The Complex Path from Mammary Ductal Hyperplasia to Breast Cancer: Elevated Malignancy Risk in Atypical Forms
Bogdan-Alexandru Gheban, Lavinia Patricia Mocan, Adina Bianca Boșca, Rada Teodora Suflețel, Eleonora Dronca, Mihaela Elena Jianu, Carmen Crivii, Tudor Cristian Pașcalău, Mădălin Mihai Onofrei, Andreea Moise-Crintea, Alina Simona Șovrea

TL;DR
This paper reviews how certain benign breast lesions, especially those with atypia, have a higher risk of turning into breast cancer, and explores the factors that drive this progression.
Contribution
The paper provides a comprehensive synthesis of risk factors, molecular mechanisms, and environmental influences specific to atypical ductal hyperplasia progression.
Findings
Atypical ductal hyperplasia is linked to low-grade ductal carcinoma in situ through genetic and epigenetic changes.
Environmental factors like obesity and endocrine disruptors increase progression risk via inflammation and genomic instability.
Stratified surveillance and biomarker-based interventions are recommended for high-risk hyperplasia.
Abstract
Background: Mammary ductal hyperplasia represents a spectrum of benign proliferative breast lesions, some of which pose elevated risks for malignant transformation into ductal carcinoma in situ and invasive breast cancer. This narrative review explores why only specific types, particularly those with atypia, exhibit higher progression potential, synthesizing epidemiologic, histopathologic, molecular, and environmental insights. Methods: We reviewed key literature from databases, including PubMed, focusing on classification, risk stratification, genetic/epigenetic mechanisms, tumor microenvironment dynamics, and modifiable factors influencing progression. Results: Benign breast lesions are categorized into non-proliferative, proliferative without atypia, and proliferative with atypia, such as atypical ductal hyperplasia and atypical lobular hyperplasia. Atypia represents a morphologic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
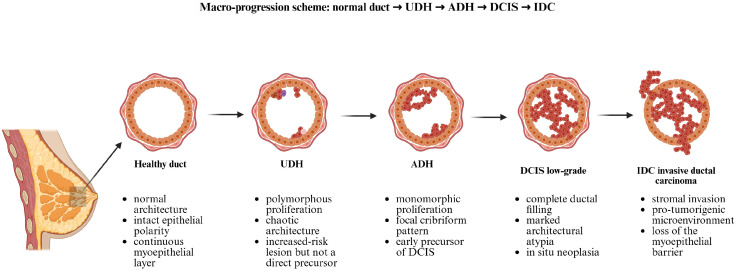 Figure 1
Figure 1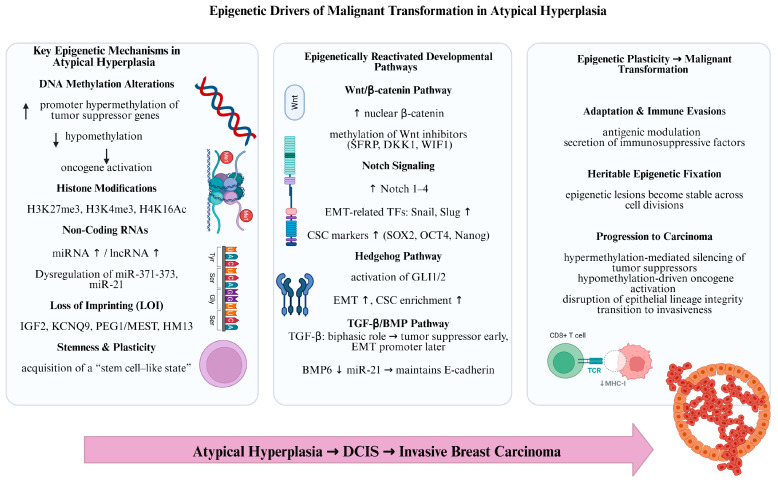 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBreast Lesions and Carcinomas · Breast Cancer Treatment Studies · Cancer Risks and Factors
1. Introduction
Mammary ductal hyperplasia (MDH) is a condition characterized by the abnormal growth of cells within the breast’s milk ducts. This condition can vary in severity, and while it is considered a benign lesion, it is often regarded as a potential precursor or risk factor for the development of breast carcinoma, primarily when associated with other pathological changes. The relationship between different types of mammary hyperplasia and the subsequent development of carcinoma is complex, influenced by both genetic and environmental factors.
Recent studies have further elucidated the progression risks associated with MDH, particularly emphasizing the role of atypical ductal hyperplasia (ADH) as a high-risk lesion. A 2023 study highlights that ADH confers a 4- to 5-fold increased risk of developing ductal carcinoma in situ (DCIS) within five years, positioning it as a marker of elevated breast cancer susceptibility rather than a direct precursor, with the risk manifesting bilaterally in the breasts [1]. A 2020 analysis indicated that the breast cancer development rate during surveillance after ADH surgery was 7.3% over a mean follow-up of 63.9 months, with palpable lesions showing significantly higher rates of progression [2]. A 2020 focused review on B3 lesions with atypia supports a low-grade progression model for ADH, noting the need for careful risk assessment to avoid overdiagnosis [3]. A 2021 review on high-risk breast lesions advocates for multidisciplinary management, including risk assessment tools and chemoprevention, to mitigate progression in atypical hyperplasia (AH) cases [4].
This narrative review synthesizes epidemiologic, histopathologic, molecular, and environmental insights to explore why only specific types of hyperplasia, particularly those with atypia, exhibit higher malignant potential. By delineating the mechanisms underlying differential progression risks, we aim to inform stratified surveillance strategies, biomarker-driven interventions, and lifestyle modifications for high-risk patients.
2. Materials and Methods
This narrative review was conducted to synthesize current evidence on the progression of MDH to breast cancer, with a focus on differential risks among hyperplasia subtypes and underlying mechanisms. The review adheres to general guidelines for narrative reviews in medical literature, emphasizing transparency in the literature selection and synthesis without formal meta-analysis or systematic appraisal tools such as PRISMA.
2.1. Literature Search Strategy
A comprehensive literature search was performed between January 2025 and November 2025. PubMed/MEDLINE served as the primary database, supplemented by manual searches of reference lists from key articles for additional relevant publications. The search strategy employed a combination of Medical Subject Headings. Key search terms included: (“ductal hyperplasia” OR “atypical ductal hyperplasia” OR “atypical hyperplasia” OR “flat epithelial atypia” OR “usual ductal hyperplasia” OR “proliferative breast disease”) AND (“breast cancer risk” OR “malignant progression” OR “precursor lesion” OR “genetic alteration” OR “PIK3CA” OR “tumor microenvironment” OR “stromal remodeling” OR “immune infiltration” OR “angiogenesis” OR “diet” OR “obesity” OR “endocrine disruptors” OR “environmental factors”). No date restrictions were imposed on the electronic search to capture foundational studies; however, priority was given to articles published after 2020 to emphasize recent advancements in molecular and environmental insights.
2.2. Inclusion and Exclusion Criteria
Studies were included if they were peer-reviewed articles published in English and provided relevant data on the histologic classification, risk stratification, genetic/epigenetic mechanisms, tumor microenvironment dynamics, dietary/obesity influences, or environmental factors related to the progression from mammary hyperplasia to carcinoma. Exclusion criteria encompassed conference abstracts, case reports, editorials, letters, non-English publications, and studies lacking direct relevance to hyperplasia progression.
2.3. Study Selection and Data Synthesis
Initial search results yielded over 220 references, which were screened for relevance by title and abstract. Full-text articles were retrieved for detailed evaluation, resulting in 209 citations ultimately included. Data were synthesized narratively, grouping findings thematically (e.g., classification, genetic drivers, microenvironment, and modifiable factors) to highlight mechanistic coherence and translational potential. Figures illustrating key concepts were generated using BioRender software under a publishing license CC BY 4.0.
3. Mammary Ductal Hyperplasia (MDH) and Its Types
Breast carcinogenesis is often framed as a multistep evolution from benign epithelial alterations toward DCIS and eventually invasive breast carcinoma. Yet in practice, most benign epithelial proliferations remain morphologically stable throughout a woman’s lifetime. Discriminating between proliferative lesions that are genuinely at risk and those that are innocuous is fundamental to guiding surveillance, biopsy decisions, and chemoprevention strategies. A proper conceptual schema subdivides benign mammary epithelial alterations into three broad categories:
- Nonproliferative changes;
- Proliferative hyperplasia without cytologic atypia;
- Proliferative hyperplasia with cytologic atypia [5].
In what follows, we review key entities in each category, summarize the epidemiologic evidence on differential cancer risk, and explore plausible mechanistic explanations for why only specific hyperplastic lesions tend to evolve.
3.1. Classification of Benign Breast Lesions and Associated Risks
3.1.1. Nonproliferative Changes
Nonproliferative lesions are those lacking significant epithelial proliferation. Representative lesions include simple cysts, ductal ectasia, apocrine metaplasia, mild epithelial hyperplasia without proliferation, and stromal fibrosis [6]. In large cohorts, women whose benign biopsy reveals only nonproliferative changes do not display a significantly increased risk of subsequent breast cancer relative to the background population. Indeed, nonproliferative disease is used as the baseline reference in risk stratification in many studies [7]. In select analyses, marginal associations between specific nonproliferative features (e.g., microcalcifications, mild epithelial change) and later cancer risk have been reported. However, effect sizes are modest and often attenuate with multivariable adjustment. In the classic Hartmann et al. cohort of 9087 women with benign breast disease, nonproliferative lesions had a relative risk (RR) for breast cancer of 1.27 (95% CI 1.15–1.41) compared to the general population, which is lower than in proliferative disease without atypia (RR ~ 1.88) or AH (RR ~ 4.24) [3].
A recent systematic review emphasized that the breast cancer risk associated with benign breast disease is heterogeneous, primarily determined by the extent of epithelial proliferation and the presence or absence of cytologic atypia. While benign breast disease in general correlates with an elevated incidence of breast cancer compared to women without such findings, the magnitude of this increase is substantially lower for nonproliferative lesions than for either proliferative lesions without atypia or AH [8]. Consequently, nonproliferative disease represents the lowest-risk category within the benign breast disease spectrum.
Thus, the consensus is that nonproliferative lesions are low-risk and do not demand intensive follow-up beyond standard screening, unless modified by other risk factors (e.g., family history, dense breasts, reproductive and hormonal factors, or genetic predisposition) [9,10].
3.1.2. Proliferative Hyperplasia Without Atypia
This category encompasses lesions with increased epithelial cell numbers but without definite cytologic atypia. Key entities include: usual ductal hyperplasia (UDH), sclerosing adenosis, radial scars, complex sclerosing lesions, intraductal papillomas without atypia, complex proliferative lesions, or multifocal proliferative foci.
Epidemiologic evidence indicates that proliferative lesions without atypia confer a modest increase in breast cancer risk, typically in the range of 1.5- to 2.0-fold compared to nonproliferative disease. For example, in a large case–control study nested within benign breast disease cohorts, proliferative lesions without atypia had an adjusted odds ratio (OR) of approximately 1.45 relative to nonproliferative disease. [11]. Other classic studies similarly estimate a RR of 1.8–1.9 [3].
With a focus solely on certain lesional entities, UDH is characterized by a haphazard proliferation of heterogeneous epithelial cells forming slit-like spaces, bridging projections, and irregular architecture. Because of its disorganized morphology and lack of monoclonal dominance, UDH is generally considered a marker of increased risk rather than a direct precursor lesion. [3,12]. Currently, no reliable prognostic indicators exist to determine which patients with UDH will progress to invasive breast carcinoma. The associated risk level is comparable to that conferred by specific reproductive characteristics, such as early onset of menarche or delayed menopause. It therefore does not justify the modification of routine mammographic screening intervals [13].
Radial scars and complex sclerosing lesions often harbor adjacent proliferative changes, and some series report modest independent risk contributions or occasional occult carcinoma [14]. Although once considered potential precursors, current evidence does not confirm a premalignant nature for radial scars. Their modestly increased breast cancer risk (RR of approximately 1.4–1.7) likely reflects coexisting proliferative changes rather than intrinsic malignancy. Larger lesions (>0.6 cm), more common in women over 50, may coexist with AH or carcinoma [15]. Optimal management of mammographically detected radial scars and complex sclerosing lesions remains debated. Lesions showing epithelial atypia on core biopsy carry a malignancy risk warranting excision [16,17]. In contrast, recent studies suggest that lesions with concordant imaging and pathologic aspects lacking atypia might be managed conservatively [18,19,20].
To sum up, proliferative disease without atypia constitutes an intermediate-risk stratum but, by itself, lacks many of the hallmarks of progression to carcinoma, such as architectural disorder or clonal expansion.
3.1.3. Proliferative Hyperplasia with Atypia
This is the high-risk category, containing principally ADH and atypical lobular hyperplasia (ALH), and, in some classification schemes, flat epithelial atypia (FEA). These lesions combine proliferation with cytologic uniformity, partial loss of polarity, and architectural disturbance.
AH is identified in 4 to 10% of benign breast biopsies performed for imaging or palpable lesions [5,9]. Multiple long-term follow-up cohorts and meta-analyses demonstrate that proliferative hyperplasia with atypia confers a 3- to 5-fold RR of future breast carcinoma compared to nonproliferative disease [3,21,22,23]. In the Mayo cohort, the estimated cumulative incidence of breast cancer at 25 years in women with AH was approximately 29% [9]. Recent large-scale analyses using contemporary imaging and biopsy data refined these estimates. In the Breast Cancer Surveillance Consortium cohort, the 10-year incidence of invasive carcinoma after ADH was approximately 5 to 6%, slightly lower than previous figures, with risk modestly higher in excisional than in core needle biopsies (6.7% vs. 5%) [24]. These findings suggest that smaller ADH foci, increasingly detected by modern imaging, may carry a lower absolute risk than in earlier cohorts. However, ADH remains a significant marker of long-term susceptibility [25].
ALH is the lobular counterpart of ADH, arising in the terminal duct-lobular units. It is characterized by a monomorphic proliferation of discohesive, dyshesive cells (often due to loss of E-cadherin expression) filling part of the lobule. ALH and the related LCIS are collectively referred to as lobular neoplasia, which confers a risk of subsequent carcinoma similar to that of ADH [25].
The often-quoted approximately 1% annual risk for AH is a useful heuristic, though actual risk is modulated by age, family history, breast density, and other lesion features [25].
3.1.4. Subtype Nuances: FEA, ADH vs. ALH, and the Extent of Atypia
FEA represents clonal proliferation of mildly atypical epithelial cells lining terminal duct lobular units, typically associated with microcalcifications and incidentally discovered during mammographic screening [26]. Although limited data exist, evidence indicates that FEA may occasionally precede invasive breast carcinoma [27]. Nonetheless, the probability of progression is very low, and its risk magnitude is considerably lower than that of ADH or ALH [28]. Consequently, FEA should not be managed identically to these higher-risk atypical proliferations.
Several retrospective analyses have reported that up to one-third of FEA lesions identified on core needle biopsy may be more advanced at the time of surgical excision. However, these findings are inconsistent due to small sample sizes, variable histologic criteria, and differences in radiologic–pathologic correlation, resulting in wide variability in reported upgrade rates. Current evidence suggests that surgical excision is not uniformly required after a diagnosis of pure FEA, particularly when post-biopsy imaging confirms complete removal of the calcifications associated with the lesion. Instead, careful radiologic–pathologic concordance is advised to determine appropriate follow-up and to ensure that management remains individualized and evidence-based [29,30].
ADH is frequently conceptualized as a direct, though non-obligate precursor of low-grade DCIS, whereas ALH is considered a risk indicator or a marker of a susceptible field [31]. In practice, both types are associated with subsequent ductal carcinoma, and their clinical outcomes are often similar [27].
The question of whether the extent (number of foci) of atypia modulates risk has been explored. In the nested case–control study by Collins et al., among women with ADH, increasing the number of foci (≥3) did not significantly increase OR beyond that of 1–2 foci (OR 2.7 vs. 3.5, p = 0.58) [32]. In ALH, there was a trend toward higher OR with ≥3 foci (OR 8.0) vs. 1–2 foci (OR 5.2), though this was not statistically robust (p = 0.66). Other series suggest some stratification by extent, but the evidence is not consistent enough to uniformly guide management [33]. In another systematic review of determinants of breast cancer risk in women with atypia, use of menopausal hormone therapy, timing of follow-up, and other factors were also modifiers of risk [8].
3.1.5. Upgrade Rates and Interpretive Caution
A recurring complication is that many cases of AH diagnoses on core-needle biopsy are upgraded to DCIS or invasive carcinoma on subsequent excisional pathology. This upgrade phenomenon clouds the boundary between actual progression versus initial under-sampling of coexistent malignancy [21]. Meta-analyses of core-biopsy-diagnosed ADH yield upgrade rates to malignancy in the range of 10–30%, depending on biopsy technique, lesion characteristics, and institutional practice [34]. Disentangling upgrades from genuine evolution remains an ongoing challenge, requiring rigorous pathological and imaging correlation and thoughtful clinical decision-making.
Therefore, the histologic classification of benign breast lesions remains highly relevant to stratified surveillance. For nonproliferative disease, standard screening per population guidelines is generally appropriate. In proliferative diseases without atypia, a modest risk elevation may justify closer surveillance in the context of additional risk factors. On the other hand, because AH confers substantial long-term risk, many centers recommend enhanced surveillance (annual mammography plus supplemental MRI) and consideration of risk-reducing interventions. Excision of the atypical lesion does not eliminate the residual elevated risk conferred by the breast environment; therefore, continued vigilance is required [22,35].
3.2. Mechanistic Frameworks Underlying the Progression of Breast Lesions
Although the above classification and epidemiologic data stratify risk, they do not explain why only particular lesions (especially those with atypia) tend to evolve toward malignancy. Below, we synthesize current mechanistic hypotheses, enriched by recent molecular studies, and present them graphically in Figure 1.
3.2.1. Degree of Atypia
The presence of cytologic and architectural atypia is perhaps the single most important histologic feature distinguishing benign proliferations that merely signify elevated risk from those that embody early steps in the neoplastic continuum. Within AH, the severity of cytologic atypia may correlate with proximity to malignant features (e.g., higher nuclear grade, more mitotic activity, loss of polarity). Lesions with more pronounced atypia may be closer to the threshold and thus more likely to cross into carcinoma, especially with additional alterations [36].
3.2.2. Morphologic and Histopathologic Continuum: ADH as a “Mini-DCIS”
ADH is generally regarded as a transitional lesion within the biological spectrum of breast epithelial proliferations, representing an intermediate step between UDH and low-grade DCIS. One of the most compelling arguments that atypia represents a transitional state is the striking morphologic and structural affinity between ADH and low-grade DCIS. In fact, many pathologists conceptually regard ADH as a limited, partial, or incipient DCIS, fulfilling many of the same architectural and cytologic criteria, but lacking the extent or completeness of ductal involvement [37].
Microscopically, ADH is characterized by a relatively uniform (monomorphic) epithelial population showing mild to moderate nuclear enlargement and minimal pleomorphism. The architectural arrangement often mirrors that of low-grade DCIS, displaying cribriform, micropapillary, or solid patterns; however, these features are typically focal, involving only a limited number of ductal spaces. A partial loss of polarity and reorganization of epithelial layers may also be observed, features that distinguish ADH from the irregular, haphazard cell stratification seen in UDH. From an immunophenotypic standpoint, ADH shares several markers with low-grade DCIS [38]. The atypical cells commonly express estrogen and progesterone receptors and demonstrate a relatively low proliferative index (Ki-67), though this is higher than that observed in non-atypical proliferations. Molecular studies further reveal overlapping genetic and epigenetic alterations between ADH and DCIS, reinforcing the notion that ADH represents an early, quantitatively limited precursor in the neoplastic progression toward DCIS [39].
Because ADH clones possess many (though not all) features of low-grade DCIS, they are anatomically and biologically poised for further progression. Indeed, the threshold from ADH to low-grade DCIS is sometimes subtle and somewhat arbitrary (extent, degree of involvement, continuity) in diagnostic practice [21,40]. Thus, from a morphologic standpoint, the presence of atypia pushes a lesion from the benign proliferative region toward the in situ neoplastic region on the histologic continuum.
3.2.3. Epidemiologic and Clinical Evidence: Progression Risk Linked to Atypia
From the population and cohort studies, the differential risk conferred by AH (vs. proliferative disease without atypia) is pronounced and consistent. Whereas non-atypical proliferative disease confers a RR of approximately 1.5 to 2, AH confers approximately 4–5× risk of future carcinoma [9]. Moreover, long-term follow-up data support that AH is a strong independent predictor of later DCIS and invasive carcinoma, even after adjustment for confounders (age, family history, breast density) [9]. This suggests that atypia is not merely a marker but is likely part of the causal pathway.
Observational cohorts of ADH (particularly “pure ADH” without concurrent lesions) show meaningful rates of later carcinoma, consistent with a subset of these lesions evolving over many years [21]. Additionally, in analyses of biopsy upgrade rates (concurrent, initially occult carcinoma), ADH diagnoses on core biopsy are upgraded to DCIS or invasive cancer in a variable proportion (10–30%) depending on biopsy technique, lesion features, and institutional practice. This suggests that in many cases, ADH is a “step on the ladder” adjacent to more advanced neoplasia [21].
In conclusion, atypia is a powerful discriminator of risk and a clinically actionable histologic marker of progression potential.
3.2.4. Microenvironmental and Epithelial Constraints: How Atypia May Loosen Barriers
The molecular architecture of atypical lesions (discussed below) suggests a “halfway house”, offering sufficient neoplastic alteration to confer a selective advantage but not yet enough to cross the barrier into invasion. Therefore, while molecular alterations are necessary, they may be insufficient for progression without concomitant loosening of architectural or microenvironmental constraints.
The progression from benign epithelial proliferation to neoplasia also depends on changes within the cellular microenvironment that relax usual growth constraints. Atypical epithelial proliferation may represent a stage at which these regulatory barriers begin to fail, allowing for architectural disorganization and early invasive potential. As epithelial cells acquire atypical features, the structural integrity and functional role of the myoepithelial layer and basement membrane may become compromised. These barriers, normally responsible for maintaining ductal polarity and inhibiting stromal invasion, can appear attenuated, fragmented, or locally absent, diminishing their suppressive influence on epithelial expansion [41].
In parallel, reciprocal communication between the epithelial and stromal compartments is altered. Atypical cells may produce or respond abnormally to growth factors, cytokines, and matrix metalloproteinases (MMPs) that remodel the extracellular matrix, increase matrix stiffness, and recruit fibroblasts and immune cells, collectively creating a pro-tumorigenic milieu [42]. Within these altered microenvironments, the spatial organization of cells and local mechanical or metabolic gradients may further relax normal controls such as contact inhibition and nutrient diffusion, favoring the outgrowth of more resilient subclones [43].
Additionally, atypical epithelial populations often display increased resistance to apoptosis and enhanced survival under oxidative or replicative stress, enabling their persistence in settings where non-atypical cells would undergo programmed cell death [44]. Overall, atypia may therefore represent not only a morphological manifestation of genetic instability but also a biologically active phase in which epithelial and stromal interactions evolve to permit escape from architectural suppression—effectively bridging the transition from hyperplasia to neoplasia. As such, atypia may not only reflect accumulated mutations but also actively mediate escape from architectural suppression—providing a bridge from hyperplasia to neoplasia. Once a lesion surpasses the threshold of DCIS, the transition to invasion shares many overlapping features and constraints, and the presence of atypia earlier can be seen as a prelude.
Among untreated DCIS, 25–60% are believed to progress to invasive carcinoma over years to decades, depending on grade, molecular subtype, and microenvironment [45]. However, not all DCIS progress to invasive disease. Microenvironmental suppression, immune surveillance, and stromal constraints continue to play a role. In that respect, atypia marks the early loosening of barriers, but further events are required to progress to full invasion [46]. Thus, atypia marks the entry point into the neoplastic continuum, with molecular/architectural permissiveness setting the stage for subsequent progression into full in situ and invasive disease [47].
In AH, reciprocal interactions between epithelial and stromal compartments play a pivotal role in fostering a pro-tumorigenic microenvironment. Atypical epithelial cells secrete or aberrantly respond to growth factors, cytokines, and MMPs, which activate stromal fibroblasts into cancer-associated fibroblasts (CAFs) [47]. These CAFs, identifiable by α-SMA expression on immunohistochemistry, remodel the extracellular matrix by increasing collagen deposition and stiffness, thereby disrupting basement membrane integrity and promoting epithelial–mesenchymal transition (EMT)-like changes [48]. This bidirectional crosstalk also recruits immune cells, such as regulatory T cells and macrophages, which suppress anti-tumor immunity and enhance hypoxia-induced angiogenesis via vascular endothelial growth factor (VEGF). Consequently, these alterations relax normal growth constraints, such as contact inhibition, and create metabolic gradients that favor the clonal expansion of resilient subpopulations, bridging the progression from hyperplasia to neoplasia [21,49].
In conclusion, not all atypical lesions are destined to progress. The molecular and morphological heterogeneity within AH means that some lesions may remain indolent, others may regress, and only a subset evolves. The presence of atypia is necessary but not sufficient; additional factors (extent, molecular burden, microenvironment, host context) also influence the trajectory.
Given the importance of atypia, the pressing translational challenge is to differentiate which atypical lesions will progress from those that will remain stable. Biomarkers such as gene-expression risk scores, mutational burden, spatial heterogeneity metrics, and imaging correlates are being explored [49,50].
3.2.5. Specific Molecular Mechanisms from Hyperplasia to Neoplasia
The progression from MDH to breast cancer entails molecular alterations disrupting homeostasis, fostering genomic instability, and enabling transformation, with variations by subtype risk. Low-risk subtypes, like UDH, show minimal, reversible changes without clonal dominance. Genetic changes are rare, including sporadic somatic mutations or copy-number variations such as 1p/8p losses, and are not associated with broad instability [21]. Epigenetic modifications, such as minor promoter hypermethylation of suppressors, fail to drive progression [49]. The tumor microenvironment (TME) maintains homeostasis, with scant stromal remodeling, minimal immune infiltration, and hypoxia-induced angiogenesis, rendering low-risk lesions mere risk markers with a <2-fold RR compared to the general population [21]. High-risk subtypes, such as ADH and ALH, manifest amplified hallmarks bridging benignity and malignancy via low-grade pathways. Drivers include recurrent PIK3CA mutations, activating PI3K-AKT-mTOR for survival/proliferation. CNVs feature prominent 16q loss and 1q gains, mirroring DCIS and ER+ invasive ductal carcinoma. Epigenetic changes amplify miR-21 (inhibiting apoptosis) and hypermethylation of APC/TWIST1, boosting EMT [45]. The TME promotes progression through CAFs secreting hepatocyte growth factor, regulatory T-cell immune suppression, hypoxia-driven VEGF angiogenesis, and MMPs’ extracellular matrix degradation. These explain the 3- to 5-fold RR, facilitating clonal expansion and senescence escape, while enabling PIK3CA-targeted therapies [45].
4. Genetic Causes and Mechanisms of Mammary Gland Hyperplasia: From Atypia to Malignant Transformation
The likelihood that a given hyperplastic lesion will progress to cancer correlates with the accumulation of specific genetic mutations and epigenetic modifications. Recent research has revealed how these molecular events, from driver gene mutations (e.g., in PIK3CA, BRCA1/2, TP53) to DNA methylation changes and microRNA dysregulation, can cooperate to drive some hyperplasias along the trajectory to malignancy [51,52].
As we have seen, UDH is a proliferation of ordinary ductal epithelium without atypia and is not considered an actual tumor. Generally, it is polyclonal. Molecular analysis shows that UDH lesions typically lack the consistent clonal genetic alterations seen in atypical lesions [48]. Nevertheless, modern sequencing has detected low-frequency activating mutations in pro-growth pathways, even in some UDH cases: PIK3CA mutations or alterations in the PI3K-AKT-mTOR pathway [53]. These early mutations may provide a growth advantage but by themselves are insufficient for malignancy, as UDH generally retains normal architecture and growth control.
From a molecular point of view, ADH is a clonal neoplastic proliferation and is considered a genetically advanced precursor lesion [48]. ADH frequently harbors driver mutations commonly seen in luminal-type breast cancers, such as PIK3CA mutations (the catalytic subunit of PI3K), which are present in a large fraction of ADH lesions [45]. Copy number aberrations characteristic of low-grade carcinomas are also observed; classic examples include gains on 1q and losses on 16q. Loss of heterozygosity on chromosome 16q, which harbors the CDH1 tumor suppressor gene, has been detected in some ADH cases. 16q loss is a hallmark of the low-grade pathway and is also frequent in concurrent DCIS or invasive cancers of the luminal A subtype. Overall, the genetic profile of ADH often mirrors that of low-grade DCIS, supporting its role as an immediate precursor [48]. Table 1 summarizes key genetic alterations in ADH and other hyperplasias, while Table 2 provides a more detailed view of key genetic drivers from AH to cancer.
Genetically, lobular neoplasia (ADH and ALH) is defined by inactivation of the CDH1 gene, which encodes E-cadherin. E-cadherin loss is observed in more than 90% of ALH/LCIS, either through mutations in the CDH1 gene or 16q22 loss of heterozygosity [45,48]. This loss abrogates cell–cell adhesion and is a defining early step in the lobular lineage of breast cancer development [48]. In addition to CDH1, lobular neoplastic lesions often harbor mutations in PI3K pathway genes similar to those in ADH. For example, PIK3CA mutations are found in a substantial subset of ALH/LCIS and invasive lobular carcinomas [48]. Recurrent mutations in the AKT1 gene (another PI3K pathway oncogene) have also been reported in lobular cancers and may be present even at the in situ stage. Remarkably, lobular neoplasia can be multifocal and bilateral; genetically, independent ALH/LCIS foci in the same breast may arise from a field effect of genetically altered but morphologically normal cells distributed through the lobe. The presence of early BRCA2 or TBX3 gene mutations in morphologically normal epithelium has been documented in some cases [48]. This field of altered lobules can give rise to clonally related neoplastic lesions along both ductal and lobular pathways (if additional ductal or lobular lineage-specific hits occur) [48].
While genetic mutations lay the foundation for neoplastic transformation, epigenetic modifications add an additional layer of dysregulation in MDH. Epigenetic changes, heritable alterations in gene expression that do not stem from DNA sequence mutations, are now understood to occur early in breast lesion development [49]. These include DNA CpG Island methylation, post-translational histone modifications, and deregulated expression of non-coding RNAs such as microRNAs. In hyperplastic breast tissue, epigenetic alterations can silence tumor suppressor pathways or activate oncogenic programs, thereby complementing genetic drivers in promoting cell proliferation and abnormal survival. Importantly, epigenetic changes are reversible in principle, and their presence in precursor lesions raises the possibility of early intervention to halt or reverse progression (though therapeutic implications are beyond the scope of this review) [68].
Aberrant DNA methylation is one of the earliest and most consistent epigenetic abnormalities observed during breast carcinogenesis. Even at the stage of hyperplasia, especially in AHs, specific genes acquire promoter hypermethylation, leading to transcriptional silencing. For example, the promoters of tumor suppressor genes such as RASSF1A and APC are frequently hypermethylated in pre-invasive breast lesions, including ADH and DCIS. RASSF1A is a cell cycle regulator frequently silenced in breast cancer, and studies have detected RASSF1A methylation in a significant fraction of AHs and carcinoma in situ. Similarly, TWIST1, HIN1, p16INK4a, and BRCA1 are among other genes reported to undergo methylation-associated silencing early in the progression sequence [69]. In lobular neoplasia, in addition to global methylation changes, CDH1 promoter methylation can serve as a second hit in cases lacking a CDH1 mutation; in fact, CDH1 promoter hypermethylation is found in a subset of LCIS/ILC, contributing to E-cadherin loss [59]. Overall, the presence of DNA methylation changes in a hyperplastic lesion is a red flag for a biologically advanced state.
MicroRNAs (miRNAs) are 20–22 nucleotide non-coding RNAs that post-transcriptionally regulate sets of target genes. Altered miRNA expression profiles are well documented in breast cancers, and emerging evidence indicates that miRNA dysregulation starts at pre-cancerous stages [70]. In fact, certain miRNA changes in hyperplasias may signal the development of malignancy and serve as early biomarkers. It was noted that oncomiRs such as miR-21 and miR-155, which promote proliferation and inhibit apoptosis, are progressively upregulated from normal epithelium to ADH to DCIS [69]. These miRNAs are key regulators of breast tumorigenesis: for instance, miR-21 suppresses PTEN and other tumor suppressors, while miR-155 modulates TP53 and SOCS1, thereby driving cell growth. On the other hand, some tumor-suppressor miRNAs are found to be downregulated in precursors. A striking recent example is miR-1297, which is frequently underexpressed in FEA lesions. Scafetta et al. compared miRNA profiles of normal breast epithelium, FEA, and DCIS; miR-1297 was significantly downregulated in FEA and DCIS compared with normal breast epithelium [49]. MiR-1297 normally targets the oncogenic receptor Ephrin-A2 (EPHA2), so its loss may unleash pro-proliferative signaling. Functional experiments confirmed that restoring miR-1297 in breast cells inhibited growth, whereas loss of miR-1297 enhanced proliferation and altered 3D acinar morphology. Thus, miR-1297 downregulation represents an early event in mammary neoplastic transformation, potentially driving the progression of FEA toward malignancy [49]. This illustrates how even minute non-obligate lesions can harbor significant regulatory changes.
4.1. Mechanism of Malignant Transformation
The process by which ADH transitions into carcinoma involves the accumulation of multiple genetic mutations in key pathways governing cellular growth, survival, and apoptosis [71]. These mutations enable hyperplastic cells to evade normal regulatory controls, fostering an escalating potential for invasive behavior. This lesion bridges benign proliferations and DCIS, with features such as cribriform patterns, micropapillae, and uniform nuclear enlargement, all of which are discernible under microscopy [72]. The existing literature indicates that malignant transformation is not obligatory but is often driven by molecular aberrations, which can manifest as synchronous or metachronous carcinomas [25]. For instance, upgrades from ADH to DCIS or IDC occur in 22% to 65% of cases at excisional biopsy, highlighting histopathological progression marked by increased atypia and loss of myoepithelial integrity [73].
Additionally, the hypothetical multistep model of breast carcinogenesis proposes progression from normal epithelium to invasive carcinoma through intraductal hyperplasia (IDH), with and without atypia, including ADH and DCIS, and cytogenetic and molecular-genetic analyses reveal accumulation of genetic alterations [50]. Comparative genomic hybridization (CGH) and fluorescence in situ hybridization (FISH) studies reveal DNA amplification in the chromosomal region 20q13 in early IDH stages, indicating early cytogenetic changes in presumptive precursors [74].
4.1.1. Gene Expression Changes
The onset of atypia in ADH is frequently associated with alterations in gene expression of oncogenes (e.g., HER2/ERBB2, Cyclin D1) and tumor suppressor genes (e.g., TP53, BRCA1/2), which histopathologically manifest as proliferative and architectural abnormalities. Transcriptional profiling of matched normal, ADH, and carcinoma samples reveals upregulation of ERBB2, FOXA1, and GATA3 in ADH, correlating with estrogen receptor positivity and luminal phenotypes observable on immunohistochemistry (IHC) [75]. These changes are evident in H-E (hematoxylin-eosin) sections as monotonous cell populations with nuclear enlargement and rare mitoses, distinguishing ADH from UDH by uniform spacing and polarization. In synchronous ADH, gene expression patterns cluster with those of low-grade DCIS, exhibiting enriched pathways in membrane transport, fatty acid metabolism, and phenylalanine metabolism, which may contribute to the cribriform architecture and micropapillary formations observed microscopically [76].
Matrix metalloproteinase-1 (MMP-1) emerges as a potential progression biomarker, with elevated expression in synchronous versus pure ADH, aligning with basement membrane discontinuities visible on periodic acid-Schiff (PAS) staining [25]. Furthermore, hub gene analyses identify RRM2, TOP2A, PBK, MELK, and NUSAP1 as progressively upregulated from normal mammary epithelium to ADH, DCIS, and IDC, which is associated with cell cycle deregulation and proliferation. Histopathologically, this correlates with increased Ki-67 positivity in atypical cells, indicating heightened mitotic activity in transitioning lesions [77]. In ADH, including both ductal and lobular variants, gene signatures exhibit shared alterations, such as downregulation of immune-related genes and upregulation of proliferation markers, which are evident in multifocal lesions characterized by expanded ducts and acini on H-E [50]. ERBB2 overexpression, even without amplification, is noted in ADH, with IHC revealing moderate cytoplasmic and membranous staining in atypical epithelial cells, potentially driving the transition to HER2-positive IDC as seen in 7% of subsequent carcinomas [78].
These expression shifts are more pronounced in premenopausal women, in whom higher-grade features, such as punctate necrosis, predict malignant progression, underscoring hormonal influences on histopathological progression [79]. Cyclin D1 gene amplification and protein overexpression occur in benign breast disease and ADH, with frequencies similar to normal tissue in non-atypical hyperplasias (amplification 15–19%, overexpression 13%) but higher in ADH (amplification 27%, overexpression 57%), approaching DCIS (35% amplification, 50% overexpression) and IDC levels (25% amplification, 64% overexpression). Assessed via differential Polymerase Chain Reaction (PCR) and IHC, these changes precede histologic alterations but increase in atypical lesions [80]. BigH3 protein expression decreases progressively from benign tissues to DCIS, lobular carcinoma, and IDC, with benign tissue showing a 23-fold increase compared to infiltrating colloid carcinoma, correlating with malignancy in tissue microarray IHC of 192 cases [81].
In vitro hypoxia models show increased Hypoxia-Inducible Factor-1 alpha (HIF-1α), Glucose transporter 1 (GLUT1), and Carbonic Anhydrase IX (CAIX) expression, with in vivo IHC absent in normal, DH, or ADH but present in DCIS (GLUT1 56.8%, CAIX 25.0%) and IDC (GLUT1 44.1%, CAIX 30.5%), higher in high-grade lesions (p = 0.001 for GLUT1, p = 0.036 for CAIX in DCIS). Notch1 and JAG1 hypomethylation inversely correlate with protein overexpression in IDC (Notch1 88.7%, JAG1 89.9%) versus ADH (36.0%, 45.0%), and are associated with lymph node metastasis and tumor, node, metastasis (TNM) stage [82]. In postmenopausal macaques, estrogen (E2) increases proliferation, epithelial area, and progesterone receptor expression (p < 0.05), with greater columnar cell hyperplasia in E2-treated groups (p < 0.05) and greater ESR1 and Ki67 in lesional tissue. MiR-205-5p downregulation in metastatic 21T series correlates with higher histopathological grades (EG III) and invasion rates [83]. HER-2 amplification and overexpression are absent in ADH. Still, they are characteristically present in high-grade DCIS, implicating a contributory role in the clonal proliferation underlying neoplastic progression [84].
4.1.2. Loss of Tumor Suppressor Genes
The inactivation of genes such as PTEN, p16, and RB often leads to uncontrolled cell division and contributes to the progression of malignancy, as evidenced by histopathological findings including loss of cellular polarity and increased nuclear pleomorphism. Loss of heterozygosity (LOH) at tumor suppressor loci is an early event in ADH, targeting regions such as 16q (harboring CDH1/E-cadherin) and 17p (TP53), and is observed in 42% of pure ADH cases via allelic imbalance analysis [85]. This manifests histopathologically as disrupted myoepithelial layers, with reduced p63 and smooth muscle actin (SMA) expression on IHC, facilitating epithelial-stromal interactions in atypical ducts. In paired UDH and ADH samples, shared LOH at 11p15.5, 13q14, 16q24.3, and 17p13.1 suggests a continuum, with ADH showing additional losses that correlate with cribriform patterns and partial ductal involvement [86]. RB1 and BRCA1/2 regions are implicated, with 16q deletions being common in low-grade pathways, leading to E-cadherin loss and lobular-like features in some ADH lesions, which are visible as expanded acini with uniform cells on H-E [87]. PTEN inactivation, though less frequent in pure ADH, accumulates in synchronous cases and is associated with nuclear atypia and mitoses that border on DCIS criteria [88]. Histopathological grading reveals that subsequent carcinomas from ADH exhibit 31% grade 1, 43–53% grade 2, and 16–26% grade 3, with suppressor losses contributing to this spectrum rather than exclusively low-grade disease. p16 alterations, often epigenetic, are noted in AH, correlating with heterogeneous CK5/6 negativity on IHC, which distinguishes it from UDH’s mosaic pattern and indicates suppressor-mediated dedifferentiation [89].
In high-risk families, ADH prevalence reaches 39%, with BRCA1/2 mutations enhancing suppressor loss, as seen in prophylactic mastectomy specimens with multifocal atypia and microcalcifications [90]. These inactivations drive morphological shifts, such as the formation of rigid bridges and solid growth, underscoring their role in malignant evolution. Fragile Histidine Triad gene (FHIT) and WW domain–containing oxidoreductase (WWOX) expressions decrease from normal to ADH, DCIS, and IDC, with higher detectable rates in benign tissue/ADH (rate ratios 2.95–4.58 for mRNA/protein) versus in situ/invasive stages [91]. Accumulation of chromosomal imbalances has been documented in premalignant breast lesions, including recurrent gains on chromosomes 3p and 8q, as well as losses on 16q, in ADH, which may implicate tumor suppressor genes in early neoplastic changes [92]. LOH at 11q13 has been observed in approximately 9% of ADH cases, 0% of low-grade DCIS, and 35% of high-grade DCIS, indicating a potential role for this alteration in facilitating the transition to invasive carcinoma. In intraductal hyperplasia, mutant p53 and cyclin D1 overexpression are more frequently identified in DCIS than in precursor lesions, with LOH at multiple loci associating suppressor gene inactivation with disease progression [93].
4.1.3. Genetic Instability
Atypical lesions tend to harbor genomic instability, including chromosomal aberrations and mutations in critical loci, particularly those involved in DNA repair and cell cycle regulation (e.g., BRCA1 and TP53 mutations), which are histopathologically evident as aneuploidy and clonal expansions [94]. Aneuploidy is detected in 15–44% of ADH by flow cytometry and in all cases via Fluorescence In Situ Hybridization (FISH) with multiple probes, higher than in non-atypical lesions but lower than in carcinomas. Comparative Genomic Hybridization (CGH) identifies copy number aberrations such as 16q loss and 1q gain in pure ADH, mirroring DCIS/IDC, with histopathological correlates including ductal spaces filled with monotonous cells and pseudo-lumens [95]. Next-generation sequencing in small cohorts reveals shared aneuploidy (e.g., 1q gain) and somatic mutations, but with intra-lesional heterogeneity in 46% of multifocal ADH, manifesting as mixed clonal proliferations on H-E. TP53 mutations accumulate in synchronous ADH, associating with nuclear pleomorphism and mitoses, while BRCA1 defects enhance instability, as seen in high-risk lesions with frequent LOH [52].
Centrosomal abnormalities, with increased α- and γ-tubulin expression on IHC, are observed in ADH and carcinoma, indicating mitotic spindle defects that correlate with aneuploidy and invasive foci. Molecular profiles reveal advanced changes, including gross chromosomal rearrangements and epigenetic alterations in AH, with histopathological features such as calcifications and multifocality predicting greater instability and upgrade risk [96]. In progression models, instability drives diploid ADH to aneuploid high-grade breast carcinoma, though most ADH-associated cancers are ER-positive and moderate-grade, suggesting a low-grade pathway with occasional escalation. Allelic imbalance frequencies are <5% in benign tissue and ADH, 20% in DCIS, 25% in invasive carcinomas, with significant differences between ADH and DCIS (p < 0.0001), suggesting instability predetermines biology at the in situ stage [97].
Genetic alterations are absent in fibroadenomas, even associated with cancer, but LOH in DCIS within fibroadenomas, indicating no role in carcinogenesis [98]. c-erbB-2 amplification is absent in hyperplasia and ADH, but present in high-grade DCIS (10–40% in IDC), highlighting instability in the precursor to invasive transition [99].
4.1.4. Microenvironmental Factors
The surrounding tissue microenvironment, including stromal changes and inflammatory responses, can promote the progression of atypical lesions into invasive cancer, with histopathological evidence of field cancerization and multifocality [100]. Ipsilateral predominance of subsequent breast carcinoma (2:1 ratio) suggests a microenvironmental field effect, persisting post-excision, as observed in cohorts with 80% ipsilateral events in the first 5 years [31].
Stromal alterations, such as desmoplasia and calcification, are common in ADH, as revealed by H-E staining, which shows periductal fibrosis and immune infiltrates that promote progression [101]. Lobular involution extent inversely correlates with risk, with absent involution (SIR 7.66) associating with denser stroma and atypical expansions [102]. Inflammatory responses, including downregulation of immune genes in AH, contribute to evasion, as evidenced histopathologically by reduced lymphocytic aggregates in progressing lesions [103].
Microenvironmental hormones, like in premenopausal women, heighten the risk of higher-grade carcinomas, with features like punctate necrosis on core biopsy predicting upgrade [104]. Field cancerization explains the multifocal appearance of ADH (higher risk if calcified), with non-clonal origins in cancer-prone tissue, as seen in heterogeneous lesions separated by stroma [105]. Tamoxifen’s risk-reduction benefits support modulation of the microenvironment, with histopathological reversal of atypia in treated cases [106].
Overall, these factors, along with genetic changes, drive transformation, advocating for microenvironment-targeted interventions. Extra-tumoral tissue in breast carcinoma patients shows ductal hyperplasia effects similar to non-neoplastic tissue under oral contraceptive use, indicating stromal-inflammatory influences [107]. In progression, hypoxia shifts HIF-1α’s role from a proliferative response in early stages (DH, ADH) to tumor promotion, with GLUT1/CAIX in DCIS/IDC adapting to hypoxia/acidosis to promote aggressive phenotypes [108]. E2 in macaques induces columnar cell changes/hyperplasia, with greater Estrogen Receptor-A1 (ESR1)/Progesterone Receptor (PGR)/Ki67 in lesions, suggesting a hyper response in terminal ductal lobular units [109].
5. Epigenetic Modifications in Atypical Hyperplasia and Their Role in Malignant Transformation
5.1. General Features
Epigenetic reprogramming plays a key role in mammary tumorigenesis, favoring the progression from AH to invasive carcinoma by altering gene expression, promoting a stem cell-like state, and enabling EMT [110]. The main epigenetic mechanisms comprise alterations in DNA methylation, histone modifications, non-coding RNA expression, loss of genomic imprinting, and reactivation of developmental pathways such as Wnt/β-catenin and Notch signaling, which could disrupt tumor suppressor genes and promote oncogenes [111]. Another important epigenetic mechanism is cell plasticity and stemness [112]. Thus, epigenetic reprogramming may confer stem cell-like properties to tumor cells, enabling limitless self-renewal and supporting tumor heterogeneity. Still, these identical flexible epigenetic changes offer therapeutic opportunities, as preventing epigenetic converters may reestablish normal gene expression, making tumors responsive to therapy and improving the efficiency of immunotherapies [113]. Figure 2 offers an overview of the main epigenetic changes in AH.
5.2. The Main Epigenetic Mechanisms Involved in Breast Tumorigenesis
5.2.1. Loss of Imprinting
It was found that genomic imprinting has a pivotal role in development and evolution. Loss of imprinting (LOI) represents an initial phase alteration in neoplasia. Imprinting is defined as the monoallelic expression of genes in a parent-of-origin-specific approach [114]. It was shown that in diploid eukaryotes, the maternal and paternal copies of major genes are expressed at equal levels. Concerning imprinted genes, nonetheless, just one allele is transcriptionally dynamic [52]. Imprinting models are variable among tissues [115]. Most imprinted genes are regulated by imprinting control regions, which are generally modulated by DNA methylation, yet H3K27me3 also contributes to this regulation [116,117]. Moreover, as the number of imprinted genes is fundamental, disruption of imprinting is associated with some human imprinting syndromes and could lead to cancer by promoting oncogenic or suppressing antitumor processes [118,119].
LOI develops in a biallelic expression from the stimulation of the dormant allele. Research in mice revealed that demethylation of imprinted genes leading to LOI made cells more sensitive to cellular transformation and carcinogenesis [52]. Initiation of a growth-promoting allele leads to aberrant cell multiplication, a trigger of cancer [120].
LOI represents an essential abnormality in breast neoplasia growth and evolution, concerning the aberrant expression of imprinted genes linked to epigenetic dysregulation, usually DNA hypomethylation [31]. Reports have discovered imprinted genes such as HM13, KCNQ9, IGF2 and PEG1/MEST that display LOI in breast cancer, directing to modified gene expression responsible for tumor progression, with possible diagnostic and therapeutic values [31,100].
Research highlighted that HM13 shows LOI and expression upregulation in mammary tumors, involving DNA demethylation and possibly related to better prognosis in different types of cancer [121].
LOI at the KCNQ9 locus has been involved in the overexpression of the TASK3 potassium channel and is linked to triple-negative breast cancer [122]. The connection between KCNQ9 hypomethylation and triple-negative breast cancers indicates that targeting the KCNK9 DMR may represent a probable approach for inhibiting this form of breast cancer [102].
Persistent LOI of PEG1/MEST has been detected in invasive breast carcinomas, indicating its role in the evolution from hyperplasia to invasive cancer [31]. LOI of the IGF2 gene could determine its overexpression, supporting the growth of tumor-initiating cells and overall chromatin variability [123].
5.2.2. Reactivation of Developmental Pathways
Several of the best-described signaling pathways influencing self-renewal and differentiation in mature stem cells, like Wnt/β-catenin, Notch, Hedgehog and TGF-β/BMP pathways are commonly regulated in cancer via epigenetic processes [91].
Abnormal epigenetic changes in ADH could restart developmental paths, for example Notch signaling and Wnt/β-catenin signaling, that are usually suppressed in mature cells but are reactivated through cancer development [91,93].
Notch signaling controls cell multiplication, differentiation and influences cell outcome and programmed cell death [93]. Notch signaling is controlled via Notch receptors (1–4) and ligands for instance Delta-like ligand (DDL 1/3/4) and Jagged 1/2 [124]. Notch signaling collaborates with transcription factors of EMT (Snail and Slug) and stemness (SOX2, Nanog, and OCT4), in addition to accelerating the acquisition of both EMT and stemness [125]. Notch, in combination with TGF-β, induces Slug expression to modulate EMT. Furthermore, Notch signaling controls distinct target genes associated with cancer stem cells (CSCs) and facilitates chemoresistance [126]. For example, IL-6, a Notch target gene, sustains the self-renewal capacity of CSCs, and Notch-regulated activation of PKB protects cells from programmed cell death, thereby promoting survival [106]. Inactivation of Notch signaling destroys precursor cells that are like CSCs, which promote the function of Notch in stemness and correlate with chemoresistance. MDR1 is highly expressed in CSCs, during which the Notch signal is linked to NF-κB and related PI3K/Akt activation, thereby controlling MRP2, a carrier that facilitates stemness maintenance [91].
Wnt signaling pathway, which regulates the expression of target genes via β-catenin, represents a key mechanism of stem cells and CSCs and is abnormally activated through the growth of several human cancers [93]. Acquisition of function mutations of the CTNNB1 gene (encoding β-catenin) and deficit-of-function mutations of AXIN and other genes were observed to be the principal processes responsible for Wnt signaling alteration in cancers [127]. Several genes implicated in the Wnt/β-catenin signaling pathway are methylated and suppressed in mammary gland cancer, including the Wnt inhibitors SFRP1-5, WIF1, and DKK1, as well as the SRY-box containing gene 17 (SOX17) and APC [107]. Current findings indicate that the Wnt/β-catenin pathway may also be regulated by histone mutations in cancer [93]. Overexpression of EZH2 in the mammary gland leads to nuclear accumulation of β-catenin and activation of the Wnt pathway, as well as intraductal epithelial hyperplasia [128]. Moreover, the transcriptional suppression of the Wnt antagonist DACT3 was associated with histone modifications at H3K4me3 and H3K27me3 [129]. Additionally, Dkk-1-mediated suppression of expression was caused by reduced H4K16Ac and increased H3K27me3, plus the recruitment of Suz12, SirT1, EZH2, and BMI1 to its promoter [130]. Also, miRNAs play a key role in modulating various components of the Wnt/β-catenin pathway. It was shown that β-catenin/Lef1 transactivates the miR-371-373 group associated with CSCs self-renewal, and, in turn, these miRNAs control the Wnt/β-catenin signaling by directing DKK1 expression [131].
Hedgehog (Hh) signaling modulates the proliferation of stem and precursor cells in several tissues, and alterations in this pathway have been implicated in tumor growth [93,132]. Hh signaling pathway cooperates with Wnt-β-catenin signaling, inducing tumorigenesis and cancer severity [133]. This mechanism could regulate both CSC and EMT pathways by increasing key transcription factors (Slug, ZEB1, Snail, ZEB2, FOXC2, and TWIST2) and stem cell markers (CD44, BM1, and CD133) [113]. Enhancement of abnormal Hh signaling mechanisms in destructive, metastatic, and chemoresistant mammary gland cancer types leads to reduced E-cadherin expression and increased expression of Twist, FOXC2, SIP1, Snail, vimentin, N-cadherin, and fibronectin [105].
BMP/TGF-β signaling pathways control numerous biological processes like proliferation, differentiation, and programmed cell death [91]. Deregulating the molecular effectors of TGF-β signaling could lead to cancer [93]. TGF-β functions as a tumor suppressor in tumor induction. The initial phases of tumor development and the deactivation of the TGF-β tumor suppressor pathway represent the principal phase in the growth of many tumors [134]. Still, in the final phase, it produces tumor progression, EMT, and metastasis [93]. BMP-6 acts as a suppressor of mammary cancer EMT by saving E-cadherin expression. Recent findings suggest that this mechanism is facilitated by BMP-6-stimulated transcriptional suppression of miR-21, which is overexpressed in destructive mammary gland neoplasia [135].
5.3. Epigenetic Plasticity
Epigenetic plasticity enables cells to dynamically alter gene expression patterns in response to the evolving TME, facilitating adaptation and survival under stressful conditions such as hypoxia or nutrient scarcity. This adaptability also allows cancer cells to circumvent intrinsic cellular checkpoints that typically restrict aberrant proliferation, including apoptosis and cell cycle arrest mechanisms [136]. Furthermore, it promotes immune evasion, enabling tumor cells to persist and thrive by modulating surface antigens or secreting immunosuppressive factors. The underlying processes involve key epigenetic modifications, such as histone acetylation or methylation, aberrant DNA methylation at promoter regions, and dysregulation of noncoding RNAs, which collectively enhance this phenotypic flexibility during early neoplastic stages [137].
These epigenetic alterations can become heritably fixed through successive cell divisions, stabilizing a malignant phenotype that supports sustained tumor growth. In the context of breast cancer, such changes drive progression from premalignant lesions like ADH by repressing tumor suppressor genes (e.g., via hypermethylation) or activating oncogenes, thereby disrupting lineage integrity and promoting invasive characteristics [138]. Environmental factors, including dietary influences or endocrine disruptors, can further amplify these epigenetic shifts, underscoring the interplay between extrinsic cues and intrinsic molecular reprogramming in cancer evolution [139].
Table 3 highlights some known epigenetic alterations in different hyperplasia subtypes, while Table 4 provides more details on the molecular and cellular effects.
6. Tumor Microenvironment Changes in Driving Malignant Transformation of Atypical Hyperplasia
The TME in breast ADH encompasses a multifaceted, evolving landscape comprising neoplastic epithelial cells, stromal fibroblasts, immune cells, vascular networks, and extracellular matrix (ECM) components. These components interact synergistically to modulate the progression from premalignant states, such as ADH, to DCIS and ultimately to IDC. Subtle TME modifications in ADH, such as early stromal fibrosis, immune cell recruitment, and alterations in the basement membrane, are detectable through IHC and serve as harbingers of malignant progression. The existing literature highlights that these TME shifts, including disruption of the myoepithelial layer and activation of fibroblasts, are pivotal in facilitating invasion. For instance, reduced myoepithelial continuity in ADH lesions, as assessed by the co-expression of alpha-smooth muscle actin (α-SMA) and p63 via multiplex IHC, correlates with an increased risk of progression to DCIS. Moreover, gene expression profiling and spatial analyses reveal transcriptional reprogramming in the stroma that supports epithelial invasion. This review synthesizes histopathological evidence from recent studies, highlighting how TME dynamics drive the malignant transformation of ADH, with implications for biomarker development and targeted interventions to halt disease progression.
6.1. Key TME Components and Their Impact on Malignant Transformation
6.1.1. Stromal Remodeling
Remodeling in ADH involves the activation and phenotypic shift in fibroblasts, leading to desmoplasia and a supportive niche for epithelial proliferation and invasion. Histopathologically, early ADH exhibits periductal fibrosis with an increased density of α-SMA-positive CAFs, which are observable on IHC as spindle-shaped cells encircling atypical ducts [145]. These CAFs secrete cytokines and growth factors, such as hepatocyte growth factor (HGF), which upregulate the MET receptor in epithelial cells, fostering morphological changes indicative of invasive potential. In coculture models mimicking ADH progression, premalignant cells like MCF10DCIS display upregulated HGF/MET signaling when interacting with mammary fibroblasts, resulting in altered acinar structures in three-dimensional (3D) cultures, characterized by delayed cavitation and increased apoptosis upon HGF blockade [146].
This stromal-epithelial crosstalk is evident in vivo, where high HGF expression in the stromal compartment correlates with basal-like subtypes and poorer survival, highlighting stromal contributions to ADH malignant transformation. Furthermore, myoepithelial cells in ADH undergo phenotypic alterations, as indicated by IHC, which reveals reduced continuity—measured as increased distances between α-SMA/p63 co-positive cells—compared to normal tissue. This discontinuity, significantly lower in ADH bordering DCIS, permits greater epithelial-stromal interactions, potentially allowing immune cell access and promoting progression [41]. In obesity-associated models, high-fat diet (HFD)-induced stromal changes accelerate ADH formation, with H-E sections showing expanded atypical ducts and DCIS-like lesions; weight loss reverses this by reprogramming the kinome, downregulating PKC-α and MEK3 while upregulating AMPKα, as detected by proteomic arrays in unaffected glands [147]. Additionally, inhibitor of differentiation-1 (Id-1) overexpression in ADH, assessed by IHC, correlates with morphologic progression, driving shifts in carcinoembryonic antigen-related cell adhesion molecule 1 (CEACAM1) from apical to cytoplasmic/membranous patterns, disrupting cell adhesion and facilitating stromal invasion [148]. Stromal fibroblasts in high-grade ADH-adjacent areas lose CD34 expression while gaining α-SMA, as shown by IHC, and inversely correlate with microvessel density (MVD), indicating a desmoplastic response that stiffens the matrix and aids epithelial migration [149]. These histopathological features underscore stromal remodeling as a critical driver, and early interventions targeting CAFs may arrest ADH progression.
6.1.2. Immune Cell Infiltration
The immune landscape in ADH’s TME features progressive infiltration of lymphocytes and macrophages, which can either suppress or promote malignant transformation depending on their phenotype and spatial distribution [150]. Histopathologically, H-E staining of ADH biopsies reveals periductal lymphocytic aggregates, with higher stromal lymphocyte counts predicting upgrade to DCIS or IDC. In B3 lesions, including ADH, elevated stromal lymphocytes, quantified per international guidelines, combine with patient age and lesion type to yield a predictive model for upgrade, suggesting immune surveillance failure in progressing cases [151]. Multiplex IHC further delineates this, showing increased tumor-infiltrating lymphocytes (TILs) in TP53-mutated ADH, with CD3+CD8+ T cells associated with favorable outcomes and CD3+Foxp3+ regulatory T cells (Tregs) linked to recurrence. Spatial proximity analyses via IHC demonstrate that closer T cell-tumor cell interactions correlate with reduced invasive risk, implying immune-mediated containment in non-progressing ADH [41].
In hypoxic conditions, myeloid cells release S100A9, which fosters systemic immunosuppression, as evidenced by altered CD4+/CD8+ ratios in lymphoid organs and reduced T cell proliferation in cocultures [152]. Histologically, ADH progressing to DCIS shows enhanced stromal immune infiltrates, including macrophages, on H&E, contributing to ECM degradation via protease secretion [153]. In basal-like precursors, immune evasion is marked by increased polymorphonuclear neutrophils (PMNs) in the stroma, as detected by Ly6G+ IHC, which inhibit T cell function and promote atypical hyperplasia [130]. These findings are compounded in obesity, where HFD elevates inflammatory cytokines, leading to denser immune infiltrates in ADH lesions, which are reversible with weight loss, restoring lean phenotypes and reducing pre-neoplastic inflammation. Overall, histopathological evidence points to a shift from anti-tumor to pro-tumor immune profiles in progressing ADH, with TIL density and subtype serving as prognostic indicators [154].
6.1.3. Hypoxia and Angiogenesis
Hypoxic niches in expanding ADH lesions trigger angiogenesis, providing sustenance for proliferation and invasion. Histopathologically, central necrosis in atypical ducts on HE signals hypoxia, with IHC showing nuclear hypoxia-inducible factor-1α (HIF-1α) stabilization in epithelial and stromal cells. This induces vascular endothelial growth factor (VEGF) expression, which correlates with elevated MVD as measured by CD31 IHC, particularly in high-grade ADH transitioning to DCIS [155]. Dynamic contrast-enhanced MRI (DCE-MRI) quantifies this angiogenic switch, revealing increased vessel permeability and a higher vascular fraction in atypical hyperplasia, which persist through progression and are confirmed by microvessel counting [156]. In transgenic models, hypoxia-driven small extracellular vesicles (sEVs) containing HIF-1α promote angiogenesis, as evidenced by CD31 staining showing increased microvessel density and in vivo probes confirming vascular proliferation [133].
Angiogenesis precedes significant stromagenesis in ADH, as evidenced by VEGF-positive epithelial cells and immature vessels on IHC, fostering a permissive environment for invasion.
In obesity contexts, HFD exacerbates hypoxic signaling, accelerating angiogenic changes in ADH, while weight loss mitigates this via kinase modulation, reducing vascular support for progression [157]. HGF/MET pathways also contribute, with upregulated signaling in ADH, leading to tortuous vessels observable in 3D models. These histopathological markers highlight hypoxia and angiogenesis as early drivers, with therapeutic targeting of HIF-1α or VEGF potentially preventing ADH malignant evolution.
6.1.4. Extracellular Matrix (ECM) and Cell–Cell Interactions
ECM remodeling in ADH disrupts the structural integrity of the epithelium, enabling epithelial invasion. Histopathologically, Masson’s trichrome staining reveals disorganized collagen in periductal stroma, with IHC showing elevated MMPs from activated fibroblasts degrading basement membranes. Basement membrane discontinuities, visible on PAS staining, coincide with myoepithelial loss, marked by reduced α-SMA/p63 expression, facilitating EMT-like changes with vimentin positivity in atypical cells. RNA sequencing of ADH tissues identifies upregulated ECM-related genes, correlating with increased colony formation and migration in Matrigel assays, indicative of invasive potential [158].
Cell–cell adhesions weaken, with IHC demonstrating E-cadherin loss and CEACAM1 redistribution from apical to cytoplasmic patterns in progressing ADH (p < 0.05), driven by Id-1 overexpression. In 3D cultures, HGF/MET activation induces morphological shifts, such as epithelial budding and ECM invasion, reversible by signaling inhibition [159]. Obesity amplifies ECM stiffening via kinase upregulation, promoting the ADH-to-DCIS transition, while weight loss restores matrix homeostasis. These changes, including enhanced integrin-mediated signaling, underscore ECM and adhesion disruptions as key histopathological features of ADH transformation [160].
6.1.5. Exosomes and Tumor Signaling
Exosomes in ADH’s TME mediate intercellular signaling, transferring oncogenic cargo to remodel the niche. Although direct visualization is challenging, indirect evidence from IHC indicates exosome-driven alterations, such as nuclear accumulation of HIF-1α in recipient cells [129]. Hypoxic sEVs from premalignant cells induce AH, as evidenced by H-E staining, which reveals ductal expansion and increased Ki-67+ proliferation. These vesicles reprogram progenitors, disrupting the polarity of cytokeratin 8/14 and promoting EMT via RNA-seq-identified pathways [161]. In progression models, exosomal HIF-1α accelerates oncogene-driven tumorigenesis, and plasma HIF-1α levels correlate with recurrence in luminal cancers. Stromal responses, including α-SMA upregulation, suggest exosome-mediated CAF activation, though limited histopathological data exists [144]. Targeting exosomal cargo may offer novel strategies to interrupt ADH signaling networks [162].
In conclusion, the histopathological evolution of the TME in ADH—encompassing stromal activation, immune shifts, hypoxic angiogenesis, ECM degradation, and exosomal communication—critically drives malignant transformation. Integrated analyses from IHC, imaging, and molecular profiling provide robust evidence for these mechanisms, advocating for TME-targeted biomarkers and therapies to mitigate progression risks in high-risk patients.
7. Diet and Environmental Factors Influencing the Transformation of Atypical Ductal Hyperplasia (ADH) into Breast Cancer
7.1. Dietary Factors Influencing ADH Progression
According to the American Cancer Society, about 30% of the breast cancers in postmenopausal patients are related to modifiable risk factors, including diet [163]. Moreover, some research suggests that a healthy diet could prevent breast cancer onset, improve therapeutic outcome and decrease the risk of recurrence among these patients [164].
High-fat diet is an important risk factor for the onset and progression of breast cancer, through multiple pathogenetic mechanisms, involving increased systemic inflammation and oxidative stress [165]. Increased intake of total and saturated animal fat, found in red meat and high-fat dairy, was linked to the risk of breast cancer due to activation of pro-inflammatory mechanisms such as the NF-kB signaling, with downstream expression of cytokines, growth factors, and other molecules capable of supporting tumor progression [159,166]. The association between intake of monounsaturated fatty acids and breast cancer appears to depend on the source (olive oil or margarines) and the food processing methods. Intake of olive oil could have a beneficial effect by reducing insulin resistance, whereas hydrogenated oleic acids found in margarines seem to increase breast cancer risk. Polyunsaturated fatty acids can exert different effects on breast cancer, depending on the position of the double bond [167]. Thus, ω-6 fatty acids, such as linoleic acid and arachidonic acid, may promote mammary tumorigenesis by producing eicosanoids and by undergoing oxidation, which results in enhanced cell damage [168]. By contrast, marine ω-3 fatty acids seem to have a protective effect [146].
A high-fat diet can disrupt the gut microbiota, increasing the risk of malignancies, by the activation of the “gut-bone marrow-tumor” axis. Altered microbiota can produce large amounts of leucine, capable of activating the mTORC1 signaling in myeloid progenitor cells, resulting in differentiation of polymorphonuclear myeloid-derived suppressor cells (PMN-MDSCs). An increased number of PMN-MDSCs infiltrating the tumor promotes breast cancer growth and metastasis [169].
High-fat diets may promote tumor cell metastasis by preactivating platelets and endothelial cells and by forming premetastatic niches. Within niches, the crosstalk between activated platelets, endothelial cells and cancer cells promotes metastasis [170].
Additionally, high-fat diets are a major driver of obesity, which itself increases breast cancer risk and progression through complex signaling pathways affecting cell growth and angiogenesis [171].
Obesity is a high-risk factor for breast cancer and poor prognosis in women after menopause [172]. Moreover, the risk of triple-negative breast cancer and mortality post diagnosis is significantly increased in both pre- and postmenopausal obese women, compared with normal-weight women [173].
Potential pathogenic mechanisms underlying these associations include: increased estrogen levels, dysregulated insulin and Insulin-like growth factor (IGF) signaling, altered adipokines released by adipose cells, and chronic systemic inflammation indicated by high levels of C-reactive protein [41,174].
In postmenopausal women, the link between obesity and high estrogen levels is more significant, since, after menopause, the adipose tissue is the main source of estrogen. Excess estrogen derived from androgen aromatization in the adipose tissue, associated with obesity-related decrease in sex hormone binding globulin causes a hormonal imbalance, which is linked to the high risk of breast cancer [175,176]. A meta-analysis reported that obese or overweight breast cancer patients develop larger and higher-grade malignancy tumors, more frequent positive lymph nodes and distant metastasis, lower survival rate and worse overall prognostic [177]. This could be explained by the estrogen’s ability to fuel the growth of hyperplastic cells and increase the risk of DNA mutations [178].
Imbalance in adipokines due to increased mass of adipose tissue is also responsible for direct activation of breast cancer cells, contributing to tumor growth and invasion. Low adiponectin levels and high leptin levels were linked to poor prognosis of breast cancer. Adiponectin has a protective role by inhibiting cancer growth [179]. High leptin levels secreted by excess adipose tissue activate multiple mechanisms, including Jak2/Stat3, MAPK, PI3K-AKT, thus increasing tumor cell proliferation and invasion [180].
Chronic systemic inflammation maintains a pro-inflammatory state in adipose tissue, leading to increased secretion of inflammatory cytokines that promote tumor progression and metastasis. In obese individuals, adipose tissue is infiltrated by numerous macrophages, which form crown-like structures surrounding degenerating adipocytes. The presence of such structures in the mammary adipose stroma has been associated with increased levels of blood glucose, insulin, triglycerides, C-reactive protein and IL-6 [181]. Moreover, these patients had a higher risk of developing breast cancer, tumor metastasis and a lower survival rate [182,183]. The mechanisms underlying the poor prognosis in obese patients in association with the accumulation of macrophages and crown-like structures in the mammary gland include: the activation of pathways such as: NF-κB, hypoxia-inducible factor 1-alpha (HIF-1α), and PI3K-AKT-mTOR.
NF-κB, expressed in human breast cancer tumor cells, supports cancer cells’ survival and differentiation into an aggressive phenotype, and is also responsible for the ET resistance in ER+ breast tumors [184]. Expression of HIF-1α in adipose tissue macrophages has been detected in patients with insulin resistance and altered glucose metabolism [185].
PI3K-AKT-mTOR pathway controls cell growth and survival and has been demonstrated to cross-talk with estrogen receptor (ER) pathway; thus, alterations of PI3K-AKT-mTOR pathway are present in ER+ breast cancer [186].
Insulin resistance, which often accompanies obesity, leads to elevated levels of insulin and IGF, known as potent mitogens. Insulin and IGF-related tumorigenesis occurs by activation of oncogenic pathways such as PI3K/Ras-Raf in the affected cells, resulting in cell proliferation and ADH progression [187].
According to data published by the World Cancer Research Fund, a higher risk of breast cancer is linked to alcohol consumption, even in low amounts. Pathogenic mechanisms underlying the association between ethanol intake and mammary tumorigenesis include: oxidative stress by accumulation of reactive oxygen species (ROS), systemic inflammation due to increased production of pro-inflammatory cytokines, and insulin resistance [188]. Breast cancer initiation in patients who consume ethanol is the consequence of hormone level alterations, mainly increasing estrogen by activation of aromatase [189]. Thus, alcohol was found to promote the development of ER+ breast tumors, but not ER-cancers. Additionally, alcohol is converted into acetaldehyde, a carcinogenic metabolite that causes DNA damage and induces epigenetic modifications by abnormal DNA methylation, both of which contribute to cancer initiation and progression [165].
Reducing high-fat foods, especially processed meats, fried goods, and high-fat dairy, and replacing them with unsaturated fats, fruits, vegetables, and the correct supplementation with vitamins and minerals is a key lifestyle recommendation for breast cancer prevention and outcome management [142].
Phytoestrogens are plant-derived compounds found in various foods: fruits, vegetables, grains, legumes such as soybeans, flaxseeds, tofu, and lentils [190]. Due to their molecular structure, these compounds perform a wide range of pharmacologic effects, acting as antioxidants, anti-inflammatories, anti-tumoral agents, and can also be effective as endocrine modulators. Upon binding to the estrogen receptors, phytoestrogens can mimic or regulate the function of estrogen in the body [154]. Phenolic phytoestrogens contain two main groups of active pharmacologic substances: flavonoids (including flavone, isoflavone, flavonol phytoestrogens, dihydroflavone phytoestrogens) and non-flavonoid compounds (mainly terpenoids) [191]. Some studies suggest that phytoestrogens might have a protective role against breast cancer by acting as antagonists to estrogen in the breast tissue, potentially reducing the risk of malignancy in estrogen-sensitive cancers [154]. This protective role is explained by the ability of phenolic compounds to bind to the estrogen receptor and to prevent the interaction of endogenous estrogen with the G protein-coupled estrogen receptor (GPER) [192]. Moreover, the hydroxyl group in xenoestrogens plays a role in performing estrogen-like effects [193]. However, the effect of phytoestrogens on the progression of ADH is still debated, as some studies suggest that they may stimulate estrogen receptors in certain contexts, promoting cell proliferation in hormonally driven breast tumors.
Vitamins can also have anti-cancer properties. Studies suggest that vitamin D may inhibit tumor cell proliferation in mammary hyperplastic lesions, whereas vitamins C, E, and carotenoids exert antioxidant properties [194]. Dietary fibers are beneficial for breast cancer prevention by reducing glucose absorption and glycemic levels, and by regulating the balance of gastrointestinal microbiome [195].
7.2. Environmental Factors and Toxins Influencing ADH Progression
Although ADH already reflects a significant factor for the malignant transformation, other environmental and toxic factors such as endocrine disruptor chemicals, air pollution, pesticides, radiation exposure, etc., may have an important role in how ADH lesions evolve [196].
7.2.1. Ionizing Radiations
One of the most powerful environmental factors influencing ADH Progression is ionizing radiation, which directly induces double-stranded DNA breaks and also, in mammary epithelial cells, chromosomal instability. Studies show that even low doses of radiation can cause the activation of DNA repair and cell cycle genes (e.g., BRCA1). This leads to a proliferative effect in breast tissue [197].
Studies in mice have suggested that radiation exposure causes the activation of genes that are involved in proliferative and metabolic pathways that play a role in carcinogenesis in the mammary glands [198].
7.2.2. Chemical Endocrine Disruptors
Endocrine disruptors are natural or synthetic substances that are able to affect the physiological functions of the endocrine system [199]. These chemicals (EDCs) represent a class of anthropogenic compounds (e.g., bisphenol A, phthalates, and perfluoroalkyl substances) that mimic or antagonize natural hormones. Low-dose exposure to BPA and DDT significantly increases aromatase (CYP19A1) biosynthesis and 17β-estradiol production in mammary epithelial cells, leading to sustained activation and proliferation of Erα [200].
Bisphenol A (BPA) is used to produce polycarbonate plastics (e.g., plastic bottles), and recent studies discovered that exposure to BPA, due to the ability to mimic estrogen, can increase the growth of cancer cells in the breast [201].
7.2.3. Air Pollution
Airborne pollutants and particulate matter (PM) contain a multitude of compounds such as polycyclic aromatic hydrocarbons (PAHs) and metals which can stimulate the progression of mammary cancer [202].
Exposure to PM2.5 can lead to oxidative damage to the DNA and can also increase the expression level of oxidation markers. Cellular responses to these factors are activation of transcription factors, release of inflammatory mediators, etc., which can lead to cellular apoptosis. Studies revealed that urban residents have higher PM2.5 exposure and present a higher risk of developing breast cancer [203].
Polycyclic aromatic hydrocarbons (PAHs) are ubiquitous in the environment and are formed in the organic fuel combustion process. Studies have correlated PAH exposure with development of breast cancer [204].
7.2.4. Pesticides
Pesticides such as organophosphates, organochlorines and fungicides have been proven to increase the risk of breast cancer [205]. Organophosphates, a class of chemicals used as pesticides, are proven to act as endocrine disruptors and can increase the risk of mammary tumor development [206].
Dichlorodiphenyltrichloroethane (DDT), a synthetic organochlorine, is found to be in the adipose tissue and studies revealed that higher levels of DDT was found in women with breast cancer compared to women without cancer and a lower DDT level [207].
7.2.5. General Indications
Lifestyle modifications like weight control and limiting environmental toxin exposure vary in effectiveness for reducing progression from atypical ductal hyperplasia (ADH) to breast cancer in specific populations. In postmenopausal women at higher risk from adipose-derived estrogen, weight management via calorie restriction and exercise lowers incidence by 20–50% [208]. A Mediterranean diet, featuring antioxidants from fruits, vegetables, and omega-3s, adds 10–20% to risk mitigation by curbing inflammation and insulin resistance compared with high-fat diets. For ADH/ALH patients under 45, restricting alcohol to under one drink daily effectively cuts estrogen-driven ER+ proliferation, reducing risk by 10–20%. Vulnerable groups exposed to endocrine disruptors (e.g., BPA, pesticides) gain from avoidance like BPA-free items and organics, potentially lessening genomic instability, though evidence is mainly epidemiological. Tailored combinations yield the greatest benefits in postmenopausal obese and younger high-risk individuals [209].
8. Conclusions
This review has examined the complex spectrum of MDH and its potential role as a precursor to breast cancer, highlighting why certain subtypes, especially those with atypia, pose a higher risk of becoming malignant. By combining epidemiologic, histopathologic, molecular, and environmental data, we have emphasized the multifactorial nature of progression from benign proliferative lesions to DCIS and invasive cancer. Key findings show that benign breast lesions are categorized into nonproliferative changes (low risk), proliferative hyperplasia without atypia (moderate risk), and proliferative hyperplasia with atypia (high risk). Atypical ductal hyperplasia and ALH serve as crucial transitional states, both morphologically and genetically bridging benignity and malignancy. These atypical forms display early cancer-related changes, such as PIK3CA mutations, BRCA1/2 dysfunction, and epigenetic modifications, including miR-21 upregulation, that disturb cellular balance and promote genomic instability. The tumor microenvironment further increases this risk through stromal remodeling, immune dysregulation, hypoxia-driven angiogenesis, and ECM breakdown via MMPs. Modifiable factors worsen these processes: obesity and high-fat diets sustain chronic inflammation through adipokines such as leptin, while environmental exposures to ionizing radiation, endocrine disruptors, air pollutants, and pesticides cause DNA damage and hormonal imbalance, primarily affecting high-risk hyperplasia.
Stratified risk assessments enable targeted monitoring: women with atypical hyperplasia might benefit from increased mammographic screening, chemoprevention, or preventive procedures, while women with non-atypical lesions need routine surveillance. Lifestyle changes, such as weight management and reduced toxin exposure, can help prevent progression. Biomarker-based strategies, including molecular testing for PIK3CA or TP53, could tailor treatment and identify those who may benefit from early intervention. Environmental links, while strong, often rely on epidemiological links rather than direct mechanistic proof. Future research should focus on long-term cohort studies that integrate omics data to understand progression pathways better. Ultimately, this review advocates for a comprehensive approach to MDH management, combining genetic risk, environmental factors, and personalized prevention strategies to lower breast cancer rates.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Miceli R. Mercado C.L. Hernandez O. Chhor C. Active Surveillance for Atypical Ductal Hyperplasia and Ductal Carcinoma In Situ J. Breast Imaging 2023539641510.1093/jbi/wbad 02638416903 · doi ↗ · pubmed ↗
- 2Yoon J.H. Koo J.S. Lee H.S. Kim E.K. Kim M.J. Park V.Y. Moon H.J. Factors Predicting Breast Cancer Development in Women During Surveillance After Surgery for Atypical Ductal Hyperplasia of the Breast: Analysis of Clinical, Radiologic, and Histopathologic Features Ann. Surg. Oncol.2020273614362210.1245/s 10434-020-08476-932314161 · doi ↗ · pubmed ↗
- 3Ugurlu M.U. Yoldemir T. Gulluoglu B.M. Assessment and management of B 3 breast lesions with atypia: A focused review Climacteric 202023172310.1080/13697137.2019.166063731566023 · doi ↗ · pubmed ↗
- 4Bahl M. Management of High-Risk Breast Lesions Radiol. Clin. N. Am.202159294010.1016/j.rcl.2020.08.00533222998 · doi ↗ · pubmed ↗
- 5Kumar V. Abbas A.K. Aster J.C. Robbins and Cotran Pathologic Basis of Disease 10th ed.Elsevier Philadelphia, PA, USA 2020
- 6Klassen C.L. Hines S.L. Ghosh K. Common Benign Breast Concerns for the Primary Care Physician Cleve Clin. J. Med.201986576510.3949/ccjm.86a.1710030624185 · doi ↗ · pubmed ↗
- 7Hartmann L.C. Sellers T.A. Frost M.H. Lingle W.L. Degnim A.C. Ghosh K. Vierkant R.A. Maloney S.D. Pankratz V.S. Hillman D.W. Benign breast disease and the risk of breast cancer N. Engl. J. Med.200535322923710.1056/NEJ Moa 04438316034008 · doi ↗ · pubmed ↗
- 8Burke A. O’Driscoll J. Abubakar M. Bennett K.E. Carmody E. Flanagan F. Gierach G.L. Mullooly M. A Systematic Review of Determinants of Breast Cancer Risk among Women with Benign Breast Diseasenpj Breast Cancer 2025111610.1038/s 41523-024-00703-w 39955290 PMC 11829998 · doi ↗ · pubmed ↗