Metformin for Longevity and Sarcopenia: A Therapeutic Paradox in Aging
Song-Yi Han, Mukesh Kumar Yadav, Jing-Hua Wang

TL;DR
Metformin may help protect against aging-related muscle loss in some people but could hinder muscle growth in others, highlighting the need for personalized approaches.
Contribution
The paper identifies a therapeutic paradox in metformin's effects on muscle aging, suggesting context-dependent outcomes in different populations.
Findings
Metformin is linked to reduced sarcopenia in insulin-resistant older adults through metabolic and anti-inflammatory effects.
In healthy older adults, metformin may suppress muscle hypertrophy by activating AMPK and inhibiting mTORC1.
The drug's effects highlight a paradoxical role in aging, requiring precision strategies for optimal geropharmacology.
Abstract
Metformin is a first-line oral antidiabetic agent that has attracted increasing interest as a potential geroprotective therapy due to its ability to improve metabolic homeostasis, reduce oxidative stress, and attenuate chronic inflammation. However, its role in skeletal muscle aging and sarcopenia remains controversial. Observational and epidemiological studies suggest that metformin use is associated with a lower prevalence of sarcopenia, particularly in metabolically compromised or insulin-resistant older populations, where improvements in systemic metabolism and inflammatory burden may indirectly support muscle quality and function. In contrast, randomized interventional trials in metabolically healthy older adults indicate that metformin can blunt resistance exercise–induced muscle hypertrophy and protein synthesis, likely through sustained activation of AMP-activated protein kinase…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
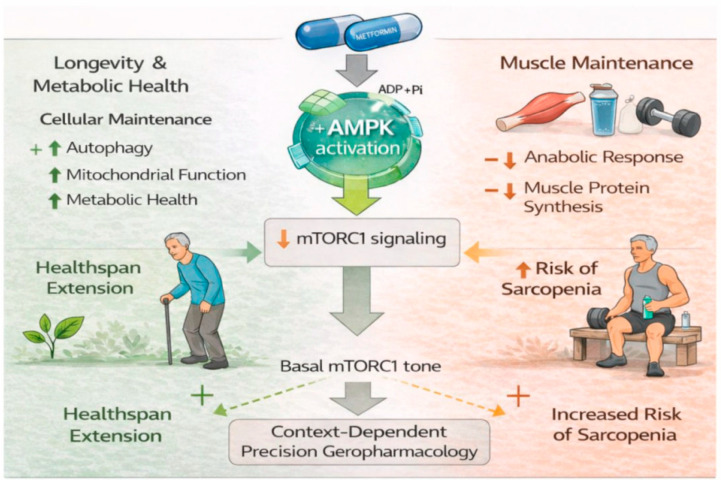 Figure 1
Figure 1- —National Research Foundation of Korea
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMetabolism, Diabetes, and Cancer · Nutrition and Health in Aging · Muscle Physiology and Disorders
1. Introduction
Initially prescribed for type 2 diabetes mellitus (T2DM), metformin has recently gained significant attention as a potential geroprotective agent [1]. It produces multiple effects by inhibiting mitochondrial complex I, decreasing hepatic gluconeogenesis, activating AMP-activated protein kinase (AMPK), and influencing inflammatory and oxidative stress pathways [2,3,4]. The Targeting Aging with Metformin (TAME) trial and related studies have expanded its reputation beyond glycemic control to possible anti-aging uses [5]. However, skeletal muscle (the largest metabolic organ) complicates this issue. Age-related muscle loss, or sarcopenia, not only leads to frailty and disability but also worsens metabolic disorders [6]. Intriguingly, while large observational studies suggest protective effects, intervention trials in older adults show that metformin may inhibit muscle gains from resistance training [7,8]. These seemingly contradictory findings suggest that the effects of metformin on aging are not uniform but instead vary systematically across disease environments and experimental conditions.
As shown in Table 1, a representative overview of selected recent studies illustrates the heterogeneous effects of metformin across aging-related conditions. Meta-analyses and clinical studies report inconsistent or modest benefits in neurodegenerative and cognitive disorders, with several investigations indicating no overall protective effect or even potentially adverse associations in specific subpopulations [9,10,11,12]. In addition, preclinical models demonstrate relatively consistent beneficial effects, including reduced tau pathology, enhanced autophagy, and improved cellular stress resistance, largely mediated through AMPK activation, mTORC1 suppression, and anti-inflammatory pathways [13,14,15]. Notably, these effects vary according to disease stage, age, experimental model, and dosage, underscoring the absence of a uniform therapeutic profile. Accordingly, this heterogeneity indicates that metformin may not represent a “universal” solution for extending lifespan.
2. The Paradox: Restrained Muscle Adaptation and the AMPK-mTOR Signaling Dilemma
In a randomized controlled trial (the MASTERS study), older adults who received 1700 mg/day of metformin during 14 weeks of progressive resistance training gained significantly less muscle mass and cross-sectional area compared to the placebo group [8]. Transcriptomic profiling further revealed attenuated activation of anabolic and mitochondrial biogenesis pathways in the metformin group [19]. Mechanistically, this paradox can be partially explained by dysregulated AMPK–mTORC1 crosstalk [20]. Metformin activates AMPK, which, while beneficial for metabolic regulation, concurrently inhibits mTORC1, the central driver of muscle protein synthesis [21,22]. In healthy muscle, this may blunt hypertrophic adaptation to mechanical loading. Therefore, the same signaling pathway that provides metabolic protection may impair muscle growth when anabolic stimulation is required.
Furthermore, in vitro studies confirm this concern. Metformin exposure in C2C12 myotubes increases FoxO3a expression and nuclear localization and enhances its binding to the myostatin promoter, thereby activating a FoxO3a-dependent catabolic program that is classically linked to induction of the muscle atrophy-related E3 ubiquitin ligases MuRF1 and Atrogin-1 [23,24]. These molecular patterns are consistent with disuse or fasting-induced atrophy, raising the possibility that chronic metformin use under specific conditions could mimic aspects of catabolic stress [25]. Even more patently, in clinical trials combining metformin with resistance exercise, older subjects exhibited reduced gains in strength and muscle hypertrophy, despite preserved insulin sensitivity [8,26]. This dissociation is consistent with what may be conceptualized as a ‘geroprotection paradox’, in which cellular signaling is improved while tissue-level adaptation is weakened.
In support of this concept, the mechanistic evidence summarized in Table 2 shows the strongly context-dependent and often bidirectional effects of metformin on skeletal muscle aging. Across in vivo, in vitro, and limited clinical observations, metformin is frequently associated with partial improvements in muscle quality, cellular stress resistance, and metabolic maintenance, such as enhanced autophagy, mitochondrial function, and reduced inflammatory burden, particularly in aging or metabolically compromised models, largely through AMPK activation and related pathways [27,28,29]. However, these cellular benefits are not consistently accompanied by enhanced muscle mass or anabolic capacity, as metformin exposure in several settings is linked to attenuated myogenic signaling, altered satellite cell–related markers, activation of catabolic transcriptional programs, or negligible effects on fat-free mass in randomized trials [30,31,32]. Importantly, these outcomes vary substantially with experimental model, age, metabolic status, treatment duration, and dosage, emphasizing the absence of a uniform therapeutic profile and providing putative mechanistic support for the dissociation between improved cellular homeostasis and blunted hypertrophic adaptation described above.
Beyond signaling mechanisms, population characteristics further shape the apparent paradox observed across study designs. The apparent discrepancy between observational and interventional findings can be largely attributed to differences in the populations studied and the outcomes assessed. Observational studies predominantly involve older adults with metabolic comorbidities, in whom sarcopenia develops in parallel with systemic metabolic stress and inflammation; in this context, metformin may indirectly preserve muscle quality and reduce sarcopenia risk through its metabolic and anti-inflammatory effects. By contrast, interventional trials typically enroll metabolically healthy, physically active older adults undergoing resistance training, where muscle adaptation relies on intact anabolic signaling and transient mTORC1 activation. In this setting, chronic AMPK activation by metformin may attenuate mTORC1-driven protein synthesis and blunt hypertrophic responses despite improved metabolic indices. Taken together, these findings are not contradictory but complementary, reflecting fundamental differences in biological context and study design. This perspective sets the stage for a broader discussion of context-dependent modifiers, including nutrition, exercise modality, and the gut microbiome.
3. Understanding Context-Dependent Metformin Effects in Aging
One key reason for the inconsistent findings across studies in Table 2 is that the effects of metformin strongly depend on context. Its influence differs with age, metabolic health, nutritional status, physical activity level, and gut microbiota. Together, these factors influence the balance between muscle-building and muscle-breaking pathways in aging muscle, which ultimately determines whether metformin is beneficial or harmful.
In metabolically unhealthy or diabetic individuals, insulin-sensitizing and anti-inflammatory properties of metformin may partially restore impaired anabolic responsiveness [38]. By improving systemic glucose handling, reducing low-grade inflammation, and alleviating lipid-induced metabolic stress, metformin may indirectly enhance muscle quality, even in the absence of robust gains in muscle mass [39]. Such effects are consistent with observations in sarcopenic or metabolically compromised models, where improvements in muscle function and cellular integrity often outweigh changes in absolute muscle quantity [36]. In contrast, in metabolically healthy or physically trained older adults, chronic activation of AMPK by metformin may blunt the mTORC1 signaling required for resistance exercise-induced muscle hypertrophy [8]. Because adaptive muscle growth relies on transient mTORC1 activation, sustained AMPK dominance may attenuate training responsiveness, providing a mechanistic basis for reports of diminished hypertrophic adaptations in this population. Interpretation is further complicated by sex-specific and dose-dependent factors. Age-related changes in pharmacokinetics, renal clearance, and muscle perfusion may alter tissue exposure to metformin, while hormonal differences between males and females could modulate downstream signaling responses [40,41,42]. These variables are rarely harmonized across studies, contributing additional layers of heterogeneity.
Nutritional status also represents a central determinant of this balance. Adequate protein intake is required to elicit postprandial mTORC1 activation and sustain muscle protein synthesis in older adults, who already exhibit age-related anabolic resistance [43]. Therefore, under conditions of insufficient protein or energy intake, metformin-mediated suppression of basal mTORC1 tone may further inhibit anabolic signaling capacity, thereby predisposing aging muscle to protein loss.
Moreover, the interaction between metformin and exercise is also critical. Aerobic exercise, which relies primarily on mitochondrial oxidative metabolism, may act synergistically with the mitochondrial and redox-modulating effects of metformin [44]. In contrast, resistance training relies on short-lived mTOR activation for muscle remodeling, a process that metformin can suppress [45]. However, muscle-intrinsic signaling pathways alone may not fully account for the divergent muscular outcomes observed across populations [46,47].
Metformin is well known to remodel gut microbial composition and function, including enrichment of short-chain fatty acid (SCFA)–producing taxa and alterations in bile acid metabolism, both of which can influence host energy homeostasis, inflammation, and muscle metabolism [46]. Exercise itself is a potent modulator of the gut microbiome, and the combined effects of metformin and specific exercise modalities may generate distinct microbial and metabolite profiles that differentially impact muscle anabolic signaling [47,48]. Although these interactions remain incompletely characterized, they introduce an additional layer of biological variability that may help explain population-specific responses to metformin.
Building on these observations, the gut microbiome can be further conceptualized as a signaling intermediary linking metformin exposure to skeletal muscle adaptation. Metformin-induced shifts in microbial composition, including increased SCFA production and remodeling of bile acid–transforming communities. These changes may influence host metabolism through elevated circulating SCFAs and modified bile acid pools, which regulate systemic inflammation, insulin sensitivity, and energy homeostasis. From a muscle signaling perspective, SCFAs such as butyrate and propionate may indirectly modulate skeletal muscle anabolic responsiveness by attenuating chronic low-grade inflammation and improving insulin signaling, thereby lowering basal metabolic stress in aging muscle. In parallel, and through partially overlapping mechanisms, microbiota-driven alterations in bile acid composition may influence muscle metabolism via FXR- and TGR5-dependent pathways, with downstream effects on mitochondrial function, oxidative capacity, and AMPK activity. Collectively, these microbiome-derived signals may shift the balance between AMPK-mediated metabolic stress responses and mTORC1-dependent anabolic signaling in skeletal muscle.
Importantly, this framework does not propose a deterministic causal pathway, but rather a hypothesis-driven model in which gut microbiota composition acts as a contextual modifier of metformin’s muscular effects. Depending on baseline metabolic status, exercise context, and microbial configuration, metformin-induced microbiome remodeling may either support muscle maintenance in metabolically compromised individuals or fail to compensate for AMPK-driven suppression of hypertrophic signaling in physically active older adults. As such, this model highlights the gut–muscle axis as a testable mechanistic layer within precision geropharmacology and underscores the need for future studies integrating microbiome profiling with muscle signaling and functional endpoints.
Overall, these observations argue strongly against a “one-size-fits-all” strategy for metformin use in sarcopenia and highlight the necessity of precision-based approaches that consider metabolic status, nutritional adequacy, exercise type, age, and sex. The growing recognition of context-dependent responses to metformin highlights the need for future precision geropharmacological approaches supported by actionable biomarkers. Potential candidates include baseline AMPK-mTORC1 signaling activity, systemic inflammatory markers (e.g., CRP and IL-6), metabolic indices of insulin resistance, and emerging gut microbiome-derived signatures. These biomarkers may enable stratification of individuals who derive metabolic benefit from metformin versus those at risk of impaired anabolic adaptation, and should be evaluated in future studies.
4. Redefining Geroprotection in Muscle Aging
The binary view of metformin should be abandoned as either a “beneficial” or “harmful” drug for skeletal muscle. Instead, a nuanced framework is needed that integrates dose–response relationships, timing of administration, and combined lifestyle interventions. Emerging evidence suggests that intermittent or lower-dose metformin may mitigate the hypertrophy-blunting effect while preserving mitochondrial and anti-inflammatory advantages [49]. Alternatively, co-treatment strategies that transiently relieve AMPK-mediated suppression could achieve balanced outcomes, such as metformin cycling or combination with nutraceutical AMPK modulators [50,51].
Moreover, hybrid compounds like RJx-01 (metformin + galantamine) have demonstrated synergistic effects in preclinical models, improving autophagy, neuromuscular junction integrity, and muscle force generation beyond what either compound achieves alone [27]. Such findings indicate that polypharmacological approaches may overcome metformin’s intrinsic limitations.
Importantly, future research must differentiate between “muscle aging” (cellular senescence, mitochondrial decline, autophagic failure) and “sarcopenia” (loss of mass and function). While these phenomena overlap, metformin may target the former more effectively than the latter. Longitudinal human trials that integrate muscle omics, imaging, and functional endpoints are urgently needed to determine whether metformin delays true sarcopenia or merely improves metabolic biomarkers.
One promising strategy to reconcile the metabolic benefits of metformin with its potential to blunt anabolic adaptation is the exploration of intermittent or cycling dosing paradigms. From a mechanistic perspective, the beneficial effects of metformin on mitochondrial function, inflammation, and metabolic homeostasis are largely mediated through AMPK activation and downstream stress-response pathways, which may not require continuous pharmacological pressure to be sustained. In contrast, chronic AMPK dominance may interfere with the transient mTORC1 activation required for resistance exercise–induced muscle protein synthesis. Intermittent exposure to metformin could therefore represent a theoretical means to retain metabolic protection while permitting periodic restoration of anabolic responsiveness during phases of mechanical loading or nutritional sufficiency. In addition, co-treatment strategies warrant consideration as a conceptual way to balance metformin’s pleiotropic effects. Rather than opposing metformin’s action directly, rational combinations may aim to temporally or contextually offset its anabolic suppression. For example, pairing metformin with nutritional, exercise-based, or pharmacological interventions that support mTORC1 signaling, satellite cell activity, or neuromuscular adaptation could help preserve muscle adaptability without negating metabolic benefits. Importantly, these approaches should not be viewed as fixed therapeutic prescriptions, but as testable hypotheses that underscore the necessity of context-aware intervention design. Within a precision geropharmacology framework, such strategies emphasize timing, physiological state, and adaptive capacity as critical determinants of outcome, reinforcing the view that metformin’s effects on aging muscle are modifiable rather than intrinsically paradoxical.
5. Re-Evaluating the “Anti-Aging” Potential of Metformin
Interest in metformin as a potential universal anti-aging drug has sometimes exceeded the strength of the available evidence. Its use in large longevity trials, such as the TAME study [5], highlights the need to evaluate not only effects on lifespan but also on functional healthspan, especially skeletal muscle function. If a longevity drug weakens muscle adaptation, it may help people live longer but make it harder for them to stay independent and mobile, which are essential for healthy aging. This paradox reflects a broader tension in geroscience: interventions that suppress anabolic signaling to extend lifespan may simultaneously impair tissue repair and regeneration [52]. Because skeletal muscle relies heavily on mechanical loading and anabolic signaling, these trade-offs are particularly important. The effects of Metformin on muscle are therefore unlikely to be uniformly “anti-aging” and instead appear to depend on context, acting primarily as a modulator of cellular energy sensing.
6. Conclusions and Perspective
Taken together, the dual role of metformin as both a metabolic protector and a potential inhibitor of muscle adaptation challenges the notion of universal geroprotection (Figure 1). Consequently, the future of sarcopenia research lies in precision geropharmacology, in which interventions are tailored according to metabolic phenotype, exercise context, gut microbiome configuration, and individual aging trajectories. Rather than asking whether metformin is “good or bad” for aging muscle, more relevant questions are: when, in whom, at what dose, and under which metabolism and gut microbiome context does metformin confer benefit or harm? Hence, future studies should explicitly incorporate microbiome profiling into experimental and clinical designs, alongside integrated multi-omics, imaging, and functional endpoints, to delineate the dynamic drug–exercise–muscle axis. Only by embracing this biological complexity can metformin be transformed from a blunt anti-aging intervention into a refined and personalized strategy for preserving skeletal muscle health across the lifespan.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kulkarni A.S. Gubbi S. Barzilai N. Benefits of Metformin in Attenuating the Hallmarks of Aging Cell Metab.202032153010.1016/j.cmet.2020.04.00132333835 PMC 7347426 · doi ↗ · pubmed ↗
- 2Foretz M. Guigas B. Viollet B. Metformin: Update on mechanisms of action and repurposing potential Nat. Rev. Endocrinol.20231946047610.1038/s 41574-023-00833-437130947 PMC 10153049 · doi ↗ · pubmed ↗
- 3Ansari A. Bose S. Lim S.-K. Wang J.-H. Choi Y.-H. Kim H. Combination of Scutellaria baicalensis and Metformin Ameliorates Diet-Induced Metabolic Dysregulation in Mice via the Gut–Liver–Brain Axis Am. J. Chin. Med.2020481409143310.1142/S 0192415 X 2050069 X 32907360 · doi ↗ · pubmed ↗
- 4Wang J.-H. Bose S. Lim S.-K. Ansari A. Chin Y.-W. Choi H.S. Kim H. Houttuynia cordata Facilitates Metformin on Ameliorating Insulin Resistance Associated with Gut Microbiota Alteration in OLETF Rats Genes 2017823910.3390/genes 810023928937612 PMC 5664089 · doi ↗ · pubmed ↗
- 5Rolland Y. Sierra F. Ferrucci L. Barzilai N. De Cabo R. Mannick J. Oliva A. Evans W. Angioni D. De Souto Barreto P. Challenges in developing Geroscience trials Nat. Commun.202314503810.1038/s 41467-023-39786-737598227 PMC 10439920 · doi ↗ · pubmed ↗
- 6Larsson L. Degens H. Li M. Salviati L. Lee Y.I. Thompson W. Kirkland J.L. Sandri M. Sarcopenia: Aging-Related Loss of Muscle Mass and Function Physiol. Rev.20199942751110.1152/physrev.00061.201730427277 PMC 6442923 · doi ↗ · pubmed ↗
- 7Qaisar R. Javed M. Khan I.M. Ahmad F. Karim A. Metformin improves skeletal muscle and physical capacity by stabilizing neuromuscular junction in older adults Arch. Gerontol. Geriatr.202412710558710.1016/j.archger.2024.10558739084174 · doi ↗ · pubmed ↗
- 8Walton R.G. Dungan C.M. Long D.E. Tuggle S.C. Kosmac K. Peck B.D. Bush H.M. Villasante Tezanos A.G. Mc Gwin G. Windham S.T. Metformin blunts muscle hypertrophy in response to progressive resistance exercise training in older adults: A randomized, double-blind, placebo-controlled, multicenter trial: The MASTERS trial Aging Cell 201918 e 13039 Erratum in Aging Cell 2019, 19, e 13098. https://doi.org/10.1111/acel.13098.10.1111/acel.1303931557380 PMC 6826125 · doi ↗ · pubmed ↗