Molecular Motors Orchestrate Pause-and-Run Dynamics to Facilitate Intracellular Transport
Yusheng Shen, Kassandra M. Ori-McKenney

TL;DR
Molecular motors actively control pauses during intracellular transport, revealing a new principle that helps cells move cargo efficiently.
Contribution
The discovery that pauses in intracellular transport are active, motor-driven states governed by a quantitative law.
Findings
Pauses during intracellular transport are active and driven by molecular motors, not passive drifting.
A quantitative law (Deff ∝ v²) links the diffusivity during pauses to the velocity during runs across different motors and cargo.
Reduced motor engagement leads to slower transport and longer pauses, impairing cargo delivery.
Abstract
Intracellular transport is essential for cellular organization and function. This process is driven by molecular motors that ferry cargo along microtubules, but is characterized by intermittent motility, where cargoes switch between directed runs and prolonged pauses. The fundamental nature of these pauses has remained a mystery, specifically whether they are periods of motor detachment and passive drifting or states of active motor engagement. By combining single-particle tracking with large-scale motion analysis, we discovered that pauses are not passive. Instead, they are active, motor-driven states. We uncovered a unifying quantitative law: the diffusivity of a vesicle during a pause scales with the square of its velocity during a run. This parabolic relationship, Deff ∝ v2, holds true for both kinesin and dynein motors, different cargo types, and a variety of cellular…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
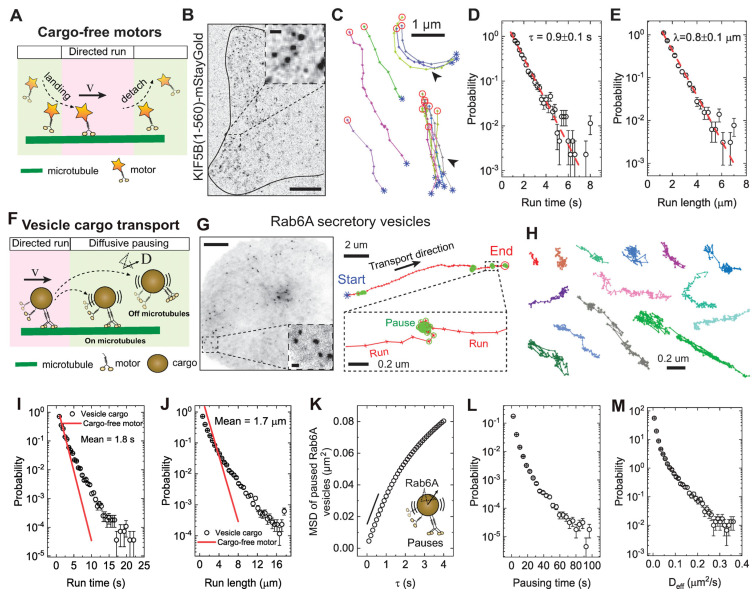 Figure 1
Figure 1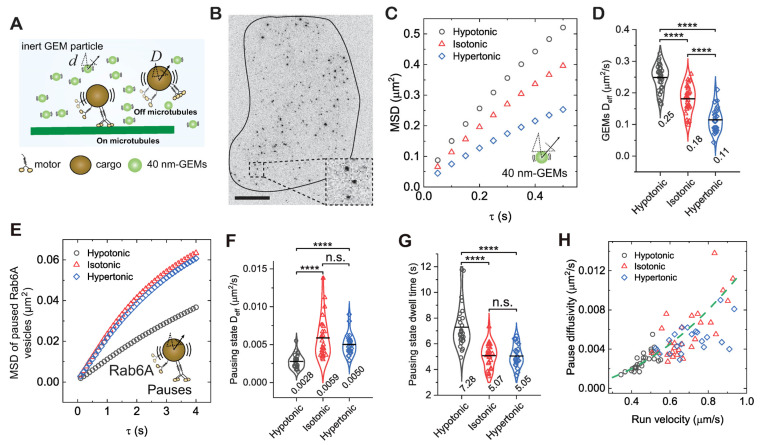 Figure 2
Figure 2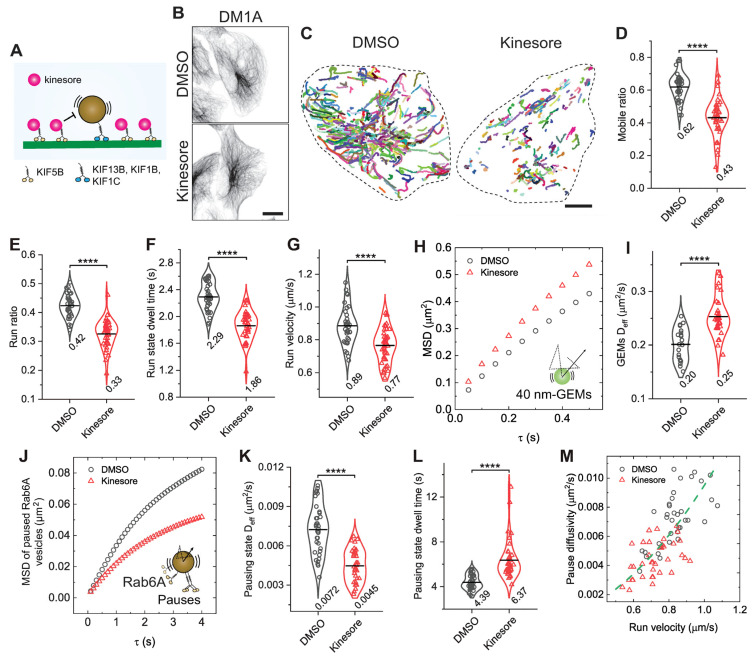 Figure 3
Figure 3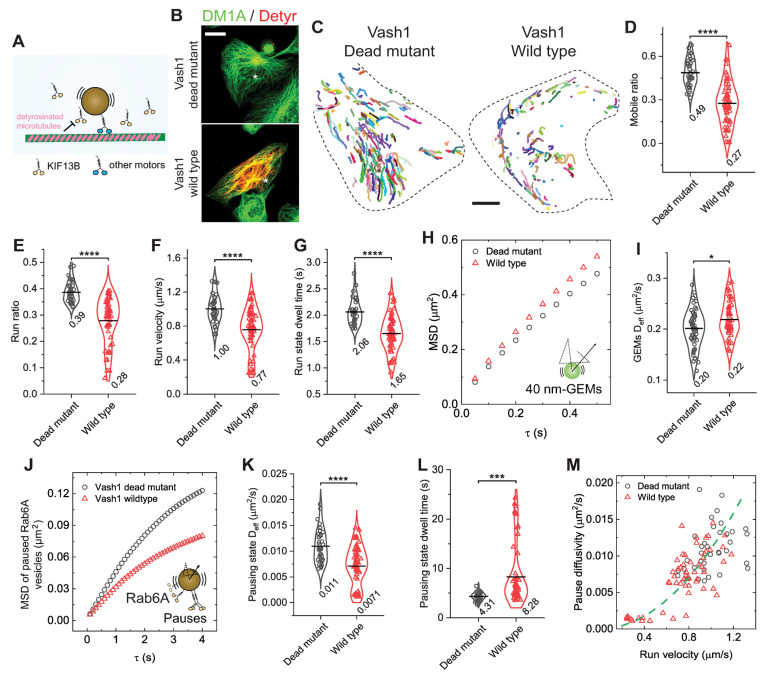 Figure 4
Figure 4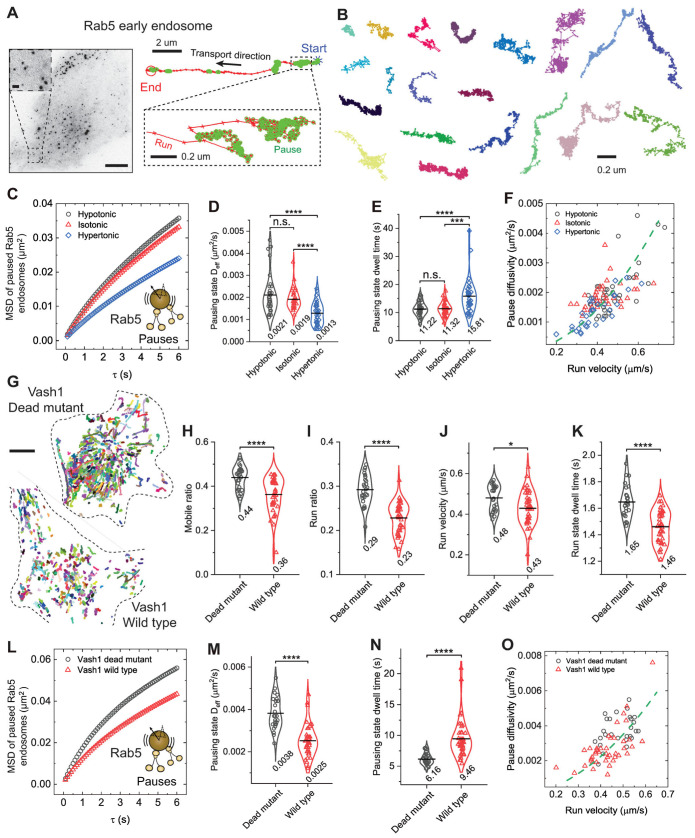 Figure 5
Figure 5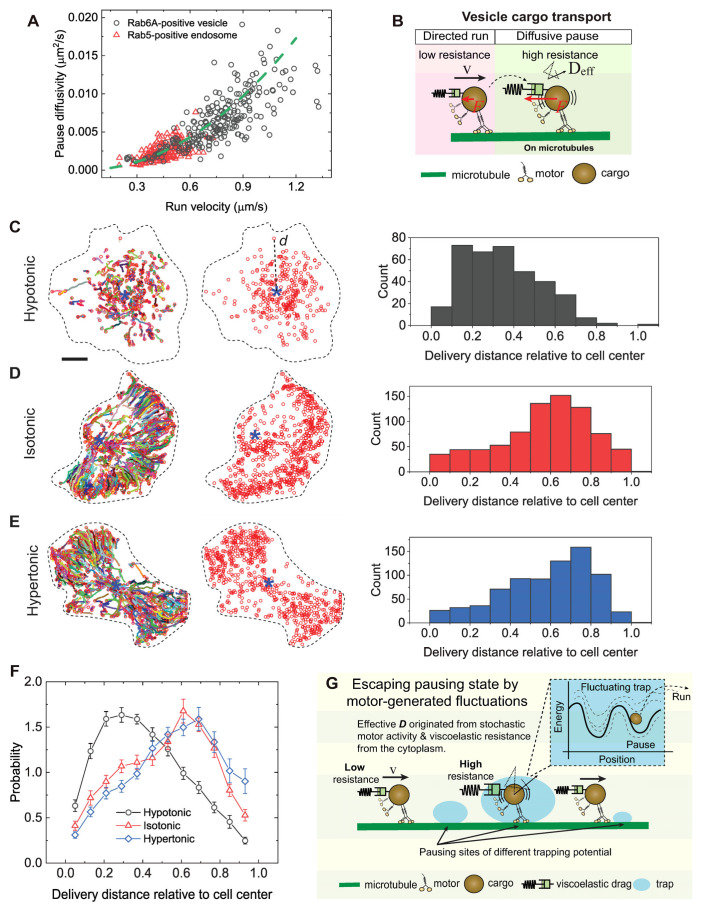 Figure 6
Figure 6- —NIH
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrotubule and mitosis dynamics · Micro and Nano Robotics · Nuclear Structure and Function
1. Introduction
Mammalian cells expend considerable energy to transport materials through their crowded cytoplasm. This active transport is powered by molecular motors, such as kinesins and cytoplasmic dynein, which move cargoes directionally along microtubule tracks to maintain cellular organization and homeostasis [1]. By converting chemical energy into mechanical work, these motors enable long-range transport that overcomes the limits of passive diffusion, particularly for large cargoes such as vesicular organelles and ribonucleoprotein complexes in the dense, viscoelastic cytoplasm [2,3,4,5]. To operate efficiently, motor–cargo complexes must adapt to a dynamic and complex intracellular landscape shaped by physical obstacles [6], membrane–membrane contacts [7,8], microtubule-associated proteins (MAPs) [9,10,11,12,13,14], and tubulin post-translational modifications (PTMs) [15,16,17,18,19], all of which modulate motor activity.
Perhaps due to this complexity, a hallmark of intracellular transport is intermittency. Cargoes, including endosomes, lysosomes, secretory vesicles, mitochondria, and autophagosomes, switch between short, directed runs and longer, seemingly stalled, diffusive pauses [20,21,22,23,24,25,26,27,28,29]. These pauses can consume up to 80% of a cargo’s travel time, presenting a paradox. While runs clearly reflect active motor-driven motion, the fundamental nature of pauses remains debated. This contrasts sharply with the behavior of individual cargo-free motors in vitro, which move processively along bare microtubules [11,30,31,32,33]. Why, then, do cargoes in cells spend most of their time in a seemingly inefficient paused state?
In cells, pauses have been attributed to various causes, including physical barriers [6,24,34], transient membrane tethers [8,23,35], regulation by MAPs and tubulin PTMs, and mechanical “tug-of-war” between opposing motors [12,21,22,27,36,37]. However, a critical and unresolved question remains: during a pause, are motor-cargo complexes actively engaged with the microtubule, withstanding viscoelastic cytoplasmic forces, or do they detach from the microtubule and passively diffuse? Directly distinguishing these possibilities in living cells has been challenging because it requires resolving the tripartite interactions of microtubules, motors, and cargoes with both high spatial and temporal resolution coupled with large-scale statistical power to analyze highly heterogeneous behaviors [24,29]. Resolving this is essential for understanding how cells precisely allocate energy to enable long-range intracellular transport.
To address this question, we combined single-particle tracking with large-scale statistical motion-state analysis of cargo-free kinesin-1 and its physiological cargo, Rab6A-positive secretory vesicles. We altered cytoplasmic fluidity, motor–cargo coupling, and motor–microtubule interactions and found that vesicle mobility during pauses is governed by motor activity in the run state and is independent of cytoplasmic viscosity. Parallel analysis of dynein-driven retrograde transport of Rab5-positive early endosomes revealed the same coordinated relationship, indicating a conserved mechanism between transport directions. Together, our live-cell observations uncover a robust quantitative coupling between pause-state diffusivity and run-state velocity, D_eff_ ∝ v^2^, that holds across motor families, perturbations, and cargo types. This unifying principle demonstrates that motor engagement directly coordinates both motility states, thereby ensuring robust cargo delivery through the complex intracellular landscape.
2. Materials and Methods
2.1. Cell Line
BEAS-2B cells (ATCC, CRL-9609) were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Thermo Fisher Scientific, Waltham, MA, USA), supplemented with 10% fetal bovine serum (FBS), 50 units/mL of penicillin, and 50 µg/mL of streptomycin. All cell cultures were maintained in a 95% air/5% CO_2_ atmosphere at 37 °C. The cell line was routinely confirmed to test negative for mycoplasma contamination. For live cell imaging, cells were seeded at a density of approximately 5 × 10^4^ cm^−2^ on a glass coverslip, which was placed in a 35 mm polystyrene tissue-culture dish. Transfections were performed by using the FuGENE 6 (Promega, Madison, WI, USA) according to the manufacturer’s instructions. Cells were generally transfected for 8 h with 1 µg of plasmids when the density reached ~80% confluency.
2.2. Antibodies and Immunostaining
For immunostaining, we used mouse monoclonal antibodies against α-tubulin (clone DM1A, T9026; Sigma-Aldrich, St. Louis, MO, USA) and rabbit polyclonal antibodies against detyrosinated α-tubulin (AB3201; Sigma-Aldrich). The following secondary antibodies were used: Alexa Fluor 488– and 647–conjugated goat antibodies against rabbit and mouse IgG (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) for immunofluorescence. For immunofluorescence cell staining, BEAS-2B cells were generally fixed in −20 °C methanol for 10 min, and then blocked with 4% bovine serum albumin (BSA; Sigma-Aldrich) in PBS at room temperature for 2 h. Next, cells were incubated with primary antibodies against detyrosinated tubulin (1:200) and α-tubulin (1:500), and corresponding secondary antibodies (1:200), each for 1 h. Stained cells were mounted onto a microscope slide with ProLongTM Gold Antifade Mountant (Invitrogen) and then examined using spinning disk confocal microscopy. The spinning disk confocal was performed on an inverted research microscope Eclipse Ti2-E with the Perfect Focus System (Nikon, Tokyo, Japan), equipped with a Plan Apo 60× NA 1.40 oil objective, a Crest X-Light V3 spinning disk confocal head (Crest-Optics, Rome, Italy), a Celesta light engine (Lumencor, Beaverton, OR, USA) as the light source, a Prime 95B 25MM sCMOS camera (Teledyne Photometrics, Tucson, AZ, USA) and controlled by NIS elements AR software version 6.22.01 (Nikon). The fluorescent images for BEAS-2B cells were collected over a stack of vertical z-sections across the entire cell’s ~4 µm thickness. The final fluorescent images and their fluorescent intensities shown in the main text are based on the z-averaged images by using Fiji software (https://fiji.sc/).
2.3. Generation of Plasmids
The expression vectors used in this study were pDeltaCMV-KIF5B(1-560)-mStayGold, pEGFP-Rab6A (Addgene, Watertown, MA, USA, plasmid 49469), pEGFP-Rab5 (Addgene plasmid 49888), pCMV-VASH1-EGFP-P2A-SVBP, pCMV-VASH1(C168A, catalytically dead mutant)-EGFP-P2A-SVBP, pCMV-VASH1-mScarlet-P2A-SVBP, pCMV-VASH1(C168A, catalytically dead mutant)-mScarlet-P2A-SVBP, pDeltaCMV-VASH1-mScarlet-P2A-SVBP, pDeltaCMV-VASH1(C168A, catalytically dead mutant)-mScarlet-P2A-SVBP and pCDNA3.1-pCMV-PfV-GS-Sapphire (Addgene plasmid 116933; for mammalian expression of 40 nm-GEMs). mStayGold was synthesized as a gBlock based on the published sequences [38]. The truncated KIF5B proteins (a.a. 1-560) were cloned into a pDeltaCMV vector (a gift from Dr. Scott Hansen for expressing proteins at low levels [39]) with a C-terminal mStayGold cassette. VASH1 and SVBPs were cloned into a pDeltaCMV vector or pCMV vector split by a P2A self-cleaving peptide with an EGFP or mScarlet cassette at the C-terminus of VASH1 [26,27]. The single amino acid mutations were generated by PCR. All cloning was performed using Gibson assembly. All constructs were verified by DNA sequencing (Plasmidsaurus, Eugene, OR, USA).
2.4. Osmotic and Drug Treatments
To manipulate the cytoplasm fluidity, cells were treated with and imaged in extracellular osmotic environments ranging from hypoosmotic to hyperosmotic conditions (250–400 mOsm). These solutions were prepared by adding D-mannitol to a hypotonic base solution (40 mM NaCl, 5 mM KCl, 1 mM CaCl_2_, 2 mM MgCl_2_, 10 mM HEPES, pH 7.4; 91 mOsm) to maintain a constant ionic strength. The final D-mannitol concentrations were 0.28 M (~400 mOsm, hypertonic), 0.2 M (~310 mOsm, isotonic control), 0.15 M (~250 mOsm, hypotonic), and 0.1 M (~200 mOsm, hypotonic) [26,40]. Cells were typically treated with the osmotic solutions 10 min before imaging. To activate kinesin-1 activity, cells were treated with kinesore (100 µM, Torcris, Bristol, UK) in culture medium for 1 h before being fixed or for 10 min before being imaged using TIRF microscopy.
2.5. Total Internal Reflection Fluorescence Microscopy
TIRF microscopy experiments were performed on an inverted research microscope Eclipse Ti2-E with the Perfect Focus System (Nikon), equipped with a 1.49 NA 100× TIRF objective with the 1.5× tube lens setting, a Ti-S-E motorized stage, piezo Z-control (Physik Instrumente, Karlsruhe, Germany), LU-N4 laser units (Nikon) as the light source, an iXon DU897 cooled EMCCD camera (Andor, Belfast, Northern Ireland, UK) with an high-speed emission filter wheel (ET480/40M for mTurquoise2, ET525/50M for GFP, ET520/40M for YFP, and ET632/60M for mRuby2; Chroma, Bellows Falls, VT, USA). The microscope was controlled with NIS Elements software (Nikon, Tokyo, Japan, version 6.22.01). All live-cell experiments were performed in a live-cell imaging chamber (H301-Nikon-TI-S-ER, Oko Labs, Naples, FL, USA) that was equipped to the microscope to provide optimal culture conditions (95% air/5% CO_2_ atmosphere at 37 °C) for cells during imaging. After cell transfection, the cell-containing glass coverslip was mounted on a coverslip holder (SC15012, Aireka Cells, Tokyo, Japan), which was finally mounted on the microscope. KIF5B(1-560) was recorded at 5 fps for 3 min. Rab6A and Rab5 vesicles were recorded at 10 fps for 3 min. 40 nm GEMs were recorded at 20 fps for 2 min. Due to the limited detection depth of TIRF microscopy (~100–200 nm), the vesicle, motor, and GEMs trajectories analyzed here primarily reflect two-dimensional motion within the imaging plane, while excursions with substantial z-displacement rapidly fall out of focus and are not included in the analysis. The recorded images have 16 bits of gray scale and a spatial resolution of 512 × 512 pixels, with the width of each pixel = 107 nm in our TIRF optical setup.
2.6. Single Particle Tracking and Analysis
Single particle tracking (SPT) was performed using a homemade tracking program written in Matlab (Mathworks, Natick, MA, USA) as previously described [26,41], which is based on the standard tracking algorithm [42,43]. With this advanced SPT algorithm, we were able to obtain the position r*(t)* at time t for KIF5B motors, Rab6A-positive secretory vesicles, Rab5-positive early endosomes and GEMs, and their trajectories were constructed from the consecutive images. This algorithm allowed us to achieve a spatial tracking resolution of ~20 nm.
To characterize the mixed-motions of Rab6A-positive secretory vesicles and Rab5-positive early endosomes, we used an algorithm that could automatically identify and extract the states of diffusive “jiggling” movement and the states of directed “runs”, as described previously [26,44,45]. Briefly, we determined the motion state of an arbitrary point in the trajectory by analyzing the turning angles, θ, around it at 5 different time steps to calculate the local directional persistence of the point. Each turning angle was discretized as 1 if θ < π/2, corresponding to forward motion without a turn, or 0 if θ ≥ π/2, corresponding to a backward movement with a turn. The five discretized values were then averaged, and a threshold of 0.5 was used to classify the local trajectory segment as a run state (⟨θ⟩ ≥ 0.5) or a paused state (⟨θ⟩ < 0.5). To suppress transient switching between states, a sliding window of 10 time steps (1 s) was applied to the resulting time series. Finally, only run-state segments with durations longer than 0.3 s were retained as true runs in the analysis. Trajectories containing no directed runs were considered immobile. The mobile ratio was defined as the ratio of the number of mobile trajectories over the total number of trajectories in a single cell. The run ratio was defined as the ratio of the total time spent in the run state over the total travel time of a vesicle. The mobile ratio and run ratio generally characterize how active the motors are in moving vesicles from the “pauses” to the “runs” states at the whole cell level. In this study, only the cell-averaged run ratio is reported. The run state dwell time is defined as the cell-averaged duration of individual run segments extracted from vesicle trajectories in a cell. Directed runs longer than 20 time-steps (2 s) were used to compute the run velocity. This is achieved by computing the mean squared displacements (MSDs), , and fitted to to obtain the run velocity . The run velocity, , was calculated for each run segment, and the reported run velocity represents the average over all extracted run segments in a cell. Diffusive pauses longer than 10 time-steps (1 s) were used to compute the effective pause diffusivity D_eff_. D_eff_ was obtained by fitting the MSDs to . Run-state and pause-state dwell time of each trajectory were obtained by averaging the dwell time of each run and pause segment, respectively. Because microtubules in cells adopt a wide range of geometries, including straight, buckled, parallel, bundled, and networked configurations, the pause–run statistics reported in this study represent ensemble-averaged behavior within intact cells. These measurements encompass the full range of microtubule geometries present under each experimental condition but do not resolve potential geometry-dependent effects.
To quantify the delivery distance for Rab6A-positive vesicles, we calculated the distance between the end position of each trajectory to the cell center . Because microtubules in BEAS-2B cells are radially organized, we defined the cell center as the position where all vesicle transport started. In each cell, we normalized d as , such that from cells of different sizes are in the same range . For the quantification of KIF5B motor run time, we only considered trajectories that lasted longer than 5 time-steps (1 s).
To study the diffusion dynamics of GEMs, we first selected the mobile trajectories from the whole set of GEM trajectories. This is achieved by computing the radius of gyration R_g_(τ) of each GEM trajectory obtained over a time period of τ,
where N is the total number of time steps in each trajectory, x_i_ and y_i_ are the projections of the position of each trajectory step on the x- and y-axis, respectively, and ⟨x⟩ and ⟨y⟩ are their mean values. Physically, R_g_ quantifies the size of a GEM trajectory generated during the time lapse τ. A cutoff value of (R_g_′)c = 0.3 was used in the experiment, below which the GEMs trajectories are treated as immobile ones [26,41]. Here, is the normalized radius of gyration, with ⟨R_g_⟩ being the mean value of R_g_. MSDs of individual mobile trajectories were then computed and fitted to to obtain the trajectory-based effective diffusion coefficients of GEMs, D_eff_, in BEAS-2B cells.
2.7. Statistics
Data are expressed as mean ± s.e.m. unless specified otherwise. Graphs were created using OriginPro (OriginLab, Northampton, MA, USA, version 2025b). Statistical tests were performed with a two-tailed unpaired Student’s t-test. The statistical details of each experiment can be found in the figure legends.
3. Results
3.1. Vesicle-Bound Motor Complexes Exhibit Distinct Transport Dynamics Compared to Cargo-Free Kinesin-1 Motors
To investigate the origins of pausing during intracellular transport, we compared the movement of cargo-free kinesin-1 motors to their physiological cargo, secretory vesicles labeled with EGFP-Rab6A. These vesicles are primarily driven anterogradely by kinesin-1 and kinesin-3 motors [25,26,46]. While kinesin-1 moves processively along bare microtubules in vitro (Figure 1A), vesicle cargoes pause frequently in cells, raising the question about the nature of these pauses [10,31,47]. To perform live single-molecule imaging of kinesin-1 in BEAS-2B cells, we tagged the truncated kinesin-1 motor, KHC(1–560), with the bright and stable fluorescent protein, mStaygold [38], and transiently expressed it at low levels using a truncated cytomegalovirus (CMV) promoter (Figure 1B) [39]. BEAS-2B cells, a human lung bronchial epithelial cell line, typically spread as a thin sheet with a large projection area (up to 100 μm × 100 μm) on glass coverslips, and their microtubules are largely arranged laterally near the basal membrane [26]. This geometry provides the advantage of collecting large datasets to capture the spatiotemporal dynamics of cargo transport across the whole cell. The truncated construct lacks the cargo-binding domain of kinesin-1, allowing us to directly examine how the in vivo microtubule landscape influences the motility of the active dimeric motor KHC(1–560) [1,48]. In BEAS-2B cells, single KHC(1–560)–mStayGold motors exhibited unidirectional, processive motility along linear tracks without pausing, showing some preferential movement along specific microtubules (Movie S1 and Figure 1C), consistent with previous findings [49,50]. By analyzing over 2000 motor tracks, we found that KHC(1–560) exhibited a characteristic run time of ~0.9 s and run length of ~0.8 µm on in vivo microtubules (Figure 1D,E), despite the heterogeneous landscape of MAPs and tubulin PTMs [26]. This suggests that single cargo-free kinesin-1 motors move rather freely along microtubules in cells, similar to their behavior on bare microtubules in vitro.
Given that single motors do not pause, we next asked why vesicles driven by multiple motors pause so frequently. We used a previously established algorithm to extract distinct motion states from vesicle trajectories [26]. For Rab6A-positive secretory vesicles in BEAS-2B cells, pauses occurred randomly along microtubules and accounted for up to 60% of the total travel time (Movie S2 and Figure 1F–H). Analysis of over 51,000 run segments from Rab6A-vesicle trajectories revealed that both run-time and run-length distributions were not a simple exponential and were significantly longer than that of single cargo-free KHC (1–560) motors, with mean values of 1.8 s and 1.7 µm, respectively (Figure 1I,J). This is consistent with transport being driven by multiple motors [25,51]. The pausing states themselves were characterized by confined, jiggling movements, and some pauses exhibited a distinct, strip-like pattern consistent with microtubule orientation (Figure 1H). These pauses exhibited diffusive-like behavior at short timescales (<1 s), with mean square displacements (MSDs) increasing linearly with delay time (τ), before transitioning to confined motion at longer timescales (Figure 1K). The pausing behavior was highly heterogeneous, with both pausing duration and diffusivity varying more than 50-fold between events (Figure 1L,M), suggesting that diverse cellular mechanisms likely regulate these pauses.
3.2. Rab6A Vesicle Pauses Are Microtubule-Bound States Coupled to Motor Activity
Because pauses during vesicle transport often occurred along linear trajectories consistent with microtubule orientation (Figure 1H), we next sought to determine whether these pauses represented states in which the motor–vesicle complex remained associated with, or became detached from, microtubules (Figure 1F and Figure 2A). If pauses represent detached states, vesicle mobility should positively correlate with cytoplasmic fluidity, similar to the behavior of sub-micron-sized, inert particles diffusing in the cytoplasm (Figure 2A) [52]. To test this hypothesis, we exposed cells to extracellular osmotic environments ranging from hypoosmotic to hyperosmotic conditions (250–400 mOsm) in order to either increase or decrease cytoplasmic fluidity [26]. We then quantified these fluidity changes in BEAS-2B cells by tracking the motion of 40 nm diameter genetically encoded multimeric (GEM) nanoparticles, a well-established probe for characterizing cytoplasmic fluidity in diverse cell types (Figure 2B–D) [53]. Consistent with previous findings [26,54], GEM diffusivity increased under hypotonic treatment and decreased under hypertonic treatment (Figure 2B–D), confirming that cytoplasmic fluidity was modulated accordingly.
We next investigated the effect of cytoplasmic fluidity on the pausing mobility of Rab6A-positive vesicles. Unexpectedly, hypotonic treatment, which increased cytoplasmic fluidity, reduced vesicle diffusivity during pauses by 52.5%, whereas hypertonic treatment, which decreased cytoplasmic fluidity, had no detectable effect compared with the isotonic control (Figure 2E,F). Notably, the reduced diffusivity under hypotonic conditions correlated with longer vesicle dwell times in the pausing state, which increased 1.44-fold (Figure 2G). The response of pausing behavior to changes in cytoplasmic fluidity correlated with that of run behavior reported previously for Rab6A-positive vesicles, where hypotonic treatment inhibited directed runs [26]. To further define the relationship between the two states, we plotted pause-state diffusivity against run-state velocity for Rab6A-positive vesicles. Strikingly, all data from different fluidity conditions collapsed onto a single parabolic curve (Figure 2H), revealing a robust positive correlation between pausing mobility and run velocity that is independent of cytoplasmic fluidity. These results indicate that the motor–vesicle complex remains associated with microtubules during pauses, and that reduced motility in the run state is coupled with reduced mobility in the pausing state, leading to prolonged trapping of vesicles in the pausing state.
3.3. Kinesin-1 Engagement Governs Vesicle Motility in Run and Pause States
To directly examine the role of kinesin-1 in coordinating vesicle motility during run and pause states, we treated cells with kinesore, a small-molecule compound that has a dual effect of activating kinesin-1 on microtubules while simultaneously disrupting its association with native cargoes [55,56], like Rab6A-positive vesicles (Figure 3A). While this severs the specific kinesin-1-cargo linkage, the net effect on the vesicle is a reduction in the number of processive, cargo-bound motors capable of generating cooperative force. Treating BEAS-2B cells with 100 µM kinesore for 1 h did not noticeably alter the radial microtubule network (Figure 3B). As predicted, acute kinesore treatment impaired the run motility of Rab6A-positive vesicles, significantly reducing all measured run parameters compared with control, with decreases of 30.6%, 21.4%, 18.8%, and 13.5% in the mobile ratio, run ratio, run-state dwell time, and run velocity, respectively (Figure 3C–G). This is consistent with the established mechanism of kinesore, which promotes kinesin-1 binding to microtubules while severing its interaction with native cargo adaptors, thereby reducing the number of motors productively engaged on the vesicle [55,56].
We next asked if reducing active kinesin-1 motors on vesicles similarly affects the pausing state. Despite kinesore increasing cytoplasmic fluidity (Figure 3H,I), Rab6A-positive vesicle diffusivity during pauses was reduced by 37.5%, while trapping times increased 1.45-fold (Figure 3J–L). Kinesore-induced increases in cytoplasmic fluidity may result from enhanced microtubule remodeling, which could actively agitate the surrounding cytoplasm [55,57]. Nonetheless, the correlation between pause-state diffusivity and run-state velocity mirrored that observed in cells under varying osmotic conditions (Figure 3M), suggesting that mobility in both states is intrinsically linked to active kinesin-1 simultaneously engaging with vesicle cargo and microtubules.
3.4. Microtubule Detyrosination Coordinates Vesicle Motility in Run and Pause States
Having established that disrupting motor–cargo interactions with kinesore impairs both run and pause states, we next asked if a similar effect would occur by perturbing the motor’s engagement with the microtubule track itself. We focused on microtubule detyrosination, a modification known to alter motor engagement, including the kinesin-3 motor, KIF13B [19,50,58,59]. Given KIF13B’s role in transporting Rab6A-positive secretory vesicles [25], we tested if detyrosination would disrupt the coordination of pause–run motility at the motor–microtubule interface (Figure 4A).
Detyrosination of α-tubulin is catalyzed by an enzyme complex composed of a vasohibin (VASH1 or VASH2) and a small vasohibin-binding protein (SVBP) [60,61]. To complement our acute pharmacological and osmotic perturbations, we also employed a slower, genetic approach by modulating the microtubule landscape through expression of VASH1. Transient expression of wild-type VASH1 in BEAS-2B cells markedly increased microtubule detyrosination compared with the catalytic-dead mutant (C168A), consistent with our previous findings (Figure 4B) [27]. We found that increasing microtubule detyrosination drastically impaired the run motility of Rab6A-positive vesicles with decreases of 44.9%, 28.2%, 23.0%, and 19.9% in the mobile ratio, run ratio, run velocity, and run-state dwell time, respectively, consistent with previous in vitro results (Figure 4C–G) [19]. While its effects are motor-specific, increasing detyrosination in cells robustly inhibits the overall anterograde motility of Rab6A vesicles (Figure 4C–G), establishing it as a functional perturbation that reduces net motor-driven force. This allows us to test whether impairing force generation at the motor-microtubule interface coordinately affects run and pause states, similar to perturbations at the motor-cargo interface. We then assessed the pausing state. Analysis of 40 nm GEM diffusion showed that increased microtubule detyrosination slightly increased cytoplasmic fluidity, in agreement with previous measurements (Figure 4H,I) [27]. Strikingly, despite the more fluid environment, vesicles on detyrosinated microtubules showed substantially reduced diffusivity by 35.5% and prolonged trapping during pauses by 1.91-fold (Figure 4J–L). This result mirrors the effect of kinesore treatment. The relationship between pause-state diffusivity and run-state velocity followed the same parabolic curve observed across perturbations (Figure 4M). Together, these results demonstrate that impairing the motor-microtubule interaction coordinately regulates vesicle motility in both run and pause states. This finding, combined with our kinesore data, reveals that the effective number of motors productively engaged in transport is the key parameter governing this coupled behavior.
3.5. Motor Engagement Coordinate Pause–Run Dynamics of Retrograde Transport
Our findings support that the run-pause dynamics of kinesin-driven vesicles are coordinated by motor engagement. We next asked whether a similar mechanism governs dynein-driven retrograde transport. To address this, we performed analogous analyses on Rab5-positive early endosomes, which are predominantly transported retrogradely by dynein–dynactin complexes in BEAS-2B cells (Movie S3 and Figure 5) [23,26,62]. Compared with kinesin-driven Rab6A-positive vesicles, Rab5-positive endosomes exhibited more frequent pauses, spending approximately 82% of their total transport time in the pausing state (Figure 5A,B and Figure S1). Similar to Rab6A vesicles, these pauses were diffusive-like and highly heterogeneous, with many trajectories displaying strip-like patterns indicative of microtubule association (Figure S1 and Figure 5B). We previously found that hypotonic treatment only modestly enhances Rab5-endosome run behavior, whereas hypertonic treatment downregulates it [26]. Consistent with a motor-engaged pausing-state, the diffusivity of Rab5-positive endosomes during pauses did not scale with the increased cytoplasmic fluidity under hypoosmotic conditions but instead remained largely unchanged, paralleling the modest increases observed in run behavior (Figure 5C–E).
We then impaired motor-microtubule engagement. Since microtubule detyrosination inhibits the landing of dynein–dynactin complexes on microtubules via the p150 CAP-Gly domain in vitro [18,63], we increased microtubule detyrosination by expressing VASH1. This efficiently impaired the run behavior of Rab5-positive endosomes with decreases of 18.2%, 20.7%, 10.4%, and 11.5% in the mobile ratio, run ratio, run velocity, and run-state dwell time, respectively (Figure 5G–K). As a direct consequence, pausing-state diffusivity markedly decreased by 34.2% and trapping times during pauses increased by 1.54-fold (Figure 5L–N). Across all perturbations, we observed a robust correlation between pause-state diffusivity and run-state velocity (Figure 5F,O). Notably, this relationship for retrograde transport of Rab5-endosomes closely paralleled that observed for kinesin-driven transport of Rab6A-positive vesicles. When data from both cargo types were visualized together, they follow a common nonlinear scaling trend that can be represented by a parabolic curve within experimental uncertainty (Figure 6A). This shared trend indicates that coordination between run and pause behaviors arises from microtubule-based motor engagement across diverse motor systems, rather than representing independent motility states.
3.6. Motor-Mediated Escape from Pauses Ensures Efficient Cargo Delivery
The robust coupling between pause and run states raised a key question: are these truly distinct states, or two manifestations of a single process where motors navigate variable local resistance? When resistance is low, vesicles move freely, like cargo-free motors. When resistance is high, a tug-of-war between motor forces and the viscoelastic cytoplasm produces jiggling, diffusive-like motion around microtubules, trapping vesicles transiently as pauses (Figure 6B). The number of engaged motors, therefore, governs a vesicle’s mobility and, even more importantly, its ability to escape these transient traps. Since vesicles spend the majority of their transit time in the pause state, an efficient escape rate is vital for delivery. To test this hypothesis, we analyzed the final delivery positions of Rab6A vesicles under osmotic conditions that differentially regulate motor activity [26]. Rab6A-positive vesicles are typically transported from the perinuclear Golgi to the cell periphery [25,26]. Under hypoosmotic conditions, which impair motor engagement, vesicles were stranded near the cell midzone. In contrast, under iso- and hyperosmotic conditions, where motor engagement is more effective, vesicles were successfully delivered to the periphery (Figure 6C–F). Given that vesicles spend most of their transit time in the pause state and that hypotonic stress disproportionately prolongs and immobilizes pauses, these results indicate that robust motor activity, which ensures both directed runs and efficient escape from prolonged pauses, is a critical determinant of successful long-range cargo delivery.
4. Discussion
Taken together, our results point to a shared quantitative scaling relationship between pause-state diffusivity and run-state velocity in BEAS-2B cells, with pause diffusivity increasing approximately with the square of run velocity (D_eff_ ∝ v^2^). While linear and quadratic descriptions are statistically indistinguishable over the limited velocity ranges accessible within individual datasets, the quadratic scaling is physically motivated by models of active particles driven by persistent, nonequilibrium forces [64,65,66]. In this framework, pausing and running are not independent motility states but reflect different manifestations of motor engagement with microtubules in the crowded cytoplasmic environment, where the same motor activity that drives directed motion also generates active fluctuations during pauses. This finding challenges the long-standing view that pauses represent cargo detachment from microtubules and passive diffusion [20,21,23,29,67,68]. Instead, our data strongly supports that pauses predominantly correspond to active, motor-engaged states where teams of motors continuously generate force against local resistance. This integrated run-pause dynamic enables vesicles to actively navigate a heterogeneous intracellular landscape, balancing directed motion with transient trapping.
The finding that kinesore, which increases kinesin-1 binding to microtubules but severs its cargo link, reduces pause mobility despite increasing cytoplasmic fluidity, is incompatible with a detachment-diffusion model. Similarly, detyrosination, which weakens specific motor-microtubule interactions, reduces pause mobility even as fluidity rises. Together, these perturbations demonstrate that pausing mobility is governed by the active forces generated by motors productively engaged with both track and cargo. This unifying principle explains the coordinated reduction in run and pause motility across all perturbations that reduce this effective motor number.
The observed correlation between motor engagement and cargo transport velocity in vivo contrasts with in vitro findings showing that increasing motor number has minimal effect on cargo velocity under dilute, low-load conditions [69,70]. This discrepancy likely arises because the intracellular environment is highly crowded and viscoelastic, presenting substantial resistive loads to moving vesicles, unlike the simplified, low-viscosity environments typically used in reconstituted assays. Indeed, intracellular cargos exhibit large velocity fluctuations, often spanning more than an order of magnitude, rather than moving at a single characteristic speed [71,72]. This interpretation is further supported by recent in vitro work demonstrating that when environmental crowding is introduced, increasing motor number can enhance cargo velocity through load sharing [73].
The prolonged pause escape times under reduced motor activity (Figure 2G, Figure 3L, Figure 4L, and Figure 5N) support a model in which motors perform the work necessary for vesicles to escape local energy barriers [74,75]. Within these traps, stochastic ATP-driven stepping of engaged motors generates fluctuating forces on the cargo. A team of motors produces larger force fluctuations than a single motor, effectively “rocking” the vesicle and increasing the probability that these forces will transiently overcome the local trapping potential. Thus, the number of engaged motors governs the escape rate by modulating the amplitude of active force noise. We conceptualize these trapping sites as potential wells of varying depths, and motor activity supplies the non-equilibrium fluctuations that drive barrier crossing and continuation of directed runs (Figure 6G).
Given that Rab6A-vesicles and Rab5-endosomes move predominantly unidirectionally (Figure 1G and Figure 5A) [26], and that inhibiting kinesin-1 with kinesore coordinately reduces both run velocity and pause diffusivity (Figure 3), the observed motor-engaged pausing is inconsistent with a symmetric tug-of-war “draw” state [22,29,36,76]. Instead, the coordinated impairment suggests that the same ensemble of engaged motors governs motility in both states, with pauses arising when their collective force is insufficient to overcome local resistive barriers. This interpretation is reinforced by the nearly 3-fold difference in pausing mobility between Rab6A-vesicles and Rab5-endosomes (Figure 2F and Figure 5D). If cargos were in a “draw” state and simply following fluctuating microtubules to which they are tethered, their pause mobilities would be similar. Therefore, the observed diffusive-like motion of pausing vesicles is also unlikely to be driven by microtubule remodeling. Instead, the resistance to motor-generated pulling forces may arise from transient tethering or obstruction by other intracellular structures [8,23,24,35,77]. In the paused state, these structures act as viscoelastic elements against which teams of motors pull while remaining bound to the vesicle. Stochastic stepping of multiple motors generates intermittent forces that rock the vesicle within these local traps. Over timescales longer than individual motor steps, this back-and-forth motion appears diffusion-like, consistent with the trajectories observed in Figure 1H and Figure 5B. While most displacements remain largely confined, vesicles occasionally exhibit persistent slip events in the direction of transport due to net motor pulling. As the number of engaged motors decreases, the net pulling force and force fluctuations are reduced, leading to smaller displacements and a lower effective diffusion coefficient. Consequently, a team with more engaged motors generates larger force fluctuations, leading to larger displacement amplitudes and thus a higher effective diffusion coefficient during the confined, rocking motion. This mechanism generates a positive correlation between run velocity and pause diffusivity, consistent with the observed trend.
Cells also appear to actively adjust motor-generated forces to adapt to environmental changes. Under hyperosmotic stress, the 39% decrease in cytoplasmic fluidity (Figure 2D) was matched by a 32% reduction in pausing mobility for Rab5-endosomes, while Rab6A-vesicles showed no change (Figure 2F). This discrepancy suggests that kinesin-driven transport may selectively upregulate force to counterbalance the elevated viscoelastic drag, potentially through enhanced recruitment of kinesin-1 by MAP7, which is enriched on microtubules under hypertonicity [10,26,77,78]. Conversely, under hypoosmotic stress, the 39% increase in fluidity (Figure 2D) coincided with a striking 47% decrease in pausing mobility for Rab6A-vesicles (Figure 2F), while Rab5-endosomes were unaffected (Figure 5D). We previously showed that hypotonic treatment disassociates MAP7 from microtubules and increases microtubule detyrosination [26]. The combined loss of the kinesin-1 recruitment by MAP7 and inhibition of kinesin-3/dynein via detyrosination likely underlies the strong suppression of motor activity and pausing mobility, overriding the changes in fluidity. This highlights how cells can tune transport by locally modulating the microtubule landscape, allowing motor-cargo complexes to adapt to a heterogeneously crowded cytoplasm. Our use of complementary perturbations (osmotic stress, acute kinesore treatment, and enzymatic microtubule detyrosination) was designed to acutely modulate the functional engagement of motors from distinct angles (cytosolic environment, motor-cargo linkage, motor-track recognition). The consistent finding across all perturbations, that reducing effective engagement coordinately impairs run and pause motility, demonstrates that this functional parameter, rather than motor abundance per se, is the key governor of the coupled transport dynamics.
5. Conclusions
Overall, the robustness of the D_eff_ ∝ v^2^ scaling across cargo types, motor families, and perturbations suggests it is a fundamental physical property of motor teams navigating a resistive environment. We speculate this relationship could be a general feature of processive molecular motors operating in confined, viscoelastic cytoplasm. Future studies could test this model in vitro using optical traps to apply calibrated loads to beads driven by defined motor teams, measuring whether the parabolic relationship between velocity under low load and diffusivity under high load is reconstituted.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Vale R.D. The Molecular Motor Toolbox for Intracellular Transport Cell 200311246748010.1016/S 0092-8674(03)00111-912600311 · doi ↗ · pubmed ↗
- 2Mogre S.S. Brown A.I. Koslover E.F. Getting around the Cell: Physical Transport in the Intracellular World Phys. Biol.20201706100310.1088/1478-3975/aba 5e 532663814 · doi ↗ · pubmed ↗
- 3Brangwynne C.P. Koenderink G.H. Mac Kintosh F.C. Weitz D.A. Intracellular Transport by Active Diffusion Trends Cell Biol.20091942342710.1016/j.tcb.2009.04.00419699642 · doi ↗ · pubmed ↗
- 4Bonucci M. Shu T. Holt L.J. How It Feels in a Cell Trends Cell Biol.20233392493810.1016/j.tcb.2023.05.00237286396 PMC 10592589 · doi ↗ · pubmed ↗
- 5Moeendarbary E. Valon L. Fritzsche M. Harris A.R. Moulding D.A. Thrasher A.J. Stride E. Mahadevan L. Charras G.T. The Cytoplasm of Living Cells Behaves as a Poroelastic Material Nat. Mater.20131225326110.1038/nmat 351723291707 PMC 3925878 · doi ↗ · pubmed ↗
- 6Ross J.L. Shuman H. Holzbaur E.L.F. Goldman Y.E. Kinesin and Dynein-Dynactin at Intersecting Microtubules: Motor Density Affects Dynein Function Biophys. J.2008943115312510.1529/biophysj.107.12001418227130 PMC 2275709 · doi ↗ · pubmed ↗
- 7Voeltz G.K. Sawyer E.M. Hajnóczky G. Prinz W.A. Making the Connection: How Membrane Contact Sites Have Changed Our View of Organelle Biology Cell 202418725727010.1016/j.cell.2023.11.04038242082 PMC 11830234 · doi ↗ · pubmed ↗
- 8Jongsma M.L.M. Berlin I. Wijdeven R.H.M. Janssen L. Janssen G.M.C. Garstka M.A. Janssen H. Mensink M. van Veelen P.A. Spaapen R.M. An ER-Associated Pathway Defines Endosomal Architecture for Controlled Cargo Transport Cell 201616615216610.1016/j.cell.2016.05.07827368102 PMC 4930482 · doi ↗ · pubmed ↗