Regulatory Potential of piRNAs Targeting Klotho and Other Genes
Anna Pyrkova, Kyrmyzy Akhmetova, Murat Zhanuzakov, Makpal Tauassarova, Aizhan Rakhmetulina, Raigul Niyazova, Saltanat Orazova, Piotr Zielenkiewicz, Anatoliy Ivashchenko

TL;DR
This study identifies piRNAs that may regulate the Klotho gene and other genes involved in metabolism and aging.
Contribution
The paper computationally predicts novel piRNA–mRNA interactions involving the Klotho gene and other metabolic regulators.
Findings
Several piRNAs were predicted to bind the Klotho (KL) mRNA in a single cluster of sites.
piR-3215034 and piR-6885965 interact with mRNAs of multiple genes in different regions.
Some piRNAs bind exclusively to the 3′UTR of FGF23 mRNA.
Abstract
Background/Objectives: piRNAs (PIWI-interacting RNAs) can significantly modify the expression of protein-coding genes by suppressing the translation process. The aim of this work was to computationally evaluate the potential interactions between piRNAs and the mRNA of the Klotho gene, as well as other genes involved in key metabolic pathways related to health and lifespan regulation. Methods: Bioinformatic analysis was conducted using the MirTarget program, which determines the quantitative characteristics of predicted nucleotide interactions between piRNAs and mRNA targets. Results: Several piRNAs (piR-44682, piR-1940042, piR-3008660, piR-3215034, piR-6885965, and piR-7980636) were predicted to bind within a single cluster of binding sites on the KL mRNA. In addition, piR-6890096 was predicted to interact with the KL mRNA through full complementarity. The mRNAs of AFF2, BCL2L11, CPT1A,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6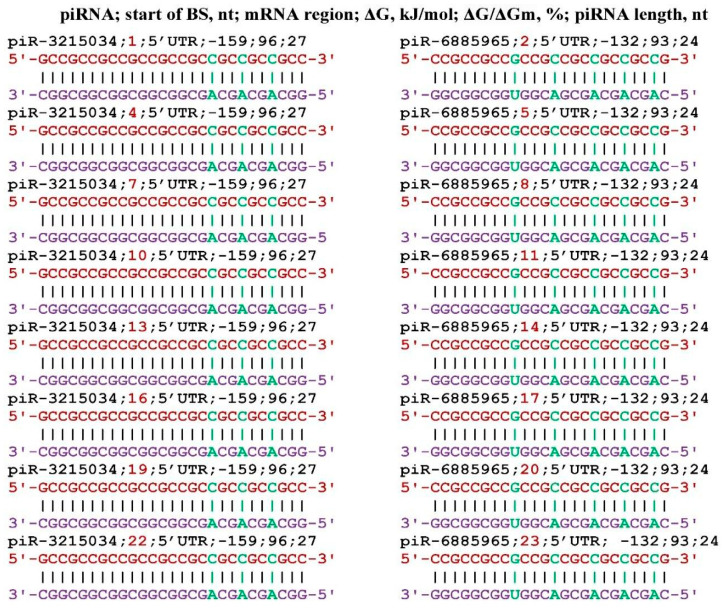 Figure 7
Figure 7 Figure 8
Figure 8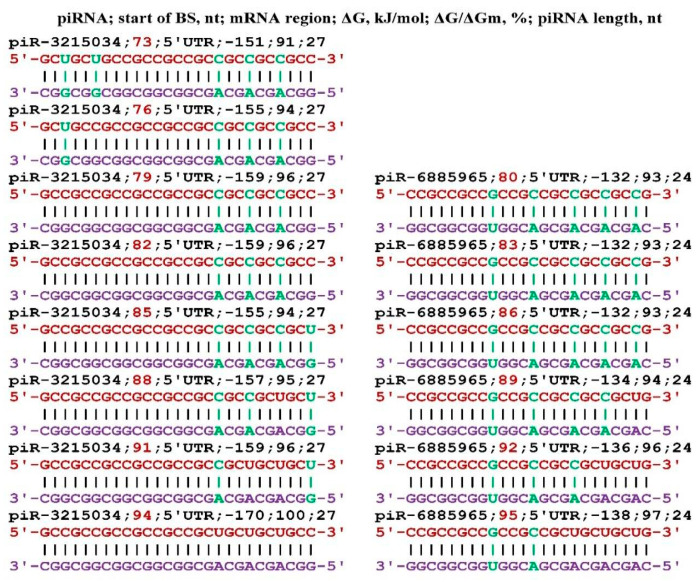 Figure 9
Figure 9 Figure 10
Figure 10 Figure 11
Figure 11 Figure 12
Figure 12- —Ministry of Science and Higher Education of the Republic of Kazakhstan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParathyroid Disorders and Treatments · Mechanisms of cancer metastasis · Fibroblast Growth Factor Research
1. Introduction
The Klotho (KL) gene belongs to a family consisting of α-klotho, β-klotho, and γ-klotho. In the present study, we focus on the α-klotho subfamily, hereafter referred to as Klotho. The KL gene is highly expressed in the brain, liver, and kidney, and has attracted substantial interest among biologists and physicians due to its involvement in diverse metabolic pathways and disease processes. Notably, reduced Klotho protein levels are associated with increased mortality in various diseases. Its molecular, physiological, and therapeutic characteristics have been extensively reviewed [1]. Growing evidence highlights the diagnostic and therapeutic potential of KL [2]. In particular, low serum Klotho levels serve as a reliable biomarker for both the severity of cardiovascular-renal-metabolic disorders and the corresponding risk of mortality [3]. Consequently, exogenous Klotho supplementation has emerged as a promising therapeutic approach. Future investigations may clarify its utility in diabetic nephropathy and other diseases [4]. Recent research has also explored epigenetic regulation of KL expression in various diseases [5,6], providing deeper insight into the mechanisms controlling its activity. The role of KL in diabetes has been increasingly recognized [7,8,9,10,11], and numerous studies have examined its function in kidney diseases [12,13,14]. Additionally, accumulating evidence supports the involvement of Klotho in cancer biology [15,16,17,18,19]. A substantial body of work has also addressed the relationship between KL and Fibroblast Growth Factor 23 (FGF23) [20,21,22,23]. Beyond disease-specific roles, the KL gene has gained particular attention as a key regulator of aging [24,25,26]. Collectively, these findings underscore the broad biological significance of KL and its potential as a therapeutic target.
miRNAs (mRNA-inhibiting RNA), small non-coding RNAs that inhibit mRNA translation, play a critical role in the post-transcriptional regulation of gene expression. Consequently, identifying miRNAs that influence KL gene expression is essential for understanding the molecular mechanisms underlying its dysregulation in disease. Several studies have reported that specific miRNAs modulate Klotho protein levels. For example, miR-34a suppresses KL expression by directly binding to the 3′-untranslated region (3′-UTR) of its mRNA [27]. miR-339 has also been implicated in the regulation of KL expression, potentially contributing to altered gene expression patterns observed in patients with schizophrenia.
Recent studies have therefore focused on elucidating the role of miR-339-5p in controlling KL gene expression [28,29]. In addition, the Klotho-derived peptide KP1 has been shown to inhibit cellular senescence by restoring endogenous KL expression, and miR-556-3p was identified as an indirect repressor of the KL gene [30]. miR-200c has been demonstrated to modulate KL expression in human kidney cells exposed to oxidative stress [31], whereas miR-379 regulates KL synthesis in the context of apoptosis [32]. Furthermore, KP1 peptide-induced suppression of cellular senescence has been linked to enhanced KL expression via post-transcriptional regulation mediated by miR-223-3p [33]. This list of miRNAs associated with KL regulation is not exhaustive and requires further verification. In the present study, we analyzed the interactions between known Klotho-related miRNAs and KL mRNA to clarify their regulatory roles.
Compared with miRNAs, piRNAs (PIWI-interacting RNAs) have been far less studied in the context of KL mRNA regulation. Like miRNAs, piRNAs are thought to regulate the expression of protein-coding genes, primarily through post-transcriptional mechanisms [34,35]. Therefore, identifying which piRNAs can directly bind to KL mRNA is an important step toward understanding their potential regulatory roles. piRNAs are capable of substantially altering gene expression by suppressing translation, and because they function as endogenous regulators, they hold promise for both diagnostic and therapeutic applications. However, their investigation is complicated by the extremely large number of known piRNAs [36]. Without computational approaches, determining which piRNAs may target KL—or other genes—is highly challenging. Furthermore, it is important to assess whether piRNAs and miRNAs that interact with KL mRNA also target additional genes, thereby potentially causing off-target effects. For these reasons, effective computational tools are essential for studying the interactions of piRNAs and miRNAs with their mRNA targets. In this study, we employed the MirTarget program to analyze the quantitative characteristics of piRNA with the mRNA of target genes. This program has demonstrated reliability and effectiveness in previous studies.
2. Materials and Methods
The nucleotide (nt) sequence of KL gene and other genes were downloaded from National Center for Biotechnology Information database NCBI (https://www.ncbi.nlm.nih.gov, 2022). The nucleotide sequences of 8,480,000 piRNAs were taken from Wang et al. publication [36]. The 3707 miRNAs were taken from Londin et al. article [37], the 2567 mature miRNA sequences were taken from the miRBase database (http://mirbase.org), and the 1036 miRNAs from the article by Backes et al. [38].
The piRNA and miRNA binding sites (BSs) in mRNA were predicted using the MirTarget program [39]. This program predicts the following features of piRNA and miRNA binding to mRNA: (a) the initiation of piRNA and miRNA binding to the mRNA from the first nucleotide of the mRNA; (b) the localization of the piRNA and miRNA BSs in the 5′-untranslated region (5′UTR), coding domain sequence (CDS), and 3′-untranslated region (3′UTR) of the mRNAs; (c) the schemes of nucleotide interactions between piRNAs and miRNA with mRNA; (d) the free energy of the interaction between piRNAs, miRNA and the mRNA (ΔG, kJ/mol); and the ratio ΔG/ΔGm (%) is determined for each site. ΔGm equals the free energy of piRNA and miRNA binding with its fully complementary canonical nucleotide sequence. Only piRNAs and miRNAs whose nucleotides interacted with mRNA using canonical (G-C and A-U) and noncanonical (G-U and A-C) nucleotides with a given ΔG value were selected from the calculated data [40,41]. The MirTarget program finds hydrogen bonds between piRNAs and miRNAs with mRNA according to the physicochemical characteristics of nucleotide interactions.
MirTarget differs from other programs in terms of finding piRNA BSs on mRNA in the following: it takes into account the interaction of piRNA and miRNA with mRNA over the entire piRNA and miRNA nucleotides sequence; it considers noncanonical pairs G–U and A–C; and it calculates the free energy of the interaction of the piRNAs and miRNA with mRNA. Note that the G, A, C, and U nucleotides, which comprise the RNA structure of microorganisms, plants, and animals, interact identically under equal conditions. Therefore, the physicochemical properties of canonical and noncanonical nucleotide pairs given above do not require additional proof of the previously established physicochemical characteristics of their interaction. The reliability of translation suppression by miRNAs that are fully complementary to mRNAs was proven by A. Fire and C.C. Mello, who were awarded the Nobel Prize in 2006 for this research [42].
3. Results
Of the entire piRNA database, only seven piRNAs (piR-44682, piR-1940042, piR-3008660, piR-3215034, piR-5194426, piR-6885965, piR-7980636) bound to the mRNA of KL gene (Figure 1). Seven piRNAs each had one BSs and piR-6885965 had two BSs. All BSs were located with a partial overlap of nucleotides, forming a cluster of BSs from 33 nt to 71 nt, only 39 nt long. Such an arrangement of BSs piRNAs in CDS mRNA leads to competition between them for binding to mRNA. piR-6890096 interacts with 3′UTR in a completely complementary manner (the value of ΔG/ΔGm is 100%), which, at its concentration comparable to that of mRNA, will lead to inhibition of the translation process.
Note that the term miRNA or piRNA binding cluster introduced by us differs from the term cluster of the location of several genes encoding miRNA in a section of the chromosome. The KL gene is many times more strongly expressed in the kidneys than in other organs, and a high concentration of piRNA is required to suppress its synthesis. Of the 7310 miRNAs, only ID00756.3p-miR is bound to the mRNA of the KL gene (Figure 1). ID00756.3p-miR can only interact with mRNA of the KCNN2, NOTCH3, and ZNF592 genes. The ID00756.3p-miR was in the BSs cluster of seven piRNAs located in the CDS mRNA KL gene; therefore, it competed with them.
Out of 8,405,000 piRNAs, only eight piRNAs were identified, of which piRNA-6885965 has two BSs in the mRNA of the KL gene. piRNA-6890096 interacted with the mRNA of the KL gene in a completely complementary manner using only canonical nucleotides. Out of 7310 miRNAs, only ID00756.3p-miR could bind to the mRNA of the KL gene. The piRNA (except piRNA-6890096) and miRNA bound in a 39 nt region (Figure 1). Here and henceforth, piRNA BSs with overlapping nucleotide sequences will be referred to as BSs clusters. As a result of this arrangement, piRNA BSs was compete each other for interaction with the target gene mRNA, and as a result, only one piRNA that binds mRNA more strongly than others or is in a significantly higher concentration will be bound for a longer time than other piRNAs. The formation of clusters of piRNA and miRNA BSs in mRNA is a kind of guarantee of the non-random association of small RNAs and their target genes.
The results obtained give reason for the possibility of specific regulation of KL gene expression using piRNAs and miRNAs. However, given that some miRNAs and piRNAs can bind to mRNAs of several or even hundreds of human genes [43], it is necessary to identify human genes that may be affected by piRNAs and miRNAs that act on mRNAs of the KL gene. That is, it is necessary to identify the possible side effect of these piRNA and miRNA on the expression of all human genes if they are used as therapeutic drugs. To this end, we studied the possible binding of ID00756.3p-miR and nine piRNAs to 17,484 human genes.
The largest number of target genes was found for piR-3215034 and piR-6885965. In the mRNA of the AFF2 gene, for each of these piRNAs, eleven and ten BSs were found, respectively, located in one cluster (Figure 2). All BSs was located the 5′UTR through three nucleotides. The free energy of piR-3215034 binding to the mRNA of the AFF2 gene varied from −155 kJ/mol to −161 kJ/mol, and the ΔG/ΔGm value varied from 94% to 97%. During the piR-3215034 interaction with mRNA, canonical nucleotide pairs was involved in the last two BSs, except for two C-A bonds. These results indicate a high efficiency of piR-3215034 influence on AFF2 gene expression. piR-6885965 is 24 nts long and, therefore, the free energy of interaction with the mRNA of the AFF2 gene was lower. Since the BSs of these piRNAs are in the same cluster, piR-3215034 has the advantage of binding. However, at a significantly higher concentration of piR-6885965, it will have an advantage in binding over piR-3215034. Therefore, when determining the effectiveness of the action of piRNAs on translation, one should take into account the free energy of their interaction with mRNA and the concentration of competing piRNAs.
Since the AFF2 and KL genes are targets for piR-3215034 and piR-6885965, it is necessary to compare the possible effect of piRNAs on these genes. It should be noted that the AFF2 gene is involved in the development of squamous cell carcinoma [44], thoracic carcinoma [45], carcinomas of head and neck [46], and renal cell carcinoma and other diseases [47,48,49]. Therefore, by suppressing the oncogenesis caused by the AFF2 gene with piR-3215034 and piR-6885965, the expression of KL gene will be simultaneously suppressed. No other piRNAs interacting with mRNA of the AFF2 gene were identified. These results clearly demonstrate that, for each miRNA or piRNA, it is necessary to determine the expression of the specific genes they may influence.
Several publications have established the involvement of the BCL2L11 gene in oncogenesis [50,51]. For piR-3215034, 12 BSs were identified, which form a cluster of BSs from 55 nt to 114 nt, 60 nt long (Figure 3). piR-6885965 also binds in this BSs cluster, but with a ΔG/ΔGm value of less than 90%, which is below the selection criterion for significant piRNAs. No other piRNAs interacting with mRNA of the AFF2 gene were identified.
The CPT1A gene is involved in fatty acid metabolism and manifests its effect during oncogenesis, diabetes, and cardiomyopathy [52,53,54,55,56,57]. Figure 4 shows the interaction schemes of piR-3215034 and piR-6885965 with the mRNA of the CPT1A gene, which show that their BSs are located in the same cluster from 99 nt to 137 nt. piR-3215034 interacts with the mRNA of the CPT1A gene with a ΔG/ΔGm value of 99%, i.e., almost canonical base pairs are formed. No other piRNAs interacting with mRNA of the CPT1A gene were identified.
High expression of the DAZAP1 gene is observed in hepatocarcinoma and can serve as a prognostic marker of the disease. Knockdown of DAZAP1 small interfering RNA markedly inhibited proliferation, migration, and invasion of hepatocarcinoma cells [58,59]. piR-3215034 and piR-6885965 can repress DAZAP1 mRNA translation (Figure 5).
The results obtained indicate that these piRNAs can significantly influence the expression of the DAZAP1 gene (Figure 5). It should be noted that both piRNAs bind to the mRNA of the gene in the same cluster of BSs located in the 5′UTR, i.e., they can stop protein synthesis before the translation process. The DAZAP1 gene is highly expressed in the testis (RPKM 19.6), and appendix (RPKM 11.8). No other piRNAs interacting with mRNA of the DAZAP1 gene were identified.
A number of publications have shown the involvement of the NDRG3 gene in the development of oncogenesis, and in most cases, increased expression is observed in cancer of various organs [60,61,62,63,64,65]. Therefore, it is important to know whether piR-3215034 and piR-6885965 can suppress the expression of the NDRG3 gene.
No other piRNAs interacting with mRNA of the NDRG3 gene were identified. The results shown in Figure 6 indicate that NDRG3 gene expression can be downregulated by piR-3215034 and piR-6885965. No other piRNAs interacting with mRNA of the NDRG3 gene were identified.
The RHOT1 gene is involved in the modification of the development of breast cancer [66], pancreatic cancer [67,68], non-small cell lung cancer, hepatocellular carcinoma [69], and the risk and occurrence of Parkinson’s disease [70]. Figure 7 shows the interaction schemes of piR-3215034 and piR-6885965 with mRNA of the RHOT1 gene. Both piRNAs have eight BSs in the 5′UTR of the mRNA RHOT1 gene located in the same BSs cluster from the first nucleotide to 48 nt. The next BS was located after three nucleotides, and the free energy of interaction was the same in each of the sites for both piRNAs. Therefore, the value of ΔG/ΔGm was the same for each piRNA. No other piRNAs interacting with mRNA of the RHOT1 gene were identified.
SKIDA1 was significantly overexpressed in all molecular subgroups, except for only two subgroups of acute myeloid leukemia. In validation analyses, SKIDA1 was associated with higher sensitivity and specificity in acute myeloid leukemia. We highlight that SKIDA1 is one of the promising markers, which has consistent overexpression among several types of acute leukemia [71,72]. SKIDA1 associated with obesity [73]. For piR-3215034, 12 BSs were identified in the mRNA SKIDA1 gene (Figure 8). At two sites, piR-3215034 interacts with mRNA almost as complementary as possible, since the ΔG/ΔGm value is 99%. A cluster of piR-3215034 BSs with 61 nt in length guarantees the binding of two of these 27 nt piRNAs at once. No other piRNAs interacting with mRNA of the SKIDA1 gene were identified.
The WBP4 gene (synonymous name FBP21) is involved in splicing and therefore affects the maturation of the mRNA of many genes involved in metabolism [74]. Eight piR-3215034 and six piR-6885965 bind to the mRNA of the WBP4 gene (Figure 9). At position 94 nt, the 5′UTR of piR-3215034 binds only via canonical base pairs. piR-6885965 binds at the 94 nt position, also with a high ΔG/ΔGm value of 97%. No other piRNAs interacting with mRNA of the WBP4 gene were identified.
The ZIC5 gene acts as a transcriptional repressor. Increased expression of this gene is observed in various types of human cancer and may contribute to cancer progression [75,76,77]. Figure 10 shows the interaction schemes of piR-3215034 and piR-6885965 with the mRNA of the ZIC5 gene, which show a high degree of influence of piR-3215034 on translation. At four positions, the ΔG value is −159 kJ/mol and the ΔG/ΔGm ratio is 96% of the maximum value. For piR-6885965, there are only four BSs in the cluster with a ΔG/ΔGm value greater than 90%. No other piRNAs interacting with mRNA of the ZIC5 gene were identified.
The transcription factor encoded by the ZSWIM6 gene is synthesized in the brain and can affect the expression of a number of genes. Mutations in this gene lead to malformations of the brain [78,79]. Figure 11 shows the binding schemes of piR-3215034 and piR-6885965 to the mRNA of the ZSWIM6 gene. piR-3215034 had seven BSs forming a cluster and piR-6885965 had only three BSs in the same cluster. Transcription factors are difficult to study because the product of their activity can be diverse and difficult to control. The importance of their biological role is undoubted. No other piRNAs interacting with mRNA of the ZSWIM6 gene were identified.
The FGF23 gene is a member of a large family of fibroblast growth factors [80], which is most associated with the KL gene in anti-aging processes. A number of publications have examined the relationship of FGF23 and KL genes in physiological processes and in various diseases [81,82]. In this regard, we studied the possible effect of piRNA on the expression of the FGF23 gene. Only piR-576442, piR-1501557, piR-1845735, piR-2069834, and piR-3029987 could bind to the mRNA of the FGF23 gene, the interaction schemes of which are shown in Figure 12. All BSs of these piRNAs were located in the 3′UTR at a considerable distance from each other, i.e., BSs did not form BS clusters.
4. Discussion
We have shown that some piRNAs are capable of suppressing KL synthesis and can suppress the expression of other genes involved in the development of various diseases. Therefore, if the concentration of these piRNAs is reduced, KL expression may increase, while the expression of these disease-related genes may also change. To increase lifespan, selective reduction in piRNAs that specifically inhibit KL expression is required. Some authors call Klotho protein a hormone, others an anti-inflammatory agent. The latter function has reason to be because the composition of Klotho protein has an increased content of phenylalanine compared to conventional proteins and is comparable to antioxidant proteins.
Common piRNAs for several genes represent a pool of gene expression regulators and maintain the expression homeostasis of these target genes. A change in the expression of any of these genes will cause a redistribution of the degree of influence of the piRNAs on other genes. Therefore, to increase longevity it is necessary to reduce the concentration of only piRNA-6890096 which suppresses the expression of the KL gene with high selectivity. Some publications have identified an anti-inflammatory effect of the Klotho protein. The latter function has reason to be because the Klotho protein has an increased phenylalanine content (6%) compared to the phenylalanine content of conventional proteins and is comparable to antioxidant proteins.
To increase lifespan, it is necessary to reduce the concentration of those piRNAs that suppress the expression of only the KL gene with high selectivity. Such a piRNA is piRNA-6890096, which binds completely complementarily to the mRNA of the gene and practically does not affect the expression of other human genes. One way to selectively reduce the concentration of piRNA-6890096 is to introduce sponging RNA containing the binding sites of this piRNA. Such sponging of mRNAs in the annular conformation can reduce free piRNA-6890096. Many publications have shown that sponging RNAs effectively reduces the effect of mRNA. Of the 7310 miRNAs, only ID00756.3p-miR can bind to the mRNA of the KL gene. Consequently, piRNA can also be synthesized for it, which will specifically reduce the concentration in blood and cells. The piRNAs identified in this work that bind to mRNAs of protein-coding genes form associations of piRNAs and the target gene, which reflect a specific relationship between several piRNAs and genes. Such associations of piRNA and the target gene, depending on a particular disease, change in different ways. Therefore, such associations can be used as a diagnostic test. A decrease or increase in the concentration of piRNA, due to the high specificity of associations, will unequivocally indicate a disease. The obtained results indicate that it is possible to identify associations of piRNA and miRNA with the KL and FGF23 genes, which will allow for the regulation of their expression. To avoid the side effects of these piRNAs and miRNAs, it is proposed to use sponging RNAs capable of highly selectively binding to piRNAs and miRNAs that suppress KL and FGF23 gene expression.
The results of this study demonstrate that identifying the interaction of a single miRNA with the mRNA of a single gene does not ensure that this miRNA can be used to regulate only that gene. A given miRNA may regulate the expression of multiple genes, potentially leading to unintended or adverse effects. For example, an miRNA may suppress the expression of an oncogene while simultaneously inhibiting a tumor suppressor gene involved in another disease. Conversely, suppression of a tumor suppressor gene may promote the expression of other oncogenes or disease-associated genes. In this way, miRNAs can influence a wide range of physiological processes.
Therefore, it is essential to determine the effects of a specific miRNA on the expression of all genes within an organism. A comprehensive assessment of miRNA function requires evaluating its impact across the entire set of protein-coding genes. However, conducting such large-scale analyses using experimental (wet-lab) methods is costly and time-consuming. Computational approaches enable the systematic identification of miRNA interactions with all genes in an organism, significantly reducing both time and cost. Moreover, computational methods enable the quantitative characterization of miRNA interactions with specific mRNAs, providing more detailed insight into the interaction properties of different miRNA–mRNA pairs.
The mechanisms underlying piRNA–mRNA interactions are similar to those of miRNAs. More than eight million piRNAs are present in the human genome, which dramatically increases the complexity, time, and cost of experimentally characterizing their interactions with approximately one hundred thousand human gene isoforms. This challenge largely explains the limited current understanding of piRNA-mediated gene regulation. Evidence indicates that miRNA and piRNA expression levels change during ontogenesis, suggesting that these small RNAs play genome-wide regulatory roles in animals, including humans. Consequently, elucidating the global influence of miRNAs and piRNAs on human gene expression represents a major objective in modern biomedical research.
5. Conclusions
This computational analysis identified predicted interactions between piRNAs and the mRNAs of protein-coding genes, forming specific piRNA-target gene associations. These interactions suggest selective associations between multiple piRNAs and their predicted target genes, which may vary depending on the specific disease. Consequently, the identified interaction patterns represent potential candidates for further investigation in diagnostic research. Changes in piRNA abundance, together with the specificity of their predicted target interactions, may serve as molecular indicators of disease; however, these findings require validation through experimental and clinical studies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kuro-o M. Matsumura Y. Aizawa H. Kawaguchi H. Suga T. Utsugi T. Ohyama Y. Kurabayashi M. Kaname T. Kume E. Mutation of the Mouse Klotho Gene Leads to a Syndrome Resembling Ageing Nature 1997390455110.1038/362859363890 · doi ↗ · pubmed ↗
- 2Li S.-S. Sheng M. Sun Z.-Y. Liang Y. Yu L.-X. Liu Q.-F. Upstream and Downstream Regulators of Klotho Expression in Chronic Kidney Disease Metabolism 202314215553010.1016/j.metabol.2023.15553036868370 · doi ↗ · pubmed ↗
- 3Ni W. Zhang Y. Yin Z. The Protective Mechanism of Klotho Gene-Modified Bone Marrow Mesenchymal Stem Cells on Acute Kidney Injury Induced by Rhabdomyolysis Regen. Ther.20211825526710.1016/j.reth.2021.07.00334466631 PMC 8367782 · doi ↗ · pubmed ↗
- 4Jiang Y. Jiang W. Li Y. Gu W. Huang H. Wei Q. Bai G. Wang J. Rizak J.D. Zhou Z. Evaluation of Klotho Gene Expression and NGAL Levels Following Acute Kidney Injury during Pregnancy Hypertensive Disorders Pregnancy Hypertens 20223016117010.1016/j.preghy.2022.08.00836272327 · doi ↗ · pubmed ↗
- 5Xia J. Cao W. Epigenetic Modifications of Klotho Expression in Kidney Diseases J. Mol. Med.20219958159210.1007/s 00109-021-02044-833547909 · doi ↗ · pubmed ↗
- 6Iijima H. Gilmer G. Wang K. Bean A.C. He Y. Lin H. Tang W.-Y. Lamont D. Tai C. Ito A. Age-Related Matrix Stiffening Epigenetically Regulates α-Klotho Expression and Compromises Chondrocyte Integrity Nat. Commun.2023141810.1038/s 41467-022-35359-236627269 PMC 9832042 · doi ↗ · pubmed ↗
- 7Aziz M.S. Aamir A.-H. Khan A. Khan Z. Shah S.Q. Safi S.Z. Batumalaie K. Alobaid H.M. Ali A. Imran M. Investigation of Klotho G 395A and C 1818 T Polymorphisms and Their Association with Serum Glucose Level and Risk of Type 2 Diabetes Mellitus Genes 202213153210.3390/genes 1309153236140700 PMC 9498819 · doi ↗ · pubmed ↗
- 8Speer T. Schunk S.J. Klotho in Diabetic Kidney Disease: More than Dust in the Wnt Kidney Int.202210246947110.1016/j.kint.2022.05.01635988933 · doi ↗ · pubmed ↗