The Synthetic Extracellular Matrix as a Maestro of the In Vitro Stem Cell Niche: Orchestrating Fate and Function
Subhajit Giri, Pratyush Rajesh

TL;DR
This paper discusses how synthetic extracellular matrices can better control stem cell behavior compared to traditional methods like Matrigel.
Contribution
The paper introduces synthetic extracellular matrices as a defined and tunable alternative to Matrigel for hiPSC culture and differentiation.
Findings
Synthetic ECM matrices offer fully defined and tunable properties for hiPSC culture.
They can be functionalized with signaling peptides and growth factors for directed differentiation.
Synthetic matrices are suitable for clinical-grade tissue engineering and regenerative medicine.
Abstract
Human-induced pluripotent stem cells (hiPSCs) have an innate ability to differentiate into the three germ layers: the ectoderm, endoderm, and mesoderm. By using targeted differentiation methods and carefully controlling growth factors, morphogens, and signaling modulators, hiPSCs can be guided to develop into specific lineage cell types. For clinical applications of hiPSCs and their derivatives, it is crucial to use xenogen-free, chemically defined culture media, reagents, recombinant growth factors, morphogens, and extracellular matrix (ECM) scaffolds. One major obstacle is the widespread use of Matrigel as an hiPSC culture matrix. Matrigel, derived from Engelbreth–Holm–Swarm (EHS) mouse sarcoma, is an extract of basement membrane material with a complex, poorly defined, and variable composition. It also exhibits batch-to-batch variability in mechanical and biochemical properties and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
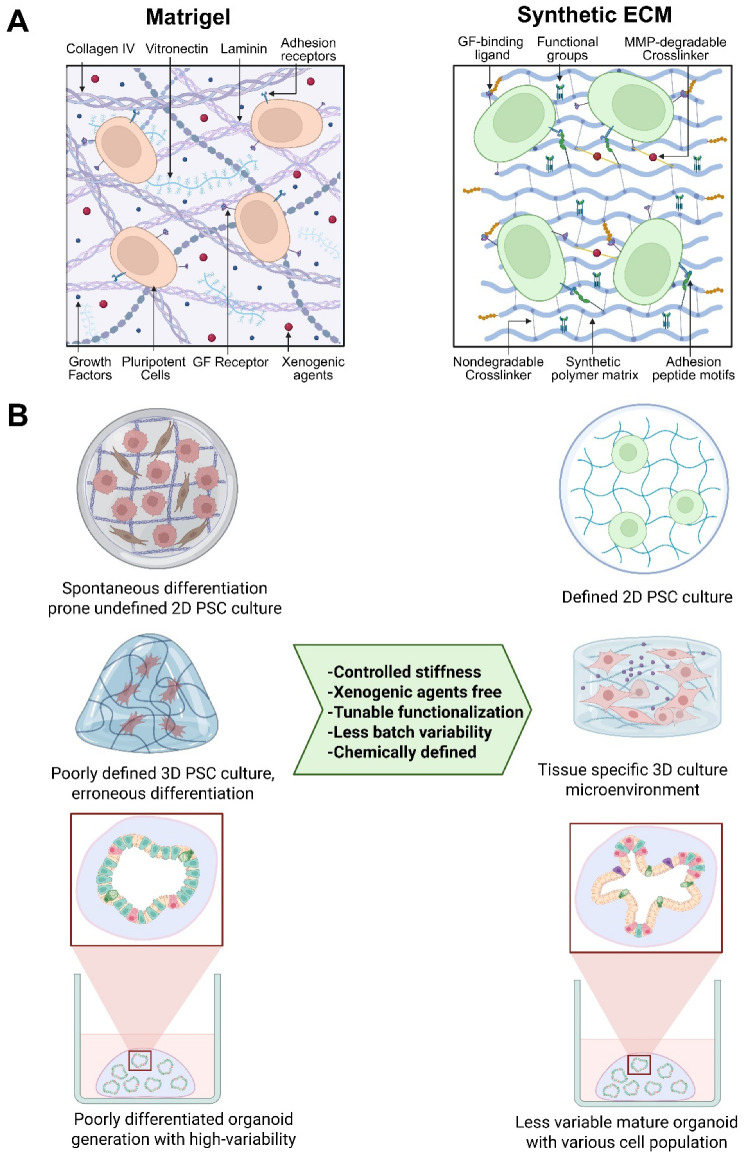 Figure 1
Figure 1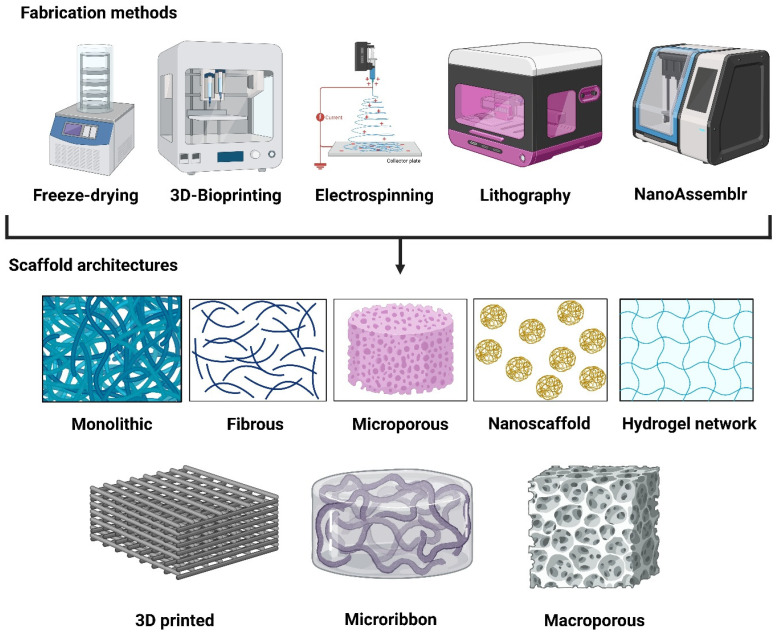 Figure 2
Figure 2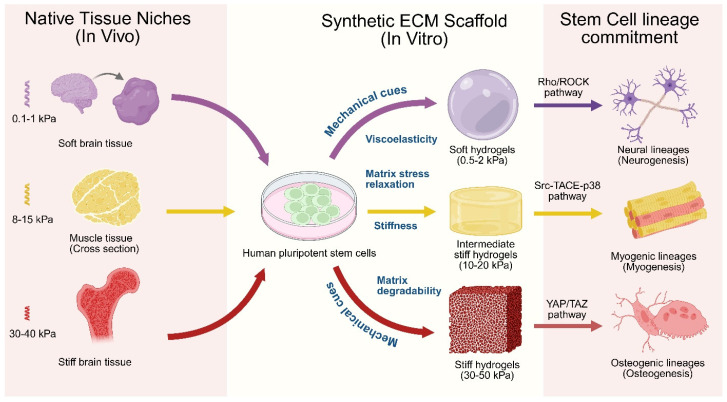 Figure 3
Figure 3- —Cancer Discovery, Medical College of Wisconsin
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPluripotent Stem Cells Research · Tissue Engineering and Regenerative Medicine · Planarian Biology and Electrostimulation
1. Introduction
The extracellular matrix (ECM) is an intricate network of fibrous proteins, proteoglycans, glycoproteins, and glycosaminoglycans that surrounds cells and serves as an anatomical scaffold, providing critical physical information that controls cellular behavior [1]. The ECM serves as a microenvironment rich in dynamic feedback signaling that regulates stem cell identity and function. The composition, organization, and stiffness of the ECM contribute to biological processes, including adhesion, migration, and differentiation [2]. The ECM consists of complexes of bioactive molecules such as growth factors, cytokines, adhesion ligands, and other signaling molecules, all of which interact with specific stem cell receptors and initiate downstream signaling pathways to modulate many of these cellular processes [3]. For instance, integrins are cell surface receptors for the ECM, such as fibronectin and laminin, and play a crucial role in regulating stem cell proliferation, differentiation, and survival [4,5]. ECM elasticity and stiffness also direct stem cell differentiation and fate determination. It has been shown that softer matrices increase the self-renewal capacity of mesenchymal stem cells (MSCs), whereas stiffer ECM scaffolds favor differentiation towards osteoblast lineages [6,7]. The ECM also acts as a depot for signaling molecules, such as growth factors and morphogens, which are sequestered and poised for release via enzymatic degradation or cell-mediated interactions, thereby generating local gradients that control stem cell migration, proliferation, and differentiation within the niche. The ECM is a dynamic entity rather than a static structure, constantly remodeled by resident cells. Different types of proteases secreted by stem cells can degrade and reorganize the surrounding ECM, altering niche composition and influencing stem cell fate and the behavior of neighboring cells. This dynamic interplay between stem cells and the ECM creates a self-regulating microenvironment that orchestrates stem cell function [8,9].
Understanding the intricate interplay between the ECM and stem cells holds immense potential for regenerative medicine. To serve this purpose, researchers can engineer synthetic ECMs. A synthetic ECM is an engineered biomaterial designed to mimic the structural, biochemical, mechanical, and functional properties of the native ECMs, while offering a precise control over composition, reproducibility, and reduced batch-to-batch variability [10]. Unlike natural ECMs, synthetic ECMs are typically composed of polymers, self-assembling peptides, biodegradable crosslinkers, and bioactive motifs, which enables tunable stiffness, degradability, and ligand presentation [11]. These matrices support cell adhesion, migration, proliferation, and differentiation with suitable physiochemical cues to the cells. Additionally, the ability to manipulate the ECM composition and signaling cues can potentially guide stem cell differentiation toward desired cell types for tissue repair and regeneration.
A tremendous number of efforts are underway to develop advanced bioinspired synthetic ECM and biomaterials to revolutionize regenerative medicine and stem cell-based therapeutic opportunities. Accumulating evidence suggests that the synthetic ECM can effectively replace the usage of Matrigel, a widely used ECM in stem cell and organoid-based research. At this inflection point, it is essential to keep track of the new inventions and knowledge base in the synthetic ECM and advanced engineered biomaterials domain. To help with this, a periodic collection of new information could be beneficial for research and for the therapeutic development community, which we tried to deliver in this review article.
2. Limitations of Matrigel as an Organoid Scaffold
Matrigel was first invented by Prof. Hynda Kleinman [12,13]. In the early stages of 3D cell culture research, ‘Matrigel’ was a gold-standard ECM for in vitro studies, revolutionizing stem cell and organoid-based research. However, its application in cell biology research and therapeutic cell manufacturing is hindered by its complexity and batch-to-batch compositional variability. Numerous proteomic profiling and protein content analysis efforts identified more than 14,000 unique peptides and 2000 proteins in Matrigel [12,13,14,15,16,17,18]. Most of these proteins are structural proteins, growth factors, transcription factors, and cytokines [15,19]. Such growth factors and cytokines in the basement membrane could alter the characteristics of cultured stem cells and interfere with the developmental trajectory during directed differentiation. Growth factor reduced (GFR) Matrigel, which is structurally similar to standard Matrigel but contains fewer and lower concentrations of growth factors, could be an alternative to address this problem [14,19]. Another significant drawback is batch-to-batch variability, which introduces additional variability into experimental results. It has been shown that insulin-like growth factor 1 and epidermal growth factor were detected at nanogram-per-milliliter levels in a single batch of Matrigel [19]. In a later study, these growth factors were not detected in four independent Matrigel batches [15]. As summarized in Figure 1A, such inconsistency in key signaling molecules, including fibroblast growth factor 2 (FGF2) and Platelet-derived growth factors (PDGFs), has been reported across multiple batches of Matrigel [15].
Matrix viscoelasticity is a crucial mechanical cue for stem cell differentiation and organoid development [6,20,21]. The mechanical properties and stiffness of Matrigel were also inconsistent, as found in batch-to-batch and intra-batch variability [22,23]. Using atomic force microscopy, the elastic modulus (‘stiffness’) of two batches of Matrigel was reported to be 400–420 Pa, whereas another batch of Matrigel was 840 Pa [24]. Heterogeneity in local stiffness in Matrigel was also reported, where in situ mechanical interferometry identified a higher elastic modulus (1–3 kPa) [23,24]. Such inconsistency of Matrigel mechanical properties can impede proper stem cell culture and differentiation, organoid development, and tissue organization [20,25,26]. Another major drawback of Matrigel is the antigenicity and the presence of xenogenic contaminations, as it is an animal-derived ECM. Viral contamination, especially the lactate dehydrogenase-elevating virus (LDHV), has been found in multiple batches of Matrigel [27,28]. LDHV is a natural mouse virus that infects macrophages and can elicit immunogenic reactions and modulate tumor behavior [29,30]. The presence of such contaminants can impede mechanistic studies in cell biology experiments and hinder the interpretation of the underlying biological events. For these reasons, the usage of Matrigel in clinical applications and therapeutic cell therapy product development is limited. Using an animal-derived ECM, such as Matrigel, in human therapies carries the risk of zoonotic pathogens that can jeopardize patient safety. Matrigel cannot be fine-tuned to the microenvironment and the scaffold architecture required for stem cell differentiation and organoid culture, thereby limiting its use across a broader spectrum.
Geltrex (Gibco) and Cultrex (R&D Systems/Trevigen) are formulated as alternatives to Matrigel with higher laminin contents, though both are derived from EHS mouse sarcoma tumors. Both Geltrex and Cultrex provide robust support to hPSC pluripotency, efficient human intestinal organoid development, and reduced viscosity for easier pipetting [31]. Geltrex excels in soft embedding for long-term brain organoid maturation up to 150 days [32]. Still, both remain animal/xeno-derived and poorly defined ECMs, restricting their use in advanced translational research and clinical applications. They exhibit low mechanical strengths and rapid degradation, and limit large organoid (>1 mm) culture due to hypoxia and nutrient diffusion issues. Overall, Geltrex and Cultrex bridge gaps but underscore the need for fully defined matrices [33,34].
3. Comparison of Matrigel, Natural ECMs, and Synthetic ECMs
Matrigel, tissue-derived natural ECMs, and fully synthetic ECMs each play distinct and complementary roles in organoid research, and the most appropriate choice depends on the specific experimental goals and translational context. Matrigel remains the most widely used matrix for early-stage exploratory studies and initial organoid establishment. Its biochemical richness, including a broad array of growth factors such as FGF-2, VEGF, and TGF-β, has made it highly effective for supporting a wide range of organoids, including intestinal, gastric, hepatic, and neural systems [14,35]. However, Matrigel’s tumor-derived, xenogeneic origin, substantial batch-to-batch variability, high cost, and lack of GMP-compliant manufacturing severely limit its suitability for clinical translation and regenerative medicine [36,37].
Decellularized, tissue-derived ECMs offer a more physiologically relevant alternative by preserving native, tissue-specific biochemical cues and age-dependent ECM signatures. These matrices often support organoid development that matches or even exceeds that achieved with Matrigel, making them particularly valuable for optimizing differentiation protocols and modeling disease in contexts where native microenvironmental signaling is critical [38,39]. For example, gastrointestinal dECM hydrogels contain a richer repertoire of tissue-specific proteins than Matrigel, which has been linked to enhanced stemness, improved differentiation capacity, long-term culture stability, and successful organoid transplantation. Despite these advantages, natural ECMs still face regulatory hurdles and retain some degree of batch variability, even when produced under GMP-compatible conditions [38].
Fully synthetic ECMs are increasingly favored for mechanistic studies and translational applications that demand precision, reproducibility, and regulatory compliance. These systems allow independent and quantitative control over key matrix parameters, including stiffness, viscoelasticity, ligand density, and enzymatic degradability, while eliminating the confounding variability inherent to undefined natural matrices [35,40]. As a result, synthetic ECMs are particularly well suited for high-throughput drug screening, the systematic interrogation of ECM–cell interactions, and the development of xeno-free, GMP-compliant scaffolds for regenerative therapies (Figure 1B). Notably, certain polyethylene glycol (PEG)-based hydrogels have been shown to support human intestinal organoid formation, viability, and differentiation at levels comparable to, or exceeding, those achieved with Matrigel [41,42].
The main limitation of synthetic ECMs lies in their reduced biological complexity, as they lack the full spectrum of endogenous growth factors and cryptic binding sites present in natural matrices. However, this apparent drawback is also a strength. The modular design of synthetic systems allows researchers to precisely introduce defined adhesive ligands and protease-sensitive motifs, enabling mechanistic insights that are difficult to obtain in complex, ill-defined environments like Matrigel. For instance, synthetic matrices with independently tunable ligand spacing and stress–relaxation dynamics have revealed fundamental mechanotransduction principles that govern stem cell fate decisions, which we discuss elaborately later in Section 7. Emerging hybrid approaches that combine synthetic backbones with defined natural components aim to balance biological functionality with experimental control, potentially bridging the gap between discovery-driven research and clinical translation. The individual strengths and limitations of Matrigel, natural ECM, and synthetic ECM are consolidated in Table 1.
4. Synthetic ECM Scaffolds
The inherent limitations of Matrigel in stem cell and organoid-based applications have prompted the search for synthetic alternatives that can be fine-tuned with greater flexibility. In this context, numerous synthetic polymer-based 2D or 3D scaffolds have been developed. They can be modified through precise fabrication processes to control molecular weight, composition, crosslinking density, matrix geometry, and mechanical and rheological properties [10,11,43,44,45]. These matrices can also be modified with biofunctional chemical groups specific to the stem cell and differentiated cell types at a given developmental stage, thereby fine-tuning the scaffold for continued proliferation or differentiation [43]. The most commonly used polymers as precursor scaffolds are polyacrylamide (PAM), PEG, polylactic acid (PLA), poly(lactic-co-glycolic acid) (PLGA), polyhydroxyethyl methacrylate (PHEMA), and polyvinyl alcohol (PVA). In contrast, their synthetic derivatives were used in most of the stem cell-based applications [46,47]. PAM is an uncharged and bioinert substance that generally does not bind to the cell surface glycoproteins. PAM hydrogels can be tuned to varying stiffness and crosslinking capacity by incorporating different cell-adhesion peptides and biofunctional groups, thereby meeting user-defined requirements. However, due to the toxicity of PAM precursors, acrylamide and bisacrylamide, and the nature of polymerization, PAM cannot be used in 3D cell encapsulation and restricts its use only to 2D culture systems. PEG is another widely used synthetic hydrogel that has been used in most studies as an artificial scaffold. PEG is hydrophilic, bioinert, highly amenable to chemical modification, and nontoxic, making it a superior scaffold for 2D and 3D cell cultures (Figure 1A). Various polymerization chemistries, including photopolymerization, Michael addition, and enzymatic reactions, can be employed to produce PEG hydrogels with incorporated reactive groups [48,49,50,51].
Synthetic ECM scaffolds are designed to recapitulate the biochemical, mechanical, and structural cues of native tissue microenvironments while offering extraordinary reproducibility and tunability. Different fabrication methods vary depending on the desired geometric architecture and tissue culture applications. The major fabrication approaches used to generate monolithic hydrogels, fibrous scaffolds, microribbons, and macroporous scaffolds are summarized in Figure 2. Monolithic hydrogels are continuous, non-porous networks typically formed through the bulk crosslinking of synthetic polymers (e.g., PEG, gelatin methacrylate, etc.) using UV or visible light photopolymerization, enzymatic reactions, or Michael-type additions, whereas rheometers and photoreactors are generally used for mechanical tuning [52]. To mimic the native ECM fibril architecture, nano- to micro-scale fibrous scaffolds are commonly fabricated through electrospinning using high-voltage power supplies, syringe pumps, and rotating collectors [53,54]. Microribbon scaffolds (μRB) consist of elongated hydrogel fragments produced by wet spinning or microfluidic extrusion followed by mechanical fragmentation and secondary crosslinking, enabling a high porosity and anisotropy (Figure 2). This type of scaffold is highly beneficial for bone and cartilage tissue or organoid culture [55,56]. In other types of scaffolds, microporous scaffolds are generated via porogen leaching, gas foaming, or cryogelation, using molds, freeze–thaw systems, and vacuum-drying equipment (Figure 2). In contrast, macroporous scaffolds with well-defined interconnected pores are often fabricated using 3D bioprinting or sacrificial templating with extrusion-based printers and temperature-controlled stages. Both such scaffolds provide 3D structural support, guide the self-assembly of stem cells to complex tissue-like structures, facilitate nutrient/waste exchange via pores, and improve organoid-based disease modeling and its applications in drug testing and regenerative medicine [57,58]. Other types of biomimetic scaffolds are nanoscaffolds that incorporate nanoscale features produced by self-assembly, phase separation, or nanolithography, using techniques such as peptide amphiphile assembly, spin coating, or atomic force microscopy-assisted patterning, as shown in Figure 2 [59,60]. Collectively, these fabrication approaches enable a precise control over scaffold mechanics, architecture, and biofunctionality to recapitulate tissue-specific ECM cues.
Scaffold architecture plays a critical role in regulating stem cell organization, fate specification, and lineage commitment by controlling 3D spatial confinement, ligand presentation, and mechanical cues within the cellular microenvironment. Certain architectural parameters such as porosity, pore size, fiber alignment, and dimensionality directly influence cell adhesion, migration, cytoskeletal organization, and other biochemical signaling pathways. Highly porous 3D-printed and electrospun scaffolds create interconnected channels that improve nutrient diffusion and facilitate cellular interactions. The porous scaffold has been proven effective for the efficient differentiation of hPSCs to pancreatic β-cell clusters and the myogenic differentiation of mesenchymal stem cells (MSCs) [58,61]. The size and shape of pores also play a critical role; for example, in bone regeneration biology, the smaller pores (~50–100 µm) support the cell attachment, while the larger ones (~200–400 µm) facilitate nutrient diffusion and angiogenesis [62]. The porous scaffolds with different pore geometries can also direct hMSCs to adopt distinct morphologies and differentiation outcomes. The octagonal architectures enhanced cell elongation and osteogenic differentiation more than hexagonal or square designs, illustrating that geometric arrangement can bias fate decisions [63]. Aligned fibrous scaffolds guide cell elongation and anisotropic organization, which efficiently differentiate cardiomyocytes from hiPSC-derived cardiac progenitor cells with improved functionality [64]. It also favors differentiation toward neural, tendon, or muscle lineages by activating the YAP/TAZ pathway and integrin-mediated signaling [65]. Similarly, electrospun and nanostructured scaffolds help hPSC aggregation, with an elevated adhesion to increased surface area contact, and promote efficient lineage commitment during differentiation [66,67]. Overall, scaffold structural parameters such as pore geometry, size, and fiber orientation act as biophysical signals that modulate stem cell organization and differentiation outcomes in tissue engineering contexts.
5. Synthetic ECM as a Superior Choice of Matrix for Human Pluripotent Stem Cell (hPSC) Culture
Human embryonic stem cells (hESCs) and induced pluripotent stem cells (hiPSCs) emerged as essential resources for modern disease modeling, drug discovery efforts, personalized medicine, regenerative medicine, and clinical applications [68,69]. hESCs/hiPSCs can proliferate indefinitely, are readily manipulated through genetic engineering, such as CRISPR gene editing, and retain the ability to differentiate into the three germ layers and, potentially, into any cell type in 2D or into specific organoids in 3D under a directed differentiation protocol. These characteristic attributes put hPSCs at the forefront of cutting-edge disease modeling, drug discovery, and clinical cell therapy [70,71]. Thriving hPSC culture depends on optimized culture protocols, chemically defined media, and matrix components in such applications. In the early days of hESC-based research, mouse embryonic fibroblasts (MEFs) served as feeder layers for hESCs. However, this co-culture system raised concerns about animal-origin contamination and spontaneous differentiation due to batch effects, which significantly affect culture efficiency. To mitigate this, the use of Matrigel as a basement membrane became popular, as it supports the proliferation and maintenance of pluripotency [72]. However, the undefined batch-to-batch variability and animal-derived xenogenic nature of Matrigel limit its clinical use in hPSC cultures, highlighting the advantages of fully synthetic matrix systems (Figure 1B).
In search of a Matrigel replacement for hPSC culture, a zwitterionic polymer, poly(2-(methacryloyloxy)ethyl dimethyl (3-sulfopropyl)ammonium hydroxide) (PMEDSAH), was first developed as a fully synthetic coating for long-term hPSC culture. It has been shown that PMEDSAH supports hPSC pluripotency and the gene expression network comparably to Matrigel, even after 20 passages [73,74]. In another study, a high-throughput screen of matrices identified a synthetic polymer poly(methyl vinyl ether-alt-maleic anhydride) (PMVE-alt-MA) that supports long-term attachment, the maintenance of pluripotency, and a normal karyotype, while reducing the spontaneous differentiation of hPSCs [75]. In addition to the matrix’s physical properties, the cell–matrix interaction properties were also investigated to develop an efficient surface coating material. It has been shown that hPSCs attach to Matrigel through integrin receptor subunits α5, α6, αv, β1, and β5 [76,77]. The fibronectin-derived three-amino-acid peptide Arg-Gly-Asp (RGD), which binds to both αvβ3 and αvβ5 integrins, provides a superior adhesion surface for hPSCs [78]. Improvements to the RGD scaffold were achieved through cyclization to cyclo(Arg-Gly-Asp-d-Phe-Lys) (cRGDfK), which is a more adhesive surface than standard laminin- or fibronectin-derived matrix peptides. In the same study, when cRGDfK was coupled with poly(acrylamide-co-propargyl acrylamide) (PAPA) brushes, it supported long-term hPSC cultures without affecting the pluripotency and karyotypic abnormalities [79]. In a subsequent study, cRGDfK was modified with 64 distinct PEG-thiol norbornene-based synthetic hydrogels and evaluated for hPSC cultures. Several candidates showed a better hPSC culture, maintenance, and preservation of the pluripotency [80].
Previous studies found that heparin sulfate proteoglycans can bind to basic fibroblast growth factor (bFGF) and protect it from proteolytic degradation, which promotes hPSC culture by maintaining the self-renewal ability and pluripotency [81,82,83]. Based on these findings, several efforts have been made to develop heparin sulfate-based synthetic hydrogels. One study developed a heparin-mimicking hydrogel, PAM6-co-PSS2, by copolymerizing poly(sodium 4-styrene sulfonate) (PSS) with PAM. This synthetic scaffold was able to support long-term hPSC culture in more than 20 passages with maintained pluripotency [84]. In 2010, a study found different proteoglycan-binding peptide sequences of GWQPPARARI, FHRRIKA, or GKKQRFRHRNRKG that can interact with cell surface glycosaminoglycans and support long-term hPSC self-renewal and pluripotency over three months [85]. In a later study, a PAM hydrogel was functionalized with a different vitronectin (VN)-derived peptide, GKKQRFRHRNRKG, and synthesized for elastic moduli (0.7, 3, and 10 kPa), and it has been shown that the rigid version (10 kPa) supports long-term hPSC cultures [86]. In addition, other VN-derived peptides were also conjugated with PAM/PEG hydrogels to produce an array of synthetic scaffolds. In a study, polyacrylic acid (PA) was used to coat the culture surface and was conjugated with a VN-derived peptide (Ac-KGGPQVTRGDVFTMP) and a bone sialoprotein-derived peptide (Ac-KGGNGEPRGDTYRAY) through EDC/NHS chemistry, which supported a long-term culture of multiple hPSC lines with chemically defined media [87]. Similarly, poly (OEGMA-co-HEMA) brushes were conjugated with VN peptides, which support the hiPSC culture over ten passages in a xeno-free and chemically defined media condition [88]. Another study used poly(vinyl alcohol-co-vinyl acetate-co-itaconic acid) (PVA) hydrogel grafted with a VN-derived peptide for hPSC cultures. It showed that it can support the hPSC culture for more than 20 passages in E8 media conditions [89]. In 2018, Sohi et al. used a chitosan film to immobilize VN-derived peptides and bFGF via an NHS-PEG-mal linker on its surface. They showed that it could support hiPSC self-renewal for an extended period in media containing FBS and bFGF [90]. In addition to these discoveries, a truncated version of human recombinant vitronectin (Vitronectin NC) was shown to be an excellent surface coating material for long-term hPSC cultures [91]. A study used carboxymethyl chitosan (CMC) on polydopamine-modified cell culture plates to immobilize the VN peptide via NHS/EDC chemistry. This coating material was proven to be supportive of hiPSC reprogramming and the long-term self-renewal (>20 passages) of multiple hPSC lines [92].
The hPSC culture system has evolved from 2D to 3D formats with advances in synthetic matrix technology. The 3D culture system promotes hPSC pluripotency and self-renewal by providing a better control over cell–cell and cell–matrix interactions, cellular morphology, and the culture microenvironment, which are crucial for hPSC growth [93,94,95,96]. In a pivotal study, the authors used an RGD-functionalized PEG hydrogel scaffold to culture hPSCs in a 3D format. They reported a 2.5-fold induction of pluripotency, reprogramming efficiency of human fibroblasts into hiPSCs, and homogeneous hiPSC colony generation [97]. In 3D scaffold preparation, electrospinning is a commonly used method. A 2014 study reported a layered gelatin nanofiber scaffold that supported an hPSC culture for more than 20 passages [98]. Later, this nanofiber was crosslinked with a microfiber layer composed of cellulose and polyglycolic acid, and the resulting scaffold supported hPSC survival for more than 2 months in mTeSR1 medium supplemented with Y27632 and methylcellulose [99]. In another study, PLGA and PMEDSAH were used to fabricate a nanofiber scaffold that supported hPSC adhesion and colony formation for two months without passaging [100]. Polystyrene electrospun fibers have also been used to develop a porous 3D scaffold that has been shown to support hiPSC culture for 10 passages [101]. The above examples of synthetic matrix hydrogels, other engineered scaffolds, and their applications in hPSC culture are summarized in Table 2.
6. Synthetic ECM as a Platform for Stem Cell Differentiation
With the advancement in our understanding of developmental biology, we are now more interested in stem cell-based organ-specific cell-type differentiation for in vitro disease modeling, drug screening, and regenerative medicine [69,102]. However, some challenges associated with stem cell culture, lineage-specific stem cell differentiation, the derivation of a homogeneous cell population, and providing a physiological microenvironment during culture and differentiation must be addressed before its clinical usage [103]. Previous evidence suggests that the surrounding microenvironment, especially the ECM composition and its physical and chemical characteristics, greatly influences stem cell differentiation towards lineage specificity [104,105]. The ill-defined composition of Matrigel is not the optimal choice for controlling microenvironmental cues during hPSC differentiation. To circumvent Matrigel-related challenges, researchers generated and tested synthetic ECMs and hydrogels to provide a tightly controlled microenvironmental scaffold for successful hPSC differentiation toward a homogeneous cell population. These synthetic ECMs and their application for lineage-specific hPSC differentiation will be discussed further.
6.1. Synthetic ECMs for Differentiation into Ectodermal Lineage
6.1.1. Neuronal Lineage
The neural ECM has a pivotal role in neuronal cell proliferation, differentiation, maturation, and migration throughout brain development [106,107]. The neuronal ECM primarily consists of hyaluronic acid (HA), heparan sulfate proteoglycans, chondroitin sulfate proteoglycans (CSPGs), laminins, and reelin [108,109]. It has previously been shown that neuronal progenitor cells (NPCs) differentiate more efficiently in softer ECMs that mimic the native brain ECM composition [110,111,112]. HA-based hydrogel scaffolds have been used to photoencapsulate ventral midbrain-derived NPCs, accelerating the maturation of mechanical properties and ultimately differentiating into neurons [113]. The synthetic hydrogel PuraMatrix was used as an ECM microenvironment to differentiate hESCs into neurons and astrocytes [114]. Later, in another study, the authors functionalized PuraMatrix with a laminin-derived peptide sequence (-GGSDPGYIGSR-) and a bone marrow homing factor peptide sequence (-GGPFSSTKT-). This modified scaffold showed an increased neuronal cell proliferation, enhanced adhesion, and elevated survival [115]. The PEG-based hydrogel was also used as a micropatterned scaffold substrate to induce single neural rosette formation and to increase radial outgrowth with peripheral neuronal differentiation [116]. In another study, the authors used a PEG hydrogel formulation that contains maleimide-functionalized PEG with integrin-binding peptides and PEG-dithiol with MMP-degradable peptides, which support human astrocyte culture and maintenance in star-shaped morphologies [117]. PEG was also conjugated with a continuous gradient of an N-cadherin (NCAM)-derived peptide sequence (HAVDI), which was associated with increased survival, neurite extension, and neural differentiation in hiPSC-derived neural stem cells (hNSCs) [118]. PLGA-based fibers were used as a mesh to generate micropatterned embryoid bodies (EBs), thereby facilitating the guided self-organization of cortical plates in brain organoids [119]. Another synthetic substrate, a polyacrylamide (PAM) hydrogel with a stiffness of 0.7 kPa functionalized with a glycosaminoglycan (GAG)-binding peptide (CGKKQRFRHRNRKG), was shown to enhance neuronal differentiation from hESCs [120]. A poly-ε-caprolactone-based microsphere was used to encapsulate guggulsterone for sustained release during hiPSC differentiation toward a neuronal fate, resulting in increased TUJ1+/Olig2+ neuronal aggregates without affecting neurite length or branching [121]. A biocompatible group conjugated conductive polymer, Poly(3,4-ethylenedioxy thiophene) doped with poly(styrene sulfonic acid) (PEDOT: PSS), was also used to stimulate neuronal differentiation from hiPSC-derived neuronal progenitor cells electrically. PEDOT: PSS has been shown to support hNSC viability, enhance adhesion, promote neuron formation with longer neurites, and cause no apparent cytotoxicity [122,123]. Similarly, another conductive polymer, polypyrrole doped with anionic dodecylbenzene sulfonate (DBS), was used as a neuronal induction scaffold, and, upon electrical stimulation, hNSCs differentiated into neurons more efficiently, exhibiting longer neurite lengths, greater branching, and an upregulated expression of neurotrophic growth factors [124,125]. An electrospun PLGA membrane was fabricated with single-walled carbon nanotubes, and, upon electrical stimulation, hiPSCs were efficiently differentiated into NSCs, with an electrical conductivity controlling neuronal maturation [126]. The applications of the above-mentioned synthetic matrices for neuronal lineage differentiation are summarized in Table 3.
6.1.2. Retinal Pigment Epithelial Cell Lineage
Retinal pigment epithelial cells arise from the developing anterior neuroectoderm, specifically from the anterior neural plate, during human development [131]. RPE cells have multimodal functions in the visual system, where they act as a physical barrier between the retina and choroidal blood vessels and play a crucial role in maintaining the photoreceptor (PR) conversion and storage of retinoids, the phagocytic removal of cellular debris, and other visual functions. RPE cell damage contributes to several degenerative eye diseases, including age-related macular degeneration (AMD). The advent of hESC/hiPSC-derived RPE cell differentiation in clinical settings provides an unprecedented opportunity for personalized curative therapies for AMD. In this context, the cGMP-compliant use of xeno-free materials, such as synthetic and defined ECMs, for RPE derivation is crucial. As in the literature, most studies have used Matrigel, laminin, poly-D-lysine, and fibronectin as ECM surfaces for hESC/hiPSC-derived RPE. The use of synthetic hydrogels and other synthetic ECMs holds promise for more efficient RPE derivation [132,133,134,135]. Scaffold-based approaches using synthetic or engineered ECM materials provide a prosthetic Bruch’s membrane that supports the formation of a polarized RPE monolayer, maintains cell–cell contacts, reduces immunogenic risk, and promotes long-term viability more effectively [136,137]. Additionally, synthetic ECM scaffolds can be designed with tunable mechanics and biodegradability to mimic native Bruch’s membrane permeability and to facilitate integration with host tissue, potentially improving transplantation outcomes and therapeutic efficacy in AMD [138].
For example, fibrin hydrogel was used to differentiate RPE from hiPSCs as a xeno-free, rapidly degradable synthetic ECM [139,140]. Another study tested four synthetic ECMs, like gelatin methacryloyl (GelMA), hyaluronic acid methacryloyl (HAMA), alginate, and fibrin hydrogels, for hESC-derived RPE adhesion and found that only fibrin hydrogel supports RPE adhesion [141]. In another study, human ARPE-19 cells were cultured on negatively charged poly (2-acrylamido-2-methyl propane sulfonic sodium) (PNaAMPS) and neutral poly(N, N-dimethylacrylamide) (PDMAAm) hydrogel surfaces and found that the usage of the PNaAMPS surface generates a very low level of reactive oxygen species (ROS) in the cultured RPE cells [142]. RGD-functionalized alginate hydrogel was also shown to elevate RPE cell differentiation in a 3D culture system [127]. PEG-grafted nanofiber surfaces have also supported RPE-cell attachment, proliferation, and maturation in vitro [128]. PEG/Gellan Gum (GG) hydrogel was also used as a biocompatible synthetic ECM surface for RPE cell attachment, survival, and proliferation [129]. In a recent study, several peptide fractions from vitronectin (PQVTRGDVFTMP) and the laminin β4 chain (PMQKMRGDVFSP) were grafted onto poly(vinyl alcohol-co-itaconic acid) (PAI) hydrogel and used as an ECM coating for hiPSC differentiation into RPE cells. The vitronectin-derived peptide fraction KVN2CK-grafted PAI hydrogel was found to support hiPSC differentiation into RPE cells, with an efficient proliferation and differentiation compared with Matrigel. These differentiated RPE cells were functional and were transplanted efficiently into a retinal degeneration rat model [130]. Table 3 summarizes the examples of synthetic ECMs that are promising candidates for RPE differentiation from hPSCs as 2D and 3D scaffolds, though they require further testing in future studies.
6.2. Synthetic ECMs for Differentiation into Endodermal Lineage
6.2.1. Hepatic Lineages
Synthetic ECMs for hepatocyte differentiation from hiPSCs and mesenchymal stem cells (MSCs) and liver tissue regeneration are becoming increasingly popular in personalized medicine. Chemically defined media and xeno-free synthetic ECMs with tunable biochemical and mechanical cues continually improve the hepatic differentiation in 2D and 3D culture formats. In a study, a collagen-coated PLGA scaffold was used to support the hepatic differentiation of MSCs in 3D, resulting in a higher hepatic marker expression and more mature metabolic functions than 2D differentiation [143]. In another study, a PEG-based 3D scaffold was conjugated with fibronectin and collagen and efficiently induced hepatocyte differentiation from MSCs, compared with the 2D differentiation system [144]. Physical parameters, such as ECM stiffness, also play a crucial role in hepatocyte differentiation, in concert with ECM composition. A softer (400 Pa) 2D micropatterned heparin hydrogel was shown to support efficient hepatocyte differentiation and maturation from MSCs, compared with a stiffer (43 kPa) hydrogel [145]. Efficient hepatocyte differentiation from resident liver stem cells was achieved in a softer PA hydrogel (400 Pa) [146]. A collagen-coated PA hydrogel with moderate stiffness (20–140 kPa) has also been employed to successfully differentiate hepatocyte-like cells (HLCs) from ESCs, with increased albumin secretion and metabolic activity [147]. A 3D synthetic ECM scaffold made of PEG-diacrylate (PEGdA)-hyaluronic acid hydrogel conjugated with hydrolytically degradable GRGDS peptides with variable stiffness was used to encapsulate and differentiate primary hepatocyte progenitor cells, HepRG, towards mature hepatocytes. The synthetic scaffold, with a stiffness of 2.8–6.17 kPa that mimics native liver stiffness, enabled the more efficient differentiation and maturation of HepRG-derived hepatocytes [148].
The porosity and pore size of the 3D synthetic ECM scaffold also play a crucial biophysical cue for hepatocyte differentiation from hPSCs/MSCs. A synthetic 3D ECM nanofibrous scaffold composed of poly(L-lactic acid)-co-poly(ε-caprolactone) (PLACL)/collagen, with 89% porosity, efficiently transdifferentiated MSCs into hepatocytes compared with its counterparts when used as a standalone scaffold [149]. A biohybrid scaffold composed of hiPSC-derived HLCs and 3D melt–electrospun poly-ε-caprolactone (PCL) was used to mature hiPSC-derived HLCs into hepatocytes with key functional attributes, including albumin synthesis, cytochrome P450 activity, and glycogen storage [150]. A highly porous electrospun poly (L-lactic acid) (PLLA)/collagen nanofiber scaffold was also used to differentiate human bone marrow-derived MSCs (hBMSCs) into hepatocytes that expressed key hepatic lineage markers and functional attributes [151]. Similarly, an electrospun poly-ε-caprolactone/collagen/polyethersulfone nanofiber scaffold was more effective than a 2D culture system at transdifferentiating hBMSCs into functional hepatocytes [152]. In a previous study, a collagen I-infused bioplotting poly-l-lactic acid (PLLA) scaffold was used to differentiate hiPSCs into hepatocytes, which produced viable, polarized hepatic cells and also formed bile canaliculi-like structures; the functional maturation was lower than that of the hepatocytes differentiated on decellularized liver ECM [153]. In a recent study, a synthetic ‘click’ hydrogel was formulated using dendritic polyglycerol-bicyclononyne (dPG-BCN) and poly(N-isopropylacrylamide)-co-polyethylene glycol azide (pNIPAAm-co-PEG-N3), which was tunable and had biochemical properties. This hydrogel was conducive to long-term hiPSC proliferation, as well as to human liver organoid differentiation and lineage specification. RGD-conjugated versions of the same hydrogel promoted the differentiation of cholangiocytes from hiPSCs via enhanced TGF-β activation, whereas the RGD-free version promoted hepatocyte lineage differentiation [154]. In another recent study, the authors fabricated Biomimesys^®^, composed of collagen I/IV fibers, chemically modified HA, and Arginylglycylaspartic acid (RGDS) adhesion peptides. This hydrogel was shown to be effective for HLO differentiation within 28 days, with upregulated liver marker gene expression, increased cellular diversity, enhanced cytochrome P450 activities, and the production of apolipoprotein (a) [155]. Advances in the development of synthetic ECM for HLO and hepatocyte differentiation from hPSCs and hMSCs will enhance the adoption of xeno-free human liver models for hepatic disease research and drug discovery (Table 4).
6.2.2. Intestinal Lineages
Human intestinal organoids (HIOs) differentiated from either hPSCs or adult Lgr5+ intestinal stem cells (ISCs) have emerged as an excellent tool for studying human intestinal development, disease modeling, in vitro drug testing, and regenerative medicine [170]. For consistent HIO generation and the use of xeno-free culture components in clinical applications, synthetic ECM with tunable stiffness, biochemical and mechanical cues, and biodegradability is gaining traction in HIO research. In 2017, a synthetic 4-arm PEG-maleimide-terminated hydrogel (PEG-4MAL) functionalized with an RGD peptide was used to differentiate HIOs from hPSCs. This fully defined hydrogel supported HIO differentiation from hiPSCs and maintained cell proliferation, apico-basal polarity, and structural complexity. The HIOs successfully engrafted onto the injured murine intestine and repaired the wound [41,42]. In a subsequent study, another synthetic PEG hydrogel was fabricated using an 8-arm vinyl sulfone-terminated PEG macromer (PEG-8VS) functionalized with a collagen I-mimicking adhesive peptide (GFOGER). It crosslinked with an MMP-degradable peptide to form primary human intestinal enteroids [156]. The enteroids retained proliferative capacity and apico-basal polarity and functionally responded to basolateral stimulation; they also expressed crypt cell- and Paneth cell-specific marker genes. Another synthetic PEG hydrogel was engineered through low-defect Thiol–Michael addition using an 8-arm PEG-VS hydrogel (2.5% w/v) containing an RGD peptide with an MMP-degradable crosslinker to grow patient-derived intestinal organoids [157]. Recently, a dynamic PEG-based hydrogel was developed using 8-arm PEG (8-PEG) as the backbone, with cytosine and vinyl sulfone covalently incorporated via Michael addition in a 50:50 ratio. This hydrogel, named Hybrid50, can reduce the stiffness of the 3D scaffold at a late stage of HIO development, thereby activating the YAP1 pathway and initiating symmetry breaking in HIOs [158]. In addition to different PEG hydrogels, a nonadhesive alginate hydrogel was used for hPSC-derived HIO differentiation and culture. This study first differentiated hPSCs toward the intestinal lineage on a 2D surface as spheroids. The spheroids were encapsulated in an alginate hydrogel to support continued differentiation and maturation toward the HIO fate. The functional and maturation profiles of these HIOs were comparable to those of HIOs differentiated in Matrigel as a 3D scaffold [169]. As a PEG-free 3D scaffold, a group recently developed a fully defined, tunable designer matrix, hyaluronan elastin-like protein (HELP), that supports the differentiation of HIOs from ISCs and the passaging of HIOs for a few generations [159]. The HELP hydrogel can be tuned for stiffness, the matrix stress–relaxation rate to support late-stage HIO development, and the matrix integrin–ligand concentration, providing layers of control for dynamic HIO development. As consolidated in Table 4, these synthetic ECMs can be used as 3D scaffolds and as a substitute for Matrigel in HIO differentiation.
6.2.3. Pancreatic Lineage
To date, 3D pancreatic organoids are developed from hPSCs, pancreatic ductal cells, and embryonic pancreatic progenitors (PPs), primarily using Matrigel as an ECM scaffold [171,172,173]. However, as the ECM composition of native pancreatic islets differs in the endocrine and exocrine parts of the pancreas, the use of defined and tunable synthetic ECMs could enhance the differentiation of hPSCs/PPs towards functional and mature pancreatic organoids [174,175]. In an early study, an enzymatically crosslinked PEG hydrogel was shown to maintain and expand PPs but failed to support further pancreatic organoid maturation [172]. Later, another group designed a 3D scaffold composed of PEG-conjugated collagen I and encapsulated hESC-derived PPs. This scaffold supported long-term islet organoid culture and maintained organoid size and morphology [160]. An activin A grafted gelatin-PLGA nanofiber scaffold was also shown to differentiate hiPSCs to pancreatic cells efficiently [161]. Other synthetic scaffolds, such as PLLA/PVA and PCL/PVA nanofibrous scaffolds, were also tested and shown to support the enhanced differentiation of insulin-producing pancreatic cells from hiPSCs [162,163].
Alongside these synthetic scaffolds, alginate-based scaffolds were also used to differentiate hPSCs into pancreatic islets. In a recent study, the authors fabricated a 3D hydrogel scaffold, namely PVAMA-AlgMA-GelMA, using PVA, alginate, and gelatin conjugated with methacrylic anhydride (MA). The 3D scaffold was further modified by coating its surface with 75% activin A and BMP4. Subsequently, hiPSCs were encapsulated in this modified scaffold and treated with retinoic acid (RA)-loaded solid lipid nanoparticles, thereby differentiating hiPSC-derived definitive endoderms into pancreatic islets [164]. Alginate fibers were also used for the intraperitoneal transplantation of hiPSC-derived PP-spheroids in diabetic mice, resulting in the reversal of hyperglycemia [176]. Differentiation efficacy was also improved for hiPSC-derived pancreatic islets when hiPSCs were encapsulated in an alginate hydrogel [177]. Recently, a Na-Alginate-Chitosan hybrid capsule hydrogel was shown to facilitate the formation of hiPSC-differentiated, mature, and functional pancreatic organoids in an all-in-water microfluidic system. The resulting organoids consist of both pancreatic α- and β-cells, exhibit an elevated expression of pancreatic hormone-specific genes and proteins, and functionally secrete insulin in response to glucose [165]. Similarly, aqueous droplet-filled alginate calcium hydrogel fibers (ADHFs) were fabricated in a microfluidic system, and pancreatic endocrine progenitors were encapsulated in the ADHFs. The resulting pancreatic organoids exhibited a high viability and functional maturation, secreting insulin [166]. Apart from alginate-based hydrogel, recently, a novel hydrogel ‘Amikagel’ was formulated through ring-opening polymerization between amikacin hydrate and poly(ethylene glycol) diglycidyl ether (PEGDE). The resulting Amikagel facilitated spontaneous spheroid formation and the differentiation of mature pancreatic β-cells from hESCs. An increased expression of key pancreatic transcription factors was reported for PDX1 and NKX6.1, enriching the committed endocrine population. Upon further differentiation, Amikagel-derived spheroids yield β-like cells with higher INS gene and C-peptide levels and show glucose-responsive insulin secretion, indicating the improved functional maturation of β-cells in vitro [167]. The applications of the above-mentioned synthetic matrices are summarized in Table 4.
6.2.4. Pulmonary Lineage
Synthetic ECMs are increasingly being adopted as alternatives to Matrigel for lung organoid culture, addressing limitations related to the undefined composition, variability, and lack of translational compatibility [178,179]. Among these, PEG-based hydrogels are the most extensively studied due to their excellent cytocompatibility, low immunogenicity, and modular chemistry that enables the independent control of stiffness, degradability, and ligand presentation [42]. Defined PEG systems, including PEG-4MAL and PEG-norbornene hydrogels functionalized with adhesive peptides and matrix metalloproteinase-sensitive crosslinkers, support human lung organoid formation comparably to Matrigel, promoting epithelial cyst growth, lumen formation, apicobasal polarization, and the expression of key lung lineage markers such as NKX2-1 and P63 [168,178]. The systematic tuning of matrix stiffness within these platforms has revealed that softer hydrogels approximating native lung mechanics preferentially enhance lung progenitor differentiation from hiPSCs, outperforming Matrigel-based cultures.
Beyond PEG, polysaccharide-based synthetic matrices such as alginate promote an increased airway epithelial diversity, yielding multiciliated, goblet, and basal cell populations organized in architectures that more closely resemble native airway tissue [169]. Additional synthetic polymers, including polyisocyanide (PIC), polyacrylamide, and polyvinyl alcohol hydrogels, offer further flexibility in fabrication and mechanical control [178]. Importantly, scaffold degradability has emerged as a critical determinant of lung organoid maturation. Non-degradable PEG-based microporous scaffolds restrict human lung organoid growth and maturation, largely maintaining cells in an immature NKX2.1^+^ progenitor state. In contrast, degradable polyester scaffolds such as PLG and PCL enable a robust organoid expansion and support the development of organized, pseudostratified airway epithelium. Following in vivo engraftment, organoids within these degradable matrices give rise to FOXJ1^+^ multiciliated cells and tissue architectures that closely resemble native airway structures [180]. Recent studies show that iPSC-derived alveolar organoids can be generated using synthetic hydrogel-based microwells and subsequently embedded within fully defined synthetic hydrogels. During both aggregation and embedding, these organoids actively secrete their own nascent extracellular matrix, creating a system that allows exogenous scaffold cues to be experimentally separated from cell-derived ECM signals [181]. Emerging synthetic, hybrid, and ECM-informed strategies continue to refine control over lung organoid development, bridging mechanistic insight with translational relevance.
A summary of synthetic scaffolds supporting hepatic, intestinal, pancreatic, and pulmonary differentiation from hPSCs is provided in Table 4.
6.3. Synthetic ECMs for Differentiation into Mesodermal Lineage
6.3.1. Cardiac Lineages
In an early effort to advance hPSC-derived cardiac tissue differentiation protocols, thiolated hyaluronic acid (HA) was crosslinked with PEG-diacylate and used as a culture surface for cardiomyocyte (CM) maturation. The collagen I-coated synthetic HA-hydrogel was beneficial for pre-cardiac cell maturation, as it enhanced the maturation of muscle fibers by 60% and resulted in a threefold higher expression of cardiac-specific markers than a static PA hydrogel [182]. The direct encapsulation and differentiation of hiPSCs in a 3D scaffold composed of a gelatin methacryloyl (GelMA) hydrogel generated contractile CMs with well-defined, aligned sarcomeres and developmentally appropriate temporal maturation [183]. In a recent study, the surface of PLA microparticles was functionalized with poly(poly(ethylene glycol) methacrylate) and poly[N-(3-aminopropyl)methacrylamide] brushes, and this modified ECM surface enhanced hiMSC adhesion and hiPSC-derived cardiomyocyte adhesion and contractility [184]. Another study fabricated a PDMS-based elastomer stencil with a honeycomb pattern for hiPSC seeding and differentiation toward CMs. The results showed that 9 kPa of substrate stiffness induced embryoid-like aggregation, which led to an efficient CM differentiation [185]. The geometric pattern and topology of the culture scaffold play a critical role in efficient CM differentiation and maturation. In a study, hiPSCs were encapsulated in a PEG-fibrinogen scaffold in three geometrical patterns, like disk-shaped micro-islands, squares, and rectangles, and differentiated to CMs directly in the 3D scaffold. The resulting CMs from the rectangle 3D scaffold showed less tissue heterogeneity, advanced maturation features, such as myofibrillar alignment and Z-line formation, and higher anisotropic contractile properties [186]. Nanogrid culture arrays with nanogrooved topographies, with groove widths of 300–2000 nm were made of polyurethane acrylate (PUA) coated with RGD peptides and have also been successfully used to differentiate mature CMs from hiPSCs [187]. Another study used polydimethylsiloxane (PDMS) substrates with a sub-micrometer 3D topography and cylindrical geometric patterns for imprinting hiPSC-differentiated CMs and showed further accelerated differentiation and maturation [188]. A PDMS gel matrix coated with 1% Pluronic F127 was also used to fabricate tissue molds in which hiPSC-derived CMs were cultured, and it showed a high cell viability, greater 3D cell alignment, and enhanced maturation [189]. The applications of the above-mentioned synthetic matrices are summarized in Table 5.
Among all these synthetic ECMs, PEG-based hydrogels functionalized with MMP-degradable crosslinkers and integrin-binding RGD peptides are emerging as highly promising synthetic extracellular matrix platforms for future cardiac therapies. These materials combine FDA-approved biocompatibility with precisely tunable mechanical properties that can be matched to the native myocardial stiffness range (~3–15 kPa). In addition to supporting more reproducible cardioid formation, PEG hydrogels are compatible with minimally invasive, catheter-based delivery approaches for myocardial infarction treatment and can be manufactured under GMP-compliant conditions at clinically relevant scales. These features position PEG-based ECMs as strong candidates for future translational cardiac regenerative applications [204,205].
6.3.2. Renal Lineages
Renal organoids, particularly kidney organoids differentiated from hPSCs, hold promise in regenerative medicine, as they can be used to study renal development and disease biology, support drug screening and development, and be engrafted in patients with chronic kidney disease. Although most kidney organoid generation protocols use different renal epithelial cells or primary human tubular epithelial cells as starting material, there is ongoing advancement in hPSC-differentiated kidney organoid development [206]. In parallel, the functional and physiological roles of scaffold ECMs are also being investigated in multiple studies. Renal organoids have successfully been generated in different natural, synthetic, and hybrid hydrogels, nanofiber scaffolds, and natural and decellularized ECM matrices, mostly from epithelial cells other than hPSCs [206]. The use of synthetic ECM scaffolds has been studied only recently; for example, one group used silk as a matrix for the differentiation of kidney organoids from hiPSCs [190]. The hiPSCs differentiated into renal epithelial cells that retained structural characteristics and supported the engraftment of renal organoids within the capsules of adult kidneys. Angiogenesis to the engraftment was elevated by using VEGF in the silk matrix, although the stromal cell differentiation was limited. In another study, the usage of a Thiol-ene crosslinked alginate hydrogel as an encapsulation scaffold for an hiPSC-differentiated kidney organoid resulted in an optimal expression of renal ECMs, like collagen 1A1, while maintaining all other aspects of kidney organoid physiology [191]. In a subsequent study, it was shown that a stiffer ECM (20 kPa) resulted in kidney organoids that lacked certain renal cell types, exhibited renal fibrosis signatures, and showed signs of epithelial-to-mesenchymal transition. However, when organoids were generated in a softer hydrogel (0.1 kPa), there was less deposition of ECM proteins, development of major renal segments, proximal tubule polarization, primary cilia formation, and functional maturation of the kidney organoid [192]. The results of this study confirm those of an earlier study, which showed that a softer, functionalized polyacrylamide hydrogel (1 kPa) enhanced the proliferation and maturation of kidney organoids compared with a stiffer PA hydrogel [193]. The applications of the above-mentioned synthetic matrices are consolidated in Table 5.
These studies indicate that carefully designed synthetic ECMs could enhance hPSC-differentiated kidney organoid maturation, engraftment, and functional repair, positing them as a promising platform for regenerative therapy (Table 5). The 3D ECM scaffolds were found to support the spatial organization of nephrons and promote the proper polarization of tubules and podocytes, making kidney organoids more mature with advanced renal function [207,208]. The properties like tunable stiffness, degradability, and biochemical cues in synthetic ECMs mimic the in vivo kidney microenvironment to promote vascularization, reduced fibrosis, and improved post-transplantation survival [209,210]. The kidney organoid embedded in the ECM scaffold can be engrafted under the renal capsule [211]. There, it can integrate with the host vasculature, show glomerular maturation, and exhibit limited urine filtration and solute handling, suggesting the potential for partial functional renal regeneration [212,213]. These synthetic ECMs can also act as a local drug or therapeutic agent delivery depot that modulates inflammation, fibrosis, and angiogenesis, further enhancing endogenous repair in chronic kidney injury models [214,215].
6.3.3. Vascular Smooth Muscle Cell (vSMC) Lineage
The vSMCs form a layer of non-striated contractile mural cells, which provide mechanical strength to the endothelial layer of the blood vessels [216]. These vSMCs retain significant phenotypic plasticity, as they can stay proliferative and ECM-generating or as a force-generating contractile phenotype [217,218]. The stiffness and architecture of the synthetic ECM scaffold affect the fate of vascular smooth muscle cells (vSMCs) by modulating key cellular signaling pathways that sense mechanical forces through integrins and the cytoskeleton. The patterned and stiffer synthetic ECM scaffold enhances the formation of focal adhesions, and increased actomyosin tension activates RhoA/ROCK, FAK/Src, and the subsequent nuclear localization of YAP/TAZ [219,220]. This activation leads to the enhanced proliferation, migration, and activation of synthetic gene programs [221]. In contrast, softer or elastin-like structures diminish stress fiber formation, fostering a contractile and quiescent phenotype with an elevated expression of smooth muscle markers [222,223]. Additionally, nanotopography and the alignment of fibers further influence the cytoskeletal structure and nuclear shape, affecting mechanosensitive transcriptional responses and the transition between contractile and synthetic states [224,225].
hPSC-derived vSMCs are an essential tool for studying human vascular development and disease [226]. For the consistent differentiation of vSMCs from hPSCs, the use of synthetic ECM matrices may be beneficial. In one study, hiPSC-derived mural cells were differentiated in a 3D-GelMa scaffold with varying degrees of stiffness, functionalization, and crosslinking density. In a 3D-GelMa scaffold with a higher stiffness and crosslinking density, hiPSC-derived mural cells and vSMCs exhibited a contractile phenotype. They expressed key developmental markers similar to the primary human vSMCs and mural cells [194]. Another study used Poly(N-isopropylacrylamide) (PNIPAAm)-PEG and alginate hydrogels as a 3D scaffold for vSMC differentiation from hPSCs and found that the hydrogel-based culture system generated vSMCs with a better contractile phenotype and highly expressed genes related to vasculature development and angiogenesis compared to the 2D culture system [195]. A 3D macroporous nanofibrous PLLA scaffold has also been used to differentiate hiPSCs into vSMCs, and the study found that the differentiated vSMCs expressed SMC-specific marker genes and SMC phenotypes compared to the vSMCs differentiated from hiPSC with an all-trans retinoid acid (RA) induction batch [196,197]. These studies showed that the biochemical and mechanical properties of the culture matrix and scaffold are important developmental cues for vSMC development, which can be fine-tuned in synthetic ECM matrices rather than in Matrigel (Table 5).
6.3.4. Osteoprogenitor Lineage
In recent years, numerous studies have sought to generate bone and musculoskeletal organoids from hPSCs and MSCs. The hiPSCs are an attractive source of starting material for personalized bone and cartilage regenerative therapy. For bone tissue differentiation and regeneration, different biocompatible and biodegradable polymeric scaffolds were investigated [227]. Fabricated bioactive hydrogel scaffolds are a great candidate for bone organoid generation, though they should (i) be biodegradable like natural ECM to promote cell adhesion, proliferation, and compatibility with osteogenic differentiation; (ii) be osteogenic, osteoconductive, and osteoinductive; (iii) be xeno-free, nonimmunogenic, and noncytotoxic; and (iv) have a porosity, stiffness and structural architecture compatible for osteogenesis [228,229,230]. Bone organoids generated using synthetic ECM scaffolds exhibit a stronger, more homogeneous mineralization and better regenerative potential than conventional matrices like Matrigel or inert porous scaffolds [227,231,232]. The Gelatin-Dextran hydrogel scaffold (G-PEG-Dx), GelMA-alendronate (GelMA-ALN), and DNA-Calcium phosphate hydrogels elevated ALP activity, RNX2/osteocalcin expression, and dense hydroxypartite-like deposition to enhance robust mineralization in bone organoids [233,234,235,236,237]. Tunable PEG-based synthetic hydrogels further enhance osteoblast differentiation and mineral deposition [238,239]. Upon engraftment, these organoids efficiently integrate with the host vasculature and form tubular-like bone structures more stable than rapidly degrading, mechanically weak traditional scaffolds.
In one study, iPSC-derived osteoprogenitor cells were encapsulated in a self-assembling synthetic peptide nanofiber hydrogel, PuraMatrix, and engrafted in a bone-injured rat model. The PuraMatrix was found to enhance vascularization and bone tissue regeneration at the injury site [198]. Two other studies supported this observation, in which PuraMatrix, as a hydrogel scaffold, promoted bone regeneration [199,200]. The iPSC-derived MSCs (hiPSC-MSCs) were also tested for osteogenic potential and compared with human dental pulp stem cells (hDPSCs) and bone marrow stem cells (hBMSCs). These cells were encapsulated in a calcium phosphate–alginate–fibrin hydrogel as an injectable scaffold. All stem cells showed a good proliferation and differentiation into the osteogenic lineage, although the osteogenic potential of hDPSCs was superior to that of the others [201]. Another study used HA- or collagen I-conjugated PEGdA and RGD-modified PEGdA hydrogels to differentiate hESCs into chondrogenic MSCs. The RGD-modified PEGdA hydrogel efficiently differentiated the neocartilage with basophilic extracellular matrix deposition compared to that of the other form of PEGdA hydrogel [202]. In a recent study, a 3D-bioprinted Ti6Al4V (3DTi) scaffold was used to rapidly differentiate osteocytes from hiPSCs via retinoic acid (RA)-induced osteogenesis, with a high reproducibility [203]. An osteogenesis-on-chip study fabricated a polymerized High Internal Phase Emulsion (polyHIPE) scaffold to differentiate woven bone organoids from hESC-derived mesenchymal progenitor cells (hES-MPs) by providing both biochemical and mechanical cues, such as flow and shear stress [240]. Because osteogenesis and bone regeneration are tightly coordinated by biochemical, mechanical, and structural signaling, the further development of tunable synthetic polymeric scaffolds will benefit future bone regeneration therapies (Table 5).
7. Synthetic ECM Properties Regulating Stem Cell Fate Specification
Individual ECM properties like stiffness, viscoelasticity, ligand presentation, degradability, and temporal dynamics systematically regulate stem cell fate decisions across diverse biological contexts. The integrated mechanotransduction pathways that translate biophysical cues into transcriptional programs bridge fundamental stem cell niche biology with rational material design principles.
Stiffness: Matrix stiffness serves as a primary biophysical determinant of human stem cell lineage specification, with substrates approximating tissue-specific elastic moduli (e.g., 0.1–1 kPa for brain, 8–17 kPa for muscle, 25–40 kPa for bone) [241], as illustrated in Figure 3. An integrin cytoskeletal-based mechanosensing mechanism regulates the fate and lineage specification [6,242]. The mechanotransduction cascade involves a physical linkage from ECM through integrin-based focal adhesions and cytoskeletal networks to the nucleus via LINC complexes [243]. The nuclear Lamin-A then systematically senses the tissue stiffness and directly couples the matrix mechanics to chromatin organization and gene expression [244]. The mechanotransduction pathways, like YAP/TAZ, FAK, and RhoA, regulated by tissue stiffness directly influence stem cell morphogenesis, differentiation, and organoid formation [221,241]. Synthetic ECM with an intermediate stiffness (2 kPa) was found to be conducive to efficient floor plate patterning in hiPSC-derived neural tube organoids compared to both softer (~0.7 kPa) and stiffer (~8 kPa) ECM scaffolds. Single cell transcriptional profiling and gene ontology (GO) analyses revealed that the tissue-driven ECM production, cytoskeleton organization, and planar cell polarity pathways were upregulated in the stretched neural tube organoid [245]. A PEG-based synthetic ECM with physiological stiffness (~1.3 kPa) was found to be optimal for intestinal organoid survival compared to a softer ECM (~0.2 kPa) [246]. In contrast, the softer ECM was found to be more efficiently conducive for crypt formation than the stiffer ones [247]. The YAP/Notch signaling was found to be responsive of ECM stiffness, modulating the crypt formation and differentiation of intestinal stem cells [248]. For liver organoids, PEG-based hydrogels efficiently differentiate hepatocytes in the liver organoids compared with Matrigel. The stiffer hydrogel (1.3 kPa) was found to enhance the hepatocyte functions in terms of albumin secretion and CYP450 activity, compared to that of softer hydrogels (0.3 kPa) [249]. Recent investigations revealed that stiff ECM (~40 kPa) promotes the osteogenic differentiation of MSCs by regulating energy metabolism through YAP-mediated mechanotransduction (Figure 3). YAP was found to function as a mechano-metabolic sensor that integrates mechanical cues with metabolic signaling pathways. It enhances glycolysis, oxidative phosphorylation, and mitochondrial fusion via increased mitofusin 1/2 expression while inhibiting dynamin-related protein 1 activity [250]. A follow-up study showed that the stiff matrices (25–50 kPa) favor osteogenesis in MSCs through the induction of RUNX2/ALP via 3D chromatin remodeling and merged TADs [251].
Viscoelasticity: Dynamic viscoelastic hydrogels that recapitulate the time-dependent mechanical cues of native tissues are proven to be superior scaffolds for organoid culture, surpassing the applicability of static ECM scaffolds [241,252,253]. The viscoelasticity depends on parameters like the loss modulus and rate of stress–relaxation. Hydrogels with enhanced stress relaxation promote MSC spreading, focal adhesion formation, osteogenic differentiation, and neural stem cell (NSC) astrocytic commitment through RhoA GTPase activity, actomyosin contractility, and focal adhesion signaling pathways (Figure 3) [254,255]. Viscoelastic matrices further enhance cellular plasticity by altering chromatin accessibility through nuclear lamina wrinkling, lamina-associated domain dissociation, and chromatin compaction [256]. Viscoelastic designs, such as alginate-PEG conjugates, high-molecular-weight (MW) alginate networks, and macroporous hydrogels enable stress–relaxation control through phototuning or rapid crosslink degradation [257,258]. Light-triggered PEG tethering to alginate spatially patterns relaxation rates (82–840 s half-times), guiding iPSC proliferation and morphology in 3D. Macroporous alginate (30% porosity) with BSA achieves an ultra-fast relaxation while retaining stability, enhancing nutrient diffusion for larger organoids. Such stress–relaxation promotes embryonic and branching morphogenesis via YAP/TAZ nuclear translocation and actomyosin remodeling [259]. For example, fast-relaxing PEG-alginate hydrogels (relaxation half-time ~1 min) or phototunable viscoelastic hydrogels accelerate intestinal organoid crypt–villus patterning and branching compared to elastic gels, mimicking embryonic gut ECMs [35,260]. DNA libraries with ultrahigh-MW polymers for tunable relaxation (1–100 s) were used to fabricate DNA-crosslinked matrix (DyNAtrix) hydrogel. DyNAtrix, being a dynamic viscoelastic hydrogel, supports the culture and development of human mesenchymal stromal cells, pluripotent stem cells, canine kidney cysts and human trophoblast organoids (GATA3+/GCM1+) with enhanced viability, proliferation, and morphogenesis [261]. During active cell cycle progression, the increased cellular volume activates stretch-activated ion channels, such as TRPV4 [262,263]. Rapid stress–relaxing hydrogels were found to reduce mechanical confinement and promote cell cycle progression through a TRPV4-PI3K/Akt-p27^Kip1^ signaling pathway [264]. In similar way, rapid stress relaxation promotes osteogenic differentiation from hMSCs through TRPV4 activation and the subsequent nuclear localization of RUNX2 [265].
Ligand presentation: Ligand presentation dynamics, such as ligand nanospacing, density, clustering, and temporal kinetics, systematically regulate stem cell fate decisions [266,267]. A critical nanospacing of approximately 60–70 nm determines the integrin clustering capacity and focal adhesion assembly, where larger spacings inhibits stable adhesion complex formation despite the high ligand density [266,268]. Photocontrolled supramolecular assemblies that dynamically inflate ligand nanospacing from 1.8 to 2.6 nm facilitate stem cell adhesion, mechanosensing, and differentiation both in vitro and in vivo [269]. Furthermore, slow peptide ligand (e.g., RGD peptide) dissociation kinetics enhance stem cell spreading and multi-directional migration compared to fast dissociation rates. Higher ligand clustering levels promote focal adhesion maturation and alkaline phosphatase expression in MSCs [270,271].
Matrix degradability: The matrix metalloproteinase (MMP) activity and tissue inhibitors of metalloproteinases (TIMPs) are primarily responsible for the regulation of ECM degradation to maintain tissue stiffness and viscoelasticity parameters. Matrix degradation regulates stem cell fate decisions by creating physical space for migration, releasing sequestered growth factors (e.g., VEGF, TGF-β, FGF-2), and modulating local mechanical properties within the stem cell niche [2,272]. MMP-degradable hydrogels enhance mesenchymal stem cell (MSC) migration and iPSC reprogramming efficiency. Highly degradable gels with an initial stiffness of 0.6 kPa, which is similar to embryonic tissue, yield the highest reprogramming outcomes through enhanced matrix malleability [40,262]. MMP activity enhances stress relaxation in both fibrillar collagen and synthetic matrices without altering bulk stiffness. The embedded cells sense such degradation-mediated mechanical alterations through modulated spreading and focal adhesion dynamics [273]. Engineered MMP-responsive hydrogels incorporating cell-mediated degradable crosslinkers enable dynamic ECM remodeling that recapitulates tissue development and regeneration processes. These synthetic ECMs are also compatible to controlled bioactive molecule release and angiogenic support, bridging stem cell niche biology with rational biomaterial design [274,275].
Temporal dynamics: The temporal dynamics of ECM cue presentation critically influence stem cell fate and lineage specification. The mechanosensing timescales, which can range from seconds (mechanosensor stretching, RhoA oscillations) to hours (gene expression) to weeks (permanent differentiation), systematically regulate stem cell fate decisions and lineage commitments [255,276]. Dynamic hydrogels with reversible stiffness revealed that neural stem cells exhibit a 24 h mechanosensitive window during which ECM stiffness profoundly impacts neurogenic commitment via substrate-dependent RhoA activation [255,277]. Skeletal muscle stem cells develop mechanical memory within the first three days of culture on stiff substrates, characterized by the RhoA-mediated loss of proliferative capacity that persists even after subsequent softening, demonstrating the temporal integration of mechanical memory [278]. Engineered hydrogels with dynamically tunable viscoelasticity through reversible crosslinks enable temporal control. Fast-dissociating crosslinks permit rapid force-induced network reorganization and enhanced mechanosensing compared to slow-dissociating crosslinks. Such binding kinetics critically determine stem cell spreading, differentiation, and chromatin accessibility across minute-to-day temporal regimes [279]. Synthetic ECMs incorporating phototunable, magnetoactive, or enzyme-responsive dynamic crosslinks now enable spatiotemporal stiffness pulsing that recapitulates developmental ECM remodeling, bridging stem cell and fate plasticity.
Modern engineered hydrogel design strategies leverage these principles with independently tunable stiffness, viscoelasticity, degradability, and bioactive ligand presentation. This enables a systematic high-throughput investigation of combinatorial ECM parameters that regulate stem cell fate, accelerated mesenchymal-to-epithelial transitions, and enhanced epigenetic remodeling during reprogramming [3,40]. These integrated ECM properties collectively define specialized microenvironments where heterogeneous cell populations, basement membrane components, and dynamic ECM remodeling converge to maintain stemness or direct differentiation through spatiotemporally orchestrated biophysical and biochemical signaling (Figure 3).
8. Recent Advances and Emerging Trends in Synthetic ECM Development
Advances in synthetic ECM engineering have fundamentally reshaped human stem cell and organoid culture by overcoming the variability and translational limitations of animal-derived materials such as Matrigel [35]. Fully defined synthetic matrices now enable reproducible control over mechanical properties, degradability, and biochemical signaling, allowing the in vitro reconstruction of key aspects of native tissue organization [41]. These developments, together with progress in stem cell biology and 3D bioprinting, have enabled the generation of organoids with improved structural fidelity, functional maturation, and clinical relevance. In recent times, a myriad of new approaches is being developed to advance synthetic ECM fabrication logics based on the knowledge of mechanical, physiochemical, architectural, and biochemical influence on human organoids and hPSC-derived cell cultures. The polymeric backbone of synthetic ECM is now generated through advanced polymerization chemistries like Michael additions, Thiol-ene chemistry, Diels–Alder (DA) reactions, inverse electron demand Diels–Alder (IEDDA) reactions, copper-catalyzed azide–alkyne cycloadditions (CuAAC), strain-promoted azide–alkyne cycloaddition (SPAAC) reactions, Sequential Click Reactions, and other bio-orthogonal chemistries [280,281]. Several key innate properties like the ECM stiffness, dynamic viscoelasticity, stress–relaxation through controlled biodegradation, and bioactive ligand presentation in a spatio-temporal fashion are now considered during synthetic ECM fabrication.
Emerging ECM strategies increasingly emphasize dynamic and cell-responsive signaling. Peptide amphiphile nanofibers functionalized with laminin-derived IKVAV motifs support the long-term maturation of human iPSC-derived neurons. Molecular mobility within the scaffold enhanced signal transduction beyond static ligand presentation [282]. Synthetic matrices also reduce organoid heterogeneity by enabling precise control over stiffness and composition, improving reproducibility in high-throughput and imaging-based applications. Consistent with this, neural organoids assembled on synthetic PEG hydrogels exhibit an enhanced cellular diversity, reduced batch variability, and functional immune responsiveness compared to Matrigel-based systems [283].
The integration of synthetic ECMs with advanced 3D bioprinting technologies has further advanced organoid engineering by enabling a spatially defined control over cell placement, matrix composition, and architecture [284]. Extrusion-, inkjet-, and light-based bioprinting approaches have been used to generate complex tissue geometries, including crypt–villus intestinal structures and cardiovascular-like models [285]. In particular, sacrificial writing into functional tissue (SWIFT) enables the incorporation of perfusable vascular channels into dense, cell-laden constructs, overcoming diffusion limitations and supporting long-term tissue viability and differentiation [286,287].
Artificial intelligence and machine learning (AI/ML) are increasingly reshaping synthetic extracellular matrix (ECM) design for organoid scaffolds by enabling data-driven optimization that replaces traditional trial-and-error approaches [288]. ML-guided models can predict and tune hydrogel composition, crosslinking chemistry, and mechanical properties, accelerating the development of ECM-mimetic bioinks with improved reproducibility, scalability, and printability for 3D bioprinting [289,290,291]. In parallel, AI tools streamline the analysis of high-dimensional biological data, including single-cell transcriptomics and multi-omics integration, providing deeper insight into organoid growth and differentiation [288]. The integration of machine learning with computer vision further enables real-time monitoring and adaptive control during bioprinting, improving fabrication robustness and consistency [292]. Emerging concepts such as ‘Organoid Intelligence’ extend these advances by coupling sensor-integrated brain organoids with ML-based training frameworks, highlighting future applications in biocomputing [37]. Despite significant progress, challenges remain related to data availability, model generalizability, experimental validation, and the integration of biological knowledge into computational pipelines [293].
9. Conclusions and Future Perspectives
Concerns about interpreting cell culture data generated using Matrigel were raised nearly three decades ago, yet its use as a 2D and 3D scaffold has continued to expand across cell biology and stem cell research [19]. The early reliance on Matrigel reflected the limited availability of alternative ECMs and the technical and resource barriers associated with developing synthetic matrices. Recent interdisciplinary advances, increased collaboration, and a growing demand for stem cell-based disease modeling have alleviated many of these challenges. Consequently, synthetic ECMs have emerged as robust alternatives, offering tunable biochemical and mechanical properties, controlled degradability, and defined biofunctionalization for reproducible hPSC culture, differentiation, and organoid generation. In parallel, improved embryoid body formation methods—such as bioreactors, rotary suspension culture, and microwell aggregation—now enable more homogeneous starting populations, enhancing lineage-specific organoid development [294,295].
PEG-based hydrogels have emerged as a leading synthetic platform due to their high cytocompatibility, resistance to nonspecific protein adsorption, and modular tunability. In particular, PEG-4MAL hydrogels, crosslinked with protease-degradable peptides and functionalized with adhesive motifs such as RGD, support the robust and reproducible generation of human intestinal organoids from pluripotent stem cells without Matrigel [41,42]. These matrices not only sustain organoid growth and differentiation but also enable minimally invasive delivery and engraftment in vivo, accelerating intestinal wound repair in murine models and highlighting their translational potential. Beyond PEG, fully synthetic PIC hydrogels with tunable charge and thermo-responsive behavior further expand the design space for defined organoid matrices [253]. Matrix mechanics play a central role in stem cell fate and morphogenesis. Soft PEG hydrogels with precisely tuned elastic moduli support the three-dimensional encapsulation of human pluripotent stem cells, preserving pluripotency and promoting uniform, lumenized aggregate formation, with intermediate stiffness favoring epiblast-like organization and a defined lower threshold required for cell viability [296]. In parallel, decellularized ECMs derived from native tissues retain tissue-specific biochemical and mechanical cues that are difficult to recapitulate synthetically. Pancreatic and kidney dECMs have been shown to enhance endocrine lineage assembly, vascularization, and the maturation of corresponding organoids, addressing persistent challenges in organoid size and functional integration [297,298]. Recent work further demonstrates that organoid formation can be driven by epithelial cell-secreted ECM, with endogenous laminin deposition establishing a de novo stem cell niche independent of exogenous matrix supplementation [299]. This finding provides a potential route toward eliminating xenogeneic materials and addressing regulatory barriers. Looking forward, the integration of synthetic ECM platforms with artificial intelligence and machine learning is poised to accelerate the optimization of bioink composition, printing parameters, and tissue architecture [300]. Key challenges remain, including scalable bioprinting, cost reduction, improved vascularization, and standardized protocols that balance synthetic control with biological complexity [301]. In this context, synthetic ECMs are best viewed as complementary tools that broaden the experimental landscape rather than as one-size-fits-all replacements for native or tissue-derived matrices. Apparently, no single engineered material can replicate the full biochemical richness and structural complexity found across all tissue niches. Instead, synthetic systems excel in contexts where researchers need tight control—such as isolating specific variables, probing mechanistic questions, or developing platforms suitable for scalable, translational manufacturing. Conversely, natural and decellularized ECMs may still be the preferred choice when robust, tissue-specific signaling or highly physiological remodeling dynamics are essential.
Recently, the U.S. Food and Drug Administration (FDA) provided the ‘Roadmap to Reducing Animal Testing in Preclinical Safety Studies’ (https://www.fda.gov/media/186092/download?attachment, accessed on 9 February 2026) and guidelines to embrace human-relevant disease models like organoids and organ-on-chip models under ‘New Approach Methodologies’ (NAMs) [302]. This brings unprecedented interest and demand for the development of organoid-based disease modeling platforms for fundamental research in disease biology and future drug development, across both public and private research entities. Alongside efforts to improve organoid culture reagents and recombinant growth factors/morphogens, significant efforts are underway to develop synthetic matrix scaffolds as alternatives to Matrigel. These efforts led to the establishment of new biotechnology startups, including Regenity, Advanced Biomatrix, Matricel GmbH, The Well Bioscience, Tissue Regenix, BioLamina AB, Luna-Gel™, and MIMETAS, as well as market leaders such as Thermo Scientific, Corning, and Sigma-Aldrich. Many of the synthetic matrices developed by these companies are described in this review. It is essential to continue developing bioengineered scaffolds through an open collaboration among stem cell biologists, materials scientists, clinicians, engineers, and biotechnology industry stakeholders. This will streamline production and commercial availability for end users in stem cell therapy, regenerative medicine, and the pharmaceutical industry.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Frantz C. Stewart K.M. Weaver V.M. The extracellular matrix at a glance J. Cell Sci.20101234195420010.1242/jcs.02382021123617 PMC 2995612 · doi ↗ · pubmed ↗
- 2Gattazzo F. Urciuolo A. Bonaldo P. Extracellular matrix: A dynamic microenvironment for stem cell niche Biochim. Biophys. Acta 201418402506251910.1016/j.bbagen.2014.01.01024418517 PMC 4081568 · doi ↗ · pubmed ↗
- 3Watt F.M. Huck W.T. Role of the extracellular matrix in regulating stem cell fate Nat. Rev. Mol. Cell Biol.20131446747310.1038/nrm 362023839578 · doi ↗ · pubmed ↗
- 4Wang H. Luo X. Leighton J. Extracellular Matrix and Integrins in Embryonic Stem Cell Differentiation Biochem. Insights 20158152110.4137/BCI.S 3037726462244 PMC 4589090 · doi ↗ · pubmed ↗
- 5Wang L. Zheng F. Song R. Zhuang L. Yang M. Suo J. Li L. Integrins in the Regulation of Mesenchymal Stem Cell Differentiation by Mechanical Signals Stem Cell Rev. Rep.20221812614110.1007/s 12015-021-10260-534536203 · doi ↗ · pubmed ↗
- 6Engler A.J. Sen S. Sweeney H.L. Discher D.E. Matrix elasticity directs stem cell lineage specification Cell 200612667768910.1016/j.cell.2006.06.04416923388 · doi ↗ · pubmed ↗
- 7Brielle S. Bavli D. Motzik A. Kan-Tor Y. Sun X. Kozulin C. Avni B. Ram O. Buxboim A. Delineating the heterogeneity of matrix-directed differentiation toward soft and stiff tissue lineages via single-cell profiling Proc. Natl. Acad. Sci. USA 2021118 e 201632211810.1073/pnas.201632211833941688 PMC 8126831 · doi ↗ · pubmed ↗
- 8Xie Y.-H. Tang C.-Q. Huang Z.-Z. Zhou S.-C. Yang Y.-W. Yin Z. Heng B.C. Chen W.-S. Chen X. Shen W.-L. Extracellular Matrix Remodeling in Stem Cell Culture: A Potential Target for Regulating Stem Cell Function Tissue Eng. Part B Rev.20222854255410.1089/ten.teb.2021.006634082581 · doi ↗ · pubmed ↗