many AMPs is their capacity for multi-mechanistic action, which enhances their overall antimicrobial efficacy and can reduce the potential for resistance development. For instance, the human cathelicidin LL-37 demonstrates not only direct bactericidal activity but also immune-modulatory and anti-biofilm properties [364]. Aires Host-derived AMPs such as lipocalin, hepcidin, and LL-37—isolated from sources like human bone marrow and umbilical cord mesenchymal stem cells (MSCs)—exhibit potent, broad-spectrum antibacterial properties [365].
However, some therapeutic challenges remain. Certain peptides have been associated with notable nephrotoxicity, primarily linked to the high doses required for clinical efficacy [210]. Furthermore, other significant limitations hinder their clinical translation, including susceptibility to proteolytic degradation, potential host cytotoxicity, and high production costs. Several peptide-based antibacterials are successfully used in medicine. Commercially available examples include dalbavancin, daptomycin, colistin (polymyxin E), oritavancin, polymyxin B, teicoplanin, vancomycin, and telavancin [366].
6.5.5. Efflux Pump Inhibitor (EPIs)
Another strategic approach to combat antimicrobial resistance is the utilization of Efflux Pump Inhibitors (EPIs). These compounds function by obstructing the activity of bacterial efflux pumps, which are key mediators of multidrug resistance. When integrated into combination therapies with existing antimicrobials, EPIs can significantly restore and enhance drug efficacy [367].
EPIs are generally characterized as simple, stable, and affordable molecules with a favorable safety profile for human use [368]. A notable secondary benefit of certain EPIs is their additional capability to inhibit bacterial biofilm formation, thereby targeting another major resistance and virulence mechanism. Clinically relevant examples of EPI compounds include thioridazine, phenylalanine-arginine β-naphthylamide (PaβN), and the arylpiperazine derivative 1-(1-naphthylmethyl)-piperazine (NMP).
6.5.6. Bacteriophage (Phage) Therapy
Bacteriophage (phage) therapy involves the use of viruses that specifically infect and lyse bacterial hosts, representing a promising alternative for combating MDR pathogens. Often described as “bacteria eaters,” phages offer several distinct advantages over conventional antibiotics, including high host specificity, natural self-replication at the infection site, low inherent toxicity, minimal environmental impact, and the absence of cross-resistance with antibiotics. Their diversity and relative ease of isolation contribute to their therapeutic potential.
Clinically, phage therapy has demonstrated notable efficacy in treating localized and topical infections where conventional antibiotics have failed. Promisingly, combination strategies employing both phages and antibiotics have shown synergistic effects, particularly in eradicating challenging multidrug-resistant biofilms [369].
Engineered lysins and bacteriophage therapy: Modified lysins with enhanced therapeutic profiles and carefully selected bacteriophages offer highly targeted and potent alternatives to conventional antibiotics. These biological agents show significant promise against multidrug-resistant bacteria, including specific resistant strains [370,371]. The high specificity of phages can restrict their broad applicability. Additionally, bacteria may evolve resistance to phage infection. The clinical translation also faces complex regulatory and manufacturing hurdles.
6.5.7. Immunotherapy
Immunotherapy encompasses therapeutic strategies designed to enhance, modulate, or harness the body’s immune system to combat infections more effectively (e.g., immunotherapy, procurement of lymphoid tissue stem cells). By leveraging the inherent specificity and adaptability of the immune response, these strategies offer a promising avenue for addressing antibiotic-resistant infections, potentially overcoming key limitations of traditional antibiotics. These include the risk of triggering autoimmune reactions, variability in patient responses, and the typically lengthy and complex development and regulatory pathways.
6.5.8. Antivirulence Therapy
Antivirulence therapy represents a novel therapeutic approach that focuses on suppressing or eradicating the virulence factors secreted by a pathogen, rather than directly killing the bacterium itself. The primary intention is to disarm the pathogen and mitigate infection severity while reducing the selective pressure that drives the development of traditional antimicrobial resistance. However, this strategy presents certain challenges. It may not achieve complete pathogen eradication, necessitating a precise and thorough understanding of intricate virulence mechanisms. Additionally, there remains the potential for pathogens to evolve compensatory virulence pathways. Another innovative method combines antibacterial sonodynamic therapy with antivirulence immunotherapy by using engineered nanovesicles to capture bacterial toxins and generate reactive oxygen species upon ultrasound activation [372].
6.5.9. Probiotic and Prebiotic Supplementation
This strategy involves modulating the host microbiome to enhance its ability to manage and prevent pathogen colonization, thereby competitively excluding potential invaders. This strategy shows particular promise in managing infections caused by Proteus mirabilis. Specific strains of Lactobacillus, particularly Lactobacillus gasseri (L. gasseri), have demonstrated the ability to help prevent P. mirabilis-induced urinary stones. The primary mechanism involves the production of lactic acid, which directly inhibits the growth of P. mirabilis and critically blocks its urease enzyme activity. Lactic acid competes with urea for binding to urease, thereby reducing the production of ammonia and subsequent crystallization. This results in diminished biofilm formation and fewer urinary stones [373]. The efficacy of this approach can be constrained by its strain-specific nature, variable colonization success in the host, and potentially limited impact against severe, established infections.
6.5.10. Drug Repositioning
Drug repositioning is a strategy that involves identifying new antimicrobial applications for existing drugs already approved for other clinical indications. This approach, which includes combining non-antibiotic drugs with traditional antibiotics to enhance their effectiveness, offers a pathway to accelerate the development of new anti-infective therapies. A key advantage is the potential to achieve therapeutic effects with lower doses of the antibacterial agent, thereby minimizing adverse impacts on healthy cells and tissues [374]. In addition, reactive nitric oxide (NO) and reactive oxygen species (ROS) produced by metallic NPs harm bacterial cell components [375]. Repositioned agents often exhibit more modest antimicrobial potency compared to purpose-developed antibiotics, necessitating extensive clinical validation for their new indication. Furthermore, the potential for unexpected off-target side effects remains a significant consideration during development.
6.5.11. Machine Learning and Artificial Intelligence for Predicting Antimicrobial Resistance
The application of machine learning (ML) and artificial intelligence (AI) offers a transformative, data-driven alternative for combating antimicrobial resistance by enabling early prediction and informed decision-making, thereby optimizing therapy and stewardship. A pivotal challenge in managing bloodstream infections is the 24–72 h delay in obtaining culture and susceptibility results [376], a period during which clinicians often rely on empirical broad-spectrum antibiotics that can inadvertently exacerbate AMR if inappropriate [377]. Developing tools to guide more targeted treatment during this window is essential for improving outcomes and curbing resistance [378]. Toward this goal, researchers have developed supervised ML models to predict the presence of resistant organisms in blood cultures from data available at the patient’s first encounter. Utilizing algorithms such as penalized logistic regression, random forest, and XGBoost to classify key resistant pathogens (including ESBL, CRE, ampC, MRSA, and VRE), studies have demonstrated high predictive performance [379]. This approach can reduce unnecessary broad-spectrum use and associated costs by providing early, evidence-based guidance.
The promise of ML extends to other infections, such as UTIs, where predictive tools using electronic health record data have shown potential for forecasting bacterial growth and resistance patterns before culture results are available [380]. Furthermore, recent advancements include integrating socio-economic deprivation indices into AMR modeling, combining demographics, infection history, and comorbidities to offer real-time decision support for infections like sepsis [380]. Beyond clinical predictors, AI and ML are revolutionizing the analysis of genomic data for AMR. AI-driven tools can automate the identification of resistance genes from sequencing data, predict phenotypic resistance patterns, and help optimize personalized treatment strategies, significantly accelerating the interpretation of complex genomic datasets [381]. This represents a powerful future direction for resistance surveillance and precision infectious disease medicine.
6.5.12. Plant-Derived Essential Oils (Eos)
Despite the promising therapeutic potential of plant-derived essential oils (Eos) as anti-biofilm agents, their practical application faces substantial challenges. The bioactive components of Eos may exhibit dose-dependent toxicity, pose organ-specific risks, and present administration-route difficulties. Current literature underscores critical gaps, including a lack of standardized toxicological assessments, divergent safety profiles among different Eos, and insufficient in vivo validation of efficacy and safety. Collectively, these hurdles significantly impede the translation of Eos from laboratory research to broad clinical or industrial applications [382,383].
6.5.13. The Role of Nanotechnology and Biopolymers: A Case Study in P. mirabilis
Nanotechnology represents a significant innovative frontier in combating resistant pathogens. Nano-formulations offer advantages such as improved drug delivery, enhanced bioavailability, and intrinsic antimicrobial activity. A key biopolymer in this field is chitosan (CS), a naturally occurring, biodegradable, and biocompatible positively charged polysaccharide derived from chitin. Its antimicrobial activity stems from electrostatic attraction to negatively charged bacterial cell surfaces, leading to membrane damage and disruption of cellular processes [384].
The true potential of chitosan is unlocked through nanoformulation. Chitosan nanoparticles (CSNPs) exhibit enhanced efficacy due to their high surface area-to-volume ratio, increased cellular uptake, and ability to overcome bacterial resistance mechanisms [385]. This makes them promising candidates against XDR and PDR bacteria, including P. mirabilis. Applied research demonstrates this potential against P. mirabilis. For instance, a trimethoprim nanoemulsion showed an eight-fold reduction in MIC compared to the plain drug [386]. Synergistic effects have also been observed with combinations like curcumin-silver nanoparticles (Cur-AgNPs) [387].
The arsenal of effective nanomaterials is expanding. Biosynthesized ZnO-CuO nanocomposites have shown significant promise against MDR P. mirabilis [388], these nanoparticles inhibited biofilm formation in robust biofilm-producing strains over 24–48 h. Furthermore, treatment with the ZnO-CuO nanocomposite led to the downregulation of the luxS gene, a key regulator of quorum sensing [388].
A recent translational study evaluated CS, CSNPs, and CSNPs combined with ciprofloxacin (CIP) against XDR/PDR P. mirabilis isolates from livestock [79]. The key findings were (Table 4):
- Synergistic enhancement: The CSNPs + CIP combination showed a 50–58% synergistic increase in the inhibition zone area compared to CIP alone.
- Activity against extreme resistance: CSNPs were highly active (MICs: 0.067–0.081 mg/mL) against all isolates, irrespective of their XDR/PDR status or the presence of resistance genes (blaTEM, *mcr-*1, qnrA). This suggests CSNPs’ membrane-disruptive, multi-targeted action bypasses common resistance mechanisms.
- Comparative advantage: The efficacy of CSNPs was competitive or superior to other nanomaterials like AgNPs, CuNPs, ZnNPs, and Se-based composites tested against P. mirabilis [386,387,388,389,390,391].
- Anti-virulence activity: Sub-MICs of both CS and CSNPs significantly reduced the swarming motility of P. mirabilis, a key virulence factor for colonization [79]. This aligns with antivirulence strategies, where CS-based materials can attenuate bacterial pathogenicity, potentially impeding biofilm formation and increasing susceptibility to host defenses and antibiotics.
The objective of these nano-strategies is twofold: to re-sensitize MDR isolates to conventional antibiotics and to provide alternative therapeutic approaches [248]. The use of antimicrobial nanomaterials has garnered considerable attention in managing infections caused by MDR pathogens like Proteus [79,392]. Accelerating the development and regulatory approval of such novel tools is crucial to ensure effective treatment options for highly resistant strains. While antimicrobial stewardship to prevent resistance emergence is paramount, the parallel development of novel agents and alternative strategies—exemplified by CSNPs, phage therapy, and antivirulence compounds—is a vital long-term solution to this public health crisis [79,392].
6.5.14. Public Awareness and Education as Cornerstones of AMR Mitigation
Public engagement and educational efforts are basic building blocks of the struggle against AMR in a One Health approach worldwide. Such efforts are the backbone of the broader approach used to address issues of antibiotic resistance [393]. It is important to conduct targeted educational efforts aimed at various segments of society such as the general population as well as healthcare professionals and agricultural workers regarding the basics of rational use and the grave risks associated with irrational use of antibiotics.
The major learning objectives include understanding what AMR is and what its health-related meaning and significance are, promoting prudent usage in both human and veterinary health, and stressing the significance of hygiene- and food-related practices, as well as effective vaccination practices, and making one aware of how everything is interlinked in human, animal, and environmental health. The above-mentioned integrated approach to education helps bring about prudent behavior and, in this way, decreases irresponsible usage and, in this manner, establishes an anti-microbial culture in society in this context [394]. Such initiatives are further supplemented by government policies, some of which proclaim veterinary monitoring and prescription necessary in animal farming for using antibiotics [394]. Moreover, targeted policies are in place that do not allow the use of certain classes of antibiotics defined as critically important for human use, including fluoroquinolones and third-generation cephalosporins, in agricultural facilities with the aim of conserving them for human use [395,396]. Raising public consciousness and encouraging positive changes in behavior, public awareness campaigns could make a significant contribution to limiting the spread and effects of AMR, including the development of P. mirabilis resistance.
6.6. Challenges and Opportunities in Combating P. mirabilis Antimicrobial Resistance
Despite extensive worldwide efforts, there are several factors that impede the progress being made against AMR in P. mirabilis. First among these is the increased antibiotic resistance P. mirabilis exhibits upon biofilm formation, which drastically reduces the efficacy of typical antibiotic treatment schemes [141]. Second, the promising alternative bacteriophage therapy also faces severe obstacles such as the development of bacterial resistance to phages themselves and intricate regulatory pathways, which further delay its clinical use [243]. The pathogen exhibits an astonishing ability to acquire and disseminate novel resistance mechanisms rapidly, especially by means of MGEs, thereby allowing new resistance patterns to emerge and spread more rapidly than the pace of developing new therapeutic drugs. This remains a significant bottleneck in the intrinsically slow and expensive process of antibiotic discovery and development, leaving an increasingly narrow arsenal to fight MDR infections.
Adding to such treatment challenges are the systemic and surveillance gaps. There are also major gaps in integrated, One Health models of surveillance that cover the human, animal, and environmental contexts. This is because of the lack of a comprehensive approach to AMR epidemiology and the tracking of resistant strains. Moreover, a cultural change, which is a long-term and hard-won approach to counter the deeply ingrained practice of using antibiotics ineffectively in human and veterinary settings, requires an even greater approach to improve and enhance personal and environmental cleanliness and standards of infection control. The environmental release of resistant bacteria and ARGs continues to drive the cycle of AMR, which is a never-ending cycle. It is critical to tackle these complex problems in order to improve patient outcomes. The future of research needs to focus on developing innovative combination regimens involving the combination of traditional antibiotics with new strategies, including nanoparticles and phage therapy. Additionally, there is an urgent need for clinical trials aimed at validating the safety and efficacy of such emerging therapies.
However, there are significant opportunities to move forward in these challenges:
- Help the One Health approach: Interdisciplinary collaboration and a flow of data maybe the starting point for a more coordinated approach to AMR.
- Leveraging technological innovations: Advances in the areas of genomics, bioinformatics, and rapid diagnostics offer immense possibilities in identifying resistance genes quickly, tracing the path of their transmission, and elucidating their evolution.
- Funding novel therapeutic approaches: Ongoing efforts in the development of novel therapeutic modalities, such as phage therapies, antivirulence agents, and immunotherapeutic drugs, could potentially offer much in the way of novel
- Enactment of strong policy frameworks: The formulation and launch of effective international and national policies on antimicrobial stewardship, comprehensive surveillance, and the environment can help bring about major shifts. The areas of intervention are the development of “super” methods of surveillance and diagnosis that understand the zoonotic disease transmission to help shape the policy, development of infrastructure and education resources and availability of drugs in LMICs, development of guidelines based on “best practices” of existing treatments available, and investment directed at the development of “novel” therapies.
7. Concluding Remarks, One Health Implications, and Future Directions
P. mirabilis exemplifies the complex nexus of sophisticated virulence, rapid AMR acquisition, and cross-sectorial transmission that defines contemporary global health threats. This review underscores its evolution from a common uropathogen to a formidable multidrug-resistant adversary, whose clinical impact is amplified by its seamless circulation across human, animal, and environmental reservoirs. The evidence compels a unified perspective, crystallized in four key interconnected messages:
- Shared reservoirs and transmission pathways: P. mirabilis thrives in interconnected ecosystems. Genetically similar, multidrug-resistant strains circulate among humans, food-producing and companion animals, and environmental matrices. Transmission via food, direct contact, and environmental contamination creates a continuous cross-sectoral challenge.
- Convergence of virulence and resistance: The pathogen’s potent virulence factors (e.g., urease, swarming motility, robust biofilm) facilitate both severe infection and the persistence and spread of acquired AMR genes (e.g., ESBLs, carbapenemases) across all hosts.
- Unified genetic pool of resistance: Mobile genetic elements carrying identical resistance determinants (e.g., blaCTX-M, blaNDM) are found in isolates from hospitals, farms, and food products, confirming continuous gene flow across the One Health spectrum.
- Imperative for integrated solutions: Controlling resistant P. mirabilis demands coherent, cross-sectorial strategies that break down institutional silos through combined surveillance, unified antimicrobial stewardship, and innovation in therapies targeting shared vulnerabilities.
The staggering projections of AMR—including an estimated 92 million associated deaths worldwide by 2050 [397]—underscore the critical urgency of translating this knowledge into decisive, integrated action. While legislative models like the PASTEUR Act aim to stimulate antibiotic development [397,398], concerns persist regarding the sustainability of a strategy focused primarily on new drug discovery. Therefore, durable defense must be anchored in the full implementation of foundational One Health measures: eliminating antibiotics for growth promotion in livestock; reducing environmental contamination; and enforcing regulations to curb non-prescription antibiotic use [398,399,400]—a policy proven effective in settings like Saudi Arabia and Kerala, India [399,400]. The lag in adopting these evidence-based stewardship interventions remains a critical implementation gap.
Ultimately, achieving sustainable mitigation requires a multi-pronged strategy that strategically marries innovation with foundational stewardship. This involves fostering the development of alternative therapies and precision diagnostics—while diligently addressing translational challenges [401]—alongside robust, preemptive policies. Looking ahead, the path forward must be channeled into three concrete, integrated fronts:
- Advancing Integrated, Genomic-Driven Surveillance: Establishing active, real-time genomic surveillance networks across human, veterinary, and environmental sectors is paramount for detecting emerging resistance and tracking high-risk clones across the One Health continuum.
- Accelerating Targeted and Innovative Therapies: Prioritizing the development and translation of precision interventions, including anti-virulence compounds, phage-antibiotic combinations, novel anti-biofilm biomaterials, and nano-formulations (e.g., CSNPs), to bridge the gap between preclinical promise and clinical application.
- Forging Enforceable, Integrated Policy Frameworks: Implementing unified international regulations on antibiotic use in agriculture, harmonized AMR data standards, and stringent environmental controls to limit the discharge of resistant bacteria, recognizing that the broad host range of P. mirabilis makes this integration non-negotiable.
In conclusion, P. mirabilis serves as a critical test for the practical implementation of the One Health paradigm. Our collective, coordinated response across these fronts will determine our ability to safeguard antimicrobial efficacy and secure the health of interconnected human, animal, and environmental ecosystems.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yong T.S.M. Panting A.J. Juatan N. Perialathan K. Ahmad M. Ahmad Sanusi N.H. Hassan L. Jahis R. Shamsudin N. Yap S.L. Development and validation of a cognitive, affective and behaviour questionnaire on pet-associated zoonotic diseases (CAB-ZDQ)Vet. Med. Sci.202171558156310.1002/vms 3.54734137200 PMC 8464282 · doi ↗ · pubmed ↗
- 2Kirk M.D. Pires S.M. Black R.E. Caipo M. Crump J.A. Devleesschauwer B. Döpfer D. Fazil A. Fischer-Walker C.L. Hald T. World Health Organization estimates of the global and regional disease burden of 22 foodborne bacterial, protozoal, and viral diseases, 2010: A data synthesis P Lo S Med.201512 e 1001921 Erratum in P Lo S Med. 2015, 12, 10019402663383110.1371/journal.pmed.1001921 PMC 4668831 · doi ↗ · pubmed ↗
- 3WHO Antibiotic Resistance 2020(accessed on 8 July 2023)Available online: https://www.who.int/news-room/fact-sheets/detail/antibiotic-resistance
- 4van Wagenberg C.P. Havelaar A.H. Economic costs related to foodborne disease in Burkina Faso and Ethiopia in 2017 Front. Sustain. Food Syst.20237122743010.3389/fsufs.2023.1227430 · doi ↗
- 5van Wagenberg C.P. Delele T.G. Havelaar A.H. Patient-related healthcare costs for diarrhoea, Guillain Barré syndrome and invasive non-typhoidal salmonellosis in Gondar, Ethiopia, 2020 BMC Public Health 202222209110.1186/s 12889-022-14539-136384546 PMC 9670532 · doi ↗ · pubmed ↗
- 6World Bank Food-Borne Illnesses Cost US$ 110 Billion per Year in Low- and Middle-Income Countries (World Bank, 2018)(accessed on 23 October 2018)Available online: https://www.worldbank.org/en/news/press-release/2018/10/23/food-borne-illnesses-cost-us-110-billion-per-year-in-low-and-middle-income-countries
- 7Ergocun G. Unsafe Food Costs $110B Annually (World Bank. Aa Com Tr, 2018)(accessed on 31 October 2018)Available online: https://share.google/XZ Ncj E Mf 7ujw T Xz B 4
- 8Hoffmann S. Ahn J.-W. Updating Economic Burden of Foodborne Disease Estimates for Inflation and Income Growth Econ Research Report ERR-297USDA ERS Washington, DC, USA 2021 Available online: https://www.ers.usda.gov/publications/pub-details/?pubid=102639(accessed on 18 November 2021)