LC-ESI-MS/MS Analysis of Echium asperrimum from the Algerian Aurès Region: Antioxidant, Antimicrobial, Cholinesterase Inhibitory, and Antiproliferative Activities
Amina Guetteche, Hamza Fadel, Mohammed Esseddik Toumi, Khawla Kerbab, Aslı Yıldırım Kocaman, İlyas Yıldız, Süleyman Muhammed Çelik, Noor Nayel, Tevfik Ozen, Ibrahim Demirtas, Hichem Hazmoune, Ramazan Erenler, Lahcene Zaiter, Maria D’Elia, Luca Rastrelli

TL;DR
This study identifies bioactive compounds in Echium asperrimum from Algeria and shows they have antioxidant, antimicrobial, and anti-Alzheimer properties.
Contribution
The paper reports the first comprehensive analysis of Echium asperrimum's phenolic profile and its multiple bioactivities.
Findings
EAAE extract showed higher antioxidant activity than EAEE extract.
Both extracts inhibited cholinesterase enzymes similarly to the drug galantamine.
EAAE exhibited antiproliferative effects on cancer cells at specific concentrations.
Abstract
The aim of the present study was to characterize the phenolic profile of hydroethanolic (EAEE) and ethyl acetate (EAAE) extracts of Echium asperrimum and to evaluate their antibacterial, antioxidant, anti-Alzheimer-related (cholinesterase inhibitory) activity, and antiproliferative activities. The DPPH radical scavenging activity of EAEE and EAAE showed IC50 values of 32.53 ± 1.25 and 97.85 ± 2.31 µg/mL, respectively. In addition, both extracts exhibited phosphomolybdenum reduction capacity, with A0.50 values of 61.60 ± 2.97 µg/mL for EAEE and 23.20 ± 1.55 µg/mL for EAAE. Acetylcholinesterase and butyrylcholinesterase inhibition assays revealed IC50 values comparable to the reference compound galantamine. Both extracts also showed antimicrobial activity against Gram-positive and Gram-negative bacterial strains. LC-ESI-MS/MS analysis indicated that p-coumaric acid (5.12 mg/g), vanillic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
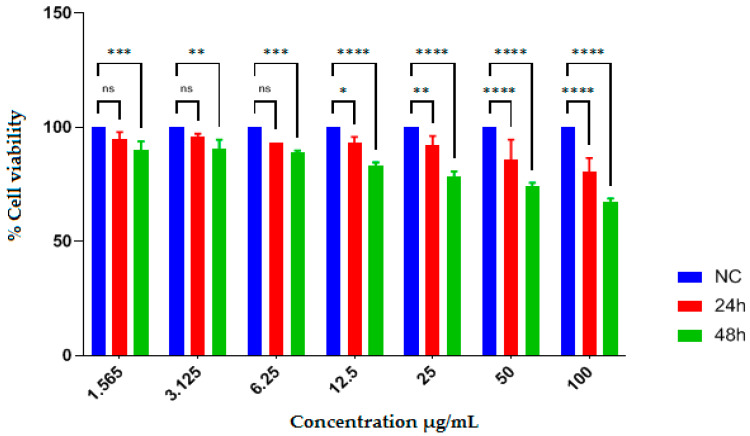 Figure 1
Figure 1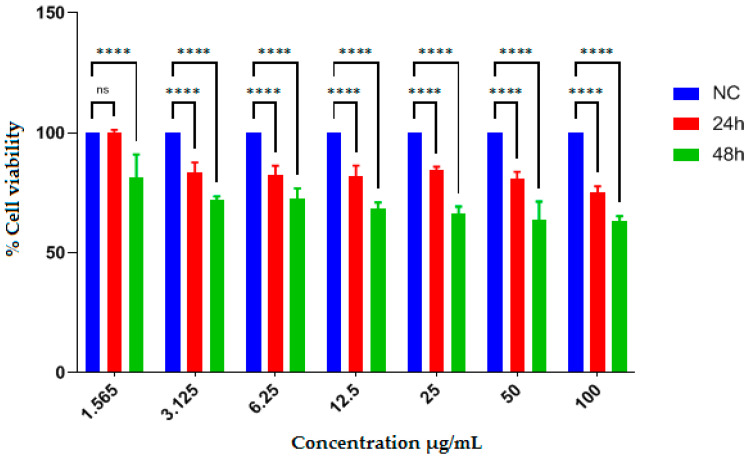 Figure 2
Figure 2- —National Recovery and Resilience Plan (NRRP)
- —Italian Ministry of University and Research
- —European Union—NextGenerationEU
- —Project title “National Biodiversity Future Center—NBFC”
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGinger and Zingiberaceae research · Medicinal Plants and Neuroprotection · Marine Sponges and Natural Products
1. Introduction
The genus Echium (Boraginaceae) comprises 67 recognized species native to North Africa, mainland Europe, and the Macaronesian region, including the Azores, Madeira, the Canary Islands, and Cape Verde. Species within this genus occur as annual, biennial, or perennial flowering plants [1].
Plants belonging to the Boraginaceae family have been traditionally used for the treatment of various ailments, including diuretic disorders, fever, asthma, kidney stones, and wound healing [2]. Since approximately 300 B.C., Echium species have played a relevant role in Mediterranean ethnomedicine due to their sedative, anti-inflammatory, antioxidant, and anxiolytic properties [3]. They have also been traditionally applied for the treatment of snakebites, hand fissures, and skin abrasions [4,5,6].
Liver cancer remains a major public health issue in the 21st century, representing such a dramatic impact on our lives [7]. Furthermore, despite their benefits, therapies are still costly, intrusive, and detrimental to the patient’s quality of life. Therefore, in order to enhance current anticancer drugs, new ones are required [8].
Owing largely to their high content of phenolic compounds, medicinal plants and their extracts represent an important resource for drug discovery, offering culinary, horticultural, and ethnomedicinal advantages with potential relevance for human health [9,10]. Antioxidants derived from medicinal plants are key components of traditional remedies, and their biological efficacy is strongly influenced by molecular complexity [11]. These compounds play a crucial role in the development of therapeutic agents and may contribute to reducing the risk of cardiovascular and other chronic diseases, as well as aging-related disorders [12]. Their biological activity is mainly attributed to strong radical-scavenging properties associated with phenolics, flavonoids, carotenoids, anthocyanins, and vitamins, which are closely linked to oxidative stress and lipid peroxidation processes [13].
From a phytochemical perspective, Echium species are known to contain a wide range of bioactive secondary metabolites, including naphthoquinones, flavonoids, terpenoids, and phenolic compounds [14]. These constituents have been associated with diverse biological effects, such as anxiolytic [15,16], antioxidant [17], anti-inflammatory [18], antibacterial [19], and antiviral activities [20]. Several reviews have addressed the geographical distribution of Echium species [21,22], as well as the pharmacological properties of specific taxa, particularly E. amoenum [23,24].
In recent years, increasing attention has been devoted to the validation of standardized plant extracts intended for nutraceutical and dietary supplement applications, particularly when such extracts reflect traditional medicinal use. In this context, Echium asperrimum is already commercially exploited through its seed oil, which is widely used as a plant-based food supplement rich in stearidonic acid (SDA) and gamma-linolenic acid (GLA). However, other aerial parts of the plant, including leaves, stems, and flowers, remain largely unexplored despite their potential as sources of bioactive compounds.
Therefore, the aim of the present study was to provide an extract-level chemical and biological characterization of E. asperrimum leaves, stems, and flowers, prepared in accordance with traditional use, in order to support their potential valorization as standardized nutraceutical ingredients.
2. Results
2.1. Antioxidant Activity
The antioxidant activities of the hydroethanolic extract (EAEE) and the ethyl acetate extract (EAAE) of E. asperrimum were evaluated using the DPPH free radical scavenging assay and the phosphomolybdenum reduction method. The results are summarized in Table 1.
Both extracts exhibited measurable antioxidant activity, although with different efficiencies depending on the assay employed. In the DPPH• scavenging assay, EAEE showed a significantly stronger radical scavenging capacity than EAAE, with IC_50_ values of 32.53 ± 1.25 µg/mL and 97.85 ± 2.31 µg/mL, respectively. This indicates a higher hydrogen- or electron-donating ability of EAEE toward the DPPH radical.
In contrast, the phosphomolybdenum reduction assay revealed a different trend. EAAE demonstrated a greater total antioxidant capacity than EAEE, as reflected by its lower A_0.5_ value (23.20 ± 1.55 µg/mL) compared with EAEE (61.60 ± 2.97 µg/mL). This result suggests that compounds enriched in the ethyl acetate fraction contribute more effectively to the overall reduction of Mo(VI) to Mo(V), highlighting assay-dependent differences in antioxidant behavior.
As expected, ascorbic acid exhibited significantly stronger antioxidant activity than both plant extracts in both assays, with IC_50_ and A_0.5_ values of 4.78 ± 0.14 µg/mL and 4.27 ± 0.05 µg/mL, respectively. Nevertheless, the observed antioxidant activities of EAEE and EAAE confirm the presence of redox-active constituents in E. asperrimum extracts.
2.2. LC-ESI MS/MS Analysis of E. asperrimum Extracts
The qualitative and quantitative profiles of phenolic compounds in EAEE and EAAE were determined by LC-ESI-MS/MS analysis. The identified compounds, their retention times, and concentrations are reported in Table 2.
From an extract-level perspective, clear differences were observed in terms of phenolic composition and abundance. In the EAEE, p-coumaric acid (0.13 mg/g), salicylic acid (0.141 mg/g), sinapic acid (0.02 mg/g), and trans-ferulic acid (0.02 mg/g) were detected as the most abundant phenolic constituents. Additional compounds were present at lower concentrations, including gentisic acid (0.092 mg/g), syringic acid (0.027 mg/g), caffeic acid (0.015 mg/g), trans-cinnamic acid (0.01 mg/g), and gallic acid (0.003 mg/g). The isoflavone biochanin A was also detected in EAEE at a low concentration (0.003 mg/g).
In contrast, the EAAE showed a markedly higher accumulation of several phenolic acids and related compounds. Vanillic acid (11.6 mg/g) was the predominant constituent, followed by p-coumaric acid (5.12 mg/g), hydroxybenzaldehyde (3.82 mg/g), gentisic acid (1.66 mg/g), and salicylic acid (1.05 mg/g). Lower amounts of caffeic acid (0.38 mg/g), sinapic acid (0.23 mg/g), trans-ferulic acid (0.22 mg/g), trans-cinnamic acid (0.36 mg/g), syringic acid (0.04 mg/g), rutin (0.02 mg/g), vanillin (0.02 mg/g), protocatechuic acid (0.01 mg/g), and protocol ethyl ester (0.01 mg/g) were also detected.
These results demonstrate a distinct distribution of phenolic compounds between EAEE and EAAE, with quantitative differences clearly highlighted by LC-ESI-MS/MS analysis.
2.3. Antibacterial Evaluation
The antibacterial activity of EAEE and EAAE was evaluated against selected Gram-negative and Gram-positive bacterial strains. The results, expressed as minimum inhibitory concentration (MIC) values, are reported in Table 3.
Both extracts exhibited antibacterial activity with MIC values ranging from 32 to 128 µg/mL, depending on the bacterial strain and extract type. Against Gram-negative bacteria, both EAEE and EAAE showed identical inhibitory activity toward Escherichia coli, with MIC values of 64 µg/mL. In the case of Pseudomonas aeruginosa, EAAE exhibited stronger antibacterial activity (32 µg/mL) compared to EAEE (64 µg/mL).
For Gram-positive bacteria, EAAE demonstrated higher inhibitory potency than EAEE. Notably, EAAE showed the lowest MIC value against Staphylococcus aureus (32 µg/mL), whereas EAEE inhibited the same strain at 64 µg/mL. The weakest antibacterial activity was observed for EAEE against Bacillus cereus, with an MIC value of 128 µg/mL.
The reference antibiotics ofloxacin and chloramphenicol exhibited markedly stronger antibacterial effects than the plant extracts against all tested strains.
2.4. AChE and BChE Activities
The inhibitory effects of EAEE and EAAE on acetylcholinesterase (AChE) and butyrylcholinesterase (BChE) activities were assessed using in vitro enzyme inhibition assays. The results are expressed as half-maximal inhibitory concentration (IC_50_) values and are summarized in Table 4.
Both extracts exhibited notable inhibitory activity against AChE and BChE. The EAAE showed stronger inhibitory potency than EAEE toward both enzymes. Specifically, EAAE inhibited AChE with an IC_50_ value of 1.33 ± 0.01 µg/mL, whereas EAEE displayed an IC_50_ value of 3.03 ± 1.09 µg/mL. A similar trend was observed for BChE inhibition, with EAAE showing an IC_50_ value of 0.03 ± 0.00 µg/mL, compared to 0.05 ± 0.03 µg/mL for EAEE.
The inhibitory activities of both extracts were comparable to those of the reference compound galantamine.
2.5. Antiproliferative Effect
The antiproliferative effects of EAEE and EAAE were evaluated using the human hepatocellular carcinoma cell line HepG2, using the MTT assay. Cell viability was assessed after 24 and 48 h of treatment at concentrations ranging from 1.565 to 100 µg/mL. The IC_50_ values are reported in Table 5, while the dose–response effects are illustrated in Figure 1 and Figure 2.
Treatment with EAEE did not result in a measurable IC_50_ value at either 24 or 48 h. At low concentrations (1.565–6.25 µg/mL), no significant reduction in cell viability was observed at either time point. At higher concentrations (12.5–100 µg/mL), a decrease in cell viability was detected, which was more pronounced after 48 h of exposure. Statistically significant reductions were observed at 50 and 100 µg/mL after 48 h (p < 0.05 to p < 0.0001). The negative control (normal control cells) maintained 100% viability, indicating that the observed effects were not attributable to the solvent (DMSO).
In contrast, EAAE showed a stronger antiproliferative effect. While no significant reduction in cell viability was detected at the lowest concentration tested (1.565 µg/mL), EAAE induced a significant, concentration-dependent decrease in cell viability starting from 3.125 µg/mL, particularly after 48 h of incubation. At higher concentrations (50 and 100 µg/mL), cell viability decreased to below 50% after 48 h, corresponding to an IC_50_ value of 83.09 ± 6.50 µg/mL.
The positive control, 5-fluorouracil, exhibited stronger antiproliferative effect than both plant extracts, with IC_50_ values of 75.93 ± 3.04 µg/mL at 24 h and 17.53 ± 3.42 µg/mL at 48 h.
3. Discussion
The present study provides the first integrated extract-level evaluation of the antioxidant, antimicrobial, cholinesterase inhibitory, and antiproliferative effects of Echium asperrimum extracts.
The overall findings indicate that E. asperrimum represents a promising source of bioactive compounds, exhibiting antioxidant, antibacterial, anti-Alzheimer-related (cholinesterase inhibitory) activity, and antiproliferative properties. These biological effects are closely associated with the phenolic profile revealed by LC-ESI-MS/MS analysis.
At the extract level, the differences in biological activities observed between EAEE and EAAE appear consistent with their distinct phenolic profiles. In particular, the higher abundance of phenolic acids such as vanillic acid, p-coumaric acid, gentisic acid, and hydroxybenzaldehyde in EAAE, widely reported as bioactive phenolics with antioxidant, antimicrobial, and enzyme-modulating properties, may contribute to its stronger antioxidant, antimicrobial, cholinesterase inhibitory, and antiproliferative effects compared to EAEE. Although no formal correlation analysis was conducted, this comparative, trend-based interpretation is commonly adopted in phytochemical screening studies. The antioxidant activity observed for E. asperrimum extracts can be primarily attributed to their phenolic composition, particularly phenolic acids such as caffeic acid, p-coumaric acid, sinapic acid, ferulic acid, and related compounds. Previous studies on Echium species have consistently demonstrated that antioxidant potential is strongly dependent on the qualitative and quantitative composition of secondary metabolites, especially polyphenols, flavonoids, lignans, echimidine, acetylshikonin, and quinic acid [25,26,27]. Variations in antioxidant efficiency among species and extracts are therefore expected and reflect differences in phenolic and flavonoid content.
Comparable antioxidant activities have been reported for other Echium species. For instance, extracts of E. arenarium showed strong radical scavenging capacity, accompanied by antibacterial and cytotoxic activities, which were attributed to a high phenolic content. This species exhibited remarkable antioxidant activity against DPPH (IC_50_ = 1.1 µg/mL), pronounced antibacterial effects against Gram-positive bacteria (Listeria monocytogenes, Staphylococcus aureus, MRSA, Enterococcus faecalis, and Bacillus cereus), and cytotoxicity toward the RAW 264.7 macrophage cell line (IC_50_ = 145.80 ± 0.84 µg/mL, SI < 10) [28]. Similarly, ethanolic extracts of E. vulgare collected in Turkey demonstrated DPPH inhibition values of 57.24 ± 0.014% for aerial parts and 69.99 ± 0.008% for roots at 500 µg/mL, while E. angustifolium extracts showed 45.32 ± 0.028% inhibition at 1000 µg/mL [29]. In addition, the ethanolic extract of E. parviflorum exhibited IC_50_ values ranging between 500 and 1000 µg/mL [29]. Moderate antioxidant activity was also reported for non-polar extracts of E. angustifolium, as assessed by DPPH and ABTS assays, with Trolox equivalent antioxidant capacities of 131.05 and 1407.24 μM TE/mg extract, respectively [26]. Furthermore, Chaouche et al. [30] reported notable antioxidant potential for methanolic root extracts of E. pycnanthum collected in southern Algeria. Actually, a number of studies have demonstrated that phenolics like vanillic (IC_50_ = 47.4 µg/mL), gentisic (EC_50_ = 0.09), and p-coumaric acids (71.2% lipid peroxidation of a linoleic acid emulsion at 45 μg/mL), and hydroxybenzaldehyde (EC_50_ = 3.4) are strong antioxidants present in plants that serve as primary defense mechanisms against oxidative stress and free radicals by neutralizing free radicals [31,32,33,34].
In addition to antioxidant effects, E. asperrimum extracts displayed relevant antibacterial activity against both Gram-positive and Gram-negative bacteria. This activity can be correlated with the presence of phenolic acids and related compounds detected in the extracts. Phenolics including p-coumaric (MIC = 256–128 µg/mL) 0.2–0.1 mg/mL, gentisic (SICs = 0.0625, 0.125, and 0.25 mg/mL), and vanillic acids (MICs = 0.8–0.2 mg/mL), as well as hydroxybenzaldehyde derivatives (MICs = 0.14–0.58 mg/mL), have been shown in numerous studies to be potent antibacterial agents by breaking down cell walls, blocking enzyme systems, and slowing microbial growth [35,36,37,38]. Additionally, previous research has demonstrated that rosmarinic acid, chlorogenic acid, p-hydroxybenzoic acid, and rutin are largely responsible for the antibacterial properties of E. italicum extracts [39]. Moreover, Shariatifar et al. [40] reported strong antibacterial activity of E. amoenum aqueous extracts against Staphylococcus aureus, Escherichia coli, and Pseudomonas aeruginosa. Notably, the antibacterial activity observed for the ethyl acetate extract of E. asperrimum appears higher than that previously reported for E. amoenum against L. monocytogenes and L. ivanovii (IC_50_ = 5 mg/mL) [41], further highlighting the relevance of this species as a source of antimicrobial agents.
The anti-Alzheimer potential of E. asperrimum extracts was demonstrated through inhibition of acetylcholinesterase (AChE) and butyrylcholinesterase (BChE), with inhibitory activities comparable to those of galantamine. According to several in vitro studies phenolics could exhibit significant anti-Alzheimer potential, primarily through antioxidant mechanisms. Szwajgier [42] reported that several phenolic acids, including homogentisic acid, 4-hydroxyphenylpyruvic acid, nordihydroguaiaretic acid, rosmarinic acid, caffeic acid, gallic acid, chlorogenic acid, homovanillic acid, and sinapic acid, exhibit notable anti-acetylcholinesterase activity. Additionally, ferulic acid, p-coumaric acid, 4-hydroxybenzoic acid, and sinapic acid have been shown to act as efficient inhibitors of both AChE and BChE in vitro [43], supporting the enzymatic inhibition observed in the present study. These findings are in agreement with previous reports on phenolic-rich plant extracts and Echium species, which have shown comparable cholinesterase inhibitor, thereby reinforcing the relevance of the present results within the existing literature.
Finally, the antiproliferative effect of E. asperrimum extracts was evaluated against the HepG2 cancer cell line, revealing stronger inhibitory effects for the ethyl acetate extract compared to the hydroethanolic extract. Phenolics determined by LC-ESI-MS/MS may have strong antiproliferative effects, which would prevent cancer cells from proliferating and cause apoptosis and cell cycle arrest [44]. Previous studies have demonstrated that p-coumaric acid exerts protective and antiproliferative effects against colorectal cancer and skin melanoma (A431 cell line) by inducing apoptosis, reducing reactive oxygen species (ROS), and modulating mitochondrial membrane potential. Moreover, p-coumaric acid inhibits NF-κB activation and downregulates inflammatory mediators such as TNF-α, IL-1β, IL-6, IL-8, COX-2, and PGE_2_ [45,46]. It has also been reported that p-coumaric acid supplementation reduces glucose-regulated protein 78 (GRP78) expression and activates unfolded protein response (UPR)-mediated apoptosis in both in vitro and in vivo colon cancer models [47]. GRP78 plays a key role in angiogenesis, epithelial–mesenchymal transition (EMT), and therapy resistance in malignant cells [47,48].
In addition, caffeic acid has been identified as an antiproliferative compound capable of inducing apoptosis in T47D human breast cancer cells through activation of the Fas/FasL death receptor pathway [49]. Rezaei-Seresht et al. [50] further demonstrated that caffeic acid modulates the expression of apoptosis-related genes, including P53, Mcl-1, and P21, leading to intrinsic apoptotic pathways in MCF-7 breast cancer cells. These findings collectively support the antiproliferative effects observed for E. asperrimum extracts in the present study. The antiproliferative effects observed in HepG2 cells are consistent with previous studies on phenolic-rich plant extracts and Echium species, which have reported growth-inhibitory activity against cancer cell lines, thereby supporting the biological relevance of the present findings. It should be emphasized that the present study aims to establish composition–activity relationships at the extract level rather than direct causal effects of individual compounds. While the relative abundance of major phenolics provides a rational basis for interpreting the observed bioactivities, further studies involving compound isolation and targeted mechanistic investigations are required to confirm these associations. Taken together, from a nutraceutical development perspective, the differences observed among antioxidant, antimicrobial, cholinesterase inhibitory, and antiproliferative activities between EAEE and EAAE are consistent with their distinct phenolic profiles, supporting a composition–activity relationship at the extract level rather than direct causal effects of individual compounds.
It should also be emphasized that the present study focuses on a standardized plant extract prepared in accordance with traditional medicinal use, rather than on isolated single compounds. Echium species are already widely exploited in the nutraceutical sector, particularly through Echium asperrimum seed oil, which is commercially available as a plant-based food supplement rich in stearidonic acid (SDA) and gamma-linolenic acid (GLA), and is used for cardiovascular health, anti-inflammatory purposes, and skin well-being.
In this context, the present work aims to provide a scientific rationale for extending the valorization of E. asperrimum beyond seed oil, by exploring the biological potential of leaves, stems, and flowers as standardized extracts. The extract-level approach adopted here is therefore consistent with real-world nutraceutical development, where complex mixtures—rather than isolated molecules—are the final products intended for human use.
4. Materials and Methods
4.1. Reagents and Standards
2,2-diphenyl-1-picrylhydrazyl (DPPH) was obtained from Sigma-Aldrich, St. Louis, MO, USA. Analytical standard ascorbic acid and BHT were obtained from Sigma-Aldrich, St. Louis, MO, USA (purity ≥ 98%). The highest quality standards for LC-ESI-MS analysis were acquired from Sigma Chemical Co. (St. Louis, MO, USA). All organic solvents and chemicals were of analytical grade. Gram-positive bacteria, such as Bacillus cereus (CCM 99) and Staphylococcus aureus (ATCC 25213), and negative bacterial strains, such as Escherichia coli (ATCC 25922) and Pseudomonas aeruginosa (ATCC 15442), were acquired from the Faculty of Agriculture at Ondokuz Mayıs University. Standard antibiotics included chloramphenicol and ofloxacin. Mueller Hinton Agar (MHA) from Himedia (Plot No. C40, Road No. 21Y, MIDC, Wagle Industrial Estate, Thane (West)-400604, Maharashtra, India), galantamine, ofloxacin, chloramphenicol from Sigma-Aldrich, AChE from Electrophorus, galantamine, ofloxacin, chloramphenicol from Sigma-Aldrich, AChE from Electrophorus electricus, and BChE from horse serum.
4.2. Plant Collection
Echium asperrimum Lam. (leaves, stems, and flowers) was collected in May 2022 from the Algerian Aurès region (Timgad area, approximately 35 km east of Batna; 35°29′05″ N, 6°28′07″ E; altitude: 1072 m). The plant material was taxonomically confirmed by Pr. Rebbas Khellaf, (an expert in regional flora at Department of Botanical Science of M’sila University, Algeria), and a voucher specimen (EA/138/VAR/05-22) was authenticated and deposited in the herbarium of the VARENBIOMOL Research Unit, Constantine 1 University (Algeria).
4.3. Extraction and Preparation of Samples
The dried plant material (leaves and flowers, 1300 g) of E. asperrimum was finely powdered and subjected to maceration at room temperature in a hydroalcoholic solvent system (ethanol/water, 70:30, v/v). The use of powdered material increased the contact surface with the extraction solvent, thereby enhancing extraction efficiency.
The maceration process was repeated four times with solvent renewal, and each extraction cycle lasted between 24 and 48 h. The combined extracts were concentrated under reduced pressure at a temperature not exceeding 35 °C to obtain the crude hydroethanolic extract (EAEE), with an extraction yield corresponding to approximately 400 mL per kg of dry plant material.
After filtration, the obtained crude ethanol extract (233.3 g) was subjected to successive liquid–liquid partitioning using solvents of increasing polarity, namely chloroform, ethyl acetate, and n-butanol. The resulting organic phases were separately dried over anhydrous sodium sulfate, filtered, and evaporated to dryness under reduced pressure to obtain the corresponding fractions, CHCl_3_ (0.73 g), EtOAc (EAAE, 0.82 g), and n-BuOH (4.71 g).
4.4. DPPH Free Radical Scavenging Assay
Antioxidant activity was evaluated using the 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging assay, following the method as described by Sharma and Bhat [51] with minor modifications. Briefly, 1450 µL of a 0.06 mM DPPH solution was mixed with 50 µL of EAEE and EAAE. Ascorbic acid was used as a positive control, while pure ethanol served as the blank.
The reaction mixtures were incubated for 30 min at room temperature in the dark, after which absorbance was measured at 515 nm using a UV–Vis spectrophotometer. A calibration curve was prepared using ascorbic acid standard solutions.
The DPPH radical scavenging activity was calculated according to Equation (1):
where A_control_ represents the absorbance of the negative control and A_sample_ corresponds to the absorbance of the tested extract.
4.5. Phosphomolybdenum Reduction Assay
The total antioxidant capacity of the samples was assessed by the phosphomolybdenum reduction assay using the ammonium molybdate method [52,53].
An aliquot of 300 µL of each sample or standard solution (BHT and ascorbic acid) at different concentrations was mixed with 2.7 mL of reagent solution to obtain a homogeneous mixture. The reaction mixtures were incubated in a shaking water bath at 95 °C for 90 min.
After cooling to room temperature, 200 µL of each mixture was transferred into a 96-well microplate, and the absorbance was recorded at 695 nm using a BIOTEK Epoch2 microplate reader. The antioxidant capacity was expressed as A_0.5_ (µg/mL). The combination of DPPH radical scavenging and phosphomolybdenum reduction assays was selected to provide complementary information on the antioxidant capacity of the extracts, covering both radical scavenging activity and total reducing potential.
4.6. LC-ESI MS/MS Analysis of Phenolic Compounds
Quantitative analysis of phenolic compounds was performed with minor modifications to the method described by Erenler and Karan [54]. Briefly, 50 mg of each extract fraction were dissolved in 1 mL of a solvent mixture consisting of acetonitrile–methanol–water (1:1:1, v/v/v) in 2 mL Eppendorf tubes. The samples were vortexed and sonicated for 20 min at 10,000 rpm.
Subsequently, 0.8 mL of n-hexane was added, and the mixtures were centrifuged at 7000 rpm for 5 min using a Hettich Universal 320 R centrifuge (Hettich GmbH & Co. KG, Föhrenstraße 12, D-78532 Tuttlingen, Germany). The lower phase was collected, diluted 1:4 with distilled water, and filtered through a Captiva Premium Syringe Layered Filter (0.2 µm, 25 mm diameter).
Chromatographic separation was achieved using a Poroshell 120 EC-C18 column coupled to a liquid chromatography–electrospray ionization tandem mass spectrometry (LC-ESI-MS/MS) system, consisting of an Agilent Technologies 1260 Infinity II liquid chromatograph interfaced with a 6460 Triple Quadrupole mass spectrometer. The analytes were detected using an electrospray ionization (ESI) source operated in both positive and negative ionization modes.
The injection volume was set at 0.4 µL, with a constant flow rate of 0.400 mL/min and a total run time of 40 min. The mobile phase consisted of solvent A (water containing 0.1% formic acid and 5 mM ammonium formate) and solvent B (acetonitrile containing 0.1% formic acid). The gradient elution program was as follows: 75% A/25% B for 5 min; linear gradient from 25% to 75% B over 15 min; 100% B at 16 and 20 min; followed by re-equilibration to 85% A/15% B from 22 to 40 min.
The nitrogen gas flow rate was maintained at 11 L/min, and the column temperature was set at 30 °C. A total of 31 standard compounds, including phenolic acids and flavonoids, were used for compound identification and quantification. Secondary metabolites were identified by comparing MS/MS spectra, retention times (RT), and mass spectral data with those of authentic standards and published literature. Detailed LC-ESI-MS/MS parameters, including precursor/product ion transitions and fragmentation patterns for the compounds listed in Table 2, are provided in the Supplementary Materials (Table S1).
4.7. Antimicrobial Evaluation
The antimicrobial activity of EAEE and EAAE was evaluated against Gram-negative bacteria, including Escherichia coli (ATCC 25922) and Pseudomonas aeruginosa (ATCC 15442), as well as Gram-positive bacteria, including Bacillus cereus (CCM 99) and Staphylococcus aureus (ATCC 25213). Chloramphenicol and ofloxacin were used as standard reference antibiotics for in vitro MIC determination and comparative purposes.
Bacterial strains were grown on sterile Mueller–Hinton agar (MHA) plates, which were prepared and allowed to solidify. The cultures were streaked onto the agar surface using a sterile inoculating loop and incubated at 37 °C for 16–18 h. Stock cultures were preserved at −80 °C in sterile glycerol solution (20%, v/v).
The antibacterial activity of the samples was determined by the broth microdilution method for the determination of the minimum inhibitory concentration (MIC), according to the protocol described by Andrews [55]. To prepare the cation-supplemented medium, CaCl_2_ and MgCl_2_ stock solutions (2 mg/mL each) were prepared. Subsequently, 400 µL of MgCl_2_ and 1 mL of CaCl_2_ were added to 100 mL of Mueller–Hinton II broth (MHB).
A bacterial suspension was prepared by transferring a portion of a fresh bacterial culture into 5 mL of sterile physiological saline solution (NaCl, 0.85%) and adjusting the turbidity to a 0.5 McFarland standard. In a sterile 96-well microplate, 100 µL of cation-supplemented MHB was added to each well. Subsequently, 100 µL of the test samples (1024 µg/mL) or reference antibiotics (ofloxacin, 2048 µg/mL; chloramphenicol, 1024 µg/mL) were added to the first well. Serial two-fold dilutions were performed by transferring 100 µL across the wells up to the 12th well, from which 100 µL was discarded.
The bacterial inoculum was prepared by adding 1 mL of the 0.5 McFarland bacterial suspension to 9 mL of cation-supplemented MHB, and 5 µL of this inoculum was added to each well. The microplates were kept at 4 °C for 2 h to allow diffusion and then incubated at 37 °C for 16–18 h.
The MIC was defined as the lowest concentration of the extract that completely inhibited visible bacterial growth, as indicated by the absence of turbidity compared with the growth control. MIC values were expressed in µg/mL.
4.8. Cholinesterase Inhibitory Activity
The anti-Alzheimer effect of the extracts was evaluated by enzymatic inhibition assays targeting acetylcholinesterase (AChE) and butyrylcholinesterase (BChE). The inhibitory activities against AChE and BChE were determined using the method described by Ellman et al. [56] with slight modifications.
Briefly, in a 96-well microplate, 20 µL of sample solutions or the reference inhibitor galantamine at different concentrations were mixed with 20 µL of AChE or BChE solution (0.03 U, prepared in 100 mM Na–K phosphate buffer, pH 8.0), 20 µL of DTNB solution (3.3 mM), and 140 µL of the same buffer. The reaction mixtures were incubated at room temperature for 15 min.
Subsequently, 10 µL of the corresponding substrate (1 mM acetylcholine iodide for AChE or butyrylcholine chloride for BChE) was added to initiate the enzymatic reaction. The absorbance was measured at 412 nm using a BIOTEK Epoch2 microplate reader (100 Tigan Street, Highland Park, P.O. Box 998, Winooski, VT 05404-0998, USA). Enzyme inhibitory capacity was expressed as the half-maximal inhibitory concentration (IC_50_, µg/mL).
4.9. Evaluation of Antiproliferative Effects
The antiproliferative effect of the hydroethanolic extract (EAEE) and ethyl acetate extract (EAAE) was evaluated against the human liver cancer cell line HepG2, using a normal liver cell line as a control. The HepG2 cell line used in this study was obtained from the cell stocks of the Cell Culture Laboratory, Research and Application Laboratory, Iğdır University.
HepG2 cells were cultured in RPMI-1640 medium supplemented with 10% (v/v) heat-inactivated fetal bovine serum (FBS) and 1% (v/v) L-glutamine at 37 °C in a humidified incubator with 5% CO_2_. Prior to treatment, cells were seeded into 96-well plates at a density of 5 × 10^4^ cells/mL and allowed to adhere and reach the logarithmic growth phase by incubation for 24 h.
Plant extracts were prepared by dissolving 10 mg of each extract in 100 µL of dimethyl sulfoxide (DMSO) to obtain stock solutions at a concentration of 100 mg/mL. To minimize solvent-induced cytotoxicity, the final concentration of DMSO in the culture medium did not exceed 0.1% (v/v). The initial working concentration was set at 100 µg/mL, followed by serial dilutions to a final concentration of 1.56 µg/mL. Cells were treated with the extracts for 24 and 48 h.
Cell viability was assessed using the MTT assay, following the method described by Bilgin et al. [57] with slight modifications. After treatment, cells were washed with phosphate-buffered saline (PBS) and incubated with 10 µL of Thiazolyl Blue Tetrazolium Bromide (MTT) solution (5 mg/mL in distilled water) for 3 h. Subsequently, the MTT-containing medium was removed, and DMSO was added to dissolve the formazan crystals formed.
Absorbance was measured at 570 nm using a microplate reader. Cell viability was calculated as a percentage relative to untreated control cells, and antiproliferative effect was evaluated by plotting dose–response curves and calculating IC_50_ values using a curve-fitting algorithm.
4.10. Statistical Analysis
Results are expressed as mean ± standard deviation (SD). IC_50_ and A_0.50_ values were calculated using linear regression analysis. One-way analysis of variance (ANOVA) was performed using GraphPad Prism version 8.0 software, and statistically significant differences between means were determined using Tukey’s post hoc test. A p-value < 0.05 was considered statistically significant. For antioxidant assays, statistical comparisons were performed between each extract and the reference compound.
5. Conclusions
This study provides the first integrated characterization of the phenolic profile and extract-level biological activities of Echium asperrimum collected from the Algerian Aurès region. The results indicate that the hydroethanolic and ethyl acetate extracts exhibit relevant antioxidant, antibacterial, cholinesterase inhibitory, and antiproliferative effects, which are consistent with their distinct phenolic compositions as revealed by LC-ESI-MS/MS analysis.
Although the present work does not aim to establish direct causal relationships at the single-compound level, it highlights E. asperrimum as a promising natural source of phenolic-rich extracts with potential relevance for pharmaceutical and nutraceutical research. Further studies will be required to clarify the contribution of individual compounds and to elucidate the molecular mechanisms underlying the observed bioactivities.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sheydaei P. Amaral M.E. Duarte A.P. Genus Echium L.: Phytochemical Characterization and Bioactivity Evaluation for Drug Discovery Plants 202514254810.3390/plants 1416254840872171 PMC 12389265 · doi ↗ · pubmed ↗
- 2Al Snafi A.E. The Pharmacology of Anchusa italica Retz. and Anchusa strigosa—A review Int. J. Pharm. Pharm. Sci.20146710
- 3Terzieva S. Grozeva N. Tzanova M. A Review of the Main Biologically Active Compounds of the Genus Echium L., Naturally Distributed in Bulgaria, and Their Pharmacological Potential Pharmaceuticals 202518161810.3390/ph 1811161841304864 PMC 12655106 · doi ↗ · pubmed ↗
- 4Jin J. Boersch M. Nagarajan A. Davey A.K. Zunk M. Antioxidant Properties and Reported Ethnomedicinal Use of the Genus Echium (Boraginaceae)Antioxidants 2020972210.3390/antiox 908072232784832 PMC 7466025 · doi ↗ · pubmed ↗
- 5Rabbani M. Sajjadi S.E. Vaseghi G. Jafarian A. Anxiolytic Effects of Echium amoenum on the Elevated Plus-Maze Model of Anxiety in Mice Fitoterapia 20047545746410.1016/j.fitote.2004.04.00415261383 · doi ↗ · pubmed ↗
- 6Potdar V.H. Kibile S.J. Evaluation of Antidepressant-like Effect of Citrus Maxima Leaves in Animal Models of Depression Iran J. Basic Med. Sci.20111447848323492865 PMC 3586848 · pubmed ↗
- 7Arzumanian V.A. Kiseleva O.I. Poverennaya E.V. The curious case of the Hep G 2 cell line: 40 years of expertise Int. J. Mol. Sci.2021221313510.3390/ijms 22231313534884942 PMC 8658661 · doi ↗ · pubmed ↗
- 8Arredondo-Beltrán I.G. Ramírez-Sánchez D.A. Zazueta-García J.R. Canizalez-Roman A. Angulo-Zamudio U.A. Velazquez-Roman J.A. Bolscher J.G.M. Nazmi K. León-Sicairos N. Antitumor Activity of Bovine Lactoferrin and Its Derived Peptides against Hep G 2 Liver Cancer Cells and Jurkat Leukemia Cells Biometals 20233663965510.1007/s 10534-022-00484-436626098 · doi ↗ · pubmed ↗