Cell Wall Proteome of Candida albicans Reveals Proteins Associated with Tolerance to Antibiofilm Activity of a Lippia graveolens Kunth Stem Extract
Alejandra García-Núñez, Ana Lilia Martínez-Rocha, Carlos Antonio Alba-Fierro, Francisco Javier Ríos-Fránquez, Juan Pablo Cabral-Miramontes, María Estela Frías-Zepeda, Miguel Ángel Escobedo-Bretado, Estela Ruiz-Baca

TL;DR
This study explores how a plant extract from Lippia graveolens affects Candida albicans biofilms and identifies cell wall proteins linked to resistance.
Contribution
The study identifies cell wall proteins associated with tolerance to a natural extract's antibiofilm activity in Candida albicans.
Findings
The Lippia graveolens extract inhibited biofilm formation in C. albicans by 77% at 1.6 mg/mL.
156 proteins were up-regulated and 94 down-regulated in the cell wall after extract exposure.
40 cell wall proteins were linked to biofilm formation, adhesion, and stress response.
Abstract
Candida albicans is an opportunistic fungus capable of forming biofilms that are resistant to conventional antifungal treatments. This study evaluated the antibiofilm activity of an ethanolic extract from Mexican oregano (Lippia graveolens Kunth) stem and its impact on the protein composition of the C. albicans cell wall (CW). The proteomic analysis was restricted to the adherent cells that persisted after treatment, representing the more tolerant subpopulation. First, the biofilm-forming capacity of 18 clinical isolates was characterized. Subsequently, the effect of the Lippia graveolens Kunth stem extract on biofilm formation was assessed in clinical isolates of C. albicans with high and moderate biofilm-forming capacities. The results showed an MIC90 of 16 mg/mL against C. albicans isolates in planktonic growth. Furthermore, the extract exhibited an antibiofilm effect, showing a 77%…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
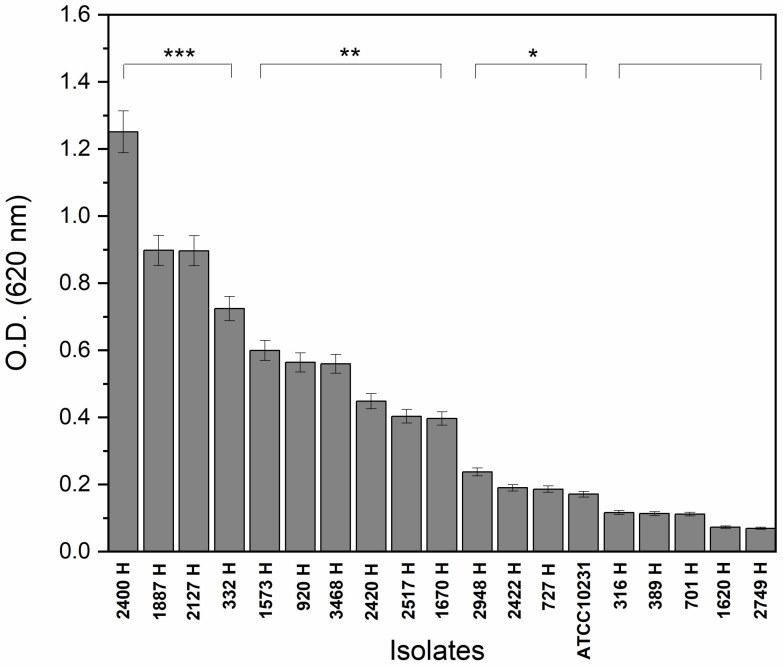 Figure 1
Figure 1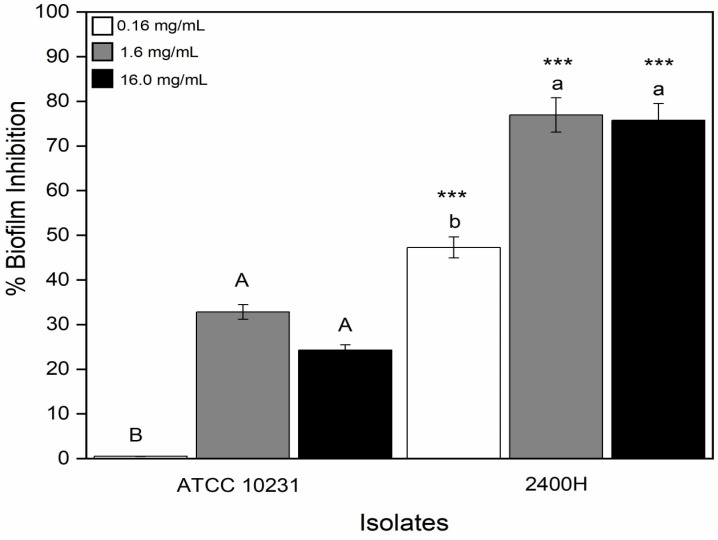 Figure 2
Figure 2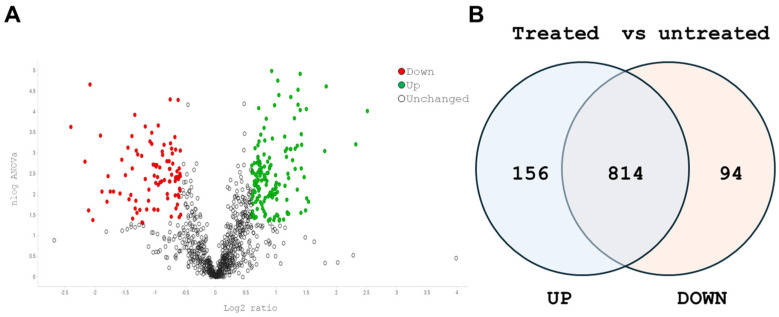 Figure 3
Figure 3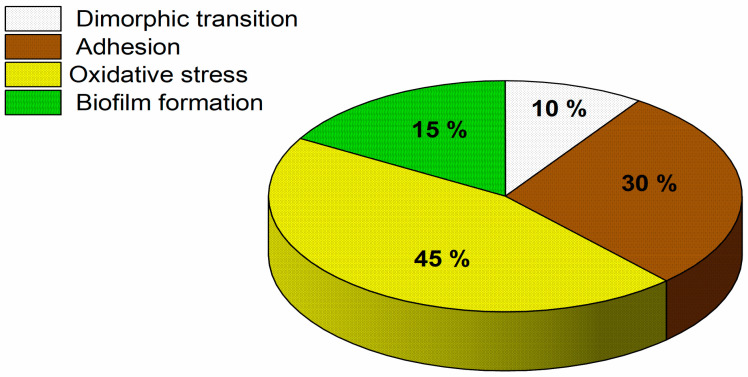 Figure 4
Figure 4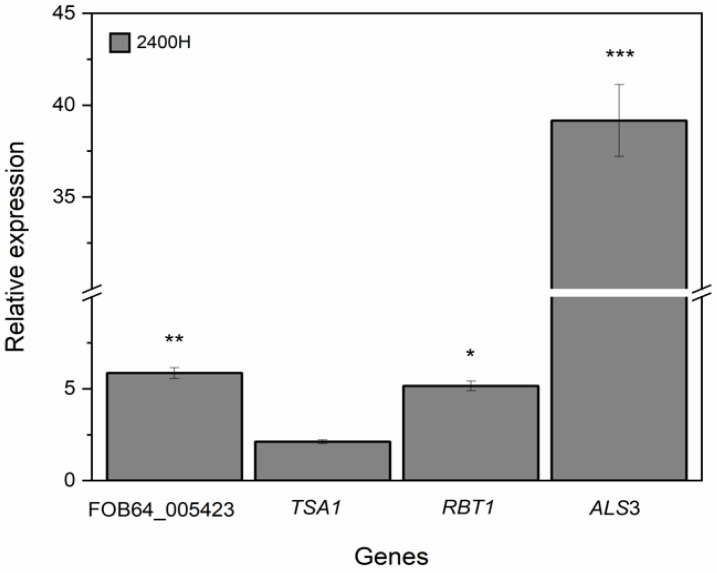 Figure 5
Figure 5- —Consejo Estatal de Ciencia y Tecnología del Estado de Durango (COCYTED)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Bacterial biofilms and quorum sensing · Legionella and Acanthamoeba research
1. Introduction
Candida albicans is a dimorphic pathogenic fungus commonly present on the surface of organs such as the mouth, skin, upper respiratory tract, intestines, vagina, etc. Candidiasis is considered the most common among fungal infections, mainly in hosts with an immunological disorder [1]. One of the most important virulence factors of C. albicans is its ability to produce biofilms, which confer resistance to hostile environments when exposed to the host’s immune response or during treatment with antifungal agents [2,3]. The biofilm is characterized by a compact community of different cell types, such as yeast, pseudohyphae, and hyphae, enclosed in an extracellular polymeric matrix, preventing or reducing drug diffusion [4]. As a worrying fact, most of the drugs used today are ineffective against C. albicans biofilms [5]; therefore, searching for new bioactive molecules with therapeutic applications against this fungus has become a relevant issue.
Plant extracts and their compounds are a valuable source of products with therapeutic potential, which can be used to treat diseases such as candidiasis [6]. The biologically active compounds include primary and secondary metabolites such as acetogenins, alkaloids, flavonoids, coumarins, lactones, anthraquinones, glycosides, tannins, and phytosterols [7,8]. The inhibitory effect on the growth and biofilm formation of C. albicans has already been demonstrated using oil extracts from plants from the Lippia genus such as Lippia junelliana, Lippia alba, Lippia salviaefolia, Lippia velutina, Lippia balansae, Lippia lasiocalycina, Lippia lupulina, Lippia citriodora, Lippia origanoides, etc. [9,10,11,12,13,14,15]. Oregano is an aromatic plant whose main species include Origanum vulgare, Lippia berlandieri Shauer, and L. graveolens Kunth, the latter commonly referred to as Mexican oregano [16]. The leaves of L. graveolens Kunth are widely used as a condiment in fresh and processed foods, as well as in the production of essential oils [17]. Secondary metabolites such as carvacrol and thymol, which are obtained from the essential oil, contain high levels of antimicrobial activity, with thymol being the most active [18]. However, although oregano essential oils obtained from leaves have shown their effectiveness in reducing the growth of C. albicans and its biofilm formation, they can show undesirable effects, such as cytotoxicity in human cell lines [19]. The stem of L. graveolens Kunth plant, a by-product of essential oil extraction, has no commercial value despite being rich in compounds such as caffeic acid and 2-hydroxybenzoic acid and flavonoids such as naringenin, taxifolin, eriodyctiol, acacetin, luteolin, quercetin 3-O-glycoside, apigenin, floridzin, and quercetin, which combined in an ethanolic extract, have been used by our research team, to demonstrate antifungal activity against Fusarium spp. [20]. Our research team also demonstrated that during murine acute toxicity measurements, even when administering L. graveolens Kunth stem extract concentrations as high as 2000 mg/Kg, the extract was classified in category 5 as nontoxic according to the guidelines set by the OECD-423 [21]. Furthermore, its potential use as a therapeutic agent against lung cancer, without affecting the viability of normal cells, was demonstrated using the same stem extract [22]. According to the latter and considering the cytotoxic issues that essential oils from leaves can present, the study of the effect of extracts from L. graveolens Kunth stem, and the resistant mechanism induced by its compounds aimed at combating fungal infections, becomes a relevant study.
In this study, we evaluated the effect of an ethanolic L. graveolens Kunth stem extract on both planktonic and biofilm-forming C. albicans cells. Furthermore, we assessed the modification in the cell wall (CW) proteome, focusing on the adherent subpopulation that persisted after treatment. This perspective adds originality to the study by examining the molecular features that support biofilm persistence and identify new therapeutic targets for developing antifungal strategies based on natural products in the face of increasing resistance and toxic effects associated with current antifungals.
2. Materials and Methods
2.1. Ethical Aspects
We used clinical C. albicans isolates (Table S1) provided by Hospital General 450 de Durango, Mexico. In all cases, the clinical isolates were obtained as part of routine diagnostic workup and used solely for research purposes without compromising the health or well-being of the donors. Since the samples were obtained as part of routine diagnostic procedures and did not contain any personal identifiable information, informed consent was not considered necessary by current regulations on microbiological research [23]. The Ethics Committee of Hospital General 450 de Durango reviewed and approved the protocol folio No. 34.
2.2. Collection of Plant Material
The plant material used in this study was the same as previously described [20]. Briefly, stems of Lippia graveolens Kunth were collected from the Oro Verde del Semidesierto Cooperative, Cuencamé, Durango. The specimen was recorded in the herbarium of the Interdisciplinary Research Center for Regional Development (CIIDIR-IPN), Durango, Mexico as Lippia graveolens Kunth (Vouchers 35542). Stems were ground to a particle size of 30 mesh with 8.5% moisture. The material was stored in sealed plastic bags and kept refrigerated at 4 °C until use.
2.3. Extract Preparation
The extracts used in this study were prepared according to the previously reported method [20]. Briefly, the stems of Lippia graveolens Kunth were macerated at room temperature (25 °C) with aqueous ethanol at concentrations of 80% and mass:solvent ratios of 1:30. The extraction was carried out for 24 h with stirring, followed by filtration, a second extraction with fresh solvent under the same conditions, and concentration using a rotary evaporator at 45 °C. The chemical composition of this extract has been previously characterized [20], revealing a profile enriched in naringenin (27%), taxofolin (21.1%), Caffeic acid (10.6%), eriodityol (18.1%), acacetin (0.7%), luteolin (8.6%), Quercetin 3-O-Glucoside (2.1%), Coumaric acid (3.8%), 2-hydroxybenzoic acid (1.7%), apigenin (1.5%), phrolidin (0.8%), quercetin (0.7%), Protocatechuic acid (0.9%), neohesperidin (0.7%), rutosidum (0.2%), quinic acid (0.3%), and 4-hydroxybenzoix acid (0.5%). The resulting aqueous extracts were lyophilized for subsequent use.
2.4. Strains and Culture Conditions
C. albicans isolates (Table S1) were routinely grown at 37 °C in a broth containing 1% yeast extract, 2% peptone, and 2% glucose (YPG). Eighteen strains of C. albicans from nosocomial candidemias used in this study correspond to clinical isolates kindly donated by the culture collection of Hospital General 450, Durango, Mexico. Species identification was confirmed by PCR amplification and sequencing of the internal transcribed spacer (ITS) region of the rDNA, according to the method described by White et al. [24]. Following molecular identification, strains were cryopreserved at −70 °C until use. The C. albicans ATCC 10231 strain was used as the reference strain.
2.5. Biofilm Formation Capacity of Clinical Isolates of C. albicans
The 18 C. albicans clinical isolates and the strain ATCC 10231 were grown in YPG medium at 37 °C and 120 rpm for 18 h. After this time, suspensions (10^6^ cells/mL) were prepared in RPMI 1640 medium and incubated in 96-well plates at 37 °C for 48 h. Subsequently, the wells were washed with phosphate buffer (0.1 M Na_2_HPO_3_•7H_2_O and NaH_2_PO_3_ at pH 7.2), stained with 0.4% crystal violet for 45 min, and washed with sterile distilled water. Subsequently, we added 95% ethanol to dry, and 100 μL of each sample was transferred to new plates to measure the optical density at 595 nm [25]. Biofilm formation was classified into four categories according to the optical density cutoff (ODc), calculated as the mean of the negative controls containing only RPMI 1640 medium (ODnc) plus three standard deviations (3 × SDnc). The categories were: negative biofilm (OD ≤ ODc), slightly positive (ODc < OD ≤ 2 × ODc), moderately positive (2 × ODc < OD ≤ 4 × ODc), and intensely positive (OD > 4 × ODc) [26]. Based on the above, C. albicans isolates were classified as high, moderate, low, or non-biofilm formers.
2.6. Evaluation of the Effect of L. graveolens Kunth Stem Extract on the Planktonic Growth of C. albicans
The C. albicans isolates were cultured independently in YPG medium at 37 °C, 120 rpm for 18 h. We then prepared suspensions of the C. albicans isolates using RPMI 1640 medium at a concentration of 10^3^ cells/mL. One hundred μL of each suspension was added to microtiter plate wells containing RPMI medium and extract, reaching a final volume of 200 μL in each well at L. graveolens Kunth stem extract concentrations of 2, 4, 8, 16, 32, and 64 mg/mL. Negative controls (0 mg/mL) were prepared using the same procedure, replacing L. graveolens Kunth stem extract with distilled water. Plates inoculated with the C. albicans isolates containing the different concentrations of the extract were incubated at 37 °C for 24 h. Due to the intense intrinsic coloration of the extract at higher concentrations, growth inhibition was subsequently evaluated visually. Fluconazole was used as a positive control according to CLSI guidelines [26].
2.7. Evaluation of the Effect of L. graveolens Kunth Stem Extract on Biofilm Formation in C. albicans Isolates
Biofilm-forming C. albicans isolate 2400H and strain ATCC 10231 were grown in YPG medium at 37 °C and 120 rpm for 18 h. Subsequently, we prepared suspensions in RPMI 1640 medium at a concentration of 10^6^ cells/mL. We added 100 μL of the suspensions to wells containing RPMI 1640 medium and L. graveolens Kunth stem extract, kindly provided by Cabral-Miramontes et al. [20], to a total volume of 200 μL and final extract concentrations of 0.16 μg/μL, 1.6 μg/ μL, and 16 μg/μL. After 24 h of incubation at 37 °C, non-adherent cells were removed by washing with phosphate buffer. Adherent cells were then mechanically scraped from each well and resuspended in phosphate buffer, and the optical density was measured at 600 nm. Negative controls were prepared using the same procedure, replacing L. graveolens Kunth stem extract with distilled water. The biofilm inhibition percentage was calculated according to the formula:
2.8. Evaluation of the Effect of L. graveolens Kunth Stem Extract on Cell Wall Proteins of C. albicans 2400H Isolate
2.8.1. Cell Lysis and Protein Extraction
Cell lysis of the highly biofilm-forming C. albicans 2400H isolate, in interaction with and without the extract, was carried out according to the method reported by [27]. After 24 h, the adherent cells were washed three times with PBS, scraped, transferred to an Eppendorf tube, and centrifuged at 3500 rpm for 5 min. The pellet cells were lysed using a homogenizer containing wash buffer (50 mM Tris-HCl, pH 7.5, supplemented with 1.0 mM phenylmethylsulfonyl fluoride (PMFS)) with SDS 0.5% and glass beads, performing five cycles of 1 min alternating with ice. The homogenate was centrifuged at 3500 rpm for 10 min at 4 °C, and the pellet with the CWPs was washed five times with wash buffer. Subsequently, an extraction buffer containing β-mercaptoethanol and 2% SDS was added according to the method reported by [28], and kept boiling for 10 min. After that, the homogenate was centrifuged at 3500 rpm for 15 min at 4 °C. The proteins in the supernatant were precipitated using 70% ethanol (1:3 v/v) and frozen at −20 °C for two hours. Finally, the proteins were centrifuged at 10,000 rpm for 15 min at 4 °C and resuspended in 100 μL of distilled water. We used the Bradford method to determine the concentration of the proteins extracted from C. albicans [29].
2.8.2. Sample Preparation and Liquid Chromatography Coupled to Mass Spectrometry (LC-MS) Analysis
CWP’s extracted from biofilms of the clinical isolate 2400 H grown in the presence or absence of the extract were digested with 50 µg of total protein by Filter-Aided Sample Preparation (FASP) [30] with UA solution (8 M urea in 0.1 M Tris/HCl, pH 8) on passivated VivaSpin^®^ 500 filters (Cytiva, Marlborough, MA, USA) [31]. BSA was added to the concentrated eluates (Waters Corporation, Milford, MA, USA) as an internal standard, achieving 25 fmol/µL in 40 µL. We used mass spectrometry, applying the LC-MS analytical method described by [32] with some modifications. Chromatographic conditions included the use of an ACQUITY M-Class UPLC system, with mobile phase A (0.1% formic acid in water) and mobile phase B (0.1% formic acid in acetonitrile) under the following gradient: 7% B (0 min), 40% B (121.49 min), 85% B (123.15–126.46 min) and 7% B (129–130 min), at a constant flow rate of 400 L/min and a temperature of 45 °C. Spectral data were collected using a Synapt G2-Si mass spectrometer (Waters, Milford, MA, USA), using nanoelectrospray ionization (nanoESI) and ion mobility separation (IM-MS). Analysis was performed with a data-independent acquisition (DIA) approach through high-definition multiplexed MS/MS (HDMS^E^) mode. The ionization source parameters included 2.75 kV at the sampling capillary, 30 V at the cone and source offset, 70 °C at the source, 0.5 bar of nanoflow gas, and 150 L/h of purge gas. Chromatograms were recorded in positive mode within the m/z range 50–2000 with a scan time of 500 ms. No collision energy was applied for low-energy chromatograms, while high-energy chromatograms used a collision energy ramp of 19–55 eV to fragment precursor ions in the transfer cell.
2.8.3. Protein Database Search and Protein Quantification
The generated MS and MS/MS spectra contained in the raw files were processed, compared, and quantified using Progenesis QI for Proteomics v4.2 software (Nonlinear Dynamics, Newcastle, UK) [33], employing an inverted Candida albicans proteome (downloaded from UniProt, UP000005640) and the bovine serum albumin (BSA) fasta database (accession P02769). Parameters included trypsin as the cleavage enzyme with one missed cleavage, carbamidomethyl (C) as a fixed modification, and amidation (N-terminal), deamidation (N, Q), oxidation (M), and phosphorylation (S, T, Y) as variable modifications. The quantification was set by automatic tolerances for peptides and fragments, with the following minimum values: 2 fragments per peptide, five fragments per protein, one peptide per protein, and a false discovery rate (FDR) ≤ 4%. Absolute quantification was conducted using the three most reliable peptides per protein (Top 3), following the method of Silva et al. [34]. All identifications reached a confidence percentage ≥ 95% (Protein AutoCurate green). Calibration of the Synapt G2-Si spectrometer was carried out with [Glu1]-Fibrinopeptide ([M + 2H]^2+^ = 785.84261) with an accuracy ≤ 1 ppm.
Absolute protein quantification was performed according to the Top3 method reported by Arnaud-Franco et al. [35] following the next equation:
where the Top3 value is the mean of the MS signal response of the three most intense tryptic peptides. Using the Top3 value of the internal standard as well as of interest proteins, it is possible to determine the amount of protein in each sample using the following equation [33,34]:
where A is the protein of interest, B is the internal standard (BSA), and “fmol_B_” is the amount injected of internal standard.
The ratio was calculated by dividing the fmol value of each characterized protein in cells treated with the extract (treatment) by the corresponding value in untreated cells (control). All proteins considered differentially expressed had at least a ratio of ±0.585 (expressed as a base 2 logarithm) and a p-value ≤ 0.05.
For processing, we used the Candida albicans *.fasta database (from UniProt, UP000005640, with 79,052 protein sequences) supplemented with BSA (downloaded from UniProt, accession number: P02769). Bioinformatics analysis of total proteins and DEPs was performed using STRING-DB v12.0 (https://string-db.org/; accessed on 3 August, 2023). The resulting functional enrichments are biological process (gene ontology), cellular component (gene ontology), KEGG pathways, and subcellular localization (compartments).
2.9. Evaluation of the Effect of the Stem Extract on the Expression Profile of the FOB64_005423, ALS3, TSA1, and RBT1 Genes
Total RNA was extracted using the Trizol method from biofilms of C. albicans 2400H, with and without exposure to L. graveolens Kunth stem extract, after 24 h at 37 °C in liquid nitrogen. RNA was then treated with RNase-free DNase I (Thermo Fisher Scientific, Carlsbad, CA, USA), and ImProm-II^TM^ was used for cDNA synthesis following the manufacturer’s instructions. RT-qPCR reactions were developed with the next gene-specific primers FOB64_005423: Fwd 5′-TGCTGGTGGTGTTAATGGTG-3′ and Rev 5′-AGCTAAAGCCAAGACTGAGG-3′, RBT1: Fwd 5′-CAAAAACTAGTGCTCTCGTCT-3′ and Rev 5′-AGACCAATAATAGCAGCACCA-3′, TSA1: Fwd 5′-CTTGAGATTGTTGGAGGCTTT-3′ and Rev 5′-AGTATTCCTTGGATGCTTCTG-3′, ALS3: Fwd 5′-CAACCAATCTCAATCGCAATC-3′ and Rev 5’-ATCAAACCACATAACCAAGTAG-3′, using 100 ng of cDNA as template and the GoTaq^®^ qPCR master mix kit (Thermofisher). The RDN5.8 gene, a ribosomal protein [36], was used as a housekeeping gene. We used the 2^−ΔΔCt^ method to analyze the data using StepOne software v2.3 (Thermo Fisher Scientific, Waltham, MA. USA) to determine the relative expression of the selected genes.
2.10. Statistical Analysis
A completely randomized design was used and analyzed using a one-way analysis of variance (ANOVA) with a post hoc Bonferroni-Holm test. Graphs were created using OriginLab 2024^®^ software. Significance levels were defined as ns (p > 0.05), * (p ≤ 0.01), ** (p ≤ 0.001), and *** (p ≤ 0.0001) compared to the control groups. All experiments were performed in triplicate using two independent biological replicates, and the data were presented as mean ± standard deviation.
3. Results
3.1. Biofilm Formation Capacity in C. albicans Isolates
To determine the biofilm formation capacity of the 18 C. albicans isolates, we used a spectrophotometric method (crystal violet). The optical density (OD) results are shown in Figure 1, wherein isolates 2400H and 2749H presented the highest and lowest biofilm formation capacity, respectively.
The C. albicans 2400H, 1887H, 2127H, 332H, 1573H, 920H, 3468H, 2420H, 2517H, and 1670H isolates presented significant differences compared to the reference strain ATCC 10231 used as the control. The isolates 2400H, 1887H, 2127H, and 332H showed more than 4-fold the OD than the control, the1573H, 920H, 3468H, 2420H, 2517H, and 1670H isolates showed an OD between 2 and 4-fold the OD than the control, while isolates 2948H, 2422H, 727H and the reference strain ATCC 10231 showed an OD higher than the control but lower than 2-fold. The isolates 316H, 389H, 701H, 1620H, and 2749H showed an OD mean value lower than or equal to that of control strain ATCC 10231.
Based on the classification criteria, from the 18 evaluated isolates of C. albicans clinical samples, 22.2% were high biofilm-forming (2400H, 1887H, 2127H, and 332H), 33.3% were moderate biofilm-forming (1573H, 920H, 3468H, 2420H, 2517H and 1670H), 16.6% low biofilm-forming (2948H, 2422H and 727H) and 27.9% non-biofilm-forming (316H, 389H, 701H, 1620H and 2749H) (Table 1). The reference strain ATCC 10231 was classified as a low biofilm-forming strain. Further experiments considered only high and moderate biofilm-forming C. albicans isolates from clinical origin and the reference strain ATCC 10231.
3.2. Effect of L. graveolens Kunth Stem Extract on the Planktonic Growth of Isolates of C. albicans
The biofilm formation capacity was evaluated in C. albicans isolates, and the L. graveolens Kunth 90% minimal inhibitory concentration (MIC_90_) was determined only in high (2400H, 1887H, 2127H, and 332H) and moderate (1573H, 920H, 3468H, 2420H, 2517H, and 1670H) biofilm-forming C. albicans isolates and the reference strain ATCC 10231. Table 2 shows that all the evaluated isolates exhibited an L. graveolens Kunth extract MIC_90_ of 16 mg/mL when incubated in RPMI media during planktonic growth. As a control, fluconazole was tested, and all isolates were susceptible.
3.3. Effect of L. graveolens Kunth Stem Extract on Biofilm Formation in C. albicans 2400H Isolate
To evaluate the effect of L. graveolens Kunth extract on biofilm formation, 2400H isolate and ATCC10231 strain were challenged with L. graveolens Kunth extract concentrations of 0.16 mg/mL, 1.6 mg/mL, and 16 mg/mL. The biofilm inhibition percentages are shown in Figure 2. At extract concentrations of 0.16 mg/mL, 1.6 mg/mL, and 16 mg/mL, 2400H isolate showed inhibition percentages of 47%, 77%, and 76%, respectively. ATCC10231 strain showed inhibition percentages of 0%, 33%, and 25% using the same concentrations. There were significant statistical differences in biofilm inhibition percentages between 2400H isolate and ATCC 10231 strain in all the tested concentrations. However, there were no statistical differences in biofilm inhibition percentages when comparing 1.6 mg/mL and 16 mg/mL concentrations in both strains, but there were when comparing 0.16 mg/mL concentration with 1.6 mg/mL and 16 mg/mL. The effect of the L. graveolens Kunth extract at 16 mg/mL was also evaluated in other biofilm-forming isolates. The results showed biofilm inhibition ranging from 18% to 77% (Figure S1).
3.4. Effect of L. graveolens Kunth Stem Extract in Cell Wall Proteins of C. albicans 2400H Isolate
To identify CWPs differentially produced in the cells that remained adherent after extract exposure, we analyzed the protein profile of the high biofilm-forming 2400H strain by comparing untreated and treated cells. The LC-MS analysis showed 1064 total proteins (Table S1) in 2400H isolate. Based on statistical significance (p < 0.05) and 0.5-fold higher or lower expression compared to control criteria, 156 and 94 proteins were up- and downregulated, respectively. As shown in Figure 3, the Volcano plot highlights proteins that were significantly up- or downregulated (Figure 3A), while the Venn diagram summarizes the total number of proteins identified, categorizing them as upregulated, downregulated, or unchanged compared to the untreated control (Figure 3B). Diverse subcellular localizations were identified in differentially expressed proteins, mainly CW-associated proteins (Figure S2). These results provide an overview of the proteomic response of the CW to the extract.
A total of 40 CWPs differentially expressed were identified; the proteins were classified into dimorphic transition, adhesion, oxidative stress, and biofilm formation functions (Figure 4). The highest percentage was represented by oxidative stress proteins (45%), while dimorphic transition proteins represented the lowest (10%).
From the 40 CWPs identified, 25 proteins were up-regulated and 15 were down-regulated (Table 3). The GPI-anchored cell wall protein RBT1, the agglutinin-like protein 3 (ALS3), the Flocculin type 3 repeat family protein (FOB64_005423), and the thioredoxin peroxidase (TSA1) were relevant up-regulated proteins implicated in the different processes.
The up-regulated CWP protein RBT1, agglutinin-like protein 3 (Als3), flocculin type 3 repeat family protein (FOB64_005423), and thioredoxin peroxidase (Tsa1) have already been reported in previous research works as relevant proteins associated with dimorphic transition, adhesion, oxidative stress, and biofilm formation functions (Table 4).
3.5. Expression Levels of FOB64_005423, RBT1, ALS3, and TSA1 Genes of C. albicans 2400H Isolate Exposed to L. graveolens Kunth Stem Extract
To validate the obtained proteomic data of the 2400H isolate, we evaluated the expression levels using RT-qPCR on FOB64_005423, RBT1, ALS3, and TSA1 genes coding for proteins reported in Table 4. Using the RDN5.8 as a housekeeping gene and the untreated cells as a control, the results showed that, L. graveolens Kunth stem extract positively regulated FOB64_005423, RBT1, ALS3, and TSA1 genes in C. albicans 2400H isolate (Figure 5). The RBT1 gene showed a relative expression of 6-fold higher than the control, while TSA1 showed a relative expression of 2.5-fold higher than the control. Notably, the gene ALS3 showed a relative expression 40-fold higher than the control.
4. Discussion
The results obtained in this research demonstrated that biofilm formation capacity among C. albicans clinical isolates differs from that of the reference strain ATCC 10231. The assay showed that after 24 h, ten C. albicans clinical isolates (2400H, 1887H, 2127H, 332H, 1573H, 920H, 3468H, 2420H, 2517H, and 1670H) exhibited higher biofilm production, and five (316H, 389H, 701H, 1620H, and 2749H) produced lower amounts of biofilm than the reference strain ATCC 10231. The latter results are consistent with studies where C. albicans isolates from candidiasis patients displayed a higher biofilm formation capacity compared to the reference strain ATCC 10231 [42]. A possible explanation for such results is that clinical experience has demonstrated that clinical isolates from fungal infections are more virulent and resistant to harsh conditions than collection strains. Previous works have found that C. albicans isolated from clinical mastitis exhibited a higher MIC_80_ than the reference strain ATCC 10231 when treated with Floribundum mundy [47].
The characterization of L. graveolens Kunth extracts has been reported in previous works, finding flavonoids such as naringenin, eriodyctiol, taxifolin, luteolin, and quercetin, and phenolic compounds such as hydroxyquinones [17,20,21]. Other research studies also reported the presence of different compounds, such as saponins and tannins [48]. The present work evaluated the effect of L. graveolens Kunth in fungal planktonic growth and biofilm formation, showing that all evaluated C. albicans isolates exhibited a MIC_90_ of 16 mg/mL. Cordoba et al. [9] determined the effect of an ethanolic extract of L. junelliana against strains of Candida krusei, C. albicans, Candida glabrata, and Candida parapsilosis, finding MIC’S_90_ of 0.00312 mg/mL, 0.40 mg/mL, 0.80 mg/mL, and 0.00312 mg/mL, respectively. Ruiz-Durán et al. [10] evaluated the antifungal activity of L. alba and L. origanoides extracts against C. albicans, C. parapsilosis and C. auris, reporting how L. origanoides extracts showed off MIC’S_50_ of 0.188 mg/mL, 0.141 mg/mL and 0.141 mg/mL for C. albicans, C. parapsilosis, and C. auris, respectively. They also reported a minimal inhibitory biofilm concentration (MIBC) of 0.188 mg/mL for C. albicans and C. parapsilosis, and 0.141 mg/mL for C. auris. More research works with different Lippia genus species as L. citriodora, L. salviaefolia, L. velutina, L. balansae, L. lasiocalycina and L. lupulina, have demonstrated antifungal effect against C. parapsilosis, C. albicans, C. krusei, and Cryptococcus neoformans, reporting MIC’S among 0.156 mg/mL and 0.250 mg/mL [11,13].
In our study, despite concentrations of L. graveolens Kunth extract being higher than MICs_90_ reported in extracts from other Lippia species, it must be considered that the extract used in this work was obtained from the plant stem, a discarded by-product of the essential oil industry, with a lower content of bioactive compounds. Furthermore, in order to increase the antifungal effect of the L. graveolens Kunth stem extract, future research related to bioformulations through emulsions or nanoemulsions could be considered, since it has been demonstrated that these formulations could increase the bioactivity of Lippia genus species extracts [12,49,50].
Some of the works mentioned above have reported the effect of natural extracts on germinal tube formation and yeast-to-hyphae transition. After adhesion, the next steps in biofilm production are germinal tube formation and hyphal elongation. In our work, compared to controls, the L. graveolens Kunth stem extract concentrations reported in this study reduced biofilm formation between 47% and 77% in the C. albicans isolate 2400H. Notably, a similar inhibitory effect was observed even at 1.6 mg/mL, a concentration tenfold lower than 16 mg/mL, which may be considered physiologically and therapeutically manageable [21,22]. At the MIC_90_, the extract completely inhibited planktonic cells but only reduced biofilms by 77%, reflecting the higher tolerance of biofilm-associated cells due to protective factors such as the extracellular matrix and reduced compound penetration [51].
A recent work [40], using 28 oleic acid-like saponins extracted from Pulsatilla chinensis, reported that C. albicans diminished its adhesion capacity and biofilm formation in the presence of concentrations between 6.25 µg/mL and 100 µg/mL of the saponin pulchinenoside B3, also reducing the expression of genes associated with yeast-to-hyphae transition. The diminishment of biofilm formation and preformed biofilm degradation has also been reported using extracts from Mentha species, showing a negative regulation in genes associated with adhesion [42]. Kim et al. [39] reported a decay in the expression levels of genes associated with adhesion and biofilm formation in C. albicans using hydroxyquinones, compounds contained in extracts of plants such as L. graveolens Kunth. A L. origanoides leaf extract has also demonstrated its effectiveness not only in C. albicans biofilm formation diminishment but also in the reduction in biofilm formation of different microorganisms such as Lactobacillus rhamnosus and Streptococcus mutans, which are associated with caries formation. However, despite the effectiveness of inhibiting biofilm formation, the extract obtained from the leaves of this plant showed cytotoxic levels similar to the compound chlorhexidine, a compound that causes undesirable effects during its prolonged use [19].
To better understand the effect induced by L. graveolens Kunth stem extract over C. albicans and the possible response mechanism of the fungus to this challenge, a proteomic LC-MS analysis was carried out in an enriched fraction from the CW obtained from the treatment-tolerant adherent cells. Through this analysis, a total of 250 CWPs were identified, but only 40 were differentially expressed. The differentially expressed proteins were classified into four functions: dimorphic transition, adhesion, oxidative stress, and biofilm formation. Different research works have evaluated through proteomic approaches the effect of plant extracts such as Cytrus hystrix and Murraya koenigii over C. albicans, finding a differential production of proteins associated with diverse functions such as mitochondrial matrix, cellular and metabolic processes, transition, antioxidant activities, adhesion, filamentous growth, etc. [38,52]. In the same sense, carrying out a proteomic study through LC-MS/MS to evaluate the effect in a whole population of C. albicans (adherent and non-adherent cells) exposed to β-citronellol, a monoterpene naturally found in the plant Citrus hystrix, Buakaew et al. [38] found that from the main 126 proteins identified, altered in response to β-citronellol, the CWPs Als2p, Rbt1p, and Pga4 and the oxidative stress response proteins Sod1p, Gst2p, and Ddr48p, contrary to our work, were downregulated. Muthamil et al. [52], working with loosely adherent planktonic cells, reported a possible mechanism of oleic acid obtained from the plant Murraya koenigii against C. albicans. Using a MALDI-TOF/TOF proteomic analysis using differentially expressed 2D SDS-PAGE spots, they established that oleic acid inhibited proteins associated with yeast-to-hyphae transition, biofilm formation, and fungal adhesion capacity, and modified the amount of ergosterol in the inner cell membrane. Later results were complemented through RT-qPCR, confirming that the genes that code for these proteins were negatively regulated. The results in the present work demonstrated a higher production of cell surface proteins associated with adhesion, oxidative stress, dimorphic transition, and biofilm formation coded by FOB64_005423, TSA1, RBT1, and ALS3 genes. The same results were further obtained by RT-qPCR, confirming opposite results to previous works [38,39,48,52]. The differences in our results compared to previous works were expected, since our proteomic analysis specifically targeted C. albicans cells that remained adherent after 24 h of exposure to the L. graveolens Kunth extract, allowing the identification of wall-associated proteins potentially involved in biofilm persistence, discarding non-adherent cells or susceptible subpopulations. However, analysis of the fraction that remained adherent allows the identification of determinants associated with tolerance and biofilm persistence. The overexpression of FOB64_005423, TSA1, RBT1, and ALS3 genes could be a compensatory stress response mechanism in the C. albicans 2400H isolate, induced by the extract, since dimorphic fungi have shown adaptation to changes in growth conditions and external factors to remodel or repair their surface, in response to environmental stress using different stress response pathways [53,54]. However, this possible mechanism needs to be confirmed with further experiments. Future studies should consider separate or comparative proteomic analyses of both adherent and non-adherent C. albicans cells after treatment, to capture the full spectrum of cellular responses to the extract. Moreover, the present findings may inform subsequent efforts aimed at identifying specific inhibitors or therapeutic combinations that more effectively target biofilm persistence.
While the present study provides molecular and proteomic evidence supporting the antibiofilm activity of the L. graveolens stem extract and its impact on CW–associated proteins, future studies could further expand these findings by incorporating complementary approaches. In particular, metabolic activity assays and advanced microscopic analyses (e.g., confocal or electron microscopy) would be valuable to correlate the observed proteomic changes with biofilm viability, architecture, and structural organization. The integration of these methodologies may offer additional insights into the phenotypic consequences of extract exposure and contribute to a more comprehensive understanding of its effects on C. albicans biofilms.
5. Conclusions
The results demonstrate that the ethanolic L. graveolens Kunth stem extract exhibits significant antifungal activity against C. albicans in both its planktonic and biofilm forms. Furthermore, these findings emphasize the potential of L. graveolens Kunth stem as an alternative source of bioactive compounds with antifungal activity, supporting their relevance in strategies aimed at controlling biofilm formation. The CW proteome revealed proteins associated with fungal dimorphic transition, adhesion, oxidative stress, and biofilm formation. These findings shed light on key tolerance mechanisms in adherent biofilm-forming cells.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Pu S. Niu S. Zhang C. Xu X. Qin M. Huang S. Zhang L. Epidemiology, antifungal susceptibilities, and risk factors for invasive candidiasis from 2011 to 2013 in a teaching hospital in southwest China J. Microbiol. Immunol. Infect.2017509710310.1016/j.jmii.2015.01.00525769664 · doi ↗ · pubmed ↗
- 2Cavalheiro M. Teixeira M.C. Candida biofilms: Threats, challenges, and promising strategies Front. Med.201852810.3389/fmed.2018.00028 PMC 581678529487851 · doi ↗ · pubmed ↗
- 3Finkel J.S. Mitchell A.P. Genetic control of Candida albicans biofilm development Nat. Rev. Microbiol.2011910911810.1038/nrmicro 247521189476 PMC 3891587 · doi ↗ · pubmed ↗
- 4Pereira R. dos Santos-Fontenelle R.O. de Brito E.H.S. de Morais S.M. Biofilm of Candida albicans: Formation, regulation and resistance J. Appl. Microbiol.2021131112210.1111/jam.1494933249681 · doi ↗ · pubmed ↗
- 5Nobile C.J. Johnson A.D. Candida albicans biofilms and human disease Annu. Rev. Microbiol.201569719210.1146/annurev-micro-091014-10433026488273 PMC 4930275 · doi ↗ · pubmed ↗
- 6Ji H.F. Li X.J. Zhang H.Y. Natural products and drug discovery EMBO Rep.20091019420010.1038/embor.2009.1219229284 PMC 2658564 · doi ↗ · pubmed ↗
- 7Abdul-Wahab S.M. Jantan I. Haque M.A. Arshad L. Exploring the leaves of Annona muricata L. as a source of potential anti-inflammatory and anticancer agents Front. Pharmacol.2018966110.3389/fphar.2018.0066129973884 PMC 6019487 · doi ↗ · pubmed ↗
- 8Hadisaputri Y.E. Habibah U. Abdullah F.F. Halimah E. Mutakin M. Megantara S. Diantini A. Antiproliferation activity and apoptotic mechanism of soursop (Annona muricata L.) leaves extract and fractions on MCF 7 breast cancer cells BCTT 20211344745710.2147/BCTT.S 31768234295188 PMC 8291383 · doi ↗ · pubmed ↗