Complement System Dysregulation in the Immunopathogenesis of Long COVID: Systematic Evidence Synthesis
Kin Israel Notarte, Jesus Alfonso Catahay, Jacqueline Veronica Velasco, Abbygail Therese Ver, Jungwook Lee, John G. Rizk, Giuseppe Lippi, César Fernández-de-las-Peñas

TL;DR
This study reviews evidence that the immune complement system may be dysregulated in people with long COVID, but findings are inconsistent and more research is needed.
Contribution
This is the first systematic review to evaluate the role of long-lasting complement activation in long COVID.
Findings
Elevated markers of classical, alternative, and terminal complement pathways were found in some long COVID patients.
Markers of the lectin pathway showed no significant differences between long COVID patients and controls.
Heterogeneous results and methodological differences highlight the need for further research.
Abstract
Background/Objective: Long COVID is an important cause of disability following SARS-CoV-2 infection; yet, its underlying mechanisms are not completely understood. One proposed mechanism is the long-lasting dysregulation of the immune complement system. This systematic review is the first to summarize the current evidence and evaluate the potential role of long-lasting complement activation in people with long COVID. Methods: A systematic electronic search on PubMed, MEDLINE, CINAHL, and Embase was conducted up to 15 October 2025, to identify studies investigating complement activation in people with the post-COVID-19 condition. The Newcastle–Ottawa Scale was used to evaluate the risk of bias and methodological quality. Results: Among the 247 studies initially identified, eleven met the inclusion criteria, comprising 1435 individuals (age: 48.5 years, 70% females) with long COVID and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
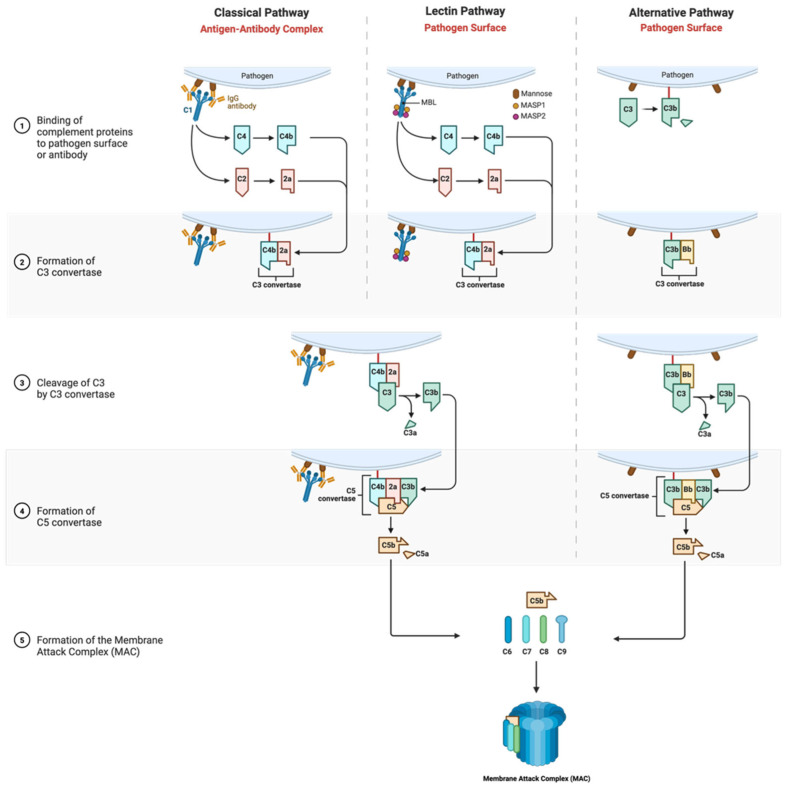 Figure 1
Figure 1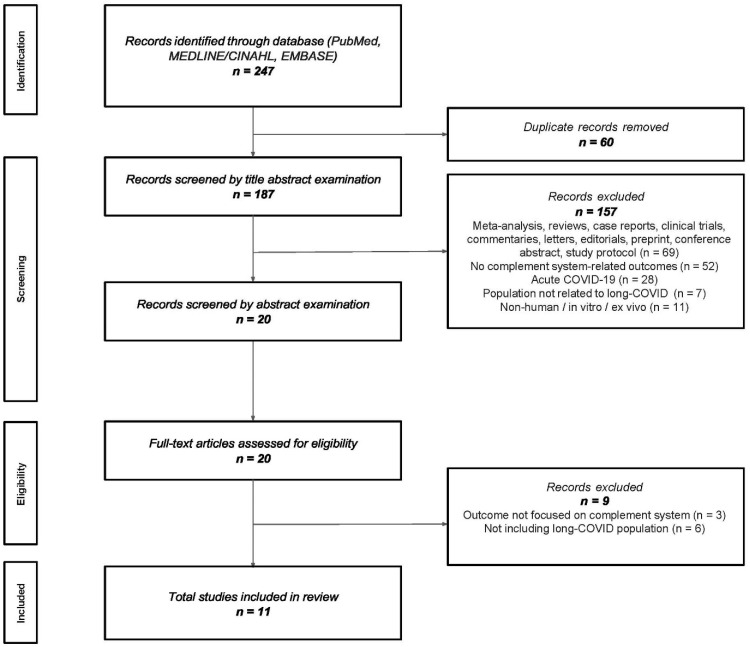 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsComplement system in diseases · Long-Term Effects of COVID-19 · Neuroinflammation and Neurodegeneration Mechanisms
1. Introduction
Long COVID, also known as post-acute COVID-19 syndrome, is characterized by persistent symptoms after the acute phase of a SARS-CoV-2 infection [1]. Commonly reported features include fatigue, dyspnea, cognitive dysfunction, and chest pain [2]. Epidemiological studies estimate that a substantial proportion of individuals experience symptoms after the initial infection, with recent data demonstrating persistence for up to two to three years [2,3]. The underlying mechanisms remain incompletely defined, but several hypotheses, including sustained organ damage, the persistence of viral RNA or proteins, the generation of autoantibodies, the reactivation of latent viruses, and chronic immune or endothelial dysfunction, have been proposed [4,5,6,7]. The complement system is an essential feature of the innate immunity that contributes to pathogen recognition, cell lysis, and the regulation of inflammation [8]. Its activation occurs throughout the classical, lectin, and alternative pathways, which are activated differently but converge to amplify inflammatory responses and generate the membrane attack complex (MAC) [9]. Specifically, the classical pathway is activated by antigen–antibody complexes, the lectin pathway by pathogen-associated carbohydrate patterns, and the alternative pathway by spontaneous activation on microbial or altered host cell surfaces (Figure 1). Regardless of the initiating stimulus, all three pathways converge at C3, leading to C5 cleavage and activation of the terminal pathway, culminating in MAC formation. In the lectin pathway, mannose-binding lectin (MBL) binds to MBL-associated serine proteases (MASPs) to initiate downstream complement activation. Collectively, these tightly regulated processes enable effective immune defense against infection while preserving host tissue integrity.
The complement system is also closely interconnected with the circulatory coagulation system, and its dysregulated activity can promote thrombo-inflammation and tissue injury [10]. Excessive complement activation has been associated with endothelial damage, microvascular thrombosis, or severe outcomes during the acute COVID-19 phase [11]. Emerging evidence suggests that complement dysregulation may also persist in individuals with long COVID, where ongoing inflammation, coagulation abnormalities, and vascular injury are present [12].
To date, no systematic review has synthesized the available evidence on the role of the complement cascade in the immunopathogenesis of long COVID. The objective of this systematic review was to address this gap by evaluating the current data and clarifying the potential contribution of long-lasting complement activation to the persistence of post-COVID-19 symptoms.
2. Methods
A systematic review of studies investigating differences in complement activation in people with post-COVID symptoms according to the 2020 Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) statement was conducted (Supplementary Materials) [13]. The review was registered in the Open Science Framework database (https://osf.io/6s45h). A meta-analysis was not conducted because studies are highly heterogenous in the complement markers assessed, the assays and biological measures used (intact complement components, activation fragments, functional assays, and omics-derived pathway signals), baseline values, follow-up timeframes, and post-COVID outcomes evaluated.
2.1. Literature Search
Literature searches for studies published up to 15 October 2025 were conducted across four databases: PubMed, Medline, CINAHL, and Embase. An experienced health science librarian assisted with the electronic searches. The reference lists of identified studies were also screened for additional studies to be included. Search terms and Boolean operators used in the literature search on each database are outlined in Table 1.
2.2. Selection Criteria
The inclusion criteria were described according to the Population, Intervention, Comparison, and Outcome (PICO) principle:
- Population: Adults (>18 years) who had been previously infected by SARS-CoV-2;
- Intervention: Not applicable;
- Comparison: Not applicable;
- Outcome: Articles investigating complement activation in individuals with post-COVID-19 condition. We used the definition by Soriano et al.: “post-COVID-19 condition occurs in people with a history of probable or confirmed SARS-CoV-2 infection, usually three months from the onset of infection, with symptoms that last for at least two months and cannot be explained by an alternative diagnosis” [14]. Articles should report post-COVID symptoms as well as measure complement activation at any follow-up period post-infection.
2.3. Screening Process, Study Selection, and Data Extraction
Cohort and case–control studies where complement activation in patients who developed post-COVID-19 condition after an acute SARS-CoV-2 infection were included. Papers that are not original articles, such as systematic reviews or meta-analyses, and editorials, were excluded. Studies were also excluded if the samples were not collected from humans. Post-mortem studies were excluded.
The titles and abstracts of each article identified during the literature search were screened by two authors. After removing duplicates, the full text of eligible articles was analyzed by the same authors. For each study, the following data were extracted: authors and country, sample size and setting, participant characteristics (age, sex, demographic, anthropometric, and clinical data), post-COVID symptoms, sample collection methods, diagnostic methods used to assess complement activation, and follow-up duration. Consensus on the selection of included studies and on data extraction should be achieved by both authors. Discrepancies at any stage of the screening process were resolved by asking a third author, if needed.
2.4. Methodological Quality/Risk of Bias
The Newcastle–Ottawa Scale (NOS) was used to evaluate the risk of bias and methodological quality of the included observational studies [15]. Scores were classified as follows: high-quality (7–9 stars), moderate-quality (5–6 stars), and low-quality (≤4 stars). Methodological quality was independently evaluated by two authors, and any discrepancies were resolved through discussion with a third researcher.
3. Results
3.1. Study Selection
The database search initially identified 247 studies. After removing 60 duplicates, 187 studies remained for the title and abstract examination, and then 157 studies were excluded at the title screening because of the type of paper (e.g., meta-analyses, reviews, case reports, clinical trials, commentaries, and editorials), population (acute COVID-19 patients or other population different from long COVID), measurements (not related to the complement system), or the papers being non-human studies. Then, 20 studies underwent an abstract and detailed full-text review. Among these, 9 were further excluded: 3 studies did not assess the complement system biology, and the remaining 6 did not include a definition of the long COVID population. Overall, eleven studies met all the eligibility criteria and were included in the review [12,16,17,18,19,20,21,22,23,24,25]. Figure 2 describes the PRISMA flow diagram.
3.2. Sample Characteristics
A total of 1435 individuals (mean age: 48.5 years old, 70% females) with long COVID and 1124 controls (mean age: 43.6 years, 60% females) were included in the studies. We found 5 studies included hospitalized COVID-19 survivors [12,19,20,21,25], 2 included non-hospitalized survivors [18,22], and the remaining 4 [16,17,23,24] did not specify the participants’ hospitalization status. The most common self-reported post-COVID symptoms included fatigue, dyspnea, brain fog, headache, insomnia, dysgeusia, hair loss, poor concentration, dizziness, sleep disturbance, or gastrointestinal problems (diarrhea, vomiting, and abdominal pain). All studies measured the complement proteins in serum or plasma, but the methods differed, including ELISA [12,18,21,22,23], mass spectrometry [12,17], immunoassay/proteomics [12,19], and other assay platforms [16,17,20,24,25]. Samples were also collected at different time points after the infection, ranging from three months to two years after SARS-CoV-2 infection (mean follow-up: 8.9 months). Table 2 summarizes the main findings of the studies included in this review.
3.3. Methodological Quality
Seven case–control (Table 3) [17,18,19,20,22,25] and four longitudinal cohort (Table 4) [12,16,21,23,24] studies were evaluated. All studies were of a high methodological quality, with scores ranging from 7 to 8 stars (7.6 ± 0.5). No disagreements were observed between the two authors.
3.4. Classical Complement Pathway in Long COVID
The activation of the classical complement pathway was investigated in nine studies [12,16,17,18,19,20,21,22,25] with heterogeneous results. For instance, elevated C2 and C5 protein levels were reported at six [12] and twelve [17] months after infection, whereas no significant differences in C3, C4, and C5 levels were observed at one [18] and two [22] years post-infection among survivors with long COVID. Similarly, elevated levels of C4a and C4b activation products were observed in patients with long COVID compared to pre-infected controls without long COVID at one-year post-infection [17,19]. In contrast, no differences in C4d product levels between people with long COVID, non-previously infected individuals, and pre-infected controls without long COVID were identified six months after infection [12]. Wei et al. observed that subjects with long COVID showed significant dysregulation in the complement cascade including C1qA, C1qC, or C4a activation products as well as C3 and C5 components compared to patients without long COVID, but these authors did not report whether this dysregulation reflected increased or decreased complement levels [25]. Baillie et al. [18] observed increased levels of the C1s/C1-inhibitor (C1s-C1INH) complex in individuals with long COVID compared with matched (by age/ethnicity/sex/infection/vaccination status) infected controls without long COVID one year after infection; however, Hurler et al. found no differences in the C1-INH product, the C1s/C1-INH complex, or pentraxin 3 (PTX3) levels among patients with long COVID, previously infected controls without long COVID, and non-infected controls; infected participants were evaluated at ~90 days post-infection [21].
Contradictory results in the same component of the classical pathway have also been reported. Aschamn et al. observed elevated levels of C1q in long COVID [17], Baillie et al. reported lower levels of C1q [18], and Hurler et al. did not find significant differences in C1q [21] between individuals with and without long COVID. It should be noted that Aschamn et al. [17] and Baillie et al. [18] measured the C1q component levels one year after infection, whereas Hurler et al. [21] evaluated C1q at approximately three months post-infection.
Further, heterogeneity in the data reported according to specific post-COVID symptoms was noted. For instance, Cervia-Hasler et al. identified increased levels of the C2 component in patients experiencing post-COVID fatigue one year after infection [12], whereas Fernández-de-las-Peñas et al. observed decreased levels in the C3 and C5 components among survivors with post-COVID fatigue or dyspnea two years post-infection [22]. Liew et al. also observed increased levels of the C1qA activation product in individuals with gastrointestinal or cognitive post-COVID symptoms, but not in those with fatigue or cardiorespiratory symptoms [20]. Similarly, Hagiya et al. analyzed the CH50 level as a global measure of the functional activity of the classical complement pathway and reported a significant association between higher CH50 levels and post-COVID brain fog (adjusted OR 1.66, 95%CI 1.04–2.66), but not post-COVID fatigue or dyspnea, at three months post-infection [16].
3.5. Lectin Complement Pathway in Long COVID
The activation of the lectin complement pathway was investigated in three studies [12,18,23], yielding heterogeneous findings. Cervia-Hasler et al. observed no differences in MBL levels, a central component of the lectin pathway, between individuals with long COVID and infected and non-infected controls [12]. Similarly, no significant difference in the serine protease 1 (MASP1)-C1INH complex was observed by Baillie et al. [18]. On the contrary, Bulla et al. reported lower MBL levels in patients experiencing post-COVID brain fog than in long COVID survivors without brain fog and with healthy (non-infected) controls [23].
3.6. Alternative Complement Pathway in Long COVID
The activation of the alternative complement pathway was investigated in three studies [18,20,24]. Baillie et al. observed higher levels of Ba and iC3b activation products as well as increased levels of regulators Factor D and properdin, while the Factor B levels were similar in individuals with long COVID compared with infected controls without long COVID [18]. On the contrary, Song et al. showed a decrease in a specific complement component (C3f) following a first or second SARS-CoV-2 infection [24]. It should be noted that C3f is a fragment resulting from the cleavage of the C3 protein, which is crucial for all (classical, lectin, and alternative) complement pathways. Liew et al. observed increased levels of COLEC12 in people with post-COVID fatigue and cardiorespiratory symptoms [20].
3.7. Terminal Complement Pathway in Long COVID
Finally, the terminal pathway was investigated in three studies [12,18,22]. Cervia-Hasler et al. found increased levels of the C5bC6 complex and decreased levels of the C7 protein in individuals with long COVID at six months post-infection [12]. Age and vaccination status were not associated with C7 protein levels [12]. Baillie et al. observed increased levels of the C5a activation product, as well as higher levels of the C9 component and terminal complement complex (TCC) in people with long COVID as compared to infected controls without long COVID [18]. On the contrary, Fernández-de-las-Peñas et al. found no overall differences in C7 protein levels between survivors with and without long COVID at two years post-infection [22]. Patients experiencing post-COVID dyspnea showed a decrease in C7 protein levels as compared to those without post-COVID dyspnea [22].
4. Discussion
The complement system is a crucial component of the innate immune system since it serves as the first line of defense against infection and facilitates immune responses. When activated, the complement system triggers a cascade of inflammatory reactions vital for immune surveillance and homeostasis. The protective capacity of this system relies on a delicate balance between its various factors and regulators. A disturbance in this network causes significant damage to the host tissue, as it is amplified by the nonspecific, target-driven cascade and further fueled by interactions with other immune and homeostatic pathways. Multiple studies have shown complement dysregulation among COVID-19 patients as a major driver of disease pathology at the acute phase of infection [11]. While it is still unclear how complement dysregulation can be sustained for long periods of time, this was originally attributed to the viral persistence in tissue reservoirs after the acute infection [26]. Nevertheless, the presence of long-lasting viral persistence in people with long COVID has not been consistently confirmed [27,28]. Additionally, since persistent inflammation has been proposed as a mechanism of long COVID and given the role of the complement system in inflammation, the persistent chronic inflammatory state found in people with the post-COVID-19 condition could lead to the long-lasting activation of the complement system [29]. In such a scenario, elevated levels of the complement system components are indicative of acute inflammation and tissue damage, whereas decreased levels of complement system components reflect uncompensated consumption potentially associated with the prolonged and unbalanced hyperactivation of the immune system [30]. Thus, the complement system has a dual role in host autoimmune propensity since its deficiency prevents the proper clearance of debris and triggers autoimmunity whereas its overactivation causes chronic inflammation. This systematic review included eleven studies investigating the complement system in individuals with long COVID. The results suggest a dysregulated complement system in individuals with long COVID; however, the findings were heterogenous, not only in the results (some studies showed increased levels, others decreased levels, and others no differences; Table 5), but also in the follow-up periods (three, six, twelve, and twenty months), as well as in complement pathways (e.g., classic, lectin, alternative, or terminal) investigated and the diagnostic methods used.
4.1. Complement System Dysregulation in Long COVID
Based on the available evidence, the current review of the literature suggests the following: (1) several markers of complement activation spanning the classical (C2, C4a, C4b, and C1s-C1INH), alternative (Ba, iC3b, and Factor D), and terminal (C5bC6, C5a, C9, and TCC) pathways were generally elevated in patients with long COVID compared with infected subjects without long COVID, whereas other markers (C3, C4, and C4d) were not significantly different between groups; and (2) immune markers spanning the lectin complement pathway (MBL, and MASP1-C1INH) were generally not significantly different between subjects with and without long COVID.
The results on the classical pathway were highly heterogeneous since increased levels of C2 and C5 components, C4a and C4b activation products, and C1s/C1-inhibitor (C1s-C1INH) complex were found in individuals with long COVID [12,17,18,19], whereas no differences in the C3, C4, and C5 components, C4d product, C1s/C1-INH complex, or PTX3 [12,18,21,22] were identified. Contradictory results on the same (C1q) component were also found. Aschamn et al. observed elevated levels of the C1q component [17], whereas Baillie et al. reported lower levels of C1q [18] in individuals with long COVID one year after the infection. In addition, one study suggested dysregulation of the complement cascade involving C3 and C5, as well as C1qA, C1qC, and the activation product C4a, in individuals with long COVID; however, it did not report the direction of change (increased vs. decreased) for these immune markers [25]. Interestingly, reduced levels of the C1q component have been associated with autoimmune diseases such as systemic lupus erythematosus [31]. Accumulating evidence suggests that SARS-CoV-2 infection can lead to a broad spectrum of immune-mediated complications, ranging from well-defined autoimmune conditions to the development of diverse autoantibodies and atypical inflammatory phenotypes [32]. However, the heterogeneous findings observed in the current review do not allow a clear determination of whether changes in the classical complement pathway primarily reflect an increased autoimmune propensity associated with lower complement levels or, instead, reflect acute immune responses during infection associated with higher complement levels.
In relation to the alternative complement pathway, individuals with long COVID showed higher levels of Ba and iC3b activation products, and higher levels of Factor D, but similar levels of Factor B [18] when compared with infected controls without long COVID. Nevertheless, Song et al. observed that the C3f component levels decreased with consecutive infections, suggesting the progressive, uncompensated consumption of this product associated with the cleavage of the C3 component [24]. Data on the terminal component pathway were heterogeneous since increased levels of the C5bC6 complex, C5a activation product, C9 component, and TCC, but lower levels of the C7 protein were observed in people with long COVID as compared to infected controls without long COVID [12,18]. Another study investigating markers of the terminal pathway revealed no differences in C7 protein levels between individuals with and without long COVID [22].
Finally, findings for the lectin pathway were relatively consistent, with two studies reporting no between-group differences in MBL or MASP1-C1INH levels among survivors with/without long COVID [12,18]. Overall, the data suggests that different complement pathways may respond differently to SARS-CoV-2 infection.
An important observation of the current review is the heterogeneous follow-up periods of the included studies leading to a potential “time–complement pathway interaction”. The fluctuating nature of the complement system is supported by two studies. Hurler et al. observed that C1q product levels were increased during the active phase of infection (days 0 and 14), but not at days 28 or 90 post-infection [21]. Similarly, Song et al. observed a gradual decrease of the complement component C3f during the recovery period after the first and second infections [24]. Hence, it is possible that the complement system is activated during the acute phase of COVID-19 as part of the natural immune response against SARS-CoV-2, leading to increased levels of complement components [11]. In individuals with persisting symptoms, sustained or excessive complement activation may result in higher levels of complement proteins and activation products, potentially reflecting ongoing immune activation and a persistent inflammatory state involving macrophages and dendritic cells [29]. In a later, prolonged stage, sustained complement hyperactivation may ultimately result in reduced circulating complement proteins and activation products due to the ongoing, uncompensated consumption over time. Longitudinal analyses within the same cohort are needed to confirm whether complement responses fluctuate across phases of long COVID.
4.2. Complement System and Post-COVID Symptomatology
Some studies have suggested that complement system dysregulation could be symptom-specific; however, the current evidence is heterogeneous due to the assessment of multiple components or activation products from different complement pathways. For example, among survivors reporting post-COVID fatigue, C2 levels were higher at six months post-infection [12], C3 levels were lower at two years [22], and CH50 levels at three months [16], and C1qA activation products at six months [20] were not different from controls. In addition, survivors experiencing post-COVID fatigue exhibited higher levels of COLEC12 (a molecule that activates the alternative complement pathway, specifically by binding to properdin independent of C3b) at six months post-infection than those without fatigue [20]. Similarly, individuals with post-COVID dyspnea showed lower levels of C3, C5, and C7 at two years post-infection [22], but had normal CH50 levels at three months post-infection [16]. The most consistent findings were observed among individuals with post-COVID brain fog: higher CH50 levels were reported at three months [16], and increased C1qA activation products at six months [20]. The differences across studies may reflect the limited number of available studies and small sample sizes. Phenotyping post-COVID patients by symptom clusters [33] may help define more homogeneous study groups for evaluating complement dysregulation in future works.
4.3. Limitations
Although most studies support a dysregulation of the complement system in people with long COVID, the findings should be interpreted cautiously. First, the evidence is based on heterogeneous study designs, small sample sizes (n < 50) in several studies [17,22,23,24,25], and different follow-ups, which restrict the temporal inference about complement system dysregulation. Second, control groups were not uniform across studies; some comparison groups included participants who had recovered from a SARS-CoV-2 infection without developing long COVID or controls who had no prior acute infection, creating a potential selection bias. Third, no study established pre-infection baseline levels or even levels of the complement system at the acute COVID-19 phase, making it difficult to identify post-infection changes from the pre-existing inter-individual variation. Fourth, the post-COVID symptomatology assessment relied on self-report with limited harmonization, raising the possibility of misclassification and weakening links between immunologic findings and clinical phenotypes. Fifth, most studies included adult populations from different clinical settings (i.e., hospitalized, non-hospitalized, or non-specified), limiting generalizability and precluding the inference to adolescents/children. Sixth, although all studies included were considered to be of a high methodological quality, it should be considered that measurement bias is not an item included in the Newcastle–Ottawa Scale. In fact, the use of different pre-analytic handling (e.g., serum vs. plasma, processing delay, freeze–thaw, and ex vivo activation) and of multiple assessment methods (e.g., ELISA assays, mass spectrometry, and proteomics), with a different sensitivity and specificity, may explain the heterogeneity and comparability of the results.
Finally, a key limitation of the published studies is that, despite the known effects of sex, age, and obesity (BMI) on complement biology, the adjustment for these factors was inconsistent. In fact, only Baillie et al. considered sex, age, and BMI as biological confounders in their analyses [18]. Females were reported to exhibit differences in complement levels and functionality compared with males, particularly in the alternative complement pathway. Gaya da Costa et al. observed that females exhibit lower levels of the C3 component and properdin regulator, and a higher factor D concentration than males, but without differences in the classical pathway activity [34]. Only Baillie et al. included sex as a confounder factor and found no effect on the differences observed in the alternative pathway between individuals with and without long COVID [18]. It should be noted that most studies did not report sex-stratified data or consider sex as a potential biological confounder. As a result, women comprised up to 70% of long COVID participants, and meaningful sex-based comparisons were not possible with the available data. In addition, aging has been associated with increased complement pathway activity, particularly in the classical and alternative pathways, but not the lectin pathway, which may contribute to higher levels of activation products (e.g., C3a and C5a) and complement components (e.g., C3, C4, and C5) [35]. Most samples of patients included in the current review were middle-aged adults (between 40–50 years), so the effect of age would be minimal. In fact, Cervia-Hasler et al. [12] and Baillie et al. [18] did not find a significant effect of age on the complement system levels. Obesity induces a state of chronic, low-grade inflammation that activates the complement system, particularly the classical pathway, by triggering the production of C3 and C1q components [36]. Only Baillie et al. adjusted for BMI as a confounder and found that it did not affect the observed differences in the alternative pathway between individuals with and without long COVID [18]. Because sex, age, and BMI were not consistently accounted for in most studies, despite their known influence on complement biology, the effect estimates may be subject to bias.
Accordingly, these limitations preclude causal conclusions about the role of complement system dysregulation in people with long COVID and highlight the need for adequately powered longitudinal studies with the standardized phenotyping of the samples, clearly defined comparator groups, baseline measurements where feasible, and, at least, a proper matching and adjustment for sex, age, and BMI.
5. Conclusions
This systematic review discusses the current insights into the relationship between complement activation and long COVID. While the evidence suggests a potential association between long COVID and complement system dysregulation, the clinical significance remains controversial due to the heterogeneous findings. Overall, abnormalities have been more consistently reported in the classical, alternative, and terminal pathways than in the lectin pathway, although the responses appear to vary across studies and timepoints. In addition, specific post-COVID symptom clusters, such as fatigue, dyspnea, or brain fog, have been linked to a distinct pattern of complement dysregulation. The limited number of studies, methodological variability, differences in study populations and control groups, heterogeneity in long COVID symptom profiles, and the lack of pre-infection baseline complement measurements underscore the urgent need to clarify whether complement dysregulation is truly associated with long COVID and to identify the mechanisms that may underline any such association. Further studies acknowledging these gaps are now essential for tailored diagnostic and treatment approaches to enhance patient care and outcomes in the context of long COVID.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Griffin D.O. Postacute Sequelae of COVID (PASC or Long COVID): An Evidenced-Based Approach Open Forum Infect. Dis.202411 ofae 46210.1093/ofid/ofae 46239220656 PMC 11363684 · doi ↗ · pubmed ↗
- 2Natarajan A. Shetty A. Delanerolle G. Zeng Y. Zhang Y. Raymont V. Rathod S. Halabi S. Elliot K. Shi J.Q. systematic review and meta-analysis of long COVID symptoms Syst. Rev.2023128810.1186/s 13643-023-02250-037245047 PMC 10220332 · doi ↗ · pubmed ↗
- 3Fernandez-de-las-Peñas C. Notarte K.I. Macasaet R. Velasco J.V. Catahay J.A. Ver A.T. Chung W. Valera-Calero J.A. Navarro-Santana M. Persistence of post-COVID symptoms in the general population two years after SARS-Co V-2 infection: A systematic review and meta-analysis J. Infect.202488778810.1016/j.jinf.2023.12.00438101521 · doi ↗ · pubmed ↗
- 4Proal A.D. Van Elzakker M.B. Aleman S. Bach K. Boribong B.P. Buggert M. Cherry S. Chertow D.S. Davies H.E. Dupont C.L. SARS-Co V-2 reservoir in post-acute sequelae of COVID-19 (PASC)Nat. Immunol.20232416161627 Correction in Nat. Immunol. 2023, 24, 1778. https://doi.org/10.1038/s 41590-023-01646-310.1038/s 41590-023-01601-237667052 · doi ↗ · pubmed ↗
- 5Xu S.W. Ilyas I. Weng J.P. Endothelial dysfunction in COVID-19: An overview of evidence, biomarkers, mechanisms and potential therapies Acta Pharmacol. Sin.20234469570910.1038/s 41401-022-00998-036253560 PMC 9574180 · doi ↗ · pubmed ↗
- 6Notarte K.I. Carandang T.H.D.C. Velasco J.V. Pastrana A. Ver A.T. Manalo G.N. Ng J.A. Grecia S. Lippi G. Henry B.M. Autoantibodies in COVID-19 survivors with post-COVID symptoms: A systematic review Front. Immunol.202415142864510.3389/fimmu.2024.142864539035011 PMC 11257835 · doi ↗ · pubmed ↗
- 7Chen B. Julg B. Mohandas S. Bradfute S.B. RECOVER Mechanistic Pathways Task Force. Viral persistence, reactivation, and mechanisms of long COVI De Life 202312 e 8601510.7554/e Life.8601537140960 PMC 10159620 · doi ↗ · pubmed ↗
- 8West E.E. Woodruff T. Fremeaux-Bacchi V. Kemper C. Complement in human disease: Approved and up-and-coming therapeutics Lancet 202440339240510.1016/S 0140-6736(23)01524-637979593 PMC 10872502 · doi ↗ · pubmed ↗