Application Method Determines Effects of Beauveria bassiana on Eucalyptus grandis Growth and Leaf-Cutting Ant Foraging
Raymyson Rhuryo de Sousa Queiroz, Thais Berçot Pontes Teodoro, Aline Teixeira Carolino, Ricardo de Oliveira Barbosa Bitencourt, Richard Ian Samuels

TL;DR
This study shows that applying the fungus Beauveria bassiana to eucalyptus plants can boost their growth and reduce ant damage, offering a greener alternative to chemical pesticides.
Contribution
The study demonstrates that foliar application of Beauveria bassiana is most effective in promoting eucalyptus growth and reducing ant foraging.
Findings
Foliar spraying of Beauveria bassiana significantly increased leaf number and shoot weight in eucalyptus seedlings.
The same treatment reduced leaf-cutting ant foraging activity by over 50%.
Soil drenching at sowing also enhanced plant height and shoot fresh weight.
Abstract
Leaf-cutting ants pose a severe threat to South American eucalyptus plantations, leading to significant economic losses. Traditional management relies on synthetic chemical baits, which are notorious for their environmental toxicity and long-term persistence in the soil. Consequently, alternative control strategies are urgently needed. While entomopathogenic fungi can be applied directly to ants or their nests, they also offer a novel approach to plant protection through plant tissue colonization. These fungi can act as endophytes, living within the plant, to provide protection and stimulate growth. However, the effectiveness of this protection and growth stimulation often depends on the inoculation method used. In this study, we tested three inoculation methods. The most promising results occurred when eucalyptus leaves were sprayed with fungal suspensions 20 days after planting. This…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
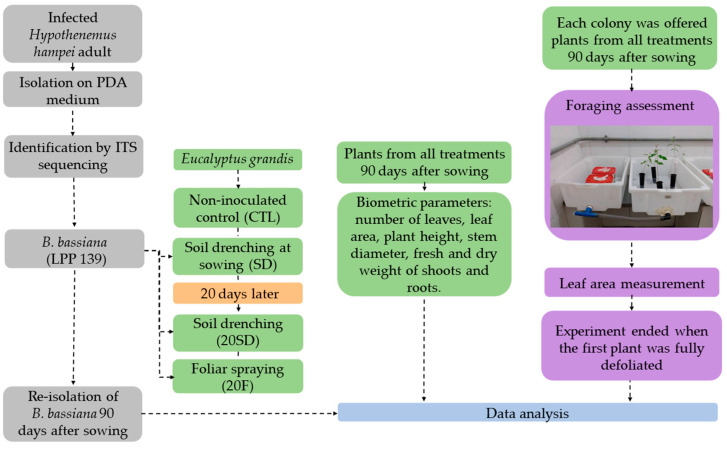 Figure 1
Figure 1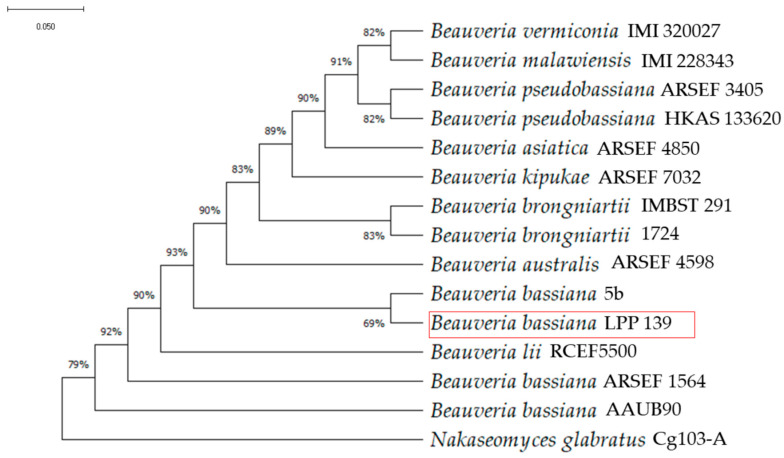 Figure 2
Figure 2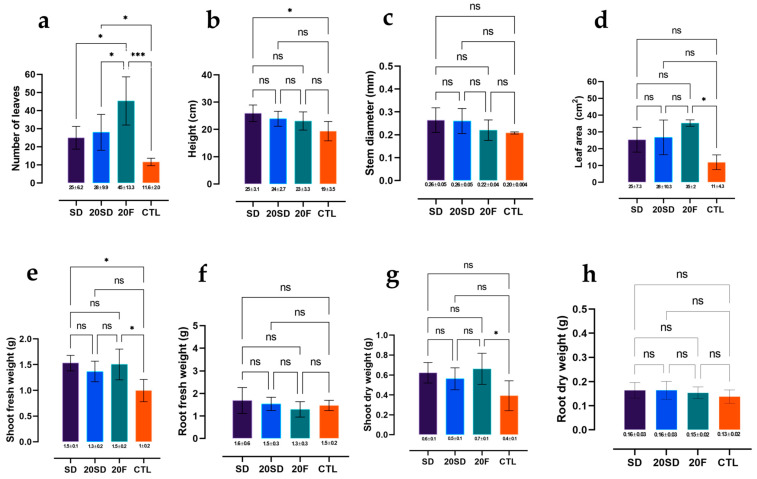 Figure 3
Figure 3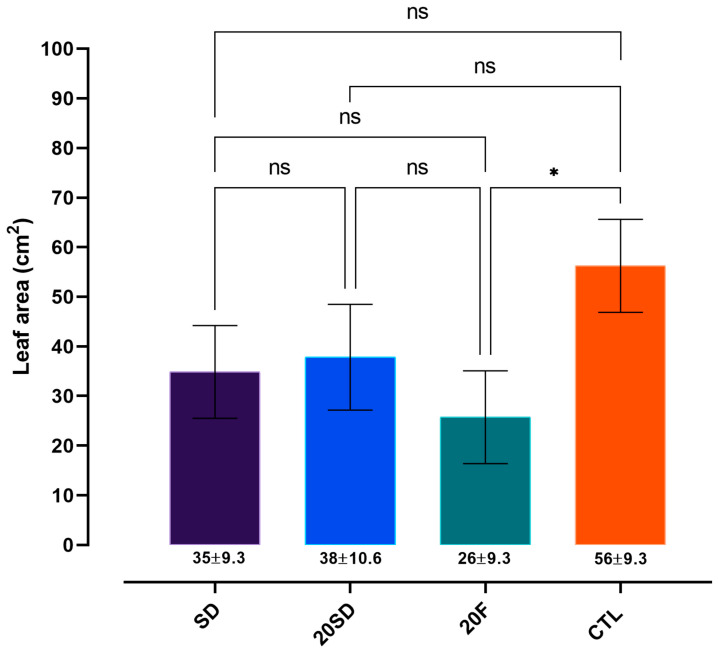 Figure 4
Figure 4- —Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ)
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
- —the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEntomopathogenic Microorganisms in Pest Control · Mycorrhizal Fungi and Plant Interactions · Fungal Biology and Applications
1. Introduction
Eucalyptus grandis Hill ex Maiden, commonly known as flooded gum or rose gum, is a tree species native to Australia [1], widely cultivated in tropical and subtropical regions due to its rapid growth, high productivity, and broad adaptation to different conditions [2,3]. Eucalyptus plantations represents a major component of Brazil’s forestry sector, supplying raw materials for construction, furniture, energy, and the pulp for paper industries [4]. As global demand for wood and wood-based products continues to rise, particularly with woodchip demand projected to grow around 40% by 2030 [5], the economic relevance of Eucalyptus plantations is expected to increase further. However, expanding production faces several challenges, including climate change [6], plant disease [7] and insect damage [8].
Among the most destructive pests in these forestry systems are leaf-cutting ants (Acromyrmex spp., Atta spp., Hymenoptera: Formicidae), which defoliate plants at all developmental stages, reducing biomass accumulation and overall productivity [9,10]. These ants harvest fresh leaves to cultivate a symbiotic fungus (Leucoagaricus gongylophorus) as their primary food source, making them highly efficient herbivores and one of the main obstacles to production in South America [11,12].
Chemical control, primarily utilizing sulfluramid-based baits, remains the most widely adopted strategy for managing leaf-cutting ants. This preference is attributed to its high efficacy, achieved through colony-level transmission once the insecticide is carried into the nest [13]. However, the synthesis of sulfluramid requires perfluorooctane sulfonyl fluoride (PFOSF), a known precursor of Persistent Organic Pollutants (POPs), and sulfluramid itself degrades in the environment, resulting in the formation of perfluorooctanesulfonic acid (PFOS). These compounds raise significant environmental concerns due to their persistence, bioaccumulation [14], toxicity to aquatic organisms [15] and mammals [16]. Consequently, global regulatory restrictions have been imposed on the use and production of sulfluramid [17].
In this context, biological control, particularly through the use of entomopathogenic and antagonistic fungi, has emerged as a promising and sustainable alternative for managing leaf-cutting ants and other agricultural pests. This strategy offers reduced environmental impact, low non-target toxicity, and potential for host specificity [18,19,20,21,22]. Compared to chemical insecticides, fungal-based control agents are considered environmentally safer because they cause less damage to the environment and non-target insects [23]. The entomopathogenic fungi (EPF) Beauveria spp., Metarhizium spp., Hirsutella spp., Lecanicillium spp., and Paecilomyces are the most widely studied genera for controlling various agricultural insect pests [23,24], including leaf-cutting ants [25,26].
Beyond their role as entomopathogens, several strains of these fungi are capable of colonizing plant tissues endophytically, providing dual benefits by promoting plant growth, enhancing resistance to biotic stress, and deterring herbivory [27,28,29,30]. Endophytic colonization typically occurs through the adhesion of fungal conidia to the plant surface, followed by germination and penetration via natural openings such as stomata, lenticels, root cracks, and germinating radicles, or through direct enzymatic degradation of the cell wall [31,32]. The promotion of plant development by endophytic Beauveria bassiana is associated with multiple physiological mechanisms, including improved nutrient uptake [33], modulation of phytohormone levels such as auxins and gibberellins [34], and stimulation of antioxidant enzyme activity [35].
As far as we know, there is little information about the cross-link between the endophytic activity of B. bassiana in Eucalyptus grandis and its impact on the foraging behavior of leaf-cutting ants in this plant species. Here, we studied the endophytic activity of B. bassiana in E. grandis via multiple inoculation methods, as well as the influence of treating plants with this fungus on plant growth parameters. Furthermore, the protective effect of endophytes on E. grandis was assessed when considering the foraging behavior of the leaf-cutting ant Acromyrmex subterraneus subterraneus.
2. Materials and Methods
2.1. Collection and Maintenance of Leaf-Cutting Ant Colonies
Six Acromyrmex subterraneus subterraneus nests were collected in the field from Bom Jardim city, Rio de Janeiro State, Brazil (22°09′07″ S and 42°25′10″ W). The nests were carefully excavated with the queens, workers, and their symbiotic fungus L. gongylophorus. The colonies were placed in 2L plastic containers and transported to the Myrmecology Unit of the Universidade Estadual do Norte Fluminense Darcy Ribeiro (UENF). The Acromyrmex species was identified according to [36]. The colonies were kept under controlled condition [12:12 h light:dark cycle, 26 ± 2 °C, and 70% ± 5% relative humidity (RH)]. The fungus L. gongylophorus was cultivated by the ants in plastic containers placed in trays and the sides of the trays were coated with talcum powder to prevent insect escape. Fresh Acalypha wilkesiana leaves were placed daily in the foraging arenas.
2.2. Fungus: Isolation and Molecular Characterization
Beauveria sp. LPP 139 was originally found infecting an adult coffee berry borer Hypothenemus hampei (Ferrari) (Coleoptera: Curculionidae, Scolytinae). To obtain a pure isolate of this fungus, insect-derived conidia derived from a moribund insect were collected directly with a sterile inoculation loop and transferred to Potato Dextrose Agar (PDA: KASVI, Curitiba, Brazil) medium supplemented with 0.05% chloramphenicol (Sigma, St. Louis, MO, USA) and incubated under controlled conditions (25 ± 2 °C, RH = 70%). After 15 days, the fungus was purified using a monosporic culture technique as described by [37].
For molecular characterization, approximately 50 mg of conidia were collected from PDA cultures and genomic DNA was extracted using the Quick-DNA™ Fungal/Bacterial Miniprep Kit (Zymo Research, Irvine, CA, USA) following the manufacturer’s instructions. DNA concentration and purity were assessed using a NanoDrop 2000 spectrophotometer (Thermo Scientific, Waltham, MA, USA). The internal transcribed spacer regions ITS-1 (TCCGTAGGTGAACCTGCGG) and ITS-4 (TCCTCCGCTTATTGATATGC) were employed for PCR amplification to facilitate fungal identification [38]. PCR reactions were prepared with 4.0 µL of dNTPs (1.25 mM), 5.0 µL of 5× buffer, 2.0 µL of MgCl_2_ (25 mM), 0.5 µL of Taq polymerase (5 U/µL), 1.0 µL of each primer (10 µM), 10.5 µL of ultrapure water, and 2.0 µL of DNA template (10 ng). Thermal cycling conditions consisted of an initial denaturation at 96 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 45 s, annealing at 55 °C for 60 s, and extension at 72 °C for 120 s, with a final extension at 72 °C for 10 min. PCR products were separated on 1% agarose gels, stained with GelRed (Biotium, Fremont, CA, USA), and visualized under UV illumination. Amplified fragments were purified using ExoSAP-IT PCR Product Cleanup Reagent (Applied Biosystems, Waltham, MA, USA) according to the manufacturer’s instructions, and subsequently sequenced by Sanger sequencing at ACTGene Análises Moleculares (Nova Alvorada, RS, Brazil).
The sequences obtained in this study were compared against 13 ITS sequences of various Beauveria species retrieved from the GenBank database (Table A1). Nakaseomyces glabratus (H. W. Anderson) Sugita & Takashima (Saccharomycetales: Saccharomycetaceae) was designated as the outgroup (Table A1). Chromatogram editing was performed using DNA Baser v.5. Sequence alignment was conducted through the Clustal W progressive method. A phylogenetic tree was generated using the maximum likelihood algorithm with nucleotide distances calculated according to the Kimura 2-parameter model. Tree robustness was evaluated via bootstrap analysis with 1000 replicates, employing the MEGA version 11 software [39].
2.3. Fungal Suspension
Beauveria sp. LPP 139 was cultivated on Petri dishes containing PDA and maintained controlled condition (25 ± 2 °C, RH = 70%). After 15 days, the conidial suspensions were prepared in sterilized distilled water containing 0.05% (v/v) Tween 80 (Quimiobrás Industrias Químicas SA, Rio de Janeiro, RJ, Brazil). For all experiments, the conidial suspension was quantified using a hemocytometer and adjusted to 1 × 10^8^ conidia/mL [40].
2.4. Treatment of Eucalyptus with Conidial Suspensions
Commercial Eucalyptus grandis seeds, LCFA 002 cultivar, obtained from Instituto de Pesquisa e Estudos Florestais (Piracicaba, Brazil), were surface-sterilized by immersion in 70% ethanol for 1 min, followed by 5% sodium hypochlorite with Tween 20. After 10 min, the seeds were washed three times with sterile distilled water [41]. For germination, the seeds were placed on moistened filter paper in Petri dishes. After six days, seedlings were transplanted into 100 cm^3^ tubes filled with commercial substrate Basaplant (Artur Nogueira, Brazil), previously autoclaved twice at 120 °C for 30 min.
Groups of E. grandis seeds/plants (n = 15 per group) were treated with the fungal suspension using three methodologies as follows: (1) soil drenching at sowing (SD); (2) soil drenching 20 days after sowing (20SD), and (3) foliar spraying 20 days after sowing (20F). For SD and 20SD, 2 mL of a conidial suspension was applied to the soil surface around the stem base of the plant. For 20F, 1.5 mL of a conidial suspension was manually sprayed onto each plant, which was the optimal volume determined to ensure adequate leaf coverage without causing runoff or drift of the suspension to the soil. Additionally, in the 20F treatment groups, the upper surface of the pots surrounding the plant stem was covered with aluminum foil prior to foliar spraying, thereby preventing contact between the soil and the fungal suspension. Following pulverization, transparent plastic bags were used to cover the plants for 24 h, in order to maintain humidity and facilitate fungal germination and colonization [42]. Plants were grown under shaded greenhouse conditions (50% shading) and irrigated as needed.
2.5. Confirmation of the Endophytic Colonization in Eucalyptus
After 90 days, five plants per group were randomly selected to assess endophytic colonization. Two g of stems, leaves, and roots were surface-washed, macerated, and serially diluted (up to 10^−2^) as described by [43] with modifications. Aliquots with 800 μL from each dilution were plated on PDA and incubated at 27 °C under a 12:12 h photoperiod. The growth of microorganisms was monitored daily, and re-isolations were performed to confirm the presence of B. bassiana following the protocol described by [44] with modification. Here, fungal growth was observed as discrete, circular colonies resembling colony forming units.
2.6. Biometric Parameters of Eucalyptus Treated with Conidial Suspension
After 90 days, biometric parameters were measured on Eucalyptus plants (N = 5 per group) exposed to the fungus. The number of leaves, leaf area (cm^2^), plant height (cm), stem diameter (mm), fresh and dry weight of shoots and roots (g) were measured. Fresh weights of the shoots and roots were taken immediately after the plants were collected from the greenhouse. Dry weights were determined after drying the plant material in a forced-air oven (Ethik 400D, Ethik Technology, Vargem Grande Paulista, SP, Brazil) at 70 °C for 24 h. The weight of the above-ground parts of the plant (i.e., leaves, stems) and roots were separately measured using an analytical balance (BEL^®^ M214A, BEL Engineering, Monza, Italy). In addition, leaf area was measured using a leaf area meter (LI-3100C LI-COR Biosciences, Lincoln, NE, USA).
2.7. Foraging Behavior Assessment
A foraging bioassay was conducted to evaluate the acceptance of treated plants by A. subterraneus subterraneus, according to [45]. The experiment followed a completely randomized design with four treatments (SD, 20SD, 20F and CTL), performed with three replicates, and conducted twice at different times, using a total of six colonies.
The experimental setup consisted of two interconnected arenas (60 cm length × 40 cm width × 18 cm height). One arena was used for foraging activity, where E. grandis seedlings were offered, while the second arena contained 4 L plastic containers with the ant colony and their symbiotic fungus garden. One seedling from each treatment was placed equidistantly within the foraging arena, allowing simultaneous choice and exposure to all treatments.
To ensure uniform foraging behavior, ants were deprived of leaves for 24 h prior to the experiment. The experiment was terminated once the ants had defoliated the first plant entirely, regardless of treatment, and this event was adopted as the endpoint criterion to ensure comparable foraging exposure among replicates. Leaf area was measured before and after exposure to the ants using an area meter (LI-3100C; LI-COR Biosciences, Lincoln, NE, USA).
2.8. Statistical Analysis
All data were submitted to Shapiro–Wilk normality test. Parametric data were subjected to one-way ANOVA followed by Tukey’s multiple comparisons test (p ≤ 0.05), and non-parametric data were submitted to the Kruskal–Wallis test followed by Dunn’s multiple comparisons test (p ≤ 0.05). All statistical analyses and graphical production were performed using GraphPad Prism 8.0.2. The complete experimental workflow, illustrating all methodological steps described above, is presented in Figure 1.
3. Results
3.1. Beauveria Isolate Identification
The ITS sequence obtained in this study was 492 bp in length, and a BLASTn search analysis revealed the highest nucleotide identity with Beauveria bassiana sequences, ranging from 91.98% to 99.22% identity and query coverage between 33% and 97%. The ITS sequence was deposited in GenBank under accession number PX591195. Also, a Bayesian phylogenetic analysis was performed, and the isolate grouped with B. bassiana reference sequences, confirming its taxonomic identity as B. bassiana (Figure 2).
3.2. Assessment of Endophytic Colonization
No fungal growth was observed from the aliquots of any of the evaluated samples obtained from plants at 90 days. Therefore, it was not possible to confirm systemic endophytic colonization using the currently applied technique.
3.3. Plant Biometric Parameters
Regarding the number of leaves (Figure 3a), comparing the groups SD vs. 20SD vs. 20F, the results were statistically different (p < 0.05) compared to each other. However, only SD had a similar result (p > 0.05) for leaf number compared with the control group. Furthermore, 20SD and 20F showed a statistical improvement in the number of leaves compared with control group (F_4,5_ = 11.05, p = 0.0419 and p = 0.0006, respectively, Figure 3a). Plants treated with fungus applied by soil drenching (SD) were significantly taller when compared with the control group (F_4,5_ = 3.415, p = 0.0353, Figure 3b).
Plants exposed to B. bassiana by foliar spray (20F) showed significantly enhancement in the leaf area compared with the control group (p = 0.0377, Figure 3d). The treatments SD and 20F significantly increased the shoot fresh weight compared with the control group (F_4,5_ = 5.110, p = 0.0154 and p = 0.0213, respectively, Figure 3e). Only plants exposed to fungus by foliar spraying presented an improvement in shoot dry weight compared with the control group (F_4,5_ = 4.090, p = 0.0236, Figure 3g). For stem diameter (Figure 3c), root fresh and dry weight (Figure 3f and h, respectively), no statistical differences (p > 0.05) were observed between all groups. Regardless of the treatment method (Figure 3b–h), no statistical differences (p > 0.05) were observed in any biometric parameters when comparing the groups treated with fungus (i.e., SD vs. 20 SD vs. 20F).
3.4. Ant Foraging Behavior
Plants exposed to B. bassiana by foliar spray (20F) showed a significant reduction in leaf area removal by A. subterraneus subterraneus compared with the control group (CTL) (F_4,3_ = 5.118, p = 0.0134, Figure 4). In contrast, no statistical differences (p > 0.05) were observed among the soil drenching treatments (SD and 20SD) or between these treatments and the control group. Similarly, no significant differences (p > 0.05) were detected among the treatments inoculated with the fungus (SD, 20SD, and 20F) (p > 0.05).
4. Discussion
Leaf-cutting ants are considered key herbivores in Neotropical terrestrial ecosystems, exerting strong ecological pressure and causing major economic losses in diverse agricultural systems [46]. Endophytic fungi have been increasingly recognized as potential allies in plant protection, as their presence can enhance host resistance and reduce herbivory [47]. Here, it was demonstrated that the application of B. bassiana to E. grandis seedlings can both promote plant growth and reduce damage to E. grandis caused by leaf-cutting ants, although these outcomes are critically dependent on the method of application. Our findings provide further evidence supporting EPF as multifunctional agents in integrated crop management.
Although endophytic colonization was not confirmed through re-isolation techniques, B. bassiana application prompted increases in the number of leaves, plant height, leaf area, shoot fresh and dry weight (Figure 3). The success of endophyte detection is highly dependent on the chosen methodology [48] and the inability to reisolate the fungus does not necessarily indicate the absence of colonization. Fungal density [49], uneven distribution within plant tissues [50], or competition with opportunistic fungi [51], can hinder recovery by cultivation-based methods. Indeed, it has been demonstrated that colonization rates of Beauveria spp. can vary significantly between cultivation-dependent (e.g., re-isolation) and cultivation-independent (e.g., PCR) methods [52].
Beneficial plant responses and negative effects on insect pests following fungal inoculation were reported, even when endophytic colonization was not assessed or confirmed [53]. In a comprehensive review, summarizing various cases in which the presence of the fungus inside plant tissues was not verified, significant physiological and ecological effects were still observed [53]. For instance, inoculation of B. bassiana in cotton resulted in reduced performance of Helicoverpa zea (Lepidoptera: Noctuidae) [54], while M. anisopliae applied to maize produced both enhanced plant growth and decreased the survival of Agriotes obscurus (Coleoptera: Elateridae) [55]. These findings suggest that the influence of entomopathogenic fungi on plant–insect interactions may occur even without explicit confirmation of endophytic colonization.
The improvements observed in several growth parameters are of significant agronomic relevance for E. grandis cultivation [56]. Among all treatments, plants subjected to foliar inoculation with B. bassiana (20F) exhibited the most pronounced increases in the number of leaves, leaf area, and shoot fresh and dry weight. This superior performance compared to both soil drenching treatments and the controls suggests that the mode of inoculation influences the plant–fungus interaction. Foliar application may promote a more direct and transient contact between fungal propagules and aerial plant tissues, which could influence physiological responses such as the activation of defense-related pathways or the modulation of growth hormones [57,58]. Furthermore*, B. bassiana* can solubilize and mobilize essential nutrients such as phosphorus, iron, and zinc in the rhizosphere, thereby improving nutrient uptake efficiency [33,59]. In addition, the fungus is capable of producing phytohormone-like compounds and secondary metabolites that modulate plant hormonal balance, including auxins and gibberellins [34]. These compounds can enhance root and shoot development, increase chlorophyll content and photosynthetic capacity [60].
Here, the significant rise in the number of leaves and leaf area in the 20F treatment expands the plant’s photosynthetic surface, which may enhance carbon assimilation and contribute to biomass accumulation and wood formation [60,61]. Additionally, increased shoot, height and biomass are recognized indicators of seedling vigor [62], and more vigorous seedlings generally exhibit higher survival and establishment rates under field conditions [63].
In our results, the reduced foraging observed on plants subjected to foliar inoculation (20F) may be associated with chemical and microbial cues rather than by direct pathogenic effects. Two complementary hypotheses can be proposed. First, the foliar application itself may have acted as a repellent treatment. Ants may have detected volatile or surface cues associated with the presence of B. bassiana, as foliar application results in a more direct deposition of fungal conidia on the leaf surface. Consequently, the leaves may have been perceived as ‘contaminated’, deterring the ants from cutting them. Leaf-cutting ants are known to exhibit a strong hygienic behaviors and avoidance of microbial contaminants to protect their symbiotic fungus garden, and even trace amounts of entomopathogenic fungi can elicit rejection behavior [64,65,66]. Second, the initial contact between the fungus and E. grandis tissues may have triggered physiological adjustments in the plant, leading to the production of deterrent volatile organic compounds (VOCs) or other chemical modifications. Certain VOCs induced by fungal endophytes possess antimicrobial properties [67], which could further discourage foraging by ants seeking to minimize microbial exposure within their colonies.
Such avoidance behavior is well documented: leaf-cutting ants rely on olfactory learning to discriminate against substrates harmful to their mutualistic fungus [68,69], a process supported by long-term memory and structural remodeling in the brain [70]. Information sharing within the colony is also highly efficient, with refuse dumps acting as “information centers” where foragers associate specific odors with contaminated or unsuitable materials [71].
Altogether, these findings suggest that B. bassiana exerts multifaceted effects on the E. grandis–ant system. The method of application modulates the primary outcome: soil drenching promotes growth and vigor, whereas foliar inoculation reduces herbivory (Figure 2 and Figure 3). Future work should focus on metabolomic and volatilome analyses to identify the specific compounds mediating these effects. Understanding this functional divergence is essential for developing tailored application strategies to optimize the use of B. bassiana in Eucalyptus cultivation.
5. Conclusions
This study demonstrated that the method of B. bassiana application is a key determinant of the functional outcome in the B. bassiana–E. grandis interaction. Soil drenching at sowing (SD) significantly increased the number of leaves, shoot height, and shoot fresh weight, indicating enhanced early vigor. Foliar application at 20 days after sowing (20F) promoted even broader effects, with significant increases in leaf number, leaf area, shoot fresh and dry weights, and also resulted in a marked reduction in leaf-cutting by A. subterraneus subterraneus. These findings showed that B. bassiana can simultaneously act as a plant growth promoter and an indirect biocontrol agent, and that the application method modulates the balance between these effects. Although these results provide valuable insights into the dual functionality of B. bassiana, further studies are needed to elucidate the mechanisms underlying its interaction with Eucalyptus plants, particularly regarding colonization dynamics, endophytic detection methods, and the physiological or chemical factors mediating ant deterrence. Such advances will be crucial for optimizing fungal application strategies and consolidating B. bassiana as a multifunctional tool in sustainable forestry systems.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Griffiths T. Forests of Ash: An Environmental History Cambridge University Press Cambridge, UK 20011248
- 2Sarre A. Global Forest Resources Assessment, 2020: Main Report Food and Agriculture Organization of the United Nations Rome, Italy 2020
- 3Kruse J. Turnbull T. Rennenberg H. Adams M.A. Plasticity of leaf respiratory and photosynthetic traits in Eucalyptus grandis and E. regnans grown under variable light and nitrogen availability Front. For. Glob. Change 20203510.3389/ffgc.2020.00005 · doi ↗
- 4IBA-Indústria Brasileira de Árvores Available online: https://www.iba.org/wp-content/uploads/2025/05/relatorio 2024.pdf(accessed on 21 October 2025)
- 5Lock P. Legg P. Whittle L. Black S. Global Outlook for Wood Markets to 2030: Projections of Future Production, Consumption and Trade Balance ABARES Report to Client Prepared for the Forest and Wood Products Australia Australian Bureau of Agricultural and Resource Economics and Sciences Canberra, Australia 202110.25814/76wa-v 824 · doi ↗
- 6Elli E.F. Sentelhas P.C. Bender F.D. Impacts and uncertainties of climate change projections on Eucalyptus plantations productivity across Brazil For. Ecol. Manag.202047411836510.1016/j.foreco.2020.118365 · doi ↗
- 7Almeida R.F. Machado P.S. Damacena M.B. Santos S.A. Guimarães L.M. Klopfenstein N.B. Alfenas A.C. A new, highly aggressive race of Austropuccinia psidii infects a widely planted, myrtle rust-resistant, eucalypt genotype in Brazil For. Pathol.202151 e 1267910.1111/efp.12679 · doi ↗
- 8Cavigliasso P. González E. Scherf A. Villacide J. Landscape configuration modulates the presence of leaf-cutting ants in eucalypt plantations Sci. Rep.2023131313010.1038/s 41598-023-40426-937573465 PMC 10423229 · doi ↗ · pubmed ↗