Immune and Tumor Microenvironment Mechanisms of Hedyotis diffusa Willd: A Scoping Review and Network Pharmacology Analysis
Soo-Dam Kim, Eun Soo Park, Jung Hyang Park, Tae-Kyung Yeo, Dong-Hyeon Kim, So-Jung Park, Hwa-Seung Yoo

TL;DR
This study explores how the herbal medicine Hedyotis diffusa Willd affects cancer growth and the immune system, using both lab research and computational analysis.
Contribution
The study provides a comprehensive synthesis of HDW's effects on the tumor microenvironment and immune responses using a scoping review and network pharmacology.
Findings
HDW suppresses cancer cell growth and reduces harmful inflammation through multiple biological pathways.
HDW enhances anticancer immune activity and inhibits tumor angiogenesis and EMT in preclinical models.
Network analysis identified key signaling pathways like PI3K-Akt and STAT3 as central to HDW's mechanisms.
Abstract
Cancer growth is strongly influenced by its surrounding environment, which includes immune cells, blood vessels, and supporting tissues. Many traditional herbal medicines are used alongside cancer care, but their effects on this tumor environment are not always clearly understood. Hedyotis diffusa Willd (HDW) is a commonly used herb in East Asian medicine, yet its actions on immune responses and the tumor environment have not been comprehensively summarized. In this study, we reviewed existing laboratory research and combined it with computational network analysis to understand how this herb may work. We found that HDW can suppress cancer cell growth, reduce harmful inflammation, and support anticancer immune activity through multiple biological pathways. These findings provide an integrated picture of how this herbal medicine may influence cancer-related immune and microenvironmental…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
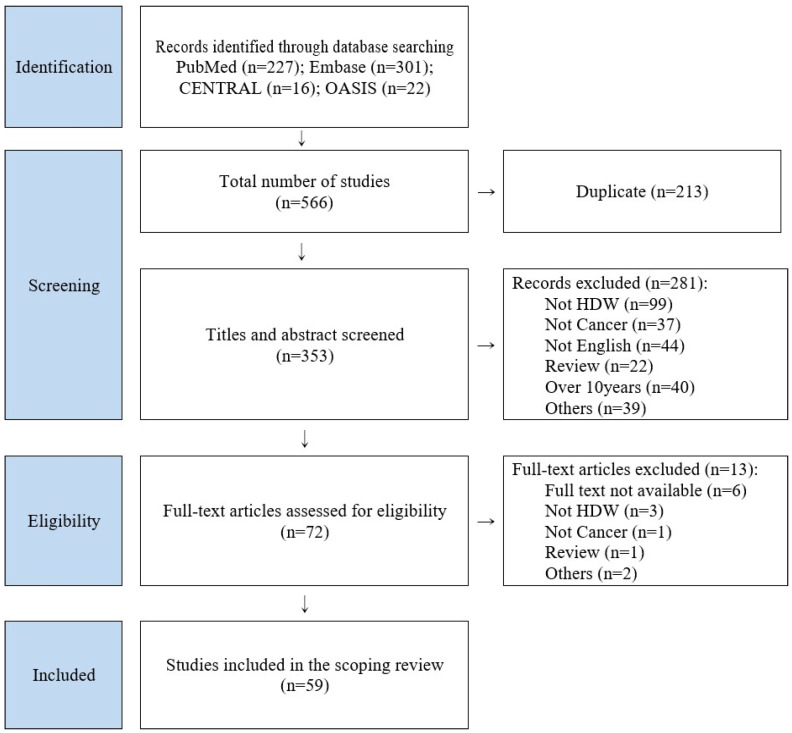 Figure 1
Figure 1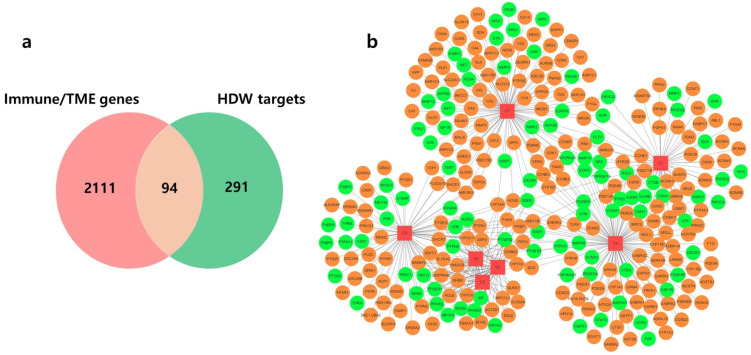 Figure 2
Figure 2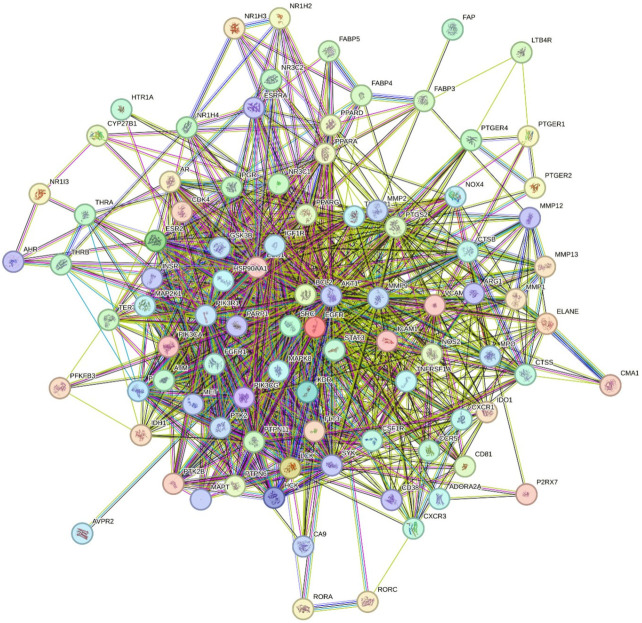 Figure 3
Figure 3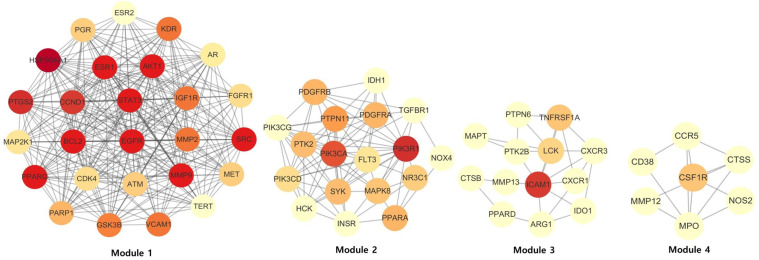 Figure 4
Figure 4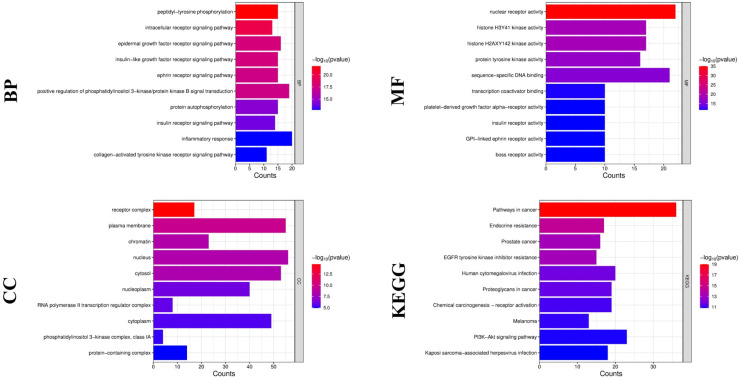 Figure 5
Figure 5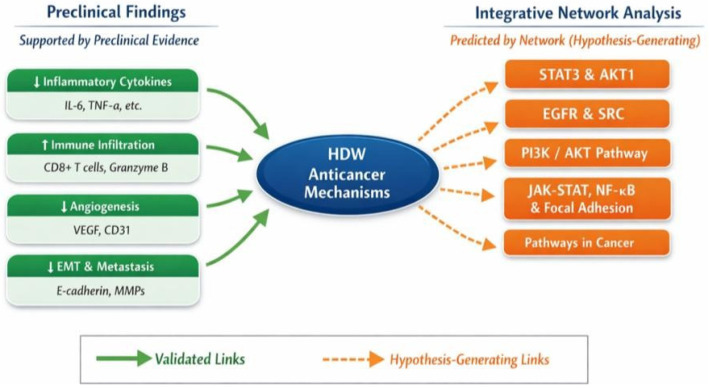 Figure 6
Figure 6- —Integrated R&D Information System (IRIS)
- —Daejeon University Research Grants
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsImmune cells in cancer · Phytochemistry and Biological Activities · Ferroptosis and cancer prognosis
1. Introduction
The concept of the tumor microenvironment (TME) has become central to modern cancer biology. Tumors develop within a complex setting that includes malignant cells, immune cells, stromal components, extracellular matrix structures, and extensive vascular support [1,2]. Disturbances within this environment, such as persistent inflammation, im-paired immune surveillance, abnormal cytokine signaling, and uncontrolled angiogenesis, promote cancer progression and allow malignant cells to evade immune control [3,4,5]. As immuno-oncology advances, therapeutic strategies increasingly concentrate on strengthening antitumor immunity. Current approaches include restoring the activity of cytotoxic T-cells, redirecting the behavior of tumor-associated macrophages, and reducing immunosuppressive features within the microenvironment [6,7,8]. These developments have encouraged researchers to explore natural compounds that can influence multiple immune-related pathways at the same time, which has increased interest in botanical medicines with broad immunomodulatory potential [9,10].
Hedyotis diffusa Willd (HDW), a herb widely used in East Asian medical systems, has traditionally been applied for its detoxifying and anti-inflammatory purposes. Modern pharmacological investigations have expanded this view by showing that HDW demonstrates antitumor activity in various in vitro and in vivo models [11,12,13]. Chemical analyses have identified triterpenoids, flavonoids, iridoids, and anthraquinones as major groups of bioactive compounds that contribute to its pharmacological effects [14,15]. Experimental studies report that HDW reduces cancer cell proliferation, induces apoptosis, limits new blood vessel formation, and modulates inflammatory responses in several malignancies, including colorectal, liver, breast, gastric, lung, and hematologic cancers [16,17,18,19,20]. Despite these diverse findings, earlier discussions have usually focused on mechanisms relevant to specific cancer types. As a result, its potential influence on immune regulation, cytokine networks, macrophage activity, and broader microenvironmental processes has not been described in a fully integrated manner.
Network pharmacology studies have attempted to explain the mechanisms of HDW, yet most previous analyses concentrated on individual diseases such as colorectal cancer, hepatocellular carcinoma, or lung cancer [21,22,23,24]. Accordingly, prior reviews and network-based analyses have largely remained disease-centered, without explicitly synthesizing immune regulation or tumor microenvironment-related mechanisms across cancer types. This fragmented approach has limited a comprehensive understanding of the immunological relevance of HDW, despite accumulating preclinical evidence suggesting its immune- and microenvironment-modulatory potential [25,26,27]. More specifically, HDW-derived flavonoid fractions have been shown to suppress pro-inflammatory cytokine production, including IL-6 and TNF-α, in macrophage models through the inhibition of NF-κB-related signaling [25]. In addition, HDW polysaccharides have been reported to activate the IL-2/IL-2R axis and enhance antitumor immune responses in murine colorectal cancer models, improving immunotherapy-related outcomes and suggesting the attenuation of immune escape-related processes within the tumor microenvironment [27]. However, these findings have not been synthesized within a cross-disease framework that links experimental observations to immune- and TME-associated molecular targets.
To address this gap, we conducted a scoping review of preclinical studies to map the immune- and TME-related effects of HDW across diverse tumor models. We then performed a targeted network pharmacology analysis focused on immune- and TME-associated molecular targets and pathways to support a systems-level interpretation of the mapped evidence. This integrated approach aims to identify shared mechanistic nodes with translational relevance and to facilitate hypothesis generation for future biomarker- or immunotherapy-oriented evaluations of HDW.
2. Methods
2.1. Study Design
This study consisted of two complementary components. First, a scoping review was conducted to map preclinical evidence on the antitumor, immune- and TME-related effects of HDW. Second, a network pharmacology analysis was performed to explore how active compounds of the herb may interact with immune- and microenvironment-associated targets at a systems level. The scoping review provided an evidence map of experimentally reported immune- and TME-related effects of HDW, which was used to contextualize the network-derived targets and pathways and to frame the network pharmacology results as hypothesis-generating.
2.2. Scoping Review
The scoping review was conducted according to the methodological framework developed by Arksey and O’Malley, and the reporting process followed the PRISMA-ScR guideline [28,29]. The review protocol was registered with the Open Science Framework (OSF) on 12 October 2025 (https://osf.io/qjmpy). In addition, the Joanna Briggs Institute methodology for scoping reviews was consulted to supplement the methodological approach [30]. The objective of this review was to organize and classify existing preclinical findings related to the biological actions of HDW, including antitumor activity, immune-associated responses, and TME-related mechanisms. The review identified the types of cancer models used, the experimental designs applied, the outcomes measured, and the main findings reported across the studies.
2.2.1. Search Strategy
A comprehensive literature search was carried out in PubMed, Embase, CENTRAL and OASIS. OASIS was included to capture Korea-indexed studies on HDW that may not be fully indexed in international databases, thereby improving coverage of East Asian evidence. The search included studies published from January 2016 to December 2025. Search terms consisted of controlled vocabulary and free-text words related to the herb and cancer research. The primary terms included “Hedyotis diffusa,” “Bai Hua She She Cao,” and cancer-related terms such as “cancer,” “tumor,” “carcinoma,” and “neoplasm.” Immune- and TME-specific keywords were not added to the core search to avoid excessive non-cancer retrieval and to preserve sensitivity, and immune/TME relevance was applied during eligibility assessment and data extraction. These terms were combined using Boolean operators appropriate to each database (Supplementary Materials Table S1). A representative full search string for PubMed was as follows: ((cancer*) OR (carcinoma*) OR (neoplasm*) OR (tumor*)) AND ((Hedyotis diffusa*) OR (Oldenlandia diffusa*) OR (Baihuasheshecao*)). Searches were performed without language restrictions at the retrieval stage. However, the final eligibility assessment was limited to studies with accessible full texts published in English.
2.2.2. Eligibility Criteria
Studies were considered eligible if they met the following conditions: (1) preclinical experiments using in vitro or in vivo models of cancer; (2) the intervention consists of HDW extracts, isolated fractions, or identified active compounds from the herb; (3) outcomes include antitumor effects or biological responses relevant to immune or microenvironmental processes. Studies involving multi-herb formulas that did not allow the evaluation of the specific contribution of HDW were excluded. Reviews, commentaries, and conference abstracts without extractable data, as well as studies without accessible full text, were excluded. Clinical studies were excluded because this scoping review focused on preclinical models to synthesize mechanistic evidence. To ensure consistent interpretation and data extraction, the final inclusion was restricted to English-language full-text articles.
2.2.3. Study Selection Process
All retrieved records were imported into a reference management program to remove duplicates. Two reviewers independently screened titles and abstracts to identify potentially eligible studies. Full texts of selected articles were then assessed for final inclusion. Disagreements were resolved through discussion or by consulting a third reviewer. The selection process was presented using a PRISMA-ScR flow diagram.
2.2.4. Data Extraction and Synthesis
A standardized extraction form was used to collect information from the included studies. Extracted data included publication year, country, cancer type, experimental model, type of HDW preparation, dose and exposure conditions, measured outcomes, and main findings. Two reviewers independently extracted data, and discrepancies were resolved through discussion and consensus. If needed, a third reviewer adjudicated unresolved discrepancies. Data were organized to identify patterns across studies, such as frequently reported mechanisms, common signaling pathways, and recurring biological responses. Because this review was exploratory, findings were synthesized descriptively. Results were summarized according to cancer types, experimental models, types of HDW preparations, and major categories of biological effects.
2.3. Network Pharmacology Analysis
The network pharmacology analysis was conducted as a hypothesis-generating approach to explore how active compounds of HDW interacted with immune- and TME-related targets at a systems level, following principles of network-based drug discovery and traditional medicine systems pharmacology [31,32]. The analysis proceeded through four main steps: identification of candidate active compounds, prediction of potential human protein targets, collection of immune- and TME-related gene sets, and construction and interpretation of protein-protein interaction (PPI) networks and enriched pathways.
2.3.1. Identification of Candidate Active Compounds
Candidate active compounds of HDW were identified using the Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform (TCMSP), accessed on 5 September 2025 [33]. All compounds registered for HDW were retrieved, and oral bio- availability (OB) and drug-likeness (DL) values were extracted. In line with previous systems pharmacology studies, compounds were retained as candidates if they satisfied the thresholds of OB ≥ 30% and DL ≥ 0.18.
2.3.2. Target Prediction for Active Compounds
Potential human protein targets for each selected compound were predicted through SwissTargetPrediction [34]. The simplified molecular input line entry specification (SMILES) for each compound was obtained from TCMSP or from the PubChem database and was submitted to SwissTargetPrediction with the species parameter set to Homo sapiens. For each compound, predicted targets with probability ≥ 0.09 were extracted, and targets with probability < 0.09 were removed to reduce noise. All predicted targets were combined into a single list, and duplicate entries were removed based on gene symbol or UniProt identifier.
2.3.3. Collection of Immune and TME-Related Gene Sets
Immune-related genes were collected from the Immunology Database and Analysis Portal (ImmPort), which provides curated gene lists associated with immune functions [35]. TME-related genes were identified from the GeneCards database using the search term “tumor microenvironment” [36]. Genes with a relevance score ≥ 2 were retained. The immune-related genes from ImmPort and the TME-related genes from GeneCards were merged into a single gene set after the removal of duplicate symbols. All databases were accessed on 5 September 2025.
2.3.4. Identification of Intersecting Targets and PPI Network Construction
Intersecting targets between the predicted HDW-related targets and the immune and TME gene set were identified using spreadsheet software. The overlap was computed based on official gene symbols, and the resulting intersecting genes were considered as candidate mediators of immune- and microenvironment-related actions of HDW. These intersecting targets were then submitted to the STRING database (Search Tool for the Retrieval of Interacting Genes or Proteins, version 12.0) to construct a PPI network, with the organism set to Homo sapiens and a minimum required interaction score of 0.4 [37]. The interaction data were exported from STRING for subsequent network visualization and analysis.
2.3.5. Network Visualization and Identification of Key Targets
The PPI network was imported into Cytoscape (version 3.10.4), an open source software platform for visualizing complex networks [38]. Topological properties such as degree and betweenness centrality were calculated using the network analysis functions within Cytoscape to identify highly connected or central nodes as potential key targets. Network clustering was performed with the MCODE plug-in to detect densely connected modules that might represent functional sub-networks [39]. Modules with relatively high scores and clear biological coherence were selected for further interpretation.
2.3.6. Functional Enrichment Analysis
Functional enrichment analysis was performed to investigate the biological path-ways and processes associated with the intersecting targets and with key network modules. The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway and Gene Ontology biological process (GO-BP) enrichment were conducted using the enrichment tools provided within the STRING platform [40,41]. Multiple testing was corrected using the Benjamini-Hochberg false discovery rate (FDR) method, and terms with FDR-adjusted p-values < 0.05 were considered significant. Pathways and biological processes with statistically significant enrichment were examined, with a focus on immune signaling, cytokine and chemokine networks, leukocyte activation, macrophage polarization, and angiogenesis. The results of the network pharmacology analysis were integrated with findings from the scoping review to propose a coherent mechanistic framework for the actions of HDW in the context of antitumor immunity and the TME.
3. Results
3.1. Study Selection
A total of 566 records were identified through database searching. After removing 213 duplicates, 353 studies remained for title and abstract screening. During this screening stage, 288 records were excluded for the following reasons: not related to HDW (n = 99), not cancer-related (n = 37), not published in English (n = 44), review articles (n = 22), published more than 10 years ago (n = 40), or excluded for other reasons (conference abstracts or protocol-only records) (n = 39). Following this process, 72 full-text articles were assessed for eligibility. Of these, 13 articles were excluded at the full-text stage for the following reasons: full text not available (n = 6), not HDW (n = 3), not cancer (n = 1), review (n = 1), and others (n = 2). Finally, 59 studies met the eligibility criteria and were included in the scoping review. A summary of the study selection process is presented in Figure 1.
3.2. Characteristics of Included Studies
Most of the eligible studies were conducted in China (n = 49), with a smaller number originating from other countries (n = 10). Across all included studies (n = 59), a majority of the included studies used in vitro experimental systems (n = 32), whereas others combined both in vitro and in vivo approaches (n = 24). Only a small number relied exclusively on animal models (n = 3). Across the studies, a wide range of cancer types was represented. The most frequently investigated models were colorectal cancer (16 studies), lung cancer, including NSCLC and adenocarcinoma (10 studies), and hepatocellular carcinoma (8 studies). Sample sizes varied substantially, particularly among animal studies, where treatment groups ranged from 4 to 15 animals. A summary of representative study characteristics is provided in Table 1.
3.3. Antitumor Activity and Growth Inhibition
3.3.1. Inhibition of Cancer Cell Proliferation and Cell Cycle Arrest
Across diverse tumor models, HDW and its derivatives consistently suppressed cancer cell growth. Crude extracts, polysaccharide fractions, total flavonoids, and single constituents such as quercetin and ursolic acid reduced cell viability and/or colony formation in breast, colorectal, hepatocellular, lung, and bladder cancer, while showing limited toxicity in non-malignant lines [42,44,45,47,51,65,66,67,76,88]. Quercetin decreased MCF-7 viability together with the downregulation of BIRC5 and CDK1 [45], and HDW-derived exosome-like particles (HDW-EVLPs) inhibited Huh-7 growth with minimal impact on WRL68 hepatocytes [13]. HDW selectively reduced A549 cell viability but spared WI-38 fibroblasts, indicating a degree of tumor selectivity [65]. Many of these studies also reported cell cycle arrest at G0/G1, G1/S, or G2/M, accompanied by changes in cyclins, CDKs, and p21, suggesting a direct interruption of cell cycle progression and DNA replication [12,22,26,44,60,62,66,67,72,73,74,83,85,86].
3.3.2. Induction of Apoptosis and Other Regulated Cell Death Pathways
Growth inhibition was closely linked to the activation of programmed cell death. Numerous HDW-based preparations triggered classical apoptosis characterized by chromatin condensation, Annexin V positivity, mitochondrial depolarization, caspase-3/8/9 activation, and PARP cleavage, together with a shift in Bcl-2 family proteins toward a proapoptotic profile (Bax/Bad upregulation with Bcl-2/Bcl-xL downregulation) [47,48,55,56,60,62,65,66,67,72,89,91]. ODE and ethanol extracts from HDW induced caspase-dependent mitochondrial apoptosis in colorectal cancer, with increased Bax, Apaf-1, and cytosolic cytochrome c [66,67]. Additional forms of regulated cell death were also identified. In bladder cancer, HDW decreased GPX4 and SLC7A11 and increased TFRC and HMOX1, together with tumor shrinkage, supporting ferroptosis-mediated antitumor effects [42]. Lung cancer models exposed to HDW-derived injections showed lipid ROS accumulation and Fe^2+^ overload consistent with ferroptotic damage [19]. Flavonoid-rich fractions and other preparations enhanced LC3B-II expression and autophagic flux while simultaneously reducing viability and tumor burden, indicating an autophagy-associated cytotoxic mechanism in selected systems [44,82]. Overall, HDW appears capable of engaging multiple, context-dependent regulated cell death pathways.
3.3.3. Suppression of Tumor Growth and Systemic Safety in Animal Models
In vivo findings generally mirrored the in vitro data. Across xenograft and syngeneic models of colorectal, hepatocellular, lung, breast, and bladder cancer, HDW-based interventions reduced tumor volume, tumor weight, or overall tumor burden relative to controls [12,19,42,44,48,51,65,66,67,72,73,82,84,91]. Polysaccharides, ethanol extracts, and aqueous ODE preparations produced marked tumor growth inhibition in mice, in some cases comparable to standard anticancer agents, and in leukemia models HDW increased survival while limiting splenomegaly [56,65,67,72]. Histological analyses showed reduced proliferation markers such as Ki-67 or PCNA and increased TUNEL-positive apoptotic cells in tumor tissue, confirming that macroscopic tumor control was accompanied by enhanced cell death [44,48,51,66,67,72,73,91]. Several studies simultaneously documented preserved body weight, stable liver and kidney function, and unremarkable organ histology, suggesting a favorable systemic safety profile at therapeutically active doses [12,19,44,51,65,67].
3.3.4. Inhibition of Metastasis, Invasion, and Angiogenesis
HDW and its constituents also constrained metastatic behavior. Extracts and isolated compounds reduced migration and invasion in colorectal, hepatocellular, lung, and ovarian cancer models, frequently in parallel with the downregulation of epithelial-mesenchymal transition (EMT) and invasion related proteins such as N-cadherin, vimentin, MMP-2, MMP-9, and uPA, and the restoration of E-cadherin expression [26,50,51,59,66,72,73,74,83,89]. OD and HDW decreased lung metastatic nodules in animal models, while a suppression of wound closure and invasion was observed in zebrafish metastasis assays [59,86]. In a circulating tumor cell model, HDW reduced adhesion to the extracellular matrix, endothelial cells, and platelets, leading to a lower lung metastatic burden without affecting body weight [90]. Vascular modulation was another recurring feature: decreased tumor burden in some studies coincided with reductions in vascular markers such as CD31, α-SMA, and HIF-1α, indicating that anti-angiogenic effects and the remodeling of the tumor vasculature contribute to overall growth inhibition [19,84].
3.3.5. Enhanced Sensitivity to Anticancer Therapies
A number of studies indicated that HDW-derived agents can act as chemosensitizers. Combinations of HDW or its constituents with cisplatin, 5-fluorouracil, arsenic trioxide, or HSP90 inhibitors produced a deeper suppression of proliferation, stronger apoptosis, and greater inhibition of colony formation than single-agent treatment [56,58,71,74,80,92]. HDW increased the responsiveness of colorectal and leukemia cells to cytotoxic drugs, and ursolic acid enhanced the antitumor efficacy of 5-fluorouracil with substantial reductions in tumor volume and weight in vivo [56,71]. Mechanistic work linked these synergistic effects to the modulation of signaling pathways associated with drug resistance and survival, including PI3K/AKT, ERK, STAT3, telomerase activity, β-catenin/CTNNB1, and DNA damage-response networks [22,55,62,67,71,80,82]. These findings suggest that HDW not only exerts direct antitumor actions but may also improve the therapeutic performance of existing anticancer regimens (Supplementary Materials Table S2).
3.4. Immune and TME Effects
3.4.1. Immune Activation and Cytotoxic Enhancement
Evidence from preclinical studies indicates that HDW and its polysaccharide fractions augment antitumor immunity by strengthening effector lymphocyte function. In a syngeneic colorectal cancer model, polysaccharides from HDW markedly increased intratumoral CD4^+^ and CD8^+^ T-cell infiltration, accompanied by elevated levels of granzyme B, TNF-α, and IFN-γ, while reducing tumor burden [27]. The same study showed that these immune responses were further intensified when HDW polysaccharides were combined with PD-1 and CTLA-4 blockade, suggesting functional complementarity with immune checkpoint inhibition [27]. Immune cell-mediated cytotoxicity was also enhanced in adoptive cell therapy models. Treatment with HDW polysaccharides increased CD3^+^CD56^+^ effector populations and augmented TNF-α and IFN-γ production, resulting in superior tumor suppression in vivo when combined with cytokine-induced killer cells [70]. Alongside immune activation, improvements in systemic inflammatory status were observed in multiple animal models, with reductions in serum IL-6 and TNF-α reported in hepatocellular and colorectal cancer studies [12,44,46,48,84]. Collectively, these data indicate that HDW enhances antitumor immunity predominantly through the amplification of cytotoxic lymphocyte activity rather than the broad stimulation of suppressive immune subsets.
3.4.2. Regulation of Inflammatory Signaling and Immunosuppressive Pathways
HDW consistently attenuated pro-tumorigenic inflammatory signaling across tumor models. Several studies documented the downregulation of key cytokines, including IL-6, IL-1β, IL-8, IL-17A, and TNF-α, indicating the inhibition of sustained inflammatory circuits that promote tumor progression [12,17,44,46,48]. These immunological changes were accompanied by the suppression of major regulatory pathways, such as NF-κB and JAK2/STAT3, which are central to tumor-associated immune evasion and cytokine production [48,55,84]. At the molecular level, ursolic acid blocked STAT3 phosphorylation and nuclear translocation and increased miR-4500 expression, thereby disrupting STAT3-driven oncogenic signaling [55]. Functional validation confirmed that miR-4500 inhibition partially reversed the antitumor effects of ursolic acid, directly linking STAT3 regulation to immune-modulatory and cytotoxic outcomes [55]. Quercetin similarly suppressed IL-6 and TNF expression at both transcript and protein levels while activating AGE-RAGE signaling, suggesting an additional immunoregulatory axis involved in HDW-mediated effects [17]. These findings support a dual role for HDW in dampening pathological inflammation while restoring immune competence within the TME.
3.4.3. TME Remodeling and Metastatic Control
HDW and its constituents exerted pronounced effects on the structural and biochemical components of the TME. Multiple studies reported the reversal of epithelial-mesenchymal transition, reflected by an increased E-cadherin and decreased N-cadherin or vimentin expression, which translated into reduced migratory and invasive phenotypes in colorectal, hepatic, lung, and breast cancer models [43,57,59,64,69,86]. Coumarin polymers further potentiated these anti-EMT effects when co-administered with HDW, highlighting a cooperative mechanism in metastatic suppression [69]. Anti-angiogenic and anti-lymphangiogenic activity was also consistently observed. HDW downregulated VEGFA, VEGF-C, VEGFR3, CD31, and α-SMA expression and impaired endothelial tube formation, thereby inhibiting vascular and lymphatic support of tumor growth [63,84,87]. These vascular effects were accompanied by the suppression of PI3K/AKT, ERK, and STAT3 signaling cascades, indicating that angiogenic regulation is mechanistically linked to growth-factor pathway inhibition [63,84,87]. Metabolic remodeling constituted another dimension of microenvironmental control. HDW partially normalized tumor-induced metabolic disturbances in rats by correcting amino acid, glycolytic, and lipid profiles, suggesting the restoration of systemic metabolic balance in tumor-bearing hosts [79]. Ursolic acid further disrupted tumor metabolism by suppressing glycolysis through the SP1-dependent regulation of CAV1, with functional knockdown experiments confirming the CAV1/SP1 axis as critical for its metabolic and antiproliferative effects [81]. In metastatic models, HDW reduced circulating tumor cell adhesion to platelets and endothelial cells through the inhibition of Src/FAK signaling, leading to decreased pulmonary metastasis without causing body weight loss [90]. Ferroptosis-related microenvironmental changes were also reported, with increased lipid ROS, Fe^2+^ accumulation, and an upregulation of HMOX1 and transferrin receptor expression in lung cancer models [19]. Taken together, these results indicate that HDW reshapes the TME through the coordinated regulation of immune infiltration, inflammatory signaling, vascular support, metabolic pathways, and extracellular matrix dynamics, thereby creating conditions that are less permissive for tumor growth and dissemination (Supplementary Materials Table S3).
3.5. Network Pharmacology Analysis Results
3.5.1. Related Targets of HDW in the Immune and TME Gene Sets
A total of 2110 immune- and TME-related targets were collected from the ImmPort (n = 1793) and GeneCards (n = 477) databases. From the TCMSP database, 37 compounds associated with HDW were initially retrieved, among which 7 compounds met the predefined pharmacokinetic criteria (OB ≥ 30% and DL ≥ 0.18). These active compounds corresponded to 291 predicted protein targets, of which 94 overlapped with the immune- and TME-related gene set (Figure 2a). The relationships between active compounds and their targets are illustrated as a compound-target network in Figure 2b. In the network, 417 red squares denote HDW-derived compounds and circles indicate predicted targets, with 418 green nodes representing immune- and TME-related genes and orange nodes indicating 419 non-immune/TME-related targets. Most targets in the network were associated with immune functions and microenvironmental regulation, supporting the involvement of 421 HDW in antitumor immunity and TME regulation.
3.5.2. PPI Network Construction and Topological Analysis
The intersecting immune- and TME-related targets of HDW were used to construct a PPI network based on STRING analysis (Figure 3). After the removal of isolated nodes, the final network consisted of 94 nodes and 1136 edges, with an average node degree of 24.2 and a clustering coefficient of 0.641, reflecting a highly interconnected network topology. Topological analysis identified several hub genes with a high degree centrality. AKT1, STAT3, EGFR, SRC, and BCL2 ranked as the most highly connected nodes, indicating their central positions within the network. Additional high-degree targets included MMP9, ESR1, PPARG, HSP90AA1, and PIK3R1, reflecting an involvement in pathways related to invasion, hormone signaling, inflammatory regulation, and cellular stress responses. Overall, the PPI structure demonstrated that HDW-related immune and TME targets form a dense interaction network characterized by multiple interconnected signaling hubs.
3.5.3. Module Detection
Module analysis using the MCODE algorithm identified four highly interconnected clusters within the PPI network (Figure 4). Module 1 contained 26 nodes and was dominated by key regulators involved in cell survival, hormone signaling, and inflammatory control. Module 2 comprised 18 targets mainly associated with growth-factor signaling and PI3K-related pathways. Module 3 included 13 nodes enriched in immune signaling and cytokine-related processes. Module 4 consisted of seven targets primarily linked to innate immune responses and inflammatory remodeling. Collectively, these findings indicate that HDW-related targets are organized into functionally distinct sub-networks rather than forming a single homogeneous interaction network.
3.5.4. Functional Enrichment Analysis of Intersecting Targets
GO function enrichment and KEGG pathway enrichment analyses were performed for the 94 intersecting targets using the DAVID database (https://davidbioinformatics.nih.gov/ (accessed on 25 October 2025)). The GO analysis comprised biological processes (BP), cellular components (CC), and molecular functions (MF), and the results are summarized in Figure 5.
In the BP category, the intersecting targets were strongly enriched in peptidyl-tyrosine phosphorylation, intracellular receptor signaling, and multiple receptor tyrosine kinase-associated pathways, including epidermal growth factor receptor, insulin-like growth factor, ephrin, and insulin signaling. The positive regulation of PI3K-AKT signaling and inflammatory response also ranked among the most significant biological processes, indicating a convergence of growth signaling and immune-related mechanisms. In the CC category, the major enriched terms included plasma membrane, nucleus, cytosol, receptor complex, chromatin, and nucleoplasm, suggesting that the intersecting targets are spatially distributed across membrane signaling compartments and nuclear regulatory structures. In the MF category, nuclear receptor activity, protein tyrosine kinase activity, sequence-specific DNA binding, and transcription coactivator binding were predominant, highlighting roles in signal transduction and transcriptional regulation.
KEGG pathway analysis demonstrated a significant enrichment in pathways in cancer, endocrine resistance, prostate cancer, EGFR tyrosine kinase inhibitor resistance, and the PI3K-Akt signaling pathway. Additional enriched pathways involved proteoglycans in cancer, focal adhesion, AGE-RAGE signaling, central carbon metabolism in cancer, and prolactin signaling, reflecting the integrated regulation of oncogenic signaling, metabolism, and microenvironmental interaction. Together, these findings indicate that HDW-related targets are concentrated within receptor tyrosine kinase-centered signaling axes and PI3K-AKT-dominated networks, linking immune regulation, metabolic control, and tumor progression through a coordinated multi-pathway framework.
4. Discussion
The present study brings together the broad preclinical literature and a systems-level network analysis to clarify how HDW modulates antitumor immunity and the TME. Contemporary cancer biology views tumors as dynamic ecosystems in which malignant cells, stromal elements, immune populations, and vascular structures co-evolve through chronic inflammation and immune escape [93]. Within this framework, the TME is not only a barrier to therapy but also a source of therapeutic vulnerabilities. Natural products remain a major reservoir of multi-target scaffolds for oncology drug discovery, and several have already entered clinical use [9,10]. Previous reviews have summarized the chemistry and broad pharmacology of HDW but have not focused systematically on immune and TME mechanisms [14,15]. By structuring heterogeneous preclinical findings and embedding them within a network of immune- and TME-related targets, our work reframes HDW as a candidate multi-component modulator of tumor ecology rather than a single-pathway cytotoxic agent. Network-based analyses are used to contextualize, rather than replace, the experimentally derived findings (Figure 6).
Across tumor types, the scoping review shows that growth inhibition and regulated cell death are among the most consistent effects of HDW. Extracts, polysaccharides, flavonoid-rich fractions, and defined constituents such as quercetin and ursolic acid reduce proliferation, induce cell cycle arrest, and trigger apoptosis or ferroptosis in colorectal, cervical, hepatocellular, lung, and prostate cancer models [11,12,16,19,42,44,48,51]. These phenotypes are accompanied by the modulation of cyclins, CDKs, Bcl-2 family proteins, and ferroptosis-related markers, suggesting that HDW interferes with survival and stress-response programs at multiple points. The network analysis helps to rationalize this diversity: intersecting immune/TME targets cluster around hub genes such as AKT1, STAT3, BCL2, and EGFR within a dense PPI network. The enrichment of PI3K-Akt-centered “Pathways in cancer” indicates that relatively few highly connected nodes may integrate upstream compound signals into broad effects on proliferation and cell death.
In vivo experiments support the translation of these molecular changes into macroscopic tumor control with an acceptable systemic safety. HDW-based preparations consistently reduced tumor volume or weight in xenograft and syngeneic models of colorectal, liver, lung, and bladder cancer, with corresponding decreases in proliferation markers and increases in apoptotic indices in tumor tissue [19,42,51,84]. Multiple studies reported preserved body weight and a largely unremarkable histology of major organs, suggesting a therapeutic window in preclinical systems [42,51,67]. At the same time, HDW enhanced the efficacy of conventional agents such as cisplatin and arsenic trioxide, producing a deeper suppression of proliferation and apoptosis induction than monotherapy [56,74]. In the network, these observations align with modules enriched for growth-factor and PI3K-related signaling, in which AKT1, EGFR, and related kinases act as central nodes that regulate both intrinsic survival pathways and responses to cytotoxic or targeted therapy [31,37,40].
The immune findings reveal that HDW not only affects tumor cells directly but also alters antitumor immunity. Polysaccharide-rich preparations increased intratumoral CD4^+^ and CD8^+^ T-cell infiltration, enhanced granzyme B, TNF-α, and IFN-γ production, and improved the performance of immune checkpoint blockade and adoptive cell therapy in colorectal cancer models [17,27,70]. Parallel reductions in IL-6, TNF-α, and other pro-inflammatory cytokines, together with the inhibition of NF-κB and JAK2/STAT3 signaling, indicate the suppression of chronic tumor-promoting inflammation [17,25,48]. Our immune- and TME-focused gene sets from ImmPort and GeneCards captured this duality by highlighting targets involved in cytokine signaling, innate immunity, and inflammatory remodeling. STAT3 and related transcriptional regulators emerged as hubs that connect inflammatory pathways with survival signaling, which may provide a plausible mechanistic context for improved cytotoxic responses and dampened pathological inflammation in the TME [17,31].
The remodeling of the structural and metabolic components of the TME is another recurring theme that becomes clearer when mapped onto network modules. Experimentally, HDW and its constituents reversed epithelial-mesenchymal transition, reduced migration and invasion, and downregulated EMT-related proteins while restoring E-cadherin in colorectal and lung cancer models [43,69,86]. Anti-angiogenic and anti-lymphangiogenic effects were demonstrated through reduced VEGF-related markers and impaired endothelial tube formation in liver and colorectal cancer [19,63,84]. Metabolomics work further showed a partial normalization of tumor-induced disturbances in amino acid, glycolytic, and lipid pathways in tumor-bearing rats [79]. Within the network, the enrichment for focal adhesion, proteoglycans in cancer, and central carbon metabolism suggests that HDW influences how tumor cells adhere, signal through the matrix, and utilize nutrients. Together, these data support a model in which HDW reshapes the microenvironmental “soil” that supports tumor “seeds,” making it less permissive for invasion, vascularization, and systemic metabolic disruption.
The compound-target network underscores that these effects arise from cooperation among multiple HDW constituents rather than the action of a single molecule. A subset of compounds that passed pharmacokinetic filters yielded a compact yet densely connected network of immune- and TME-related targets, in which flavonoids and triterpenoids such as quercetin and ursolic acid occupy central positions. Independent studies have shown these molecules to modulate STAT3, PI3K-Akt, and metabolic pathways in colorectal and other cancers, supporting their roles as key drivers within the HDW pharmacological profile [45,55,88]. Network pharmacology analyses in colorectal and prostate cancer similarly highlight PI3K-Akt, MAPK, and matrix-related pathways as recurrent nodes of HDW action [23,24]. By integrating our scoping review with these in silico results, HDW can be viewed as a modular assembly of compounds acting on overlapping sub-networks controlling cell survival, immunity, and tissue architecture.
From a translational viewpoint, this integrated picture positions HDW as a candidate adjunct for strategies that target the TME and antitumor immunity. The ability of HDW preparations to enhance checkpoint blockade, augment adoptive cell therapies, and sensitize tumors to chemotherapy resonates with current efforts to exploit microenvironmental vulnerabilities and overcome immune evasion [4,5,17]. At the same time, the evidence base is almost entirely preclinical and is characterized by a substantial heterogeneity in models, formulations, and dosing. In addition, common internal validity limitations, such as the frequent non-reporting of sample size, incomplete reporting of randomization or blinding, and variable characterization of HDW preparations across different plant species, plant parts, and developmental stages, may introduce bias and limit comparability across studies. These limitations reflect an inherent preclinical bias, and the lack of a consistent pharmacokinetic characterization of HDW preparations and their key constituents, including bioavailability, metabolism, and tissue distribution, further constrains translational inference.
As a scoping review, our work maps this landscape but does not provide pooled effect estimates or formal risk-of-bias assessments. On the network side, target prediction and enrichment depend on available databases and may miss relevant interactions. In addition, database-driven target prediction introduces uncertainty and does not account for exposure-related constraints such as bioavailability, metabolism, or tissue distribution. Therefore, the in silico results should be interpreted as hypothesis-generating rather than causal. Because the network component was intentionally centered on shared immune- and TME-related hubs across heterogeneous preclinical tumor models, we did not perform a compound-disease/drug-disease pathway analysis for specific cancer indications, and disease-specific interpretations should therefore be viewed accordingly. Future work should therefore combine standardized in vivo experiments with a focused validation of priority hubs such as AKT1, STAT3, and caveolin-1, extend the network framework to cancer-type-stratified compound-target-disease/pathway analyses, evaluate rational combinations with immunotherapy, and eventually link HDW exposure to defined immune and TME endpoints with biomarker-guided stratification in early-phase clinical settings.
5. Conclusions
This study integrates a scoping review of preclinical evidence with network pharmacology to clarify how HDW influences antitumor immunity and the TME. Across diverse cancer models, HDW consistently suppresses tumor growth while modulating immune activity, inflammatory signaling, angiogenesis, epithelial-mesenchymal transition, and metabolic pathways. A systems-level analysis indicates that these heterogeneous effects converge on a limited set of highly connected signaling hubs, particularly the PI3K-Akt-STAT3 axis, linking direct cytotoxic stress with immune activation and microenvironmental remodeling. Rather than acting as a single-pathway agent, HDW emerges as a multi-component botanical modulator that reshapes tumor-immune interactions and the surrounding microenvironment. Although the current evidence is largely preclinical and exploratory, this integrated framework provides a rationale for the further investigation of HDW as a potential adjunct strategy in preclinical and translational research targeting immune and microenvironmental vulnerabilities in cancer.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baghban R. Roshangar L. Jahanban-Esfahlan R. Seidi K. Ebrahimi-Kalan A. Jaymand M. Kolahian S. Javaheri T. Zare P. Tumor microenvironment complexity and therapeutic implications at a glance Cell Commun. Signal.2020185910.1186/s 12964-020-0530-432264958 PMC 7140346 · doi ↗ · pubmed ↗
- 2Zhong H. Zhou S. Yin S. Qiu Y. Liu B. Yu H. Tumor microenvironment as niche constructed by cancer stem cells: Breaking the ecosystem to combat cancer J. Adv. Res.20257127929610.1016/j.jare.2024.06.01438866179 PMC 12126709 · doi ↗ · pubmed ↗
- 3Nishida A. Andoh A. The Role of Inflammation in Cancer: Mechanisms of Tumor Initiation, Progression, and Metastasis Cells 20251448810.3390/cells 1407048840214442 PMC 11987742 · doi ↗ · pubmed ↗
- 4Tufail M. Jiang C.-H. Li N. Immune evasion in cancer: Mechanisms and cutting-edge therapeutic approaches Signal Transduct. Target. Ther.20251022710.1038/s 41392-025-02280-140739089 PMC 12311175 · doi ↗ · pubmed ↗
- 5Yu H. Li J. Peng S. Liu Q. Chen D. He Z. Xiang J. Wang B. Tumor microenvironment: Nurturing cancer cells for immunoevasion and druggable vulnerabilities for cancer immunotherapy Cancer Lett.202561121738510.1016/j.canlet.2024.21738539645024 · doi ↗ · pubmed ↗
- 6Yang Y. Yu Q. Zhang H. Liu Y. Wang H. Yang N. Shi Y. Zhang W. Wu Z. Huang S. Restoring tumor antigenicity activates the “bystander” T cell immune cycle J. Control. Release 202538025626810.1016/j.jconrel.2025.01.09439894265 · doi ↗ · pubmed ↗
- 7Wei G. Li B. Huang M. Lv M. Liang Z. Zhu C. Ge L. Chen J. Polarization of Tumor Cells and Tumor-Associated Macrophages: Molecular Mechanisms and Therapeutic Targets Med Comm 20256 e 7037210.1002/mco 2.7037240904700 PMC 12402616 · doi ↗ · pubmed ↗
- 8Shafqat A. Omer M.H. Ahmed E.N. Mushtaq A. Ijaz E. Ahmed Z. Alkattan K. Yaqinuddin A. Reprogramming the immunosuppressive tumor microenvironment: Exploiting angiogenesis and thrombosis to enhance immunotherapy Front. Immunol.202314120094110.3389/fimmu.2023.120094137520562 PMC 10374407 · doi ↗ · pubmed ↗