Exosomes and Triple-Negative Breast Cancer: Current Knowledge and Clinical Significance
Maria Loukopoulou, Anastasia Kottorou, Angelos Koutras, Foteinos-Ioannis Dimitrakopoulos

TL;DR
Exosomes may serve as non-invasive biomarkers for diagnosing and treating triple-negative breast cancer, but challenges remain in standardizing methods and understanding their biological roles.
Contribution
This systematic review highlights the potential of exosomal biomarkers in TNBC and emphasizes the need for standardized protocols and combinatorial biomarker panels.
Findings
Exosomal molecules show promise for early detection and tracking of TNBC progression.
Variations in methodology and sample diversity hinder generalization of findings.
Standardized protocols and combinatorial biomarker panels are needed for clinical translation.
Abstract
Exosomes, acting as vital mediators of cellular communication and carriers of diverse biomolecular cargo, are increasingly documented as important participants in cancer pathogenesis and progression. When it comes to triple-negative breast cancer (TNBC), a disease that comes with significant therapeutic hurdles, finding new, non-invasive biomarkers is absolutely crucial. This systematic review considers recent research, focusing on the role of exosomal biomarkers in diagnosing, predicting prognosis and foreseeing treatment response in TNBC patients. After an extensive search across PubMed and Google Scholar, we found many exosomal molecules showing great promise for early detection, tracking disease progression and tailoring treatments. This truly highlights liquid biopsy as a valuable, minimally invasive tool. However, there are still some big challenges to treat. These include…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
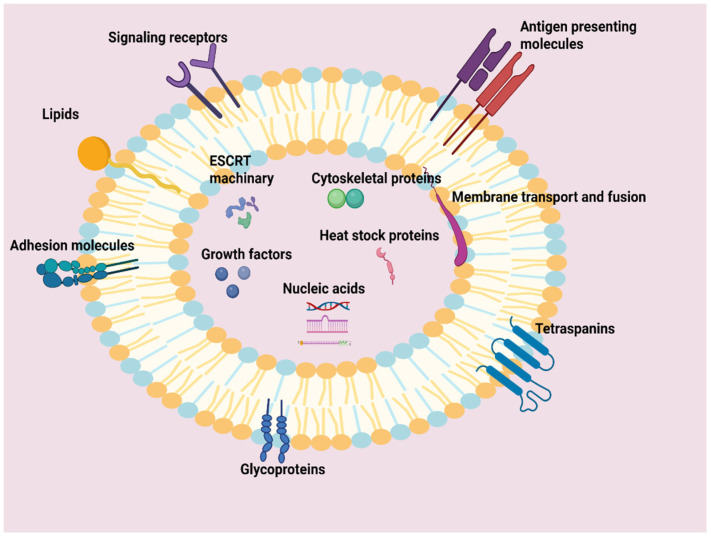 Figure 1
Figure 1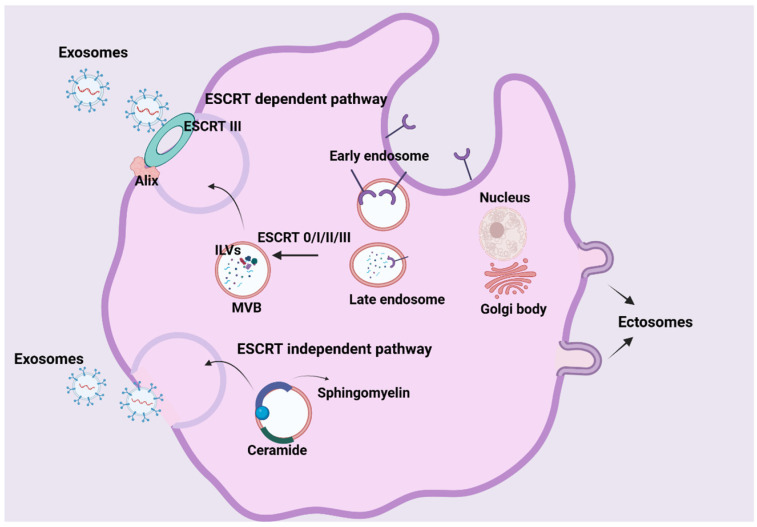 Figure 2
Figure 2|
| ||||
|
|
|
|
|
|
| miGISig | Plasma | 20 BC/10 HC | Elevated in plasma exosomes of Chinese BC patients | [ |
| miR-122-5p | Plasma | 257 BC/257 HC | Elevated in plasma exosomes of Chinese BC patients (38.9% TNBC) | [ |
| miR-3662 | Serum | 60 BC/20 HC | Increased levels in the serum of Chinese BC patients | [ |
| miR-146a | ||||
| miR-1290 | ||||
| miR-200c | Serum | 98 BC/45 benign/79 HC | Increased levels in the serum of Chinese BC patients | [ |
| miR-372 | Serum | 50 Invasive BC/12 HC | Increased levels in serum of German BC patients (21% TNBC) | [ |
| miR-92b-5p | Serum | 59 BC/53 HC | Elevated in Taiwanese BC patients vs. HC; correlated with disease stage | [ |
| miR-148a | Serum | 125 BC/50 benign/40 HC | Significantly reduced in Chinese BC patients | [ |
| miR-21 | Plasma | 16 BC/16 HC | Significantly higher in plasma exosomes of American BC patients | [ |
| miR-21 | Urine | 22 BC/26 HC | Significantly lower in Japanese BC patients than in controls | [ |
| miR-424 | Urine | 69 BC/40 HC | Highly specific/sensitive tool for distinguishing BC from HC in German patients (16 TNBC/69) | [ |
| miR-423 | ||||
| miR-660 | ||||
| let-7i | ||||
| miR-9 | Plasma | 62 BC/20 HC | Significantly elevated in early-stage South Korean BC patients (15/62) | [ |
| miR-16 | ||||
| miR-21 | ||||
| miR-429 | ||||
| miR-373 | Serum | 168 primary BC/19 benign/28 HC | Significantly elevated in German TNBC patients vs. other BC subtypes | [ |
| miR-376c | Serum | 224 TNBC/20 HC | Elevated in German TNBC patients relative to healthy controls | [ |
| miR-382 | ||||
| hsa-miR-148a-5p | Plasma | 27 BC/3 HC | Upregulated in Chinese TNBC patients vs. HC | [ |
| hsa-miR-200a-5p | ||||
| hsa-miR-210a-3p | ||||
| hsa-miR-378a-3p | ||||
| hsa-miR-483-5p | ||||
| hsa-miR-7110-5p | ||||
| hsa-miR-92b-3p | Downregulated in Chinese TNBC patients vs. HC | |||
| hsa-miR-150-5p | ||||
| miR-335-5p | Serum | 56 TNBC/HC not specified | Lower expression in Chinese TNBC patients than in HC | [ |
| miR-150-5p | Serum | 31 BC (16 LA. 15 TNBC)/16 HC | Accurately distinguished LA and TNBC subtypes in Brazilians | [ |
| hsa-miR-1180 | Tumor Tissue | 15 TNBC tumor/15 adjacent normal | Significantly elevated in Indian TNBC tumor tissue | [ |
| hsa-miR-4728 | ||||
| miR-21 | Serum | 85 TNBC/85 HC | Overexpressed in Indians TNBC patients; early detection biomarker | [ |
| Tissue | 85 TNBC/85 adjacent normal | |||
| miR-195 | Serum | 85 TNBC/85 HC | Downregulated in Indians TNBC patients; early detection biomarker | |
| Tissue | 85 TNBC/85 adjacent normal | Overexpressed in Indians TNBC patients; early detection biomarker | ||
|
| ||||
|
|
|
|
| |
| lncRNA XIST | Serum | 91 TNBC/50 HC | Distinguishes TNBC patients from HC in Chinese; decreased after tumor excision | [ |
| SUMO1P3 | Serum | 130 TNBC/60 non-TNBC/60 benign/50 HC | Higher in Chinese TNBC patients compared to non-TNBC, benign and HC | [ |
|
| ||||
|
|
|
|
| |
| circHSDL2 | Serum Exosomes and Tumor Tissue | 43 TNBC (serum)/Not specified HC (serum)/20 TNBC tumor | Significantly elevated in serum exosomes and tumor tissues of TNBC patients vs. HC | [ |
| circSIPA1L3 | Serum | 50 BC/50 HC | Significantly increased levels in the serum exosomes of Chinese BC patients | [ |
| circSTIL | Plasma | 59 TNBC/40 non-TNBC | Diagnostic value in distinguishing TNBC from non-TNBC subtypes in Chinese | [ |
| circPSMA1 | Serum and Cell Lines | 20 TNBC/20 non-TNBC | Increased expression in Chinese TNBC patients and TNBC cell lines vs. Non-TNBC patients/lines | [ |
|
| ||||
|
|
|
|
| |
| CD151 | Serum | 10 TNBC/17 HC | Higher expression in Chinese TNBC patients than in HC | [ |
| exo-AnxA2 | Serum | 169 BC/68 HC | Concentration significantly increased in serum samples from American BC patients vs. HC | [ |
| FLT1 | Serum | 56 TNBC/Not specified HC | Higher expression in Chinese TNBC patients than in HC | [ |
| GLUT-1 | BC Cell Line and non-tumorigenic breast cell line | Increased expression in BC cell lines | [ | |
| GPC-1 | ||||
| ADAM10 | ||||
|
| ||||
|
|
|
|
|
|
| miR-148a | Serum | 125 BC and 50 benign/40 HC | Lower expression in Chinese BC patients linked to worse OS and DFS | [ |
| miGISig | Plasma | 210 | Higher expression in Chinese BC patients linked to worse OS | [ |
| miR-4448 | Serum | 24 TNBC | High expression associated with longer OS in Japanese TNBC patients | [ |
| miR-2392 | ||||
| miR-2467 | ||||
| miR-4800 | ||||
| miR-21 | serum | 53 BC | Higher expression in metastatic Spanish BC patients | [ |
| miR-105 | ||||
| miR-335-5p | Serum | 56 TNBC | Low expression associated with worse OS in Chinese TNBC patients | [ |
| miR-637 | Serum | 20 TNBC | Low expression (combined with high Akt1) correlated with poor prognosis in Chinese TNBC patients with lymph node metastasis | [ |
| miR-939 | Tissue | 63 TNBC | High expression associated with recurrence risk in Italian breast cancer patients | [ |
|
| ||||
|
|
|
|
|
|
| XIST | Serum | 91 TNBC | High expression associated with worse OS in Chinese TNBC patients | [ |
| SUMO1P3 | Serum | 130 TNBC | High expression associated with worse OS in Chinese TNBC patients | [ |
| LINC00989 | Serum | 135 TNBC | Elevated in metastatic TNBC; combination with CEA, CA125 associated with shorter OS and PFS in Chinese TNBC patients | [ |
| LINC00899 | Plasma | 119 BC | Correlated with Ki-67 index, tumor size and presence/absence of lymph node metastasis | [ |
|
| ||||
|
|
|
|
|
|
| circSIPA1L3 | serum | 238 BC | Increased levels associated with shorter OS in Chinese BC patients. | [ |
| circSTIL | plasma | 49 TNBC | High expression associated with worse DFS in Chinese TNBC patients. | [ |
| circHSDL2 | serum | 43 TNBC | Promotes proliferation, infiltration and metastasis in TNBC cells. | [ |
| circMIB1 | exosomes from CAFs | 113 TNBC | Increased expression associated with shorter OS and DFS in Chinese TNBC patients. | [ |
|
| ||||
|
|
|
|
|
|
| FLT1 | serum | 56 TNBC | High expression associated with worse OS in Chinese TNBC patients. | [ |
| Akt1 | serum | 20 TNBC | High expression of Akt1 combined with low expression of miR-637 correlated with poor prognosis in Chinese TNBC patients with lymph node metastasis. | [ |
| LDHC | serum | 75 BC | Higher levels in Chinese preoperative breast cancer patients than in postoperative patients. | [ |
| AnxA2 | serum | 169 TNBC | Higher levels associated with shorter OS and worse DFS in Americans and higher levels in TNBC patients with Grade III compared to Grade II. | [ |
| EV_APRIL | serum | 190 TNBC | Higher levels in Korean TNBC patients associated with shorter OS. | [ |
| EV_CXCL13 | ||||
| EV_VEGF-A | ||||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsExtracellular vesicles in disease · Ovarian cancer diagnosis and treatment · Biomarkers in Disease Mechanisms
1. Exosomes: Silent Messengers of Cellular Communication
At the heart of cellular communication and complex pathological interplays, exosomes seem to take on the role of tiny messengers capable of revolutionizing our understanding of intercellular dialogs [1]. Exosomes are nanovesicles that possess a spherical morphology, ranging between 30 and 150 nanometers, encased within a bilayer phospholipid membrane similar to typical cellular membranes [2].
Exosomes have been isolated from all body fluids (e.g., blood, plasma, cerebrospinal fluid, breast milk and even urine), making them easily accessible for clinical study [3]. In exosome research, plasma and serum are the most commonly used sources of blood-derived samples. The choice between them can have a significant impact on exosome yield, purity and the profile of molecular biomarkers [4]. Plasma is generally preferred because clotting during serum preparation activates platelets, leading to the release of additional extracellular vesicles that may contaminate the exosome fraction [5]. These differences can affect both the reproducibility and interpretation of results, emphasizing the importance of careful sample selection and standardized collection protocols, particularly when aiming for clinical applications [6].
These nanovesicles are secreted by cells dynamically and continuously under normal physiological conditions as well as when the state of a cell is compromised. This means they are uniquely capable of mirroring the state of the parent cell [7]. Through these vesicles, valuable information travels to distant cells and tissues, contributing to the organism’s homeostasis. However, when this equilibrium is disturbed, as in cancer, they can also foster pathological processes [8].
Exosomes invariably contain a remarkably complex and diverse “cargo” depending on their functional versatility (Figure 1). The categories of this cargo are mainly proteins, nucleic acids and lipids, each responsible for a specific function or set of functions [9].
The proteins inside exosomes exhibit considerable diversity. Some proteins are universal in all exosomes and function as their “identity markers,” such as tetraspanins CD9, CD63 and CD81, RAB GTPases, Alix and HSP60/90 (heat shock proteins) [10,11]. Other proteins are more specific and indicate the type of cell comprising them, such as MHC (major histocompatibility complex) Class I and II, responsible for antigen presentation [12]. Exosomes can also carry membrane-bound signaling receptors like FasL (Fas ligand) and TNFR (tumor necrosis factor receptor), which participate in cell death pathways. Other molecules include adhesion molecules like integrins, which mediate attachment to the extracellular matrix and influence cell adhesion and migration. The diverse transport and communication functions of exosomes are the result of the cooperation of glycoproteins involved in cell communication and recognition, of cytoskeletal proteins belonging to the ESCRT (Endosomal Sorting Complexes Required for Transport) system, of membrane transport and fusion proteins, of growth factors and cytokines [13,14].
Regarding nucleic acids, exosomes act as containers storing genetic information. Exosomes can carry many types of RNA, including mRNA, microRNA (miRNA), lncRNA (long non-coding RNAs), tRNA, snRNA (small nuclear RNA), snoRNA (small nucleolar RNA) and circRNA (circular RNAs) [15]. Among these, miRNAs are the most abundant in number and are essential in the regulation of gene expression that affects processes such as hematopoiesis, angiogenesis and exocytosis. The mRNAs conveyed by them are coding for protein synthesis, making exosomes a pronounced means of transferring genetic instructions to distant cells that may contribute to protein replenishment, cell differentiation, or microenvironment modification [16,17]. Moreover, exosomes also carry PIWI-interacting RNAs (piRNAs). These non-coding RNAs play a vital role in safeguarding the genome from transposable elements, thereby ensuring genetic stability and notably influencing tumor development [18,19].
Finally, the exosomal membrane bilayer is exceptionally rich in lipids, including sphingomyelin, cholesterol and ceramides. These are not just structural components; they are crucial for maintaining the exosomal structure’s stability, facilitating their secretion and ensuring the precise sorting of their internal cargo. Furthermore, they actively contribute to intercellular signaling [20].
2. Exosome Biogenesis: A Complex Cellular Process
The production of exosomes is a strictly regulated and dynamic process, guided by specific molecular pathways. Understanding the mechanisms of exosome biogenesis and their composition is of vital importance, as it illuminates their functional activities and paves the way for future clinical applications [21,22].
Exosome formation begins with endocytosis at the plasma membrane’s surface, leading to the creation of early endosomes (Figure 2) [23,24]. These early endosomes function as sorting stations for internalized cargo, maturing and transforming into late endosomes. At this stage, they form multiple inward membrane invaginations that encapsulate specific proteins, nucleic acids and other substances, thereby generating intraluminal vesicles (ILVs), the precursors of exosomes. Subsequently, late endosomes containing multiple ILVs develop into multivesicular bodies (MVBs) [25,26].
The fate of MVBs varies. While some MVBs fuse with lysosomes for the degradation of their contents, others are destined for the secretion of ILVs into the extracellular space (Figure 2) [27]. These released ILVs are then termed exosomes. The secretion of MVBs from the cell involves three key steps: targeted transport of MVBs, tethering of MVBs to the plasma membrane, and fusion of the MVB’s limiting membrane with the plasma membrane (Figure 2) [25]. The proper functioning of this entire process relies on the surface proteins of the MVB [28].
The biogenesis of ILVs and MVBs is primarily guided by two main and distinct, yet sometimes overlapping, mechanisms: the ESCRT-dependent and the ESCRT-independent pathways (Figure 2) [29].
ESCRT-dependent pathway: This intricate pathway is precisely orchestrated by a large multiprotein complex comprising around thirty different types of proteins. The core function of the ESCRT machinery is to selectively sort specific components into ILVs. The ESCRT system itself is made up of four main complexes—ESCRT-0, ESCRT-I, ESCRT-II and ESCRT-III—each playing a unique and vital role [30].
ESCRT-0: It is responsible for cargo aggregation in a ubiquitination-dependent process, recognizing ubiquitinated cargo proteins. It contains the HRS (Hepatocyte Growth Factor-Regulated Tyrosine Kinase Substrate) protein, which can interact with STAM.
ESCRT-I and ESCRT-II: They function synergistically to induce the encapsulation of specific molecules from the endosomal membrane through budding. HRS recruits TSG101 to ESCRT-I and then ESCRT-I recruits ESCRT-III via ESCRT-II or ALIX.
ESCRT-III: It drives vesicle scission and forms a filamentous structure that promotes the inward budding and fission of ILVs from the endosomal membrane. The ESCRT-III complex is disassembled by the VPS4 complex and is subsequently recycled.
ESCRT-associated proteins: Proteins such as ALIX and VPS4 also contribute to MVB biogenesis by regulating cargo sorting, membrane remodeling and ESCRT disassembly. The ALIX protein can interact with syndecan-syntenin and participate in regulating exosome formation [10,30,31].
b.ESCRT-independent pathway: In addition to the ESCRT-dependent pathway, ILVs and MVBs can also be generated by ESCRT-independent mechanisms (Figure 2). These mechanisms involve lipids, such as sphingomyelin, cholesterol and ceramides, as well as tetraspanins and HSPs [26]. Exosomes are rich in these lipids, whose composition resembles that of membrane lipid rafts. Evidence suggests that lipid raft components play fundamental roles in ESCRT-independent ILV formation [26,32,33].
It is important to note that the ESCRT-dependent and ESCRT-independent pathways are not mutually exclusive. They can even operate within the same MVB, leading to the production of distinct ILV populations enriched with different cargo. This suggests that the pathways may intersect to some extent, which can depend on the cell type, its environment and its cargo [34].
3. Exosomes: Important Contributors in Triple-Negative Breast Cancer Pathogenesis
Triple-Negative Breast Cancer (TNBC) is a particularly aggressive and heterogeneous subtype of breast cancer. It is characterized by the absence of estrogen receptor (ER), progesterone receptor (PR) and HER2 expression, which drastically limits available targeted therapies [35]. This clinical challenge makes it imperative to delve deeper into the underlying molecular mechanisms of the disease, enabling the development of new, more effective therapeutic strategies [36].
In light of this imperative need, exosomes have emerged as important factors in the pathobiology of many tumor types [37,38,39], and recently, of TNBC. Through their ability to transfer a wide range of biomolecules and influence cell-to-cell signaling, they actively participate in shaping the tumor microenvironment (TME) and regulating the behavior of cancer cells at every stage of the disease [40,41].
Specifically, exosomes, through proteins and non-coding RNAs, actively contribute to tumor growth by enhancing cancer cell proliferation, inflammation and angiogenesis [42]. Exosomes play a significant role in TNBC metastasis, encompassing both local invasion and the establishment of distant metastatic niches. Exosomes promote epithelial–mesenchymal transition (EMT), a critical step for cancer cell invasion and dissemination [43,44]. Furthermore, by transferring biomolecules from their cells of origin, they can target specific remote tissues and create a “permissive” pre-metastatic environment, thereby facilitating the engraftment and growth of metastatic cells. A characteristic example is the involvement of exosomal integrins in predicting and shaping these metastatic sites [45,46].
Moreover, exosomes are a significant factor in the development of drug resistance, a major obstacle to effective TNBC treatment [47]. Although chemotherapy remains the primary therapeutic approach, a significant percentage of patients (approximately 70%) do not achieve a complete response and in some cases, chemotherapy-induced metastasis is even observed [48]. The development of chemoresistance by cancer cells is a complex process involving multiple mechanisms [49]. In this context, exosomes emerge as critical factors, as they actively participate in intercellular communication within the tumor microenvironment [50]. Experimental evidence shows that exosomes derived from chemoresistant breast cancer cells can transfer resistance-associated molecular cargo, including miRNAs and lncRNAs, to drug-sensitive cells, thereby modulating drug response pathways and promoting chemoresistant phenotypes in recipient cells [51,52]. Specifically, three main exosomal mechanisms contribute to increased chemotherapeutic drug efflux: the direct efflux of drugs from cells, the transfer of membrane-embedded drug efflux pumps and the transfer of drug-metabolizing enzymes, leading to their deactivation [53].
4. Exosomal Biomarkers in ΤΝBC
TNBC, as an aggressive subtype of breast cancer with limited therapeutic options, highlights the urgent need for reliable non-invasive biomarkers [54]. In this context, exosomes represent one promising approach among several emerging tools. Due to their ability to carry a dynamic cargo of biomolecules that reflects the molecular state of the tumor, they have been investigated as potential tools to support early diagnosis, provide prognostic information on disease progression and help predict treatment response, contributing decisively to personalized medicine in TNBC [55].
4.1. Exosomes as Diagnostic Biomarkers in TNBC
Early and non-invasive detection of TNBC is crucial for improving patient outcomes. Exosomes offer a unique molecular mirror of the tumor and thus, various studies have identified diverse exosomal biomolecules with diagnostic value in TNBC [56,57].
4.2. Exosomal microRNAs (miRNAs) as Diagnostic Biomarkers in TNBC
Exosomal miRNAs are key regulators of gene expression transported via exosomes, making them ideal candidates for non-invasive biomarkers [58]. Numerous studies have investigated their diagnostic value in TΝBC (Table 1a). Diverse exosomal miRNAs with a potential diagnostic role have been identified in breast cancer patients, irrespective of subtype. Specifically, the gastrointestinal system-derived miRNA (miGISig) and miR-122-5p were observed to be elevated in the plasma exosomes of patients, demonstrating effective diagnostic accuracy [59,60]. Similarly, increased levels of exosomal miR-3662, miR-146a, miR-1290, miR-200c and miR-372 were found in the serum of BC patients [61,62,63]. Exosomal miR-92b-5p in serum was observed to be elevated in BC patients compared with healthy controls and was correlated with disease stage [64], while exosomal miR-148a in serum was found to be significantly reduced in BC patients [65].
Of particular interest is miR-21, which is strongly recommended for breast cancer screening, and its diagnostic value in breast cancer has been confirmed by meta-analyses, suggesting improved accuracy through its combination with other miRNAs [66]. Indeed, miR-1246 and miR-21 were detected at significantly higher levels in the plasma exosomes of patients, with their combination serving as a superior diagnostic marker compared to their individual levels [67]. However, it is noteworthy that another study found the expression of miR-21 in urinary exosomes to be significantly lower in breast cancer patients than in controls [68]. Although both studies employed qRT-PCR for miR-21 quantification, the discrepancy in results likely arises from the different exosome sources (plasma vs. urine), underscoring the impact of sample type on exosomal biomarker levels.
Additionally, diagnostic accuracy is enhanced by the use of biomarker panels. One such panel, consisting of four urinary microRNAs (miR-424, miR-423, miR-660 and let7-i), was identified as a highly specific and sensitive tool for distinguishing breast cancer patients from healthy controls [69]. Furthermore, a panel of four exosomal miRNAs (miR-9, miR-16, miR-21 and miR-429) was found to be significantly elevated in early-stage breast cancer patients, irrespective of subtype, compared to healthy controls [70].
Beyond these general findings in breast cancer, numerous exosomal miRNAs have been specifically identified for their diagnostic role in TNBC. For instance, serum levels of sEV-miR-373 have been found significantly elevated in TNBC patients compared to other breast cancer subtypes [61]. Similarly, significantly higher levels of plasma exosomal miR-376c and miR-382 have been observed in TNBC patients relative to healthy controls [71]. A comprehensive study identified six upregulated miRNAs (hsa-miR-148a-5p, hsa-miR-200a-5p, hsa-miR-210a-3p, hsa-miR-378a-3p, hsa-miR-483-5p and hsa-miR-7110-5p) and two downregulated miRNAs (hsa-miR-92b-3p and hsa-miR-150-5p) in the plasma exosomes of TNBC patients versus healthy controls [72]. Additionally, the expression of miR-335-5p in the serum exosomes of TNBC patients has been found to be lower, while FLT1 was higher than in the control group [73]. Notably, miR-150-5p accurately distinguished Luminal A and TNBC subtypes [74]. Additionally, two exosomal oncomiRs, hsa-miR-1180 and hsa-miR-4728, were found to be significantly elevated in tumor tissue samples from TNBC patients [75]. Finally, a panel of six miRNAs (miR-21, miR-221, miR-210, miR-195, miR-145 and Let-7a) was proposed as a minimally invasive biomarker for the early detection of TNBC, with miR-21, miR-221 and miR-210 showing significant overexpression and miR-195 and miR-145 being downregulated, all correlating well with various clinicopathological and demographic factors [76].
4.3. Exosomal circRNAs and lncRNAs in Diagnosis of TNBC
CircRNAs and exosomal lncRNAs can also offer valuable information for the early diagnosis and monitoring of TNBC (Table 1b,c). The expression of circHSDL2 was significantly elevated in serum exosomes and tumor tissues of TNBC patients compared to healthy controls [77]. Concurrently, circSIPA1L3 has been recognized with high diagnostic value, showing significantly increased levels in serum exosomes of breast cancer patients more generally [78]. Correspondingly, the expression level of circSTIL in patients’ plasma exosomes showed potential diagnostic value in distinguishing TNBC from non-TNBC subtypes [79]. Furthermore, higher expression of circPSMA1 was observed in serum exosome samples from TNBC patients, as well as in TNBC cell lines, compared to non-TNBC patients and cell lines [80].
Regarding lncRNAs, lncRNA XIST in serum has emerged as a strong diagnostic indicator. Serum exo-XIST levels were capable of distinguishing TNBC patients from the healthy control group. Interestingly, these levels significantly decreased after the excision of primary breast tumors, indicating their association with tumor presence [81]. Additionally, the expression level of lncRNA SUMO1P3 in serum exosomes was significantly higher in TNBC patients compared to non-TNBC patients, patients with benign breast disease and healthy controls [82].
4.4. Exosomal Proteins as Diagnostic Biomarkers in TNBC
Exosomal protein cargo represents another promising category of biomarkers for TNBC diagnosis and several studies have highlighted the diagnostic value of some of them (Table 1d). Expression levels of the exosome tetraspanin CD151 in a serum derived from TNBC were significantly higher than those from healthy individuals [83]. Furthermore, serum exo-AnxA2 levels could serve as a suitable diagnostic tool for aggressive breast cancer, especially in TNBC patients; the concentration of exo-AnxA2 was also significantly increased in serum samples from breast cancer patients compared to healthy individuals [84]. Lastly, three exosomal membrane/surface proteins—glucose transporter 1 (GLUT-1), glypican 1 (GPC-1) and ADAM metallopeptidase domain 10 (ADAM10)—were recognized as potential breast cancer biomarkers in a study conducted on cell lines [85].
5. Exosomes as Prognostic Indicators in TNBC
Beyond their diagnostic role, exosomes are emerging as reliable prognostic indicators in TNBC. Their study allows for the prediction of recurrence, disease progression, the occurrence of metastases and estimates patients’ overall survival (OS) and disease-free survival (DFS).
5.1. Exosomal miRNAs as Prognostic Indicators in TNBC
Decreased expression of exosomal miR-148a in serum and exosomal miGISig in plasma has been closely linked to an unfavorable clinical outcome in breast cancer patients (Table 2a) [59,65]. Conversely, high expression of a panel of four miRNAs—miR-4448, miR-2392, miR-2467 and miR-4800—in serum was associated with longer OS (overall survival) in TNBC patients [86]. Εxosomal miRNA-21 and miR-105 expression levels in serum were higher in metastatic BC patients compared to non-metastatic patients and healthy controls [87].
Furthermore, the expression of miR-335-5p in serum exosomes was related to histological grade, differentiation degree and lymph node metastasis. Specifically, low expression of miR-335-5p was associated with significantly shorter OS in TNBC patients, while high expression of its target gene, FLT1, was linked to a lower survival rate [73]. Additionally, high expression of Akt1 combined with low expression of mir-637 in serum exosomes was strongly correlated with poor prognosis in TNBC patients with lymph node metastasis [80]. Finally, miR-939 in tissue exosomes emerged as a prognostic factor for DFS, particularly in combination with lymph node involvement (N), multifocal occurrence and tumor size, suggesting a synergistic risk of recurrence in patients with high miR-939 expression and lymph node involvement [88].
5.2. Exosomal lncRNAs and circRNAs as Prognostic Indicators in TNBC
Several studies have associated the increased expression of specific circRNAs with an unfavorable prognosis and more aggressive TNBC behavior (Table 2c). Specifically, the increased expression of exosomal circSIPA1L3 in serum was associated with shorter OS in BC patients [78]. Similarly, high expression levels of exosomal circSTIL in plasma were correlated with worse DFS in TNBC patients, although no association with OS was found [89]. Furthermore, exosomal circHSDL2 in the serum of TNBC patients appeared to promote proliferation, infiltration and metastasis in TNBC cells, while the increased expression of exosomal circMIB1, derived from CAFs (cancer-associated fibroblasts), was significantly associated with shorter OS and DFS in TNBC patients [77,90].
Another significant prognostic biomarker is the exosomal lncRNA XIST, whose expression levels have been shown to be significantly higher in patients with recurrent TNBC compared to those with non-recurrent TNBC (Table 2b). High expression of exo-XIST in serum was associated with worse OS, while its expression significantly decreased after primary tumor excision and increased after recurrence, indicating that exo-XIST can serve as a non-invasive biomarker for predicting the progression of TNBC patients [81].
Additionally, lncRNA SUMO1P3 in serum exosomes was closely associated with vascular/lymphatic invasion, lymph node metastasis and histological grade. Patients with high levels of lncRNA SUMO1P3 in serum exosomes had worse OS compared to those with low levels [82]. Similarly, exosomal lncRNA LINC00989 in serum was significantly elevated in patients with metastatic TNBC [91]. Higher levels of this biomarker, often in combination with clinical markers such as CEA and CA125, were significantly associated with shorter OS and PFS, establishing it as an independent prognostic factor [91]. Finally, LINC00899 was found to be reduced in plasma exosomes and breast cancer cell lines, correlating with the Ki-67 index, tumor size and the presence or absence of lymph node metastasis [89].
5.3. Exosomal Proteins as Prognostic Indicators in TNBC
Additionally, the level of exosomal LDHC (Lactate Dehydrogenase C) in the serum of preoperative breast cancer patients was significantly higher than in postoperative patients (Table 2d). Furthermore, in patients with recurrent disease, the level of exosomal LDHC was elevated, suggesting that exosomal LDHC in serum could be a promising indicator for effective monitoring and prediction of breast cancer recurrence [92]. The exosomal protein AnxA2 (exo-AnxA2) has shown significant prognostic value. Exo-AnxA2 levels were significantly higher in serum samples from TNBC patients with grade III compared to grade II patients. Breast cancer patients with high exo-AnxA2 levels in serum had significantly shorter OS and worse DFS. Moreover, the association of exo-AnxA2 with African American TNBC patients suggests its potential role as a factor contributing to TNBC aggressiveness in these women [84]. Finally, one study showed that high levels of the exosomal proteins EV_APRIL, EV_CXCL13 and EV_VEGF-A in the serum of TNBC patients were associated with significantly shorter OS. Multi-parametric analysis even identified the high level of EV_CXCL13 as an independent prognostic factor for poor OS [93].
6. Exosomal Biomarkers as Predictive Indicators in TNBC
Beyond their diagnostic and prognostic roles, exosomes and their biomolecular content may also serve as predictive biomarkers in TNBC. Their ability to dynamically reflect tumor response to various therapeutic approaches, such as chemotherapy and immunotherapy, offers an invaluable tool for treatment individualization. By studying exosomes, researchers aim to identify patients more likely to respond to specific therapies, enable early recognition of treatment resistance and monitor minimal residual disease, ultimately leading to more effective and targeted interventions [94].
6.1. Overall Exosome Concentration and Treatment Response
The overall concentration of extracellular vesicles (EVs) in plasma, among which exosomes are particularly critical, has emerged as a significant predictive indicator for the response of BC patients to neoadjuvant chemotherapy (NACT). More specifically, one study showed that EV concentration was 40 times higher in BC patients compared to healthy controls, and that an increase in its concentration after NACT was associated with treatment failure, reduced three-year DFS, and OS. Consequently, this study suggests that plasma EV concentration in BC patients may reflect the status of minimal residual disease (MRD), as well as treatment response [95].
6.2. Exosomal RNA as Predictive Indicators for Chemotherapy Response
At the level of lncRNAs, SUMO1P3 levels in serum exosomes significantly decreased in cases of chemosensitivity, while remaining unchanged in cases of chemoresistance in TNBC patients (Table 3a) [82]. Correspondingly, LINC00899 in plasma exosomes demonstrated high predictive value for pathological outcome in TNBC patients receiving neoadjuvant treatment, indicating its potential to predict chemotherapy efficacy [89]. Similarly, the expression of circSTIL from plasma exosomes significantly increased in TNBC patients without NAC compared to patients who received NAC [79].
Regarding miRNAs, several have emerged as predictive indicators for response to neoadjuvant chemotherapy (NAC). Specifically, the expression of miR-4448 and miR-2392 in serum was significantly higher in patients with pathological complete response (pCR) compared to those with non-pCR. A combined model of these two miRNAs, along with miR-2467-3p and miR-4800-3p, is reported to accurately distinguish between pCR and non-pCR patients in TNBC [86]. Likewise, the upregulation of plasma miR-127 was associated with pCR in BC. While after the first dose of NACT, high expression of miR-141 in plasma was associated with pCR, miR-34a, exo-miR182 and exo-miR-183 predicted non-pCR [96].
Furthermore, the plasma exosomal expression levels of miR-185, miR-4283, miR-5008 and miR-3613 were found to be lower in TNBC patients with no NACT response, whereas miR-1302, miR-4715 and miR-3144 were higher [97]. MiR-770 was highly expressed in chemosensitive tissues and predicted a better prognosis for TNBC, indicating its significant role in regulating chemoresistance and disease progression [98]. Additionally, hsa-miR-6831-5p in serum showed a significant difference in expression between responding and drug-resistant patients to AC-sequential T2 chemotherapy, indicating its potential as a chemosensitivity marker [99]. Finally, during neoadjuvant therapy, miRNA-21 levels in the serum of BC patients were directly correlated with tumor size and inversely with Ki67 expression [87].
6.3. Exosomal Proteins as Predictive Indicators for Chemotherapy Response
Regarding exosomal proteins, exosomal ANXA6 levels at baseline were lower in highly sensitive TNBC patients than in resistant TNBC patients when they received gemcitabine-based first-line chemotherapy (Table 3b). This suggests that serum exosomal ANXA6 levels in TNBC patients could be predictive of the response to gemcitabine-based chemotherapy [100].
6.4. Exosomal Biomarkers as Predictive Indicators for Immunotherapy Response
Immunotherapy is an emerging therapeutic approach in TNBC and exosomes may be able to predict response to it. Specifically, exosomal PD-L1 has emerged as a significant predictive biomarker (Table 3c). Raimondi et al. have reported that patients treated with atezolizumab plus nab-paclitaxel have a significantly higher number of exosomal PD-L1 mRNA copies per mL at baseline in the case of complete or partial response compared to those with stable or progressive disease [101]. This finding aligns with another study, which demonstrated that plasma exosomes enriched with PD-L1 could serve as a favorable and predictive biomarker for immune checkpoint blockade (ICB) therapies in metastatic TNBC [102].
However, the first study adds a dynamic dimension, showing that an increase in PD-L1 mRNA copies per mL after treatment was associated with significantly shorter progression-free survival and OS. This suggests that while high initial expression may be favorable for response, an increase during or after treatment might indicate the development of resistance or more aggressive disease [101]. Consequently, exosomal PD-L1 seems to be associated with treatment outcome and response to immunotherapy.
The current standard for guiding immunotherapy decisions in TNBC is tissue PD-L1 expression, which, however, has inherent limitations: it reflects expression only at the site and time of the biopsy and may not capture the spatial heterogeneity of tumors [103,104]. In contrast, exosomal PD-L1 in plasma offers a non-invasive, dynamic alternative. It allows for repeated sampling, reflects PD-L1 from multiple tumor sites, and may provide earlier insights into treatment response or the emergence of resistance [105,106]. By complementing tissue-based analyses, exosomal PD-L1 has the potential to enhance predictive accuracy for immunotherapy, although further studies are needed to determine optimal measurement methods and validate its utility across diverse patient populations [107].
7. Conclusions and Future Perspectives
Τhis review highlights the multifaceted role of exosomes as promising biomarkers in TNBC. Due to the heterogeneity, aggressive nature and lack of targeted therapies for this particular disease, the need for novel, non-invasive biomarkers is escalating [108]. Diverse RNA molecules (such as miRNAs, lncRNAs, circRNAs) as well as proteins within exosomes have been studied for their role in the early diagnosis of TNBC, the prediction of disease progression and the response to therapies. In addition, exosomes offer a minimally invasive and dynamic method for disease monitoring [109]. However, the integration of exosomal biomarkers into clinical practice faces significant challenges and limitations. Initially, a notable heterogeneity has been observed across studies, both regarding exosome isolation and analysis methodologies, the different types of biological materials (e.g., serum, plasma, urine, etc.) and the size and populations of the samples. The lack of standardization in these techniques can impact the reproducibility of results. It is also important to note that various studies quantify exosomal RNAs using different methodologies, most commonly RT-qPCR, but also next-generation sequencing (NGS) or RNA in situ hybridization (RNA-ISH). For reliable comparison of results across studies in the future, RNA levels should ideally be measured using the same technique. Moreover, an additional methodological challenge lies in the incomplete separation of exosomes from other extracellular vesicle subtypes, such as microvesicles, as well as from protein aggregates and lipoprotein particles, which often co-isolate using commonly applied techniques [110]. This overlap can affect the molecular composition detected in exosomal preparations and may partly explain the variability reported between studies, highlighting the importance of cautious interpretation of methodological differences. Furthermore, the complexity of exosomal biomarkers and their intricate networks necessitates a deeper understanding of the biological mechanisms governing their function in TNBC. Finally, the majority of studies to date concern Asian populations (primarily Chinese and Japanese), a fact that limits the generalizability of results and necessitates the performance of multi-center studies in diverse ethnic groups.
Despite these challenges, future prospects for the clinical application of exosomal biomarkers in TNBC are particularly encouraging. The next steps in research should focus on developing standardized exosome isolation and analysis protocols, as well as highly accurate and sensitive diagnostic/prognostic kits [111]. Understanding the underlying biological mechanisms through which exosomes influence TNBC development, progression, metastasis and treatment response is crucial for developing targeted interventions [112]. Additionally, research is moving towards the utilization of panels of combinatorial biomarkers over individual ones to offer greater diagnostic and prognostic accuracy [94]. Ultimately, the utilization of exosomal biomarkers will significantly contribute to achieving personalized medicine for TNBC patients, enabling early diagnosis, more accurate prognosis and more effective treatment guidance, thereby improving clinical outcomes and patient quality of life [55].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tkach M. Théry C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go Cell 20161641226123210.1016/j.cell.2016.01.04326967288 · doi ↗ · pubmed ↗
- 2Raposo G. Stoorvogel W. Extracellular vesicles: Exosomes, microvesicles, and friends J. Cell Biol.201320037338310.1083/jcb.20121113823420871 PMC 3575529 · doi ↗ · pubmed ↗
- 3de la Torre Gomez C. Goreham R.V. Bech Serra J.J. Nann T. Kussmann M. “Exosomics”—A Review of Biophysics, Biology and Biochemistry of Exosomes with a Focus on Human Breast Milk Front. Genet.201899210.3389/fgene.2018.0009229636770 PMC 5881086 · doi ↗ · pubmed ↗
- 4Taha H.B. Plasma versus serum for extracellular vesicle (EV) isolation: A duel for reproducibility and accuracy for CNS-originating E Vs biomarker analysis J. Neurosci. Res.20231011677168610.1002/jnr.2523137501394 · doi ↗ · pubmed ↗
- 5Zhang X. Takeuchi T. Takeda A. Mochizuki H. Nagai Y. Comparison of serum and plasma as a source of blood extracellular vesicles: Increased levels of platelet-derived particles in serum extracellular vesicle fractions alter content profiles from plasma extracellular vesicle fractions P Lo S ONE 202217 e 027063410.1371/journal.pone.027063435749554 PMC 9231772 · doi ↗ · pubmed ↗
- 6Sánchez-Vizcaíno Mengual E. Cordero L. Pinto H. Human Blood Exosomes: Isolation and Characterization Methods, Variability, and the Need for Standardized Protocols—A Review Biomedicines 202513297010.3390/biomedicines 1312297041462985 PMC 12731227 · doi ↗ · pubmed ↗
- 7Mukerjee N. Bhattacharya A. Maitra S. Kaur M. Ganesan S. Mishra S. Ashraf A. Rizwan M. Kesari K.K. Tabish T. Exosome isolation and characterization for advanced diagnostic and therapeutic applications Mater. Today Bio 20253110161310.1016/j.mtbio.2025.10161340161926 PMC 11950786 · doi ↗ · pubmed ↗
- 8Corrado C. Raimondo S. Chiesi A. Ciccia F. De Leo G. Alessandro R. Exosomes as Intercellular Signaling Organelles Involved in Health and Disease: Basic Science and Clinical Applications Int. J. Mol. Sci.2013145338536610.3390/ijms 1403533823466882 PMC 3634447 · doi ↗ · pubmed ↗