Steering of Crystal Cell Volumes in Apatites and Bioapatites
Andrzej Kuczumow, Agnieszka Lasota, Mieczysław Gorzelak, Paulina Wojtyła-Buciora, Przemysław Biliński, Małgorzata Bernatek, Karolina Turżańska, Jan Olszewski, Przemysław Dyndor, Maciej Jarzębski, Marek Wieruszewski, Mirosław Jabłoński

TL;DR
This paper explores how different organisms use specific apatite crystal structures in their hard tissues, and how these structures can be influenced by environmental factors.
Contribution
The paper introduces a reformulated version of Bragg’s law to classify apatite biomaterials and steer crystal cell volumes.
Findings
Organisms select specific apatites for hard tissues based on environmental adaptation.
Volume expansion in apatites is linearly coupled with the expansion of Bragg’s d parameter.
Different modes of change in Bragg’s dimension d were identified and quantified.
Abstract
The biological variability of apatites in different hard tissues of organisms was the starting point for this investigation. Materials such as whale rostrums, ganoine, and some fish bones were analyzed. It has been proven that different organisms select specific kinds of apatites for the construction of their hard organs at the level of the crystal cell. This probably results from the long-lasting adaptation of the construction to their environmental needs. The materials are characterized by the parameters Δd and ΔE, being the real and apparent deviations from Bragg’s dimension d and the energy of excitation in XRD—E. This study is based on previously published, verified results from a number of researchers and research groups. The derivation of expressions was possible due to an original approach to Bragg’s equation, finally finished in the reformulation of the law, which describes the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
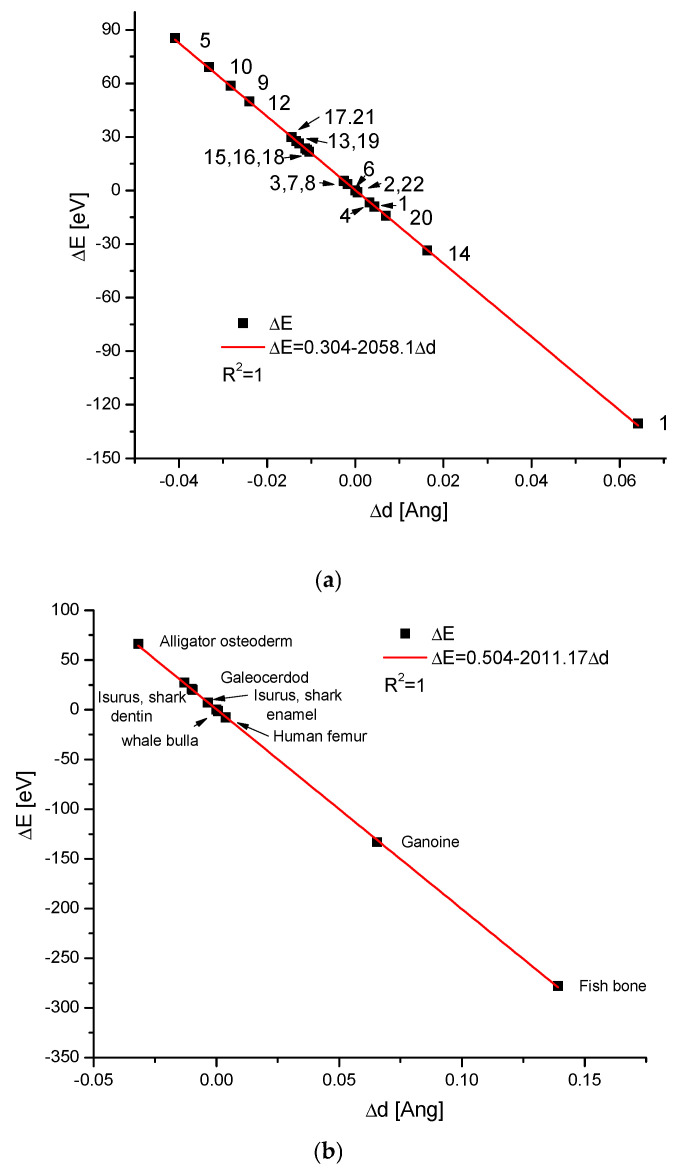 Figure 1
Figure 1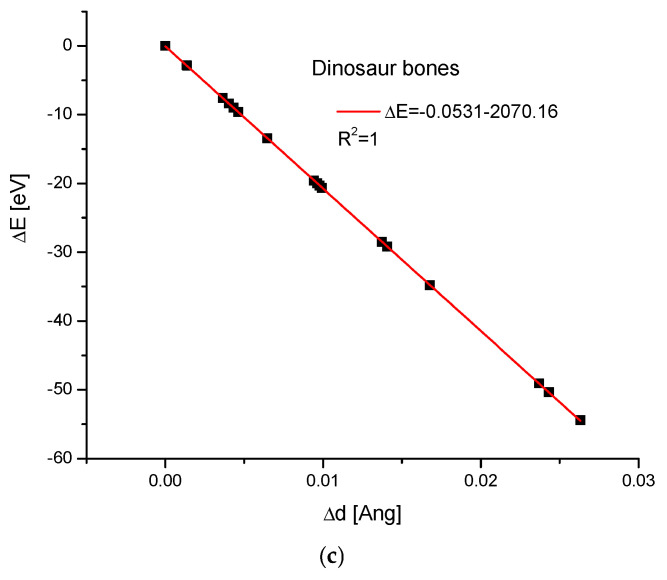 Figure 2
Figure 2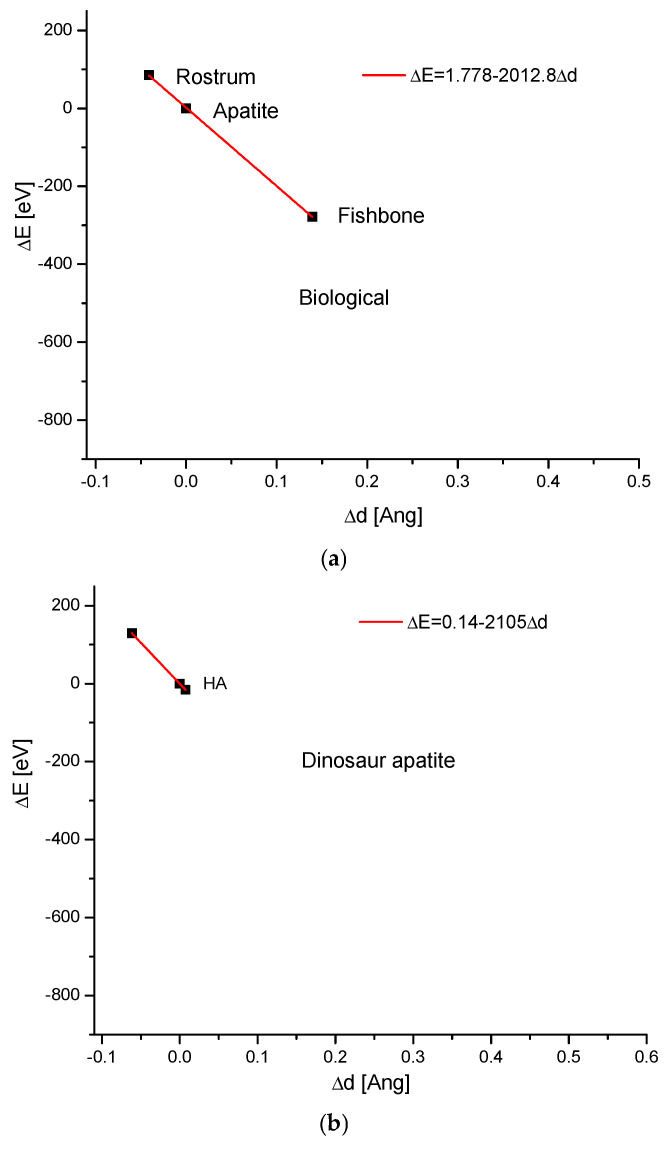 Figure 3
Figure 3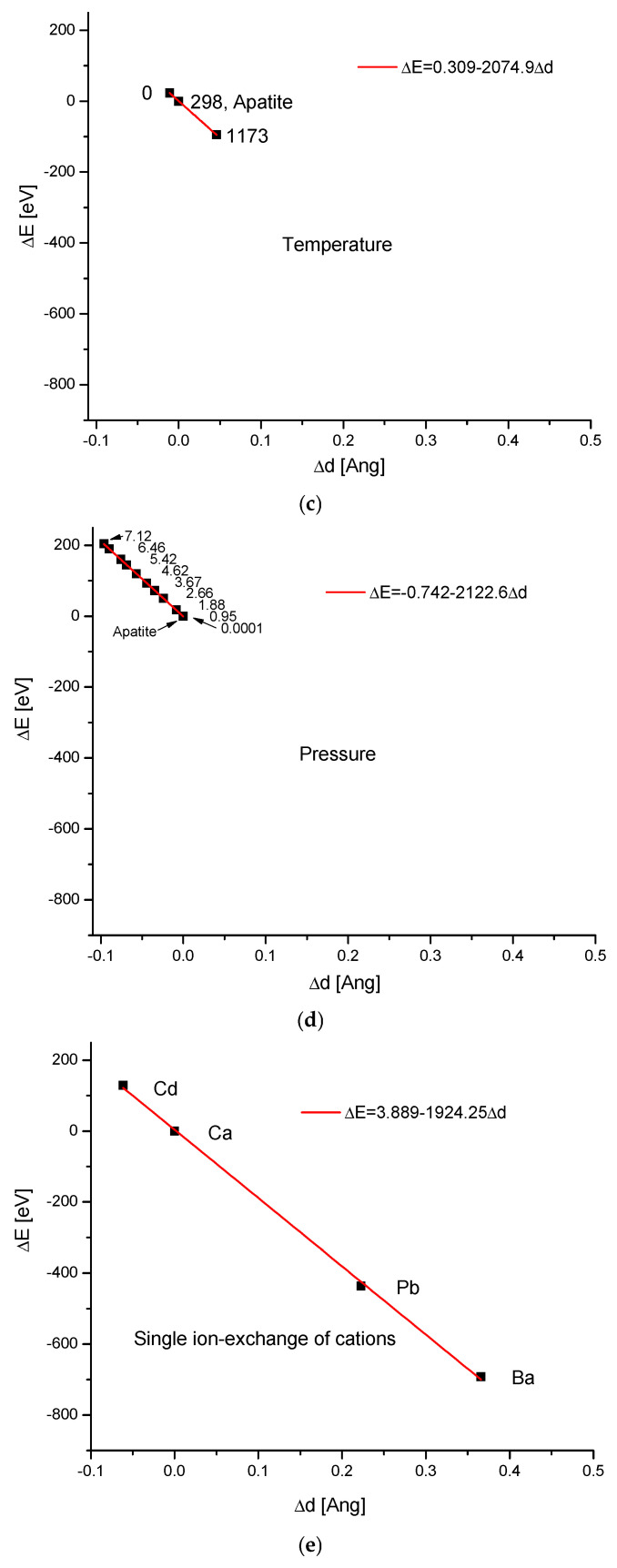 Figure 4
Figure 4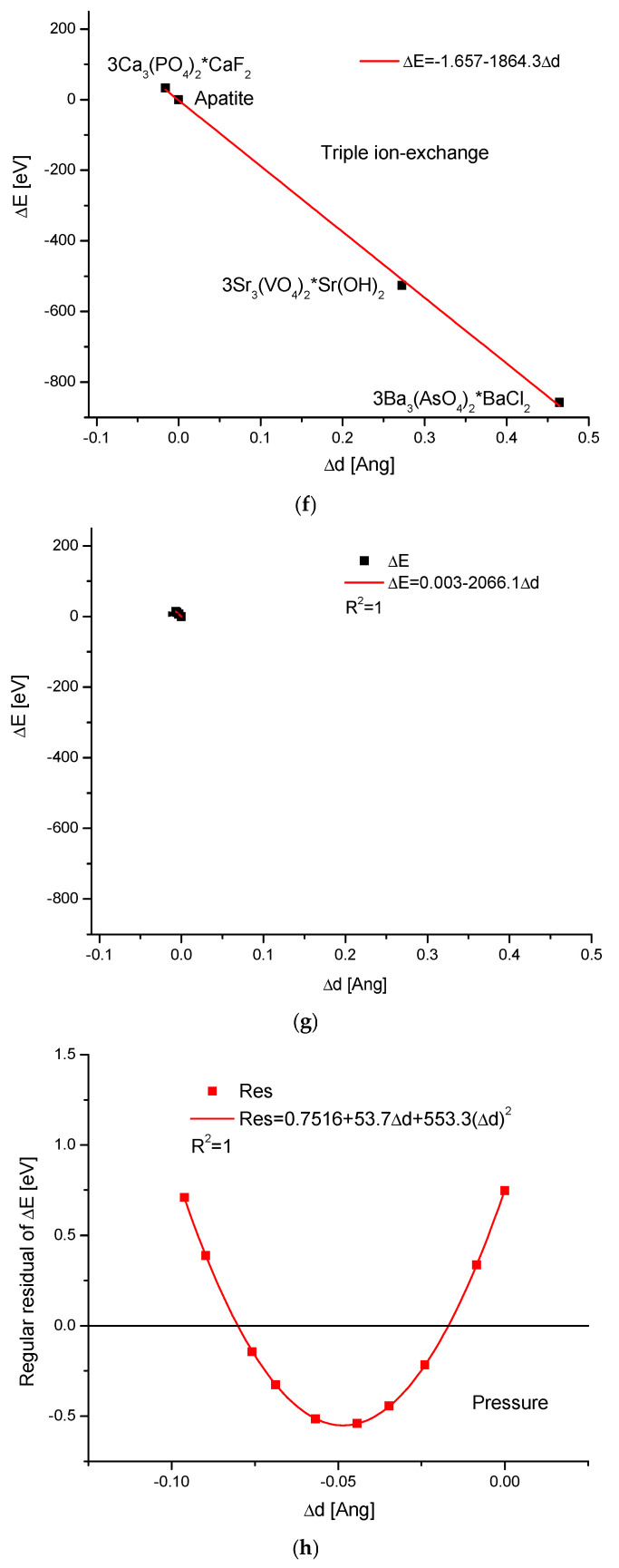 Figure 5
Figure 5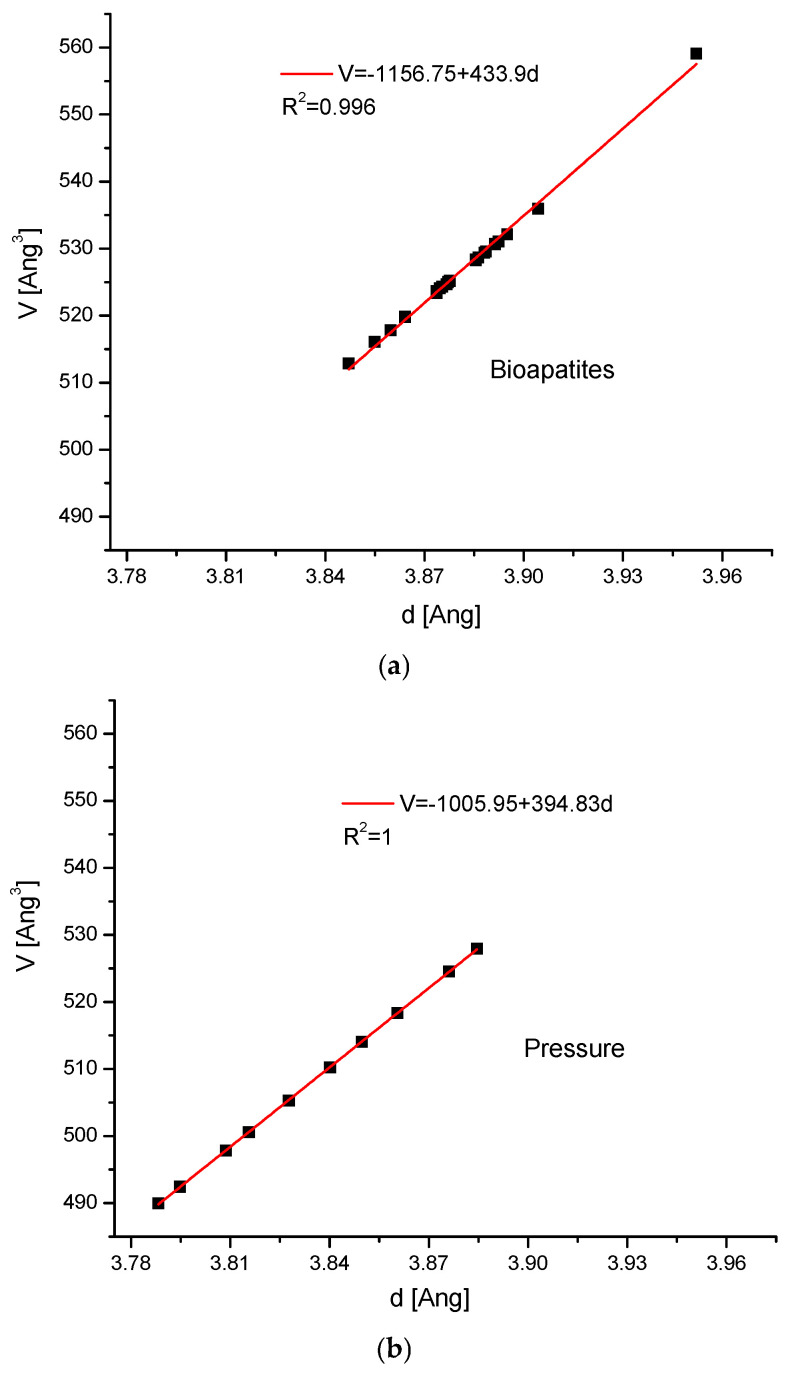 Figure 6
Figure 6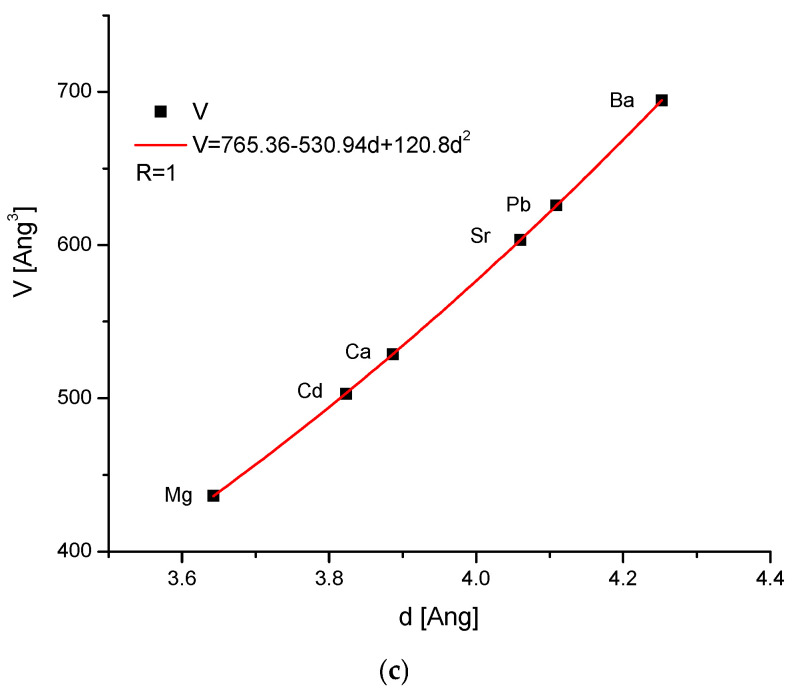 Figure 7
Figure 7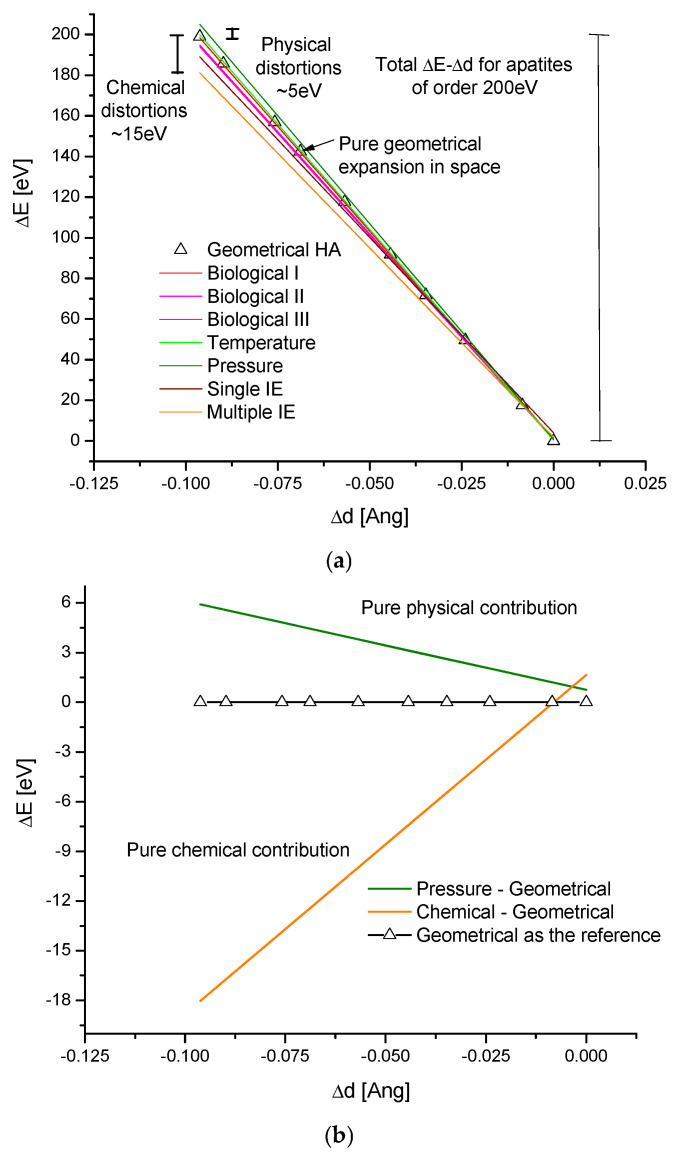 Figure 8
Figure 8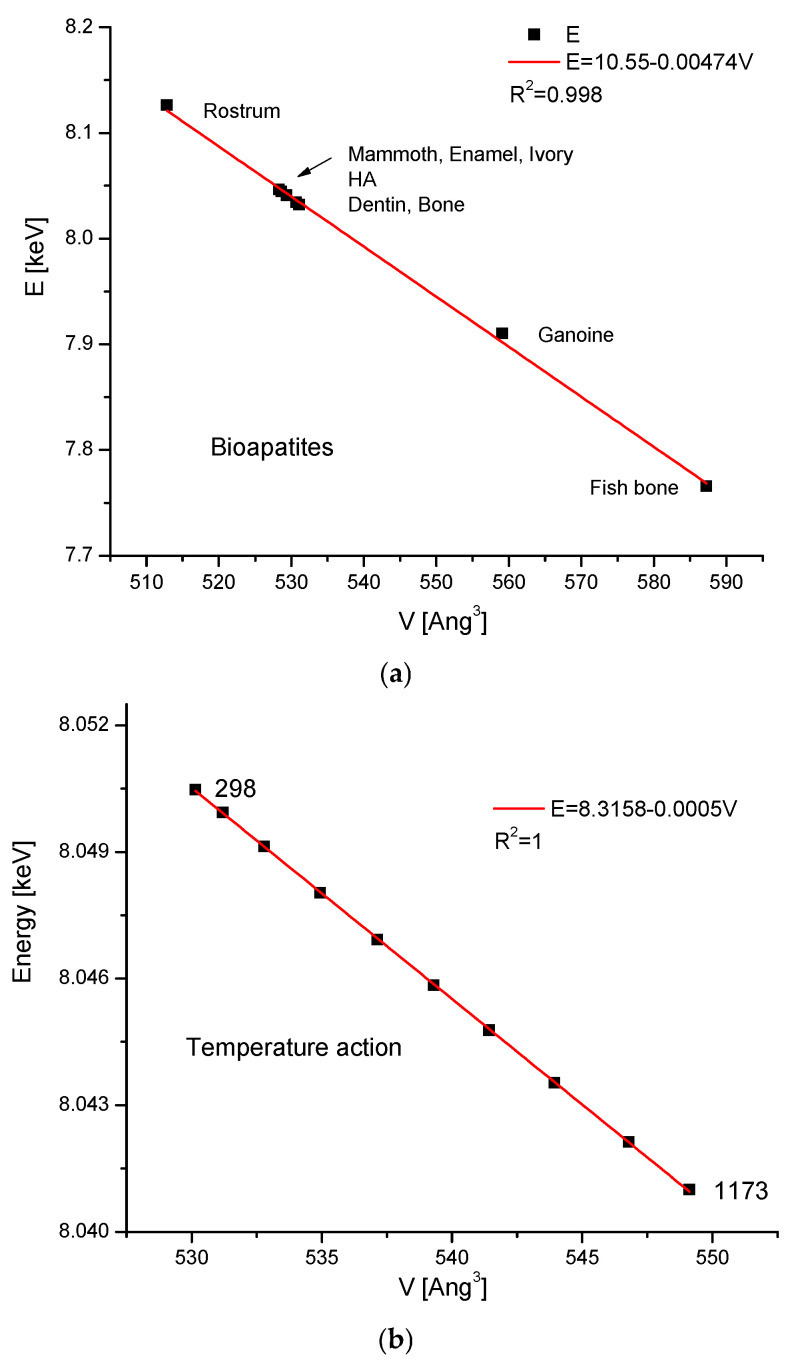 Figure 9
Figure 9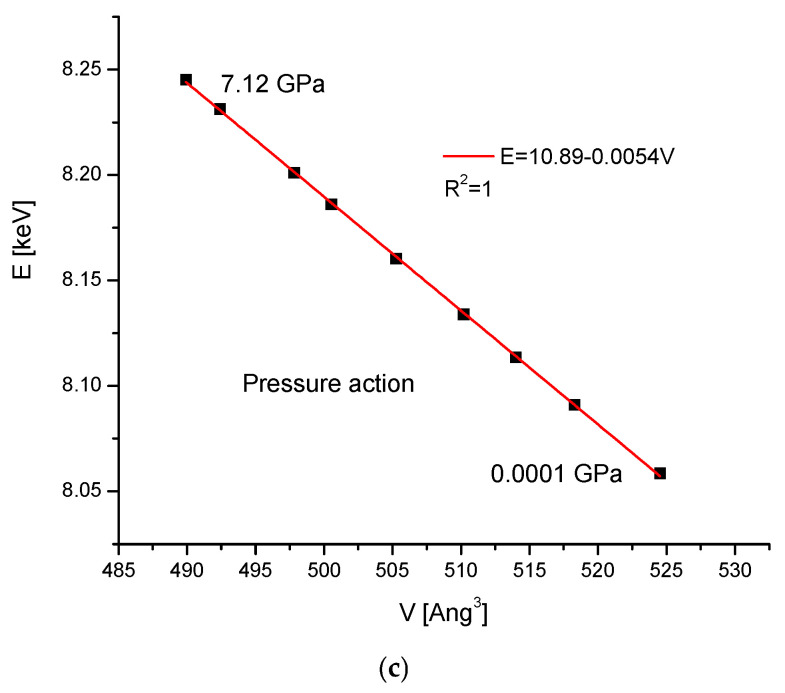 Figure 10
Figure 10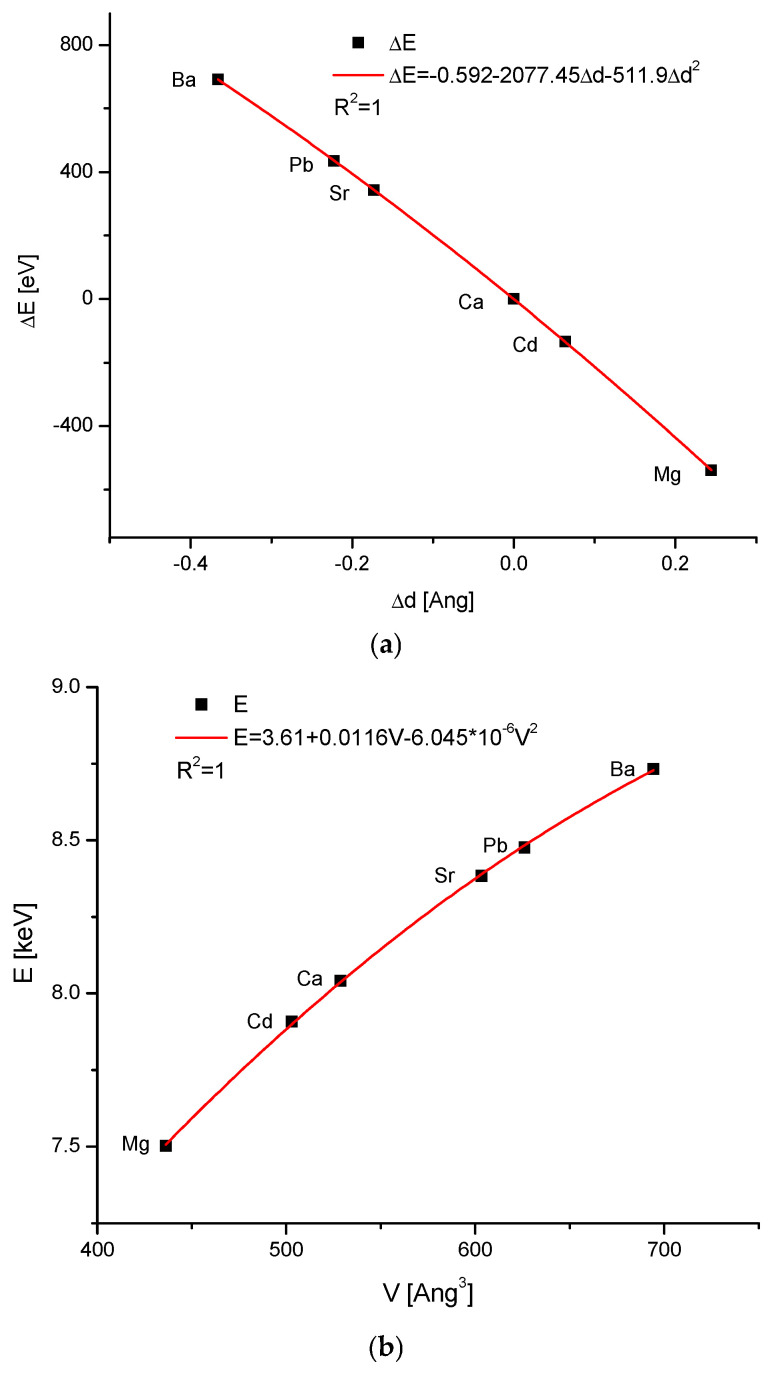 Figure 11
Figure 11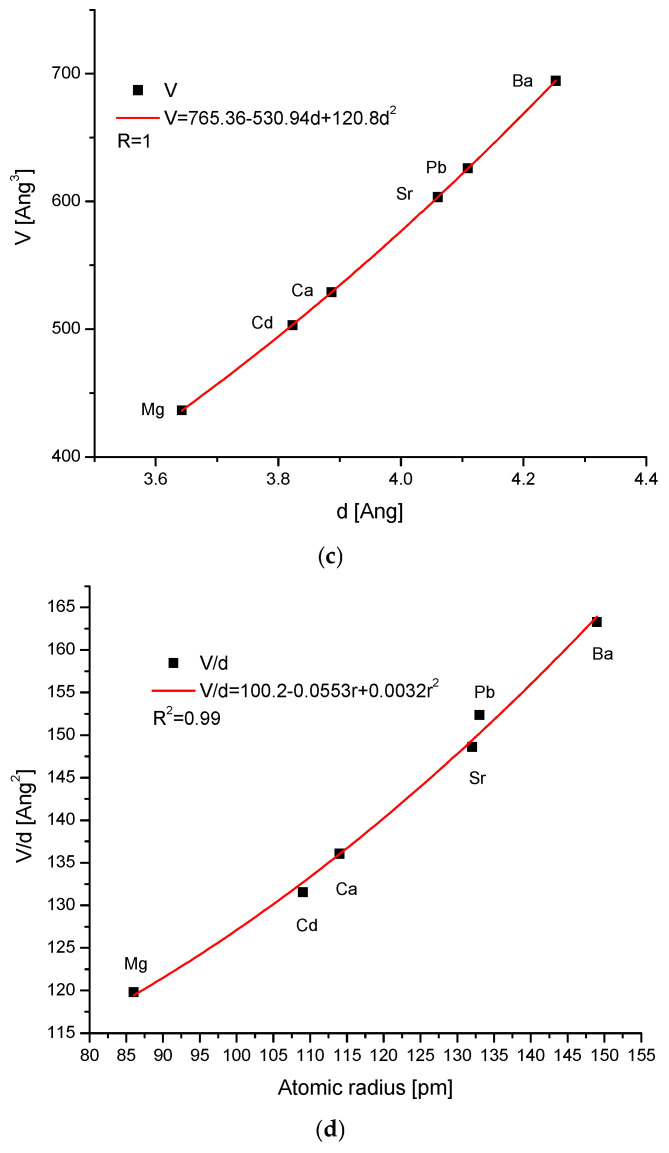 Figure 12
Figure 12Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCalcium Carbonate Crystallization and Inhibition · Bone Tissue Engineering Materials · Mineralogy and Gemology Studies
1. Introduction
There are many different hard tissues supported by apatites and functioning in nature [1]. They belong to the category of composite compounds, since they always compose smaller (enamel, rostra [2,3]) or greater (dentin, bone [4]) parts of different organic materials [5]. Due to their significance in the normal existence and functioning of organisms, they were extensively studied to explain their features [6]. One must realize that organs containing bioapatite have distinct anatomical meanings and functions (teeth, bones, tusks, femurs, antlers, rostra, bulla), and sometimes it is difficult to find a common benchmark when considering them, perhaps aside from the apatite presence. Some reviews of more exotic members of the family of hard tissues were written by Schultze [7] and Kawasaki et al. [8]. A useful review of extra-skeletal bones in mammals was provided by Nasoori [1]. A not-unimportant reason for our contribution is that the obtained knowledge could be applied in potential syntheses of new biomaterials, including ones with biomimetic reactions. Another reason for our interest is the fact that some animal wastes can be used to produce nanosized bioapatite particles on a laboratory or industrial scale [9]. There are some very interesting reviews of various biomaterials, which help provide a better understanding of them. Unfortunately, the studies mostly focus on either the morphological characteristics of tissues or crystallites [10] or on comparisons of the mechanical features of the considered substances [11,12], rather than on chemical and crystallographic parameters. The latter features seem to determine the whole rest of the behaviour of a given organ, which is not a common opinion. The apatite behaviour at the crystallographic-cell level is a basis of all the further characteristics of a given biomaterial in this text.
Since it concerns the subject of our contribution, it is important to mention the tissues under scrutiny: bone, enamel, and dentin of mammals; deer antlers; ivory of elephants and mammoths; similar tusks of walruses; rostra and bullae of, e.g., whales; enameloids of fish; carapaces of tortoises; and other similar tissues. These are characterized by the presence of apatite crystallites as the main hard components, admixed with organic matter, often but not exclusively collagenous. Given the contents, the organic part can be treated as a matrix (dentin, bone) or as a minor but still important internal component (enamel, rostra). In fact, none of the above materials are pure mineral hydroxy-, chlor- or fluorapatite, but they are all composite materials. Even if we limit consideration to the mineral parts of the tissues, we must be aware that the said apatites are enriched in sometimes considerable amounts of additional ions, such as magnesium, sodium, carbonate, and fluorine [13], and depleted of hydroxyl entities [14]. This indicates that, each time, the apatite part is different both from the original hydroxyapatite and from the apatites in the other organs of one organism (e.g., bone apatite differs from enamel and dentin apatites) and is different temporarily—ontogenetically (e.g., young and mature bones differ from each other)—and is different among species—phylogenetically (e.g., dentin apatite of mammals is still a different material than ivory apatite from elephants or mammoths). Nevertheless, similarities were also observed—Li et al. [15] suggested that whale rostrum is, on the one hand, similar, due to its great density, to enamel, while on the other hand, due to its morphological features, it is similar to very dense bone, much depleted of collagen. It is astonishing that, although scientists have studied the shapes and dimensions of apatite crystallites [4], the kinds of organic matrices, the morphology of materials, and the mechanical features of them [16], they somewhat neglected comparative studies of the single-crystallographic cells of particular apatites contributing to different organisms. Perhaps the studies on human bones and teeth are here the exceptions. Such single-crystallographic cells are, in fact, very important. We have proven in our previous contributions that only the apatite and bioapatite cell parameters determine the location of apatite within the isomorphic series of compounds [17]. This seems to involve the given bioapatite inside the whole series of bioapatites and apatites. It allows one to see clearly the ordering of all the apatites, without prejudging whether the mutual transformation of one shape into another is at all possible. One can also set the standard point, from which all the series will be considered, and the hydroxyapatite plays this role in our text. The comfortable comparative knowledge can be obtained using the special set of equations describing the apparent energetic changes in apatite cells, resulting from the changes in Braggs’ dimension d:
and, optionally,
or an equivalent one:
If we compare Equations (1a) and (1b) with (1c), respectively, the relevant values for coefficient k are as follows:
and
where ∆E expressed in keV is the energy difference resulting from the change in Bragg’s crystallographic dimension ∆d in angstroms Å, occurring under the influence of external radiation, leading to the transformation (sometimes very small) of one apatite material into another one. The K coefficient determines the compound’s membership in the given isomorphic series of chemicals. Those K coefficients have a great significance, since they determine the location of the points within the hexagonal structure, as reflected in the expansion expressed by changes in sin θ, i.e., they reflect purely geometrical swelling or squeezing. Although the energy of the X-ray tube is given in keV, we often recalculate our energy results in eV through multiplication to obtain more convenient units.
One can add several equivalent equations to the above ones:
All of them were originally derived in our previous papers.
We had the following aims to achieve in our present contribution: (i) To what level is the Bragg’s dimension of crystals characteristic of the given biomaterial of some organisms? This question can be inverted—Do the organisms choose the crystals of the selected Bragg’s dimension for their hard tissues in a way specific to given species? (ii) What are the possibilities of nature changing the Bragg’s dimensions and probing energies of excitation for bioapatites and apatites in general? (iii) Is the dimension d joined quantitatively with the volume of the crystallographic cell? (iv) Can we reformulate Bragg’s law in terms of other variables?
2. Results
2.1. Range of ΔE–Δd Diagrams for Apatites and Bioapatites
We selected a large amount of available information on the bioapatites present in the different organs of many animals. It is presented in Figure 1a, where we show the points on the diagram ΔE–Δd. Due to the large number of the points, we designed them with the numbers explained in the caption. Some geological and synthetic apatites, fossils, and calculi [18] were introduced for comparison.
This is not the only diagram concerning apatite biomaterials. Some remaining results are shown in Figure 1b. We know many other measurement results for other bioapatite samples, but they lie along the same lines. The range of specific points covers the energy values +100/−300 eV, with Δd equivalents −0.014/+0.14Å. Nevertheless, the clear densification of points lies in the region −50/+60 eV, indicating that the majority of organisms use only particles within a relatively narrow range of the parameters. The ranges lie on both sides of the hydroxyapatite position. Perhaps the main conclusion derived from the above diagrams is that animals use apatite cells with quite strictly defined parameters for themselves, and selection occurs at the level of single apatite cells, not the crystallites, as previously suggested [39,40]. Thus, the parameters of the single-crystal cell of bioapatite can uniquely determine the organ of a given species. We suppose that the assignment of the special type of apatite to a given species occurred during long-lasting adaptation in the processes of evolution, as was suggested by Weiner and Wagner, although they considered this question from quite a different point of view [41]. Please note that the densification of apatites is accompanied by alkalization during synthesis, while erosion and decay occur with acidification in further life [42]. Such environmental conditions can influence the crystal cell parameters. However, we must remember the measurement errors and the possible contamination of the samples.
The next problem to be solved is whether the range of apatite changes, as described in the framework of our formalism, ΔE–Δd diagrams, is unique only when it concerns biological actions. The range of biological variability was extracted from our data (Figure 1) and presented concisely in Figure 2a. It is somewhat surprising that the curve is asymmetric in relation to the hydroxyapatite standard. The organisms use rather less condensed forms of apatites. Figure 2b presents the data for the dinosaur fossil apatites, which are originally biological but are influenced by geological factors, including high pressure and transformation into francolite, a specific mineral containing both fluorine and carbonates, the monoclinic cousin of apatite [43]. The monoclinic form of francolite does not exclude it from the isomorphic series of apatites (observe the slopes of the curves). Nevertheless, the dinosaur bones exhibit a denser structure compared to standard hydroxyapatite.
For the comparison, the variability resulting from the heating is shown in Figure 2c. The influence of temperature is clear, but much narrower than that of biological actions. Here, the variability is one-sided: starting from the temperature 0 K, it always leads to swelling with increasing temperature. Obviously, the comeback is possible with the temperature drop, but only if the compound has not changed phase during the previous warming. Another case, evident in studies on the energy and dimensional changes in apatite crystals, relies on the pressure effect. Here (Figure 2d), the increase in pressure exerts a clear squeezing effect on the crystals.
To fully consider the allowed changes in the ΔE–Δd diagrams, we added new figures for the chemical changes, i.e., ion exchanges. The first one illustrates the case when only one type of cation is exchanged, leading to the synthesis of substituted compounds in the same crystallographic system (Figure 2e). We know from our earlier contribution that, among the three allowed kinds of ion exchange—of cations, anions, and simple chemical moieties—at the channel location, the first exerts the greatest effect. Indeed, the ranges of introduced changes in ΔE and Δd values are larger than in the previously shown cases. The next case confirms our suppositions about the prevalence of cationic exchanges—in Figure 2f, the complete exchanges involving the three allowed chemical entities are shown. The total change, although very significant, is not markedly different from that in a single cationic exchange. Undoubtedly, the ion exchange of all ions leads towards the synthesis of a totally new compound, but we—from our assumption—selected the substances organized in the same crystallographic system. Since measurements in different laboratories can be performed relative to different, locally determined hydroxyapatite standards, hydroxyapatite scattering is shown in Figure 2g. The data were taken from the classic papers mentioned in the caption. One can see that the deviations from Hughes’s results are small. This can be accounted for by a small shift along the ΔE–Δd curve. In addition, this correction is really important only for popular bioapatites. It is worth noting that the coefficient in the equation located in the figure is very close to that estimated in Equation (2), characteristic of the spontaneous expansion of the crystal cell.
Another statistical estimate of the robustness of the results relies on comparing the residuals obtained during the creation of the ΔE–Δd function, e.g., for the case when hydroxyapatite is pressurized (Figure 2h). It is an astonishing fact that the residuals, as such, also form an extremely regular structure. This seems to confirm our conviction that apatite variability is strictly quantitative.
In summary, we can state that there are the following ways to change crystal size under the influence of the factors presented in Table 1.
2.2. Passing to the Absolute Values
Now, we can pass to the absolute values of d, departing from the increments. One can ask, what happens to the volumes of the crystal cells of compounds if we pass from dimensions d(111) to the volumes? To answer this question, we will cite only three cases. The first one shows the relationships between V-d for the biominerals in Figure 4 (the reference to Figure 1a); the second one shows the relationship for hydroxyapatite under pressure (refer to Figure 2c). The third one presents the relationships connected with the ion exchange in cationic positions. The similar relationships concern all the previously considered cases.
The relationship is surprising and very simple and regular, which is not obvious if we consider the variability of parameters a and c for apatites, but more obvious if one takes into account that Bragg’s law concerns parameter d and not the ones previously mentioned:
In fact, the correction of the second order is significant only in some extreme cases, e.g., in the case of very radical full ion exchanges. In most applications, the relationship (5a) will be limited to the first two terms of the equation.
Since relationship (5a) acts also on the opposite side, we can transform Bragg’s law in a more illustrative way:
The theoretical value of V/d can be derived from the geometrical assumptions for the hexagonal system, where the volume of the cell is
and, from Equation (5),
For the (111) orientation,
Thus,
After the introduction of the above-mentioned values of a and c for our standard hydroxyapatite, the result is 135.9, which is in very good accordance with the data from Figure 4: the values are 135.97 from Figure 3a, 135.89 from Figure 3b, and 135.71 from Figure 6c, respectively. This simplifies the formulation of Bragg’s law for pure hydroxyapatite:
and, in fact,
where E is in keV, while V is in Å^3^.
Thus, Bragg’s law in such an approach as Equation (3b) (and in the simplified form (3c)) is an interplay between the apparent probing energy E and the available internal crystallographic cell volume V, controlled by the value of the sin of the angle of positive diffraction. This is a peculiar kind of geometry inside the crystal cell, where the volume is proportional to the Bragg’s dimension (here (111)). The energy is, from the assumption, the apparent probing energy, since its action is coupled to the increase/decrease in the crystal cell volume. Here, it is centred around the exciting energy of 8.041 keV for the Cu tube, but excitation using monochromatic lines from other types of X-ray tubes or synchrotron lines would form local environments around other lines. Thus, we can probe the crystals in many ways. This concerns some peculiar small spaces covering some hundreds of Å^3^ under pressures expressed in gigapascals (Figure 2d) and in the temperature range 0–1100 K (Figure 2c).
When we have the possibility of immediately determining the volume from X-ray diffraction measurements, we can use the instrument for studies in the field of volume-based thermodynamics.
3. Discussion
We managed to collect a large amount of X-ray diffraction data on the apatite molecules that constitute different organs of many animals and humans, as well as some defective excretions. The system was greatly expanded by the addition of data on various mineral apatites. We applied our earlier method of calculation for the apparent change in exciting energy in relation to the change in Bragg’s dimension d (mainly d(111)). Using the very fine linear results from Equation (1c), we proved that this equation can be supplemented by the equally simple Equation (4a), which gives the volume of the crystal cell inside. Then, Bragg’s equation can be transformed from a classical wave form into the E vs. V form in Equation (3b). In such a form, it does not immediately express the constructive diffraction phenomena, but the simple fact that each de/compression resulting in volume changes shows the apparent effect on the probing energy.
One can distinguish the geometrical, thermal, pressurized, biological, and ion-exchange-caused (chemical) volume expansions of isomorphic crystals. If we compare the coefficients of the slopes in the equations in Figure 2, some conclusions can be easily drawn.
According to Equation (2), one can describe the geometrical expansion of the hexagonal system using the coefficients from the above equations, as if only the space expansion occurred.The thermal expansion (Figure 2c) is most similar to the geometrical expansion, and we can imagine that it results from the nonuniform distribution of the thermal effects within the crystal cell, which has a different symmetry from the spherical one, and the thermal heterogeneity is most tailored to this situation.The situation during the pressure action (Figure 2d) is more distant than in the geometrical case. This results from the fact that the uniform pressure exerts a nonuniform influence on ions located at different points in the hexagonal space, thus having a variable primary energetic status. At the margin, according to our intuition, the effects of temperature and pressure would be closer to each other if we worked with other crystals in the cubic crystallographic system. Both temperature and pressure actions are of physical character, and their additional contribution is approximately ~5 eV. They are reversible unless the conditions for the phase change are met.Much more pronounced are the chemical effects. We generally consider them as ion exchanges, with the single ones leading to the substituted compounds and with the multiple ones forming quite new compounds. We must only remember that we solely consider the substances belonging to one selected crystallographic class. Here, the numerous small distortions (asymmetries) arrive. The range depends on the sizes of the introduced ions relative to those of the original ions. In general, the distortions and overcoming the internal strains decrease the energies of the order to −15 eV. The chemical influence on crystal expansion is the greatest among all the considered factors. The chemical effects are sensitive to phase changes, as seen in the transition from hexagonal to monoclinic structure in chlorapatite.The biological apatites, whose variability results from limited ion exchanges and vacancies in response to specific ion supplies, lie relatively close to the geometrical trajectory of volume expansion; what is interesting is that it is possible to observe an approximate inversion of the synthesis processes in erosion/decay-type reactions [41].
All that was said above is summarized in Figure 4. The contributions were grouped by colour. From the sizes of physical (here pressure-invoked) and chemical (here multiple-ion exchanges) changes, the geometrical contributions (the reference values determined using Equation (2)) were subtracted. The physical and chemical influences were presented in a “naked” shape (Figure 4b). This is shown in the diagrams of the ΔE–Δd type.
3.1. Intracrystalline Approach
Up to now, we have mainly concentrated on the questions of the growth increments of the essential quantities: E—the apparent probing energy, d—Bragg’s distance, and V—the volume of the crystal cell. Now, similarly to Figure 4 and Equations (3) and (4), some attention must be paid to the absolute values. The probing energy value can be easily established, since it is simply 8.041 + ΔE [keV] from the assumption. The value of 8.041 keV corresponds to the photon energy of the essential K lines in the spectrum of the Cu tube. It establishes the first level, from which we calculate the next levels by adding the corrections ΔE. One must remember that the E value is the apparent probing energy from the selected excitation source. Perhaps the values of ΔE have greater practical meaning, since they can probably reflect the real difference in energy levels between different compounds or between different states of the selected compound, and are relatively easy to experimentally verify, e.g., using calorimetric methods. However, one cannot exclude that their meaning is only theoretical and ordering in relation to isomorphic compounds.
The solution is quite obvious if one considers Equation (1c) as the differential version of another more general equation expressed in absolute values for hydroxyapatite and takes into account the relationship in Equation (4d), which leads immediately to the formulation
and gives a particularly simple relationship between the absolute values of E and V, not just between the increments. This is clearly shown in Figure 5. Equation (6) is not the basic equation. There is Equation (1), which is rigorously derived, while Equation (6) is the empirically derived integral equivalent of Equation (1). Equation (6) is indirectly drawn from Equation (1c), and its accuracy depends on how real dimensions a and c interplay, giving the value d(111).
However, joining Equations (3c’) and (6), we can obtain the especially simple version of Braggs’ law:
where one of the variables, the apparent probing energy E, is eliminated. It is the second-order equation of sin θ against V. The measurement of the diffraction angle yields the crystal cell volume as one of the two roots of this equation.
The unique method of calculating crystal cell volumes directly from X-ray diffraction measurements promises to make it easier to obtain the values necessary for thermodynamic measurements [53].
3.2. Interpretation
The irradiation of the sample allows the probe to measure the new equilibrium in the crystal cell. The initial conditions are as follows: the exciting energy is equal to 8.041, and the volume of the standard hydroxyapatite cell is 527.9Å^3^. Each deviation from the assumed volume results in a change in the apparent probing energy, since this energy is tailored to the real volume of the crystal cell. The probing energy and the cell volume are coupled by Equations (3b,c) (transformed Bragg’s equation) and (6). For more explanations—see Appendix A.
3.3. Deviations
As shown in Figure 5a, there is clear scatter in the curves which formally present the different isomorphic series, with hydroxyapatite as the member. In cases where the parameter k in Equation (1c) is close to −2068.9, we have the linear versions of Equations (1c) and (6). However, the extensive ion exchanges lead to greater deviations in the value of k. We can consider the case in which Ca in hydroxyapatite is fully substituted by divalent cations, ranging from Mg [54] to Ba.
We know that other researchers did not confirm Patel’s claims regarding the possibility of the full synthesis of Mg-apatite. Nevertheless, when we include Patel’s data in our calculations, we obtain perfect results, as if such a compound indeed exists. We decided to include the relevant data in our diagrams, with the full awareness that it may be only a result of a mathematical trick. Similarly, Drouet [55] used data on the magnesium compound to obtain quite credible data on the thermodynamic functions for MgHA.
The cases with a partial substitution (e.g., Fe, Co, Mn, and others) are not taken into account here. One can observe that even the basic curve of ΔE–Δd is slightly deviated from linearity (Figure 6a). Nevertheless, the curve is perfectly approximated with a polynomial of the second order. Moreover, the corrections in terms of first- and second-order are small, and one can observe that the essential coefficient −2077.45 is not far from the theoretical value of −2068.9. It does not prevent participating in the mentioned elements in one isomorphic series. The nonlinearity is reflected in the version expressed in the absolute values (Figure 6b), i.e., in the function E–V. The relationship expressing the V–d curve is also a function of the second order (Figure 6c). Now, we can pass to another relationship. If we consider the ionic radii [56] of the ion substituting Ca in the structure of the hydroxyapatite, then it is clear that the cations influence the volume of the crystal cell, to which they enter. In such a situation, we cannot treat the apatite cells as the invariable units, since they simply adjust their dimensions to the new elements (Figure 6c,d). This was anticipated theoretically by Wu et al. [57].
4. Materials and Methods
4.1. Materials
For the data search, we used platforms and databases such as Web of Science, Scopus, Scilit, and Google Scholar. We used available data from the world literature on widely understood apatite-type compounds with hexagonal structure, and X-ray, electron, or neutron diffraction [58] were the only instrumental methods considered for their description. The fine presentation of what can be determined using X-ray diffraction was given by Moulick et al. [59]. The hexagonal structure is considered only for the uniformity of the results and is not unique. Where it is necessary, we consider a monoclinic structure as well, after a small recalculation. Identical reasoning can be applied to other, but exclusively uniform, crystallographic classes of substances. No experiment considered in this paper was performed in the laboratories of the authors, which does not mean that they do not conduct experiments; they also conduct experiments in the field of X-ray diffraction [60,61], sometimes using very advanced X-ray 2D imaging techniques [62]. Simply, nobody in the world had the potential to make all the demanded measurements. The data are supported by the sources, which are sometimes well documented when they concern human enamel, dentin and bone, while they are much scarcer when they concern ivories, antlers, or rostra. It was a question of proper selection among the numerous data in the former case and of utilizing the available data in the latter case. Pure inorganic apatites were considered in even more detail. The special inspiration could be found in the papers [55,57,63,64,65,66,67,68]. The results were presented in the form of ΔE–Δd diagrams described by Equation (1) or other similar ones where it was relevant. The values of the ΔE and Δd parameters were always determined relative to hydroxyapatite values; thus, most results were relative. As a rule, the values of Hughes et al. [48] were applied in our contribution for this compound, and are presented in Table 2.
The dimensions d are given in angstroms Å, temperatures T as the absolute Kelvin K, and pressures P in gigapascals GPa. The results of ΔE calculations are then reported in kiloelectronvolts, but they are often recalculated to electron volts eV in our contribution.
The authors of the databases are appropriately cited in the text, and we are deeply indebted to them. Due to the huge amount of data, we could not clearly describe all the bioapatites in Figure 1a, and we decipher the particular numbers in the caption.
4.2. Methods
Supplementing previously cited equations, we should write Bragg’s equation in the energetic representation [69]:
and add the interrelations between Miller indices in the hexagonal system:
After introducing the relevant data for our hydroxyapatite standard [13], we found the theoretical values of K_1_ −2068.6eV/Å from Equation (2a) and K_2_ −2069.2eV/Å from Equation (2b). The good approximation of the constant k in Equation (1c) is the value −2068.9, and it expresses the ideal model of an isomorphic series if its members expand only due to geometrical reasons.
Since we have many coefficients in our equations, we denoted them all with K letters, each with a corresponding consecutive subscript. All the equations in our contribution, except Equations (3a) and (5), are derived by the authors. The equations of special importance are presented in bold.
Please note that the coefficients of determination R^2^ in our figures, in the majority of cases, have values of 1 or very close to 1, indicating that our equations are of functional character and not merely approximations of the experimental results. Otherwise, in many cases, the equations are rigorously derived from earlier assumptions.
The authors paid special attention to robustness and the self-tailoring of the results. At first, selecting the available results was necessary. Sometimes it was simple due to the scarcity of the materials (as for the whale rostrum); in other cases (e.g., human bones), we had to make the selection, and then the laboratory basis, the description of the experiment (Knyazev et al. [44], Matsukage et al. [45]), or the multiple uses in the scientific literature (Hughes et al. [48] for the standards) were the decisive reasons. The authors who reported a whole series of coincident results were preferred (Piga et al. [37,38]). All the deviations in the Δd value were repeated for the ΔE values due to the construction of Equation (1a–c). So, for the uniform sets of results, the local deviations were meaningless. Values very close to 1 for the coefficient of determination R^2^ result from the rigorousness of the derivation of Equation (1), and they are confirmed by the astonishing results shown in Figure 2h.
The procedure of the calculations for the construction of ΔE–Δd diagrams is very simple:
- Using the values of parameters a and c, calculate d according to Equation (4);
- Subtract the values d from d_0_ (standard), getting Δd;
- Using the latter value, calculate ΔE according to Equation (1a) or (1b);
- Furthermore, it is a trivial arrangement of the variables in the diagrams.
The paper, although based exclusively on the studies of other researchers, is absolutely original in the concepts of the authors and, to our knowledge, has no analogue in the world’s literature.
4.3. Software
Origin 9.1 was used for result evaluations and graph preparation.
5. Conclusions
We collected available data on apatite crystals in very different biostructures, including the extreme cases—whale rostrum and ganoine. The expansion of the crystallographic cells in bioapatites was compared with processes of expansion induced by thermal and pressure actions and by ion exchange, whether of single ions (substitutions) or of all ions (synthesis of a new compound). The condition was that all ΔE–Δd diagrams should concern the compounds existing in the same crystallographic system. Essentially, all the studied diagrams were very similar with respect to the parameters describing the relevant straight lines. The wide possibility of steering with the crystal cell volume was proven. We extracted subtle differences in the slopes of the lines and attributed them to differences in the physical or chemical factors reflected in the ΔE–Δd diagrams. The discoveries could be transmitted into absolute values, with the absolute probing energy E = 8.041 + ΔE if the excitation was with the Cu X-ray tube and V = K_3_ + K_4_d, or alternatively V = √3 * a^2^c/2 for the hexagonal system in configuration (111). Once the relationship between the Braggs’ dimension d and volume V was established, we proposed a new transformed version of the generalized Bragg’s law, 6.2 * K_4_ /E = (V − K_3_) * sin θ, and even a simplified version, k_7_/E = V * sin θ, expressed only in mechanical variables.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Nasoori A. Formation, Structure, and Function of Extra-skeletal Bones in Mammals Biol. Rev.202095986101910.1111/brv.1259732338826 · doi ↗ · pubmed ↗
- 2Rogers K.D. Zioupos P. The Bone Tissue of the Rostrum of a Mesoplodon Densirostris Whale: A Mammalian Biomineral Demonstrating Extreme Texture J. Mater. Sci. Lett.19991865165410.1023/A:1006615422214 · doi ↗
- 3Tang L. Zhang L. Yue M. Tian D. Su M. Li Z. New Insights into the Ultrastructure of Bioapatite After Partial Dissolution: Based on Whale Rostrum, the Densest Bone Microsc. Microanal.2019251323133010.1017/S 143192761901502231599216 · doi ↗ · pubmed ↗
- 4Elliott J.C. Calcium Phosphate Biominerals Rev. Mineral. Geochem.20024842745310.2138/rmg.2002.48.11 · doi ↗
- 5Forien J.-B. Zizak I. Fleck C. Petersen A. Fratzl P. Zolotoyabko E. Zaslansky P. Water-Mediated Collagen and Mineral Nanoparticle Interactions Guide Functional Deformation of Human Tooth Dentin Chem. Mater.2016283416342710.1021/acs.chemmater.6b 00811 · doi ↗
- 6Habraken W. Habibovic P. Epple M. Bohner M. Calcium Phosphates in Biomedical Applications: Materials for the Future?Mater. Today 201619698710.1016/j.mattod.2015.10.008 · doi ↗
- 7Schultze H.P. Scales, Enamel, Cosmine, Ganoine, and Early Osteichthyans Comptes Rendus Palevol 2016158310210.1016/j.crpv.2015.04.001 · doi ↗
- 8Kawasaki K. Keating J.N. Nakatomi M. Welten M. Mikami M. Sasagawa I. Puttick M.N. Donoghue P.C.J. Ishiyama M. Coevolution of Enamel, Ganoin, Enameloid, and Their Matrix SCPP Genes in Osteichthyansi Science 20212410202310.1016/j.isci.2020.10202333506188 PMC 7814152 · doi ↗ · pubmed ↗