Transcranial Alternating Current Stimulation for Pain: Mixed Evidence and the Path to Precision Neuromodulation
Yaser Fathi, Amin Dehghani, David M. Gantz, Giulia Liberati, Tor D. Wager

TL;DR
This review explores how transcranial alternating current stimulation (tACS) affects pain perception, finding mixed results and suggesting personalized approaches could improve its effectiveness.
Contribution
The paper proposes integrating new tACS methods like phase-specific stimulation and individualized modeling to achieve more precise and personalized pain modulation.
Findings
Out of 14 studies, 8 showed analgesic benefits or neural changes with tACS.
Somatosensory alpha oscillations were most commonly targeted in tACS pain studies.
Variability in stimulation protocols and individual differences may explain inconsistent results.
Abstract
Neural oscillations are fundamental to the integration of sensory, affective, and cognitive processes that contribute to pain perception. Transcranial alternating current stimulation (tACS) provides a valuable tool for investigating and modulating these oscillatory dynamics. In this review, we examine the effects of tACS on pain perception and pain-related oscillations in both healthy participants and individuals with chronic pain, highlighting methodological variability and mechanistic uncertainties that may contribute to mixed findings. We identified 14 studies, including 9 studies of experimental pain in healthy individuals and 5 of clinical pain disorders, comparing tACS to sham. Somatosensory alpha was the most frequently targeted oscillatory feature. Results varied considerably. Several studies reported reductions in pain, increases in alpha power, or changes in sensorimotor and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
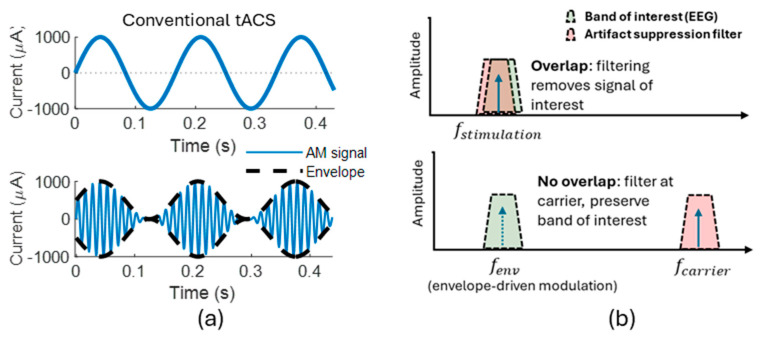 Figure 1
Figure 1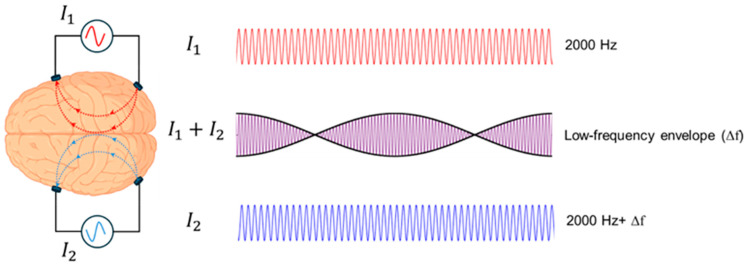 Figure 2
Figure 2- —FRS-FNRS MIS grant
- —Fondation Médicale Reine Elisabeth (F.M.R.E.)
- —National Institute of Mental Health
- —Neukom Institute for Computational Science at Dartmouth College
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTranscranial Magnetic Stimulation Studies · Pain Mechanisms and Treatments · Pain Management and Treatment
1. Introduction
Neural oscillations reflect neurophysiological mechanisms that support the integration of sensory, affective and cognitive processes involved in pain perception [1]. Human rhythms have historically been characterized by their frequency, with major frequency bands including (from low to high) delta (0.5–4 Hz), theta (4–7 Hz), alpha (7–12 Hz), beta (12–30 Hz), and gamma (30–55 Hz for low gamma and 55–200 for high gamma). They are measured using time-frequency decomposition of scalp electroencephalography (EEG) or, less frequently, intracranial EEG (iEEG). These rhythms reflect synchrony across populations of neurons and are thought to play a fundamental role in regulating which neurons are excitable at specific times, allowing the formation of integrated networks of neuronal subpopulations across the brain [2]. Alpha rhythms are found across the brain and are associated with multiple functions, including attention [3] and sensory gating, where higher alpha power is thought to reflect stronger inhibition of incoming sensory input and lower cortical excitability—providing an important framework for interpreting alpha changes in pain [4]. Alpha 1 power (7–10 Hz), measured over temporal and somatosensory regions, has been found to negatively correlate with pain ratings both at rest and during phasic thermal stimulation [5]. Alpha amplitude is also shaped by cognitive and contextual factors, decreasing with attention to pain [6] and increasing with the expectation of pain relief [7]. A slowing of the individual peak alpha frequency (PAF)—reported in chronic pain conditions [8,9,10,11,12,13]—often occurs with increased theta and gamma activity, forming a pattern known as thalamocortical dysrhythmia, which is thought to reflect altered thalamic bursting and cortical hyperexcitability. In healthy individuals, lower PAF has been associated with increased pain sensitivity [14] and reduced nociceptive facilitation [15].
These associations offer promise for understanding pain and provide targets for neuromodulatory interventions. However, there is still uncertainty about how consistently brain oscillations relate to pain. Recent large-scale studies report no significant relationship between resting PAF and pain sensitivity, supporting the conclusion that no EEG biomarker, including alpha slowing, has yet been firmly established as a reliable or generalizable marker of chronic pain [16,17]. Instead, alpha dynamics appear to reflect broader cross-frequency reorganization within cortical networks. For example, Bott et al. [18] demonstrated that expectations modulate prefrontal to somatosensory connectivity at alpha frequencies, whereas gamma band coupling encodes prediction errors, consistent with predictive coding models of pain. In clinical populations, frequency-specific patterns further highlight distributed network involvement. In chronic low back pain, theta and alpha activity in the orbitofrontal cortex were linked to localized hypersensitivity, while anterior cingulate gamma and dorsolateral prefrontal theta tracked generalized hyperalgesia [19]. In neuropathic pain, higher alpha and lower delta and theta power were associated with increased pain intensity [20]. Thus, a growing body of evidence is at odds with the simple characterization of pain as a high-theta, low-alpha phenomenon. The source locations of measured rhythms, individual differences, and pain types and states may matter in complex ways. Human iEEG work further underscores this heterogeneity in oscillatory correlates of pain across individuals [21], though there may be some consistency across individuals as well. In Shirvalkar et al.’s study [21], which involved sustained iEEG recording in individuals with chronic pain across ~120 days, the most consistent correlates of pain were decreased anterior cingulate alpha/beta power for experimental heat pain and reduced orbitofrontal delta for chronic spontaneous pain.
Given the importance of brain rhythms in circuit integration and behavior, including pain, transcranial alternating current stimulation (tACS) provides a relatively unique and important opportunity to stimulate the brain at particular frequencies [22]. tACS is a noninvasive method that delivers low-intensity sinusoidal electrical currents through scalp electrodes. When tACS is applied at a frequency close to the brain’s own ongoing rhythm, it can entrain neuronal activity by aligning the phase of neural oscillations with the stimulation waveform [23]. This effect is believed to follow the so-called Arnold tongue principle, in which the strength of entrainment increases as the stimulation frequency more closely matches the endogenous oscillatory frequency [24,25]. Another proposed mechanism involves the facilitation of synaptic plasticity via spike-timing-dependent plasticity (STDP), which may allow tACS to enhance oscillatory power at the stimulated frequency [26]. The application of tACS at the PAF has been proposed to promote effective neuronal synchronization [27], to stabilize PAF against potential slowing [28], and to augment its power and gating functions. More broadly, tACS at frequencies beyond alpha may be warranted, as pain-related cortical networks show complex cross-frequency reorganizations rather than isolated alpha-band alterations [18,19,20,21].
2. Methods (Narrative Literature Approach)
Literature was identified through iterative searches in PubMed using combinations of keywords including “tACS”, “transcranial alternating current stimulation”, “pain”, “chronic pain”, “experimental pain”, “EEG”, and “neural oscillations”. Additional articles were identified from the reference lists of the analyzed articles. Searches focused on peer-reviewed studies in the literature published up to November 2025. We included randomized, controlled studies examining the impact of tACS on either experimentally induced pain outcomes in healthy adults or clinical pain outcomes in chronic pain patient groups, relative to a control condition. Control conditions included either sham stimulation or an alternative active comparator transcranial electrical stimulation (tES) condition.
3. Results: Current Evidence on the Effects of tACS on Experimental and Clinical Pain
Across the growing literature on tACS, studies have examined whether rhythmic electrical stimulation can alter pain perception, modulate oscillatory activity, or reduce chronic pain symptoms. Although findings are heterogeneous, converging evidence indicates that tACS can modulate pain perception and pain-related neural activity under specific contextual and stimulation conditions. Specifically, outcome variability appears to depend on stimulation parameters (frequency, montage, intensity, duration), individual neurophysiology, and task.
Table 1 summarizes the key design features and outcomes across all published tACS experimental and clinical pain studies included in this review.
These studies exhibit substantial methodological variability. Sample sizes are typically small, and stimulation parameters, including frequency, intensity, montage, electrode configuration, and duration, vary widely. Power is also likely to be an issue. To detect a moderate effect size of Cohen’s d = 0.5 with 80% power, N = 34 participants are required for a within-person crossover, and N = 130 (65 per group) are required for a between-person comparison across groups. By this standard, only two studies in Table 1 are adequately powered to detect moderate effect sizes.
3.1. Evidence from Experimental Pain Studies
Alpha-frequency stimulation over the somatosensory cortex has shown mixed results. Arendsen et al. [29] reported a reduction in both pain intensity and unpleasantness during α-tACS, but only under conditions of uncertain pain expectation, suggesting an interaction between oscillatory modulation and cognitive context. In contrast, May et al. [32] found no modulation of pain or autonomic responses during α- or γ-tACS over the prefrontal cortex or S1, with Bayesian analyses supporting the null hypothesis for most conditions. Some studies targeting SM1 with α-tACS also showed dissociations between neural and behavioral effects. Peng et al. [33] reported reduced pain-evoked activity and decreased SM1-network connectivity, although pain ratings did not significantly differ from sham. Fathi et al. [28], using individualized somatosensory PAF stimulation, likewise observed no overall analgesic effect, although exploratory analyses suggested reduced sensitization in women and possible stabilization of both alpha rhythms and pain perception. Li et al. [31] reported no reduction in pain but observed disruption of laser-evoked potential habituation and changes in α_1_ and θ activity during anticipation. Mediation analyses further suggested competing influences on pain, with enhanced α_1_ oscillations tending toward analgesia, whereas increased θ activity was associated with hyperalgesia.
Some studies have shown more robust perceptual effects using longer stimulation durations and high-definition (HD) montages. Qi et al. [34] reported that 10 Hz HD-tACS over SM1 reduced capsaicin-induced pain, and resting α power and pain-related θ modulation predicted individual responsiveness. Sun et al. [35] found that HD-tACS over the dorsolateral prefrontal cortex (DLPFC) reduced pain and produced delayed increases in low γ power, although EEG was recorded only at rest.
Other frequency bands have also been explored. Ikarashi et al. [30] demonstrated that θ- and β-tACS over the DLPFC increased heat pain thresholds, with θ-tACS showing an inverted U-shaped relationship between modeled electric field strength and analgesic efficacy, suggesting that both insufficient and excessive stimulation can reduce efficacy.
Finally, interpersonal and affective factors can shape tACS outcomes. Takeuchi and Terui [36] showed that synchronous dual-brain α-tACS enhanced social touch-induced analgesia in low-empathy individuals, even though overall pain ratings did not differ across conditions. These results highlight that tACS effects may depend not only on stimulation parameters but also on social and emotional context.
Together, findings on experimental pain show that tACS can alter pain-related neural activity and occasionally reduce pain perception, but results vary considerably across studies. More consistent effects may emerge when stimulation is longer, spatially focused, tailored to individual oscillatory properties, or applied in contexts that shape pain processing, such as expectation, uncertainty or social interaction.
3.2. Evidence from Chronic Pain Studies
Clinical studies generally provide more consistent indications that tACS can influence ongoing chronic pain symptoms, although sample sizes are small and protocols vary widely.
In chronic low back pain, Ahn et al. [37] found that 10 Hz bifrontal tACS reduced pain ratings relative to sham and increased somatosensory α power, with greater α enhancement predicting larger reductions in pain and disability. Prim et al. [39] also applied 10 Hz tACS over prefrontal regions and observed autonomic modulation (increased heart rate variability) and a higher proportion of clinical responders in the active condition, although group-level pain reductions did not reach statistical significance.
Fibromyalgia studies provide mixed but informative results. Bernardi et al. [39] tailored the frequency of stimulation using 4 Hz for slow-rhythm abnormalities and 30 Hz for fast-rhythm abnormalities and observed reductions in pain intensity, improvements in cognitive and affective symptoms, and increases in α_1_ power, with effects diminishing after four weeks. In contrast, Lin et al. [40] applied 50 Hz HD-tACS over M1 and found no significant differences between active and sham overall, although active stimulation reduced fibromyalgia impact scores relative to baseline.
Clinical protocols also vary widely in stimulation frequency, intensity, montage, electrode configuration, and number of sessions, ranging from single-session interventions to multi-day treatment protocols. Despite this heterogeneity, several patterns emerge. Longer or repeated stimulation tends to show more consistent symptom improvement. Tailoring stimulation frequency based on patient stratification may yield stronger effects than using fixed frequencies. While increases in α power following bifrontal stimulation and normalization of abnormal oscillatory patterns following M1 stimulation have been associated with reductions in pain, further studies are required to confirm the robustness and generalizability of these associations.
4. Discussion: Sources of Inconsistencies, Limitations, and Going Beyond Conventional tACS
Uncertainty about the robustness and mechanisms of tES effects is not unique to pain research and has been emphasized in recent general reviews of the field. Bland and Sale [42] highlighted several fundamental challenges for tACS, including weak cortical currents, limitations in concurrent neuroimaging due to stimulation artifacts, and the difficulty of dissociating genuine neural entrainment from indirect or peripheral effects. Potential methodological solutions have also been discussed. Here, we focus on how these general challenges extend to pain research and consider recent developments in the field that may help improve mechanistic clarity and reproducibility.
4.1. Mechanistic Uncertainty Driven by Target and Frequency Variability
Pain processing is not localized in a single brain region, but it is highly distributed and involves contributions from sensorimotor, prefrontal, limbic, and cingulo-opercular circuits, each oscillating at multiple characteristic rhythms [1]. Moreover, individuals suffering from chronic pain can also exhibit elevated activity in limbic brain regions [43,44,45] and other brain network-level abnormalities [46,47,48,49,50], further complicating the search for analgesic targets and frequencies for clinical applications.
Although alpha activity prior to pain onset correlates with lower pain ratings in some studies [5,51,52], somatosensory alpha-tACS has not consistently modulated pain. For instance, rigorous sham-controlled studies of tonic pain failed to show robust analgesic effects for either somatosensory alpha- or prefrontal gamma-tACS, suggesting that simple one-to-one mappings between rhythm and perception may be insufficient [32]. Power is likely an issue, however, as mentioned above, and larger pre-registered studies must be conducted to more definitively address this issue. Complementary imaging-based work has demonstrated that alpha-tACS over M1 reduces both bilateral SM1 activity and connectivity with pain-related regions, with changes emerging most prominently after stimulation rather than during, pointing toward plasticity mechanisms that extend beyond immediate entrainment [33].
The mechanistic uncertainties surrounding target site selection and frequency tuning therefore represent a critical barrier: without biomarker-guided strategies, stimulation protocols risk inconsistency, with clinical trials reporting modest or null effects despite promising preclinical rationales [53]. Establishing causal links between frequency-specific entrainment, network-level modulation, and pain reduction is therefore essential for moving toward precision neuromodulation.
4.2. Sources of Inter-Individual Variability
A major challenge for effective tACS interventions is the substantial inter-individual variability in stimulation outcomes. This variability arises from a combination of anatomical, physiological, and contextual factors that shape the degree to which electrical fields engage targeted neural circuits. Structural MRI-informed modeling suggests that individual differences in head and brain anatomy lead to substantial variability in intracranial electric field strength and distribution, even when identical stimulation montages are used [30]. As a result, two individuals receiving the same nominal current may experience very different levels of cortical engagement, and behavioral efficacy may follow non-linear patterns, including inverted U relationships between field magnitude and analgesic effects. Such effects, however, remain to be replicated in well-powered studies.
Physiological traits further contribute to variability. Baseline oscillatory dynamics, such as resting alpha power and PAF, are thought to be predictors of pain sensitivity and of responsiveness to stimulation. In controlled experiments, resting alpha power and pain-related theta reactivity jointly mediated responses to alpha HD-tACS over SM1.
Sex and gender differences may also be a factor. Complementary work has shown sex-specific effects in which women exhibited stronger modulation of heat pain thresholds [28]. Biological rhythms, including the menstrual cycle, introduce additional variability through their effects on pain thresholds, cortical excitability, and oscillatory activity.
4.3. Outcomes Measures
Variability in outcome measures and pain models further limits comparability across studies. Experimental paradigms include tonic heat [32], laser stimulation [31,33], electrical pain [36], pressure pain [29], and capsaicin-induced sustained pain [34,35], each engaging partially distinct sensory and cognitive processes. Chronic pain studies involve different diagnoses, symptom profiles, and comorbid conditions. Neural readouts also vary widely, with some studies relying solely on behavioral ratings [29], others measuring resting EEG before and after stimulation [28,32], and still others collecting fMRI activity during pain [33]. Differences in the timing of measurements relative to stimulation further add to inconsistency. Together, these factors make it difficult to identify stimulation features that consistently influence pain or pain-related neural activity. Improved comparability could be achieved by adopting more standardized pain models and outcome measures, clearly distinguishing experimental from clinical pain contexts, and systematically aligning neural readouts and assessment time points with hypothesized mechanisms. Such methodological convergence would facilitate cross-study comparisons, meta-analytic synthesis, and a more coherent understanding of how stimulation parameters relate to pain and pain-related neural activity.
4.4. Blinding Challenges
Blinding quality is another major concern, as sensory experiences during stimulation can differ substantially between active and sham conditions. Because expectation strongly modulates pain perception, inconsistent sham procedures introduce an important confound that may obscure or exaggerate stimulation effects. Across the studies summarized in Table 1, only a limited subset of those reporting significant neural or analgesic effects also demonstrated adequate blinding as verified by formal assessment. Adequate blinding was typically defined by post-session questionnaires assessing participants’ ability to guess stimulation condition, with chance-level accuracy indicating successful blinding. Such assessments were explicitly reported in several double-blind studies, including May et al. [32], Fathi et al. [28], Qi et al. [34], Peng et al. [33], Ahn et al. [37], and Prim et al. [41], all of which employed ramp-up and ramp-down sham protocols designed to mimic initial sensory experiences. Among these, some studies reported clear neural modulation without robust behavioral analgesia [33], whereas others showed pain reduction alongside verified blinding [34,37]. In contrast, multiple studies reporting significant reductions in pain intensity or thresholds did not report formal blinding assessments or relied on single-blind or between-subject designs, including Arendsen et al. [29], Ikarashi et al. [30], Sun et al. [35], and Bernardi et al. [39]. Clinical studies involving longer or repeated stimulation sessions also appear especially vulnerable to partial unblinding, as sensory differences may accumulate over time [37,39,41]. Overall, while several studies in Table 1 report significant neural or analgesic effects, only a small number combine these findings with empirically verified participant blinding, highlighting blinding quality as a critical and inconsistently addressed source of heterogeneity in the current tACS pain literature.
In short, across the 14 sham-controlled tACS pain studies summarized in Table 1, only 6 clearly reported a participant blinding assessment, highlighting the need for more systematic evaluation and transparent reporting of blinding quality in tACS pain research. Blinding in tACS is inherently challenging because cutaneous sensations (tingling, itching, warmth, phosphenes) can differ between active and sham conditions, particularly at higher intensities. While a trade-off between maintaining blinding and ensuring effective tACS application has been discussed in [32], recent unconventional tACS approaches using high-carrier frequencies modulated at lower frequencies (see Section 4.5.1) suggest that higher intensities can be applied while preserving successful blinding [54]. We further recommend the use of an “expectation-neutral blinding assessment,” defined as allocation-belief questions designed to minimize demand characteristics, for example, by allowing “unsure” responses and confidence ratings rather than implying that active and sham conditions must feel different. This approach is motivated by evidence that participant beliefs can influence subjective outcomes [55] and that traditional end-of-study condition guesses may poorly reflect true sensory detectability [56]. Importantly, such blinding procedures should be pilot tested to verify blinding success before full-scale experiments (see [57] for an example).
4.5. Beyond Conventional tACS
Here, we review recent developments in unconventional tACS techniques and discuss their potential to address the inconsistencies and limitations outlined above.
4.5.1. Amplitude-Modulated tACS (AM-tACS)
While many studies focus on aftereffects linked to plasticity, the online effects of tACS—particularly neuronal entrainment during stimulation—are less understood. Directly tracking neural activity during tACS would help clarify these mechanisms, but concurrent EEG recordings are technically challenging due to strong stimulation artifacts. Beyond this, current tACS protocols are typically fixed throughout stimulation, whether individualized or not, whereas optimal neuromodulation likely requires parameters that adapt dynamically to ongoing brain activity. Real-time monitoring would enable such adaptive or closed-loop frameworks by aligning stimulation parameters to moment-to-moment oscillatory dynamics.
To overcome stimulation artifacts and enable online monitoring of neuronal entrainment, Witkowski et al. [58] introduced amplitude-modulated tACS (AM-tACS), where the intended low-frequency stimulation pattern is embedded within a high-frequency carrier signal. The high-frequency carrier is set sufficiently high to avoid phase locking of neuronal activity to the carrier. For recordings, applying an appropriate low-pass filter removes the high-frequency components of the stimulation signal. (Figure 1). However, nonlinearities in stimulation and recording hardware can introduce residual artifacts, and additional denoising approaches have been developed to more reliably recover the neural activity [54,59,60].
While AM-tACS provides a valuable tool for artifact-free recording, its efficacy as a neuromodulatory technique relative to conventional tACS requires further validation [61].
4.5.2. Phase-Dependent and State-Dependent Modulation in tACS
While tACS is often assumed to entrain brain oscillations at their endogenous frequency, recent evidence indicates that it interacts with ongoing neural activity rather than merely amplifying it. Using electrophysiological recordings in awake rhesus monkeys, Krause et al. [62] demonstrated that tACS does not simply impose rhythmicity on neurons but instead competes with intrinsic oscillations for control of spike timing. These recordings revealed that neurons with weak baseline entrainment became more rhythmic during tACS, whereas strongly entrained neurons often lost synchrony. The findings explain variability in human tACS/EEG studies and suggest opportunities for both strengthening and desynchronizing neural activity.
In humans, Fiene et al. [63] provided complementary evidence by pairing visual flicker with occipital tACS at different phase shifts. They showed that steady-state visual responses were modulated in a phase-dependent manner and that the optimal phase shift correlated with each participant’s cortical response delay. These findings highlight that the effectiveness of tACS depends critically on the timing of stimulation relative to ongoing brain rhythms. Haslacher et al. [54,60] provided evidence that the impact of tACS critically depends on its phase relationship to ongoing oscillations. They showed that stimulation at specific phases could enhance or suppress both neural activity and perceptual dominance and later demonstrated that working memory performance was maximized at distinct phase lags that aligned with changes in frontoparietal alpha synchrony. Another example showing the importance of timing is provided by Schreglmann et al. [64], who demonstrated that non-invasive cerebellar tACS phase-locked to the tremor rhythm in essential tremor patients suppressed tremor amplitude by disrupting the temporal coherence of pathological oscillations, with the effective phase lag varying across individuals.
Together, these studies demonstrate that the efficacy and even the direction of tACS effects (enhancement or suppression) are not fixed but depend on phase alignment with ongoing neural activity and brain state, varying not only across individuals but also across time within the same individual. For pain modulation, this implies that the analgesic effect of, for example, alpha-tACS may depend on applying the stimulation at the correct phase of the individual’s ongoing sensorimotor alpha rhythm. Current protocols do not account for this, potentially explaining why some participants respond and others do not. At the same time, state-dependent suppression can be therapeutically exploited. For instance, in neuropathic pain conditions characterized by thalamocortical dysrhythmia, tACS could be tuned to selectively suppress aberrant low-frequency theta or delta oscillations.
4.5.3. Temporal Interference Stimulation (TIS)
In general, noninvasive brain stimulation techniques lack the capability to target deep brain regions effectively without affecting surrounding areas. As pain encompasses sensory, affective, and cognitive dimensions, effective modulation may require selectively targeting brain regions involved in these processes. One such region is the insula, a key hub for integrating sensori-discriminative and affective-cognitive aspects of pain [65,66,67]. The posterior insula is more strongly associated with processing nociceptive input [68], whereas the anterior insula is most closely associated with affective and evaluative aspects [69]. Dysregulation of insular activity is often observed in chronic pain [66], and lesions in the posterior insula can abolish the unpleasantness of pain while preserving sensory detection [70]. Furthermore, iEEG evidence suggests that nociceptive pain preferentially modulates theta and alpha oscillations in the dorsal posterior insula [71], indicating that this subregion could be a promising stimulation target.
Targeting the insula for pain treatment has been investigated in several studies. A recent rodent study showed that repetitive insular cortex stimulation alleviated neuropathic pain by normalizing synaptic plasticity, specifically reducing phosphorylated CRMP2, AMPAR, and NR2B expression, and dampening insular hyperactivity [72]. A pilot intracranial study in humans showed that anterior insula stimulation raised heat pain thresholds without adverse effects, supporting its role in the affective-cognitive modulation of pain [73]. Together, these converging lines of evidence underscore the insula as a promising therapeutic hub for targeting both sensory and affective dimensions of chronic pain.
However, invasive insular stimulation for pain remains limited in scope, and its clinical application is constrained by surgical risks and outcome variability. A noninvasive method capable of selectively modulating the insula without affecting the surrounding cortex would therefore offer a safer and more accessible therapeutic option while also enabling deeper investigation into the insula’s mechanistic role in pain. The same principle applies to other regions important in pain construction.
Temporal interference stimulation (TIS) builds on the principle of amplitude-modulated tACS, where two electrode pairs deliver high-frequency currents with slightly different frequencies, creating a low-frequency envelope at their intersections that can modulate neural activity (Figure 2). Grossman et al. demonstrated in mice that such deep activation is possible without engaging in the overlying cortex and that by adjusting current ratios, the stimulation locus can be shifted [74], opening the possibility to selectively target subregions such as specific insular subregions. This steerability and focus make TIS a particularly attractive approach for noninvasively modulating deep structures involved in pain. A systematic review of human TIS studies [75] suggests that although preliminary findings indicate frequency- and target-specific modulation of cortical and subcortical activity, the evidence remains limited by small sample sizes and methodological heterogeneity. Larger, well-controlled trials are needed to establish clinical efficacy.
4.6. Recommendations for Future Studies
Taken together, inconsistent findings likely arise from a combination of heterogeneous stimulation parameters, variable sham procedures, insufficient exploration of dose–response and duration effects, limited verification of neural target engagement, diverse pain models, and substantial inter-individual differences. Future research would benefit from systematic testing of stimulation duration and intensity, clearer reporting of modeled intracranial electric fields, standardized sham protocols, larger and adequately powered samples, and direct measures of neural engagement during or immediately after stimulation. Incorporating individualized oscillatory biomarkers, anatomical modeling, and repeated session designs will also help clarify the conditions under which tACS reliably influences pain networks. Addressing these methodological challenges in a coordinated, evidence-based manner is essential to determine whether tACS can provide consistent and meaningful pain modulation. Emerging tACS techniques are increasingly designed to target complex oscillatory dynamics and interactions [76]. Beyond the approaches discussed in this study, other strategies may help advance our understanding of pain and the role of oscillations in its processing. One example is cross-frequency coupled tACS (CFC-tACS) [77], which, rather than applying a single frequency, targets coupled oscillations to modulate their interactions. Another example is broadband tACS, which uses non-sinusoidal patterns that deliver a range of frequencies instead of a narrow sinusoid, thereby mimicking the spectral complexity of natural neural activity. Applying these techniques to pain requires systematic investigation to evaluate their potential contributions to pain modulation mechanisms.
5. Conclusions
Across experimental and clinical studies, tACS can modulate oscillatory activity within pain-related networks, but its behavioral effects on pain perception remain variable. Evidence suggests that stimulation outcomes depend on the interaction between stimulation parameters, individual neurophysiology, and contextual brain state. Studies using longer and more focal stimulation, individualized frequency alignment, verification of neural target engagement, and integration with behavioral or cognitive interventions such as expectancy training or pain distraction tend to show more reliable neural and, in some cases, analgesic effects. Progress toward clinical translation will require adequately powered and preregistered studies that combine electric-field modeling with real-time verification of target engagement. Integrating phase- and state-dependent stimulation strategies, including adaptive or closed-loop approaches, represents a critical next step toward precision neuromodulation for pain.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ploner M. Sorg C. Gross J. Brain Rhythms of Pain Trends Cogn. Sci.20172110011010.1016/j.tics.2016.12.00128025007 PMC 5374269 · doi ↗ · pubmed ↗
- 2Fries P. Rhythms for Cognition: Communication through Coherence Neuron 20158822023510.1016/j.neuron.2015.09.03426447583 PMC 4605134 · doi ↗ · pubmed ↗
- 3Foxe J.J. Snyder A.C. The Role of Alpha-Band Brain Oscillations as a Sensory Suppression Mechanism during Selective Attention Front. Psychol.2011215410.3389/fpsyg.2011.0015421779269 PMC 3132683 · doi ↗ · pubmed ↗
- 4Jensen O. Mazaheri A. Shaping Functional Architecture by Oscillatory Alpha Activity: Gating by Inhibition Front. Hum. Neurosci.2010418610.3389/fnhum.2010.0018621119777 PMC 2990626 · doi ↗ · pubmed ↗
- 5Nir R.R. Sinai A. Moont R. Harari E. Yarnitsky D. Tonic Pain and Continuous EEG: Prediction of Subjective Pain Perception by Alpha-1 Power during Stimulation and at Rest Clin. Neurophysiol.201212360561210.1016/j.clinph.2011.08.00621889398 · doi ↗ · pubmed ↗
- 6May E.S. Butz M. Kahlbrock N. Hoogenboom N. Brenner M. Schnitzler A. Pre- and Post-Stimulus Alpha Activity Shows Differential Modulation with Spatial Attention during the Processing of Pain Neuroimage 2012621965197410.1016/j.neuroimage.2012.05.07122659486 · doi ↗ · pubmed ↗
- 7Huneke N.T.M. Brown C.A. Burford E. Watson A. Trujillo-Barreto N.J. El-Deredy W. Jones A.K.P. Experimental Placebo Analgesia Changes Resting-State Alpha Oscillations P Lo S ONE 20138 e 7827810.1371/journal.pone.007827824147129 PMC 3795660 · doi ↗ · pubmed ↗
- 8Walton K.D. Dubois M. Llinás R.R. Abnormal Thalamocortical Activity in Patients with Complex Regional Pain Syndrome (CRPS) Type I Pain 2010150415110.1016/j.pain.2010.02.02320338687 · doi ↗ · pubmed ↗