A New Species of the Genus Scincella Mittleman, 1950 (Squamata: Scincidae) from the Hengduan Mountains, Sichuan Province, Western China
Fan Liu, Hongli Pu, Songwen Tan, Jiahao Chen, Bing Lyu, Guocheng Shu, Yayong Wu, Bingjun Dong, Peng Guo

TL;DR
A new skink species, Scincella heishuiensis, was discovered in the Hengduan Mountains of China, increasing the known diversity of the genus.
Contribution
The formal description of a new Scincella species based on molecular and morphological data from the Hengduan Mountains.
Findings
Scincella heishuiensis is a new species distinguished by unique morphological and genetic traits.
The new species forms a sister group with S. wangyuezhaoi and is genetically distinct by 5.45% in 16S rRNA.
The discovery increases the number of Scincella species in China to 17.
Abstract
The skink genus Scincella, a member of the family Scincidae, is characterized by a wide distribution and conservative morphology. This study integrates molecular and morphological data to conduct phylogenetic analyses and comparative morphology of Scincella population in the Hengduan Mountains. Our findings include the discovery of a new species and the clarification of its phylogenetic relationships within the genus. Significant morphological differences were observed between specimens collected from Heishui County and other known Scincella species. When combined with phylogenetic analysis results, these specimens were identified as a new species, formally described as Scincella heishuiensis sp. nov. Phylogenetic reconstruction reveals that Scincella heishuiensis sp. nov. forms a sister group with S. wangyuezhaoi while exhibiting distinct morphological differentiation from all other…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
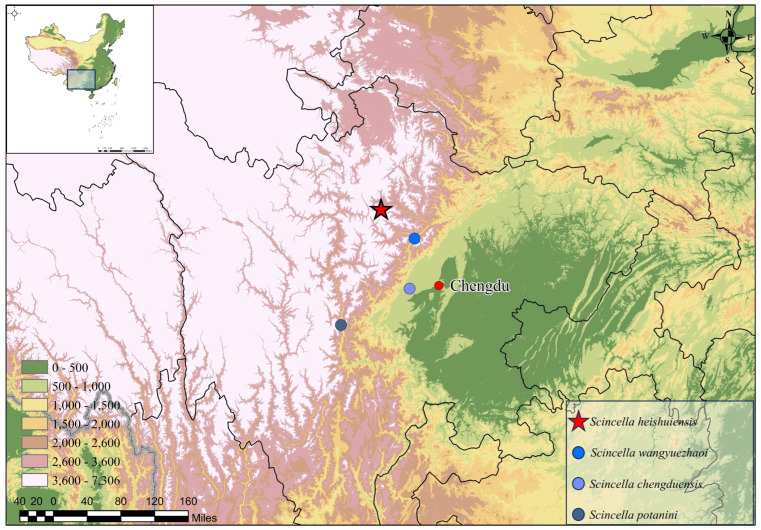 Figure 1
Figure 1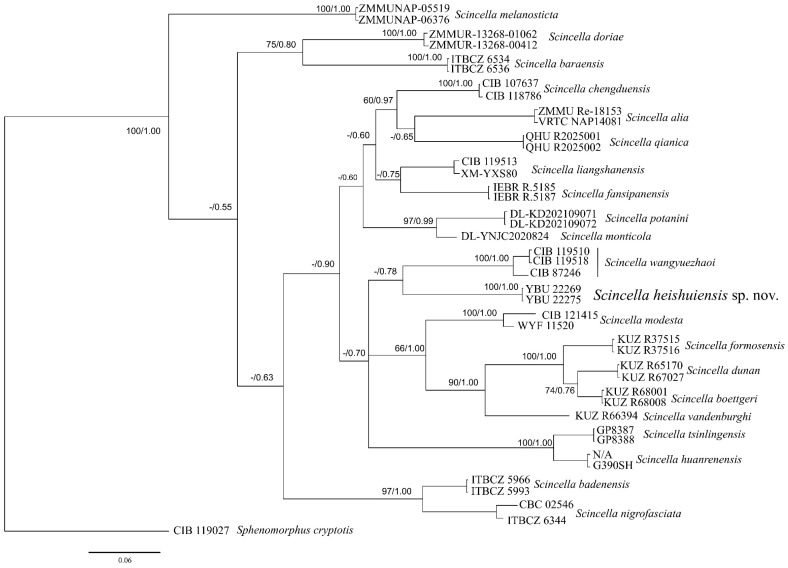 Figure 2
Figure 2 Figure 3
Figure 3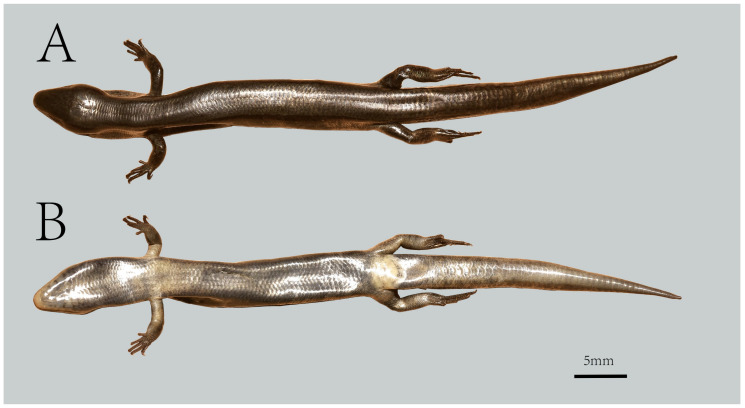 Figure 4
Figure 4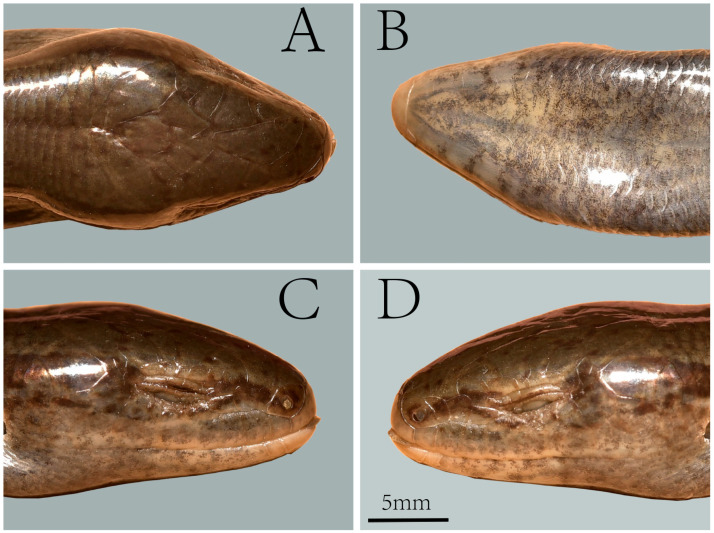 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7- —Second Tibetan Plateau Scientific Expedition and Research (STEP) program
- —Scientific Research Foundation of Yibin University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAmphibian and Reptile Biology · Animal Behavior and Reproduction · Venomous Animal Envenomation and Studies
1. Introduction
The Hengduan Mountains (HDM), located on the eastern Qinghai–Xizang Plateau (QXP), are characterized by dramatic elevational gradients that create distinct vertical ecological zones, harboring exceptionally high biodiversity. This region has been recognized as the original and evolutionary center for several taxonomic groups [1]. Recent taxonomic investigations have revealed numerous new species in the area, including snakes [2,3,4], lizards [5,6], and frogs [7]. The increasing description of new taxa in this region indicates a significant underestimation of its biodiversity [8].
The genus Scincella Mittleman, 1950, which belongs to the family Scincidae, was originally described based on the type species S. lateralis. Wang et al. divided Chinese species into three groups based on morphological comparisons and recognized 14 species in China [8]. Che et al. moved three Scincella species inhabiting Xizang, China to Asymblepharus [9], which was established by Eremtschenko [10], and would later be treated as a subjective junior synonym of Ablepharus [11]. In recent years, several new species have been described sequentially in Western China [5,6,12]. Currently, the genus Scincella comprises 50 species and exhibits a broad distribution across East Asia, Southeast Asia, and North America [13].
In China, the HDM represents the primary distribution area for Scincella species, with 10 species recorded in this region: S. schmidti, S. liangshanensis, S. wangyuezhaoi, S. potanini, S. monticola, S. tsinlingensis, S. modesta, S. doriae, S. barbouri and S. reevesii [5]. However, recent studies have identified several potential taxonomic issues [5,6], indicating the possibility of the presence of a substantial number of cryptic species in the area.
During recent field surveys in the HDM, we collected several Scincella specimens, including a distinctive population from Heishui County, Sichuan, China. Morphological analyses revealed significant differences from congeners, which were further supported by molecular phylogenetic evidence showing substantial divergence from known species. Based on integrated morphological and molecular phylogenetic analyses, we formally describe these specimens as a new species. Subsequent phylogenetic analysis based on three mitochondrial DNA genes (COI, 12S rRNA, and 16S rRNA) confirmed that this population from Heishui County, Sichuan Province, belongs to the genus Scincella. The analysis also revealed that it forms a distinct sister lineage to S. wangyuezhaoi from Wenchuan County, Sichuan Province. Further examination of the scalation, coloration, and morphology of the Heishui population, along with comparisons to other species, demonstrated clear morphological distinctions from both its sister species and existing species within the genus Scincella. Therefore, we describe the skinks from Heishui County in the northern HDM of China as a new species of Scincella.
2. Materials and Methods
Specimen collection. The specimens of Scincella sp. were collected from Heishui County, Sichuan, China (Figure 1). After the livers were taken, the specimens were fixed in a 10% formaldehyde solution for three days and then transferred to 80% ethanol for long-term preservation. The liver samples were kept at 85% ethanol in −20°. All specimens and samples are deposited at Yibin University (YBU).
2.1. Morphology Analyses
The two unidentified specimens were morphologically examined and recorded following Zhao et al. and Nguyen et al. [14,15,16,17,18]. Body dimensions were taken with a digital caliper to the nearest 1 mm. Symmetric mensural head characters were taken on the right side of the head only, unless this was damaged, in which case they were done on the left. Meristic characters were recorded on both sides, and the average was used in the analysis. All data were collected by the senior author to avoid inter-observer bias [18,19].
The characters measured and recorded and their abbreviations are as follows: snout–vent length (SVL, distance from tip of snout to posterior edge of vent); tail length (TAL, distance from posterior margin of vent to tip of tail); axilla–groin distance (AGD, distance between posterior edge of forelimb insertion and anterior edge of hindlimb insertion); forelimb length (FLL, measured from forelimb insertion to tip of finger IV or longest digit); hind-limb length (HLL, measured from hind-limb insertion to tip of toe IV or longest digit); toe IV length (T4L, measured from the most basal part to tip of toe IV); finger IV length (F4L, measured from the most basal part to tip of finger IV); midbody scale-row count (MBSR, number of longitudinal scale rows measured around widest point of midbody); dorsal scale rows between dorsolateral stripes (DBR, number of dorsal scale rows at midbody between dark dorsolateral stripes, following Inger; enlarged, differentiated nuchal count (NU, X pairs or absence); paravertebral scale-row count (PVSR, number of scale rows counted between parietals and the just posterior margin of hindlimbs); ventral scale-row count (VSR, number of scale rows counted between gulars and precloacal); superciliary count (SC, left/right); supralabial count (SL, left/right); infralabial count (IfL, left/right); superciliary count (SC, left/right); supraocular count (SO, left/right); enlarged temporal count (TEM, left/right); number of enlarged, undivided lamellae beneath finger IV (F4S, left/right); number of enlarged, undivided lamellae beneath toe IV (T4S, left/right); frontoparietal in contact with each other (FP); prefrontal in contact with each other (PF); chin-shields: paired large scales behind mental or postmentals; gulars: many minor scales in the center of the ventral head (number of scale rows counted between the first scale behind the chin-shields and the middle of the forelimb); appearance of dorsolateral stripes (ADLS).
2.2. Molecular Analyses
Total DNA was extracted from the liver tissue of two samples using the M5 HiPer Universal DNA Mini Kit (Mei5 Biotechnology Co., Ltd., Beijing, China). Three mitochondrial DNA fragments were amplified, 12S ribosomal RNA (12S) using primers L1091-F and H1478-R (5′–3′: AAACTGGGATTAGATACCCCACTAT/GAGGGTGACGGGCGGTGTGT), 16S ribosomal RNA (16S) using primers 16SL-F and 16SL-R (5′–3′: TGTTTACCAAAAACATAGCCTTTAGC/TAGATAGAAACCGACCTGGATT), cytochrome c oxidase subunit I (COI) using primers Chmf4 and Chmr4 (5′–3′: TYTCWACWAAYCAYAAAGAYATCGG/ACYTCRGGRTGRCCRAARAATCA) [20]. PCR cycling parameters followed the original protocols from these studies. PCR products were purified using commercial kits prior to sequencing. Bidirectional sequencing was performed by Sangon Biotech (Shanghai, China). Phylogenetic analyses incorporated 21 Scincella species and one outgroup (Sphenomorphus cryptotis) [21], with sequences retrieved from GenBank (Table 1). Sequence editing was conducted in Lasergene v15.1, followed by Clustal W alignment and quality verification in MEGA v11.0 [22]. Raw nucleotide sequences were manually validated using SeqMan v.7.1.0.44 [23].
Prior to phylogenetic analyses, the optimal substitution model for each gene partition was selected using PartitionFinder 2.1.1 [24] under the Akaike Information Criterion. Phylogenetic trees were constructed using Maximum Likelihood (ML) and Bayesian Inference (BI) methods. BI analyses were conducted in MrBayes 3.2.2 [25], using three heated chains and one cold chain, with three independent runs of 10 million generations each (sampling every 2000 generations). The initial 25% of samples were discarded as burn-in, achieving a potential scale reduction factor (PSRF) < 0.005 across all runs. ML analyses were performed in IQ-TREE [26] with 1000 standard bootstrap replicates (BS). Bayesian posterior probabilities (BS) ≥ 0.95 and PP ≥ 95 are considered to indicate strong support for tree nodes [27]. Phylogenetic trees were visualized and annotated in FigTree 1.4.3 [28]. Given the difficulties in collecting samples of Scincidae species, coupled with the numerous discoveries of new species within the genus Scincella in recent years, and considering that this specimen morphologically most closely resembles the recently described S. wangyuezhaoi, this study exclusively selected species from the genus Scincella to construct the phylogenetic tree.
The uncorrected p-distance between species was calculated for 16S and COI sequences in MEGA v11.0 with 1000 bootstrap replications [22] respectively.
3. Results
3.1. Morphology Examination
The newly collected specimens exhibit morphological affinities with Scincella by sharing key scalation features, including well-developed eyelids, absence of supranasal, and one pair of preanal. Meanwhile, they can be distinguished from congeners by the following unique characters: four dorsal scale rows between dark dorsolateral stripes at midbody, specific midbody-scale-row count, presence or absence pattern of postnasal scales, and loreal scale number.
3.2. Molecular Phylogeny
The final aligned dataset consists of 1462 nucleotide positions (490 bp from 16S RNA, 353 bp from 12S RNA, 619 bp from COI). The best-fit nucleotide substitution model (GTR + I + G) was determined for the combined dataset. All novel sequences generated here have been deposited in GenBank (Table 1).
Both BI and ML analyses yielded largely consistent phylogenetic topologies, with minor discrepancies in node support values observed at certain positions (Figure 2). All recognized Scincella species, including the two newly collected specimens, formed a strongly supported monophyletic group (100 BS and 1.0 PP). The new specimens constituted a distinct clade with high support indices (BS = 100, PP = 0.99), exhibiting a sister-group relationship with the clade containing three S. wangyuezhaoi individuals.
Based on 16S rRNA sequences, interspecific genetic distances among Scincella species ranged from 2.19% (S. tsinlingensis vs. S. huanrenensis) to 10.18% (S. huanrenensis vs. S. gemmingeri). Based on COI sequences, the distances ranged from 7.6% (S. dunan vs. S. boettgeri) to 23.1% (S. melanosticta vs. S. vandenburghi). The newly proposed taxon (comprising the two novel samples) exhibited genetic distances of 5.45% (16S) and 15.2% (COI) relative to S. wangyuezhaoi (Table 2 and Table 3).
Integrating morphological comparison and molecular phylogeny, we confirm that the specimens collected from Heishui County, Sichuan Province, China, represent a distinct species. We formally describe it herein.
3.3. Taxonomic Account
Scincella heishuiensis sp. nov.
Figure 3, Figure 4, Figure 5 and Figure 6.
Scincella wangyuezhaoi Jia, Ren, Jiang & Li, 2023 [5].
Holotype. YBU 22269, an adult male, collected from Mt. San’ao, Heishui County, Aba Tibetan and Qiang Autonomous Prefecture, Sichuan Province, China (32.02° N, 102.92° E, 2900 m a.s.l), in June 2022.
Paratype (n = 1). YBU 22275, an adult male, the same locality and date as the holotype.
Diagnosis. Scincella heishuiensis sp. nov. exhibits the following characteristics which can distinguish from its congeners: (1) four dorsal scale rows between dorsolateral stripes; (2) 5–7 superciliary; (3) 28 midbody scale rows; (4) 24–25 gulars; (5) during the breeding season, the ventral scales and preanal exhibit a reddish-brown color; and (6) a black lateral stripe extends from behind the eye to the posterior third of the tail near the cloaca.
Description of the holotype. YBU 22269, an adult male. The body is slender and of medium size, with a snout–vent length (SVL) of 40.90 mm, and TAL of 39.25 mm. The neck is indistinct from the head. The axilla–groin distance (AGD) 29.65 mm, (AGD/SVL 0.72). The tail is short and thick, with a length (TAL) of 39.25 mm (TAL/SVL 0.96). The forelimb length (FLL) is 8.20 mm, and the hind limb length (HLL) is 12.87 mm. Forelimb digit lengths: IV > III > II > V > I; Hindlimb digit lengths: IV > III > II > V > I. Fingers are slender, F4L 2.58 mm, T4L 4.96 mm. Subdigital lamellae single, arranged in an overlapping series: F4S 8/8, T4S 13/13.
The rostral is blunt and rounded. The supranasal is absent. Two prefrontals, rhomboid-shaped, separated by the frontonasal, with the two scales contacting only at one point. Two frontoparietals, irregularly pentagonal, adjacent and in contact with each other. One parietal, the largest on the dorsal head, pentagonal in shape, with a width approximately two-thirds of its length; its lateral margins contact only two supraoculars and one prefrontal. Six supraoculars, with second being the largest and sixth the smallest. Four supraorbitals, with the second being the largest. The nasal is oval, the nostril positioned at its center. Postnasals absent. The tympanic cavity recessed, but the tympanum not exposed; scales surrounding the tympanum are slightly smaller than the ventrolateral scales. Two loreals, square-shaped, the first being the largest and contacting second and third supralabials. Supralabials 7-7; the first contacts the nasal, and the second largest. Infralabials 6-5, slender and elongated, with the first being the smallest. Four pairs of cervical scales, all elongated (approximately three times longer than wide), the first pair the largest. Temporals 4 (3 + 1), the dorsal largest and contacts the parietal. The lower eyelid has a distinct transparent disk (window), and both eyelids are accompanied by minute scales. The gular scales are relatively small and off-white in color. Three pairs of chin shields, the first pair in medial contact, the second pair separated by a small scale, and the third pair separated by a small scale. The mental scale is broad, occupying the anterior edge of the lower jaw; there is one larger postmental; the eyes are medium-sized, with a flat, circular transparent palpebral window in the center of the lower eyelid; the ear opening is oval, larger than the palpebral window but smaller than the eye diameter, and the anterior edge of the ear opening lacks flap-like protrusions; the tympanic membrane is recessed. The gulars number 25.
The body scales are smooth. The dorsal scales are rhomboidal, arranged in straight, imbricate rows, significantly larger than ventral and lateral scales; DBR 4 + 2 (1/2); MBSR 28. Ventral scales are diamond-shaped, arranged in straight, non-imbricate rows; PVSR 69; VSR 67. The tips of the fingers and toes (excluding claws) do not contact each other.
Coloration. The dorsal head and tail regions display a distinct bronze sheen, with a dark bronze patch extending between the nostrils and eyes. All dorsal head scales bear black spots. The dorsum features scattered black spots, while the regenerated tail exhibits a characteristic copper–black coloration. The anterior body and tail bear irregular dark dorsal stripes composed of melanin-rich scales, including prominent ventrolateral stripes. A continuous lateral stripe extends from the posterior orbit to the tail base. The ear opening is positioned obliquely below the eye, near the cervical junction. Ventral coloration includes a silver-gray throat and uniform reddish-brown abdomen devoid of patterning.
In preservation, the head and dorsum adopt a uniform brown hue with intensified lateral dark stripes. The dorsum and venter retain smooth integument texture, while the tail is lateral and ventral surfaces develop dark brown to black spotting. Throat and belly are bright gray, limbs are bronze, and the caudal region near the cloaca is pale gray.
3.4. Description of the Paratype and Variation
The paratype specimen YBU 22275 exhibits morphological similarity to the holotype across multiple diagnostic characters including SL, MBSR and DBR. A detailed comparison of the specimens is listed in Table 4.
Etymology. The species is named after its type locality in Heishui County, Sichuan Province, China. We suggested the common names “黑水滑蜥” in Chinese and “Heishui ground skink” in English for this species.
Distribution and ecology. Currently, Scincella heishuiensis sp. nov. has only been found in Heishui County, Sichuan Province, China. The type specimens were collected in mid-June 2022. The new species is active on rock piles in woodland areas with exposed rocks and scattered low shrubs. It is commonly found under rocks or moss on sunny days (Figure 7). This species is sympatric with Rhabdophis nuchalis, Protobothrops jerdonii, and Gloydius swild.
3.5. Comparisons with Congeners
Scincella heishuiensis sp. nov. can be distinguished morphologically from closely related homologous species in western Sichuan; the diagnostic characteristics are summarized in Table 4.
In phylogenetic trees, the new species is closely related to S. wangyuezhaoi, S. modesta, S. tsiningensis, and S. huanrenensis. Besides significant genetic divergence, the new species is morphologically distinguished from its congeners by a combination of following characters. Scincella heishuiensis sp. nov. can be distinguished from S. wangyuezhaoi and S. modesta by the significantly diagnostic character dorsal scale rows between dorsolateral stripes (DBR), 4 (vs. 6); and from S. tsiningensis and S. huanrenensis by fewer paravertebral scale rows (PVSR), 60–69 (vs. 70–90 in S. tsiningensis and 66–84 in S. huanrenensis). It can also be distinguished from S. huanrenensis by its ventral scale rows (VSR), 64–67 (vs. 52–60) [29,30].
The new species differs from S. doriae and S. formosensis by having fewer DBR (4 vs. 6). It is different from S. monticola, S. qianica, and S. doriae by exhibiting 28 MBSR (vs. 23–24 in S. monticola, 26 in S. qianica, 30–32 in S. doriae) [30,31,32,33,34]. The new species differs from S. doriae and S. schmidti by having enlarged undivided lamellae beneath toe IV 13–14 (vs. 11–12 in S. schmidti; 16–18 in S. doriae). The new species differs from S. barbouri in PVSR 60–69 (vs. 70–79 in S. barbouri). The new species can be further differentiated from S. alia by VSR 64–67 (vs. 44–52). Scincella heishuiensis sp. nov. can be differentiated from S. schmidti and S. potanini by fewer enlarged undivided lamellae beneath toe IV (8–10 vs. 11–12 in S. schmidti and 10–13 in S. potanini) [16,32,35,36]. The new species differs from S. barbourin in nuchals 2–3 (vs. 4–5). The new species can be distinguished from S. przewalskii by having a distinct black lateral stripe, whereas S. przewalskii has a pale green one. The new species has a dirty white belly, which distinguishes it from S. przewalskii (slate gray) and S. tsinlingensis (bluish gray). Scincella heishuiensis sp. nov. differs from S. modesta by having different anterior dorsolateral scale arrangement (straight vs. wavy in). The new species differs from S. przewalskii by having four supraoculars (vs. three) [37,38,39,40,41] (Table 4).
4. Discussion
With the description of a new cryptic species from western China, based on genetic and morphological evidence, the genus Scincella now comprises 51 recognized species, with 17 occurring in China. Molecular phylogenetic analyses revealed that the new species is sister to S. wangyuezhaoi (posterior probability [PP] = 0.78), a species recently described from Wenchuan County, Sichuan Province. The two species exhibit a genetic divergence of 5.45% (p-distance based on the 16S fragment). Geographically, their type localities are separated by only 100 km. Despite their proximity, they show significant genetic divergence and substantial morphological differentiation.
The Hengduan Mountains, one of the world’s youngest mountain systems, serve as a biodiversity hotspot, harboring more than half of China’s Scincella species, including S. schmidti, S. liangshanensis, S. wangyuezhaoi, S. potanini, S. monticola, S. tsinlingensis, S. doriae, and S. reevesii [40,41,42]. The region’s high habitat heterogeneity and riverine isolation have significantly contributed to speciation processes. As a widespread group of lizards in China, Scincella species exhibit complex systematic and evolutionary patterns. For instance, S. tsinlingensis and S. huanrenensis share a close phylogenetic relationship but occupy entirely distinct and geographically separated ranges. Similar challenges arise in other species within the genus, such as the S. potanini–S. monticola species complex. Second, there are two different reproductive models within this genus. Populations in high-altitude regions are predominantly lecithotrophic viviparity, while those in low-altitude areas are primarily oviparous [35,43]. Additionally, our research suggests the likely presence of a substantial number of cryptic species in this area [44]. Given these patterns, a key question emerges: what mechanisms drive such rich species diversity within this relatively small region? To address this, comprehensive sampling and multi-locus genetic data—particularly genomic data—are essential for unraveling the evolutionary history and speciation processes of these organisms.
5. Conclusions
We describe a new skink species, Scincella heishuiensis sp. nov., from the HDM in China, based on two specimens from San’ao Snow Mountain, Heishui County, Sichuan. The species is formally delineated using integrated morphological and genetic evidence, underscoring its distinctiveness from its congeners. This discovery not only contributes to the documented biodiversity of HDM, a globally recognized hotspot, but also suggests that the region may harbor additional cryptic diversity. The findings prompt further exploration of the drivers of Scincella diversification within this complex mountainous terrain and provide a scientific foundation for enhancing local conservation strategies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Guo P. Zhu F. Liu Q. Wang P. Che J. Nguyen T.Q. Out of the Hengduan Mountains: Molecular phylogeny and historical biogeography of the Asian water snake genus Trimerodytes (Squamata: Colubridae)Mol. Phylogenetics Evol.202015210692710.1016/j.ympev.2020.10692732771547 · doi ↗ · pubmed ↗
- 2Liu Q. Zhong G.H. Wang P. Liu Y. Guo P. A new species of the genus Hebius (Squamata: Colubridae) from Sichuan, China Zootaxa 2018448338539410.11646/zootaxa.4483.2.1030313795 · doi ↗ · pubmed ↗
- 3Guo P. Liu Q. Zhu F. Liu Q. A new member of the genus Sinonatrix (Serpentes: Colubridae) from western China Zootaxa 2019462353554410.11646/zootaxa.4623.3.531716249 · doi ↗ · pubmed ↗
- 4Shi J.S. Liu J.C. Giri R. Owens J.B. Santra V. Kuttalam S. Selvan M. Guo K.J. Malhotra A. Molecular phylogenetic analysis of the genus Gloydius (Squamata, Viperidae, Crotalinae), with descriptions of two new alpine species from Qinghai-Tibet Plateau, China Zoo Keys 202110618710810.3897/zookeys.1061.7042034707454 PMC 8505391 · doi ↗ · pubmed ↗
- 5Jia R.W. Gao Z.Y. Huang J.J. Ren J.L. Jiang K. Li D. Li J.T. A new species of the genus Scincella Mittleman, 1950 (Squamata: Scincidae) from Sichuan Province, Southwest China, with a diagnostic key of Scincella species in China Asian Herpetol. Res.2023142440
- 6Jia R.W. Gao Z.Y. Wu D.H. Ren J.L. Jiang D.C. Wu W. A new species of the genus Scincella Mittleman, 1950 (Squamata: Scincidae) from Sichuan Province, Southwest China Asian Herpetol. Res.20241511512910.3724/ahr.2095-0357.2024.0016 · doi ↗
- 7Ji F.R. Shi S. Ma S. Shen C. Chang L.M. Jiang J.P. A New Species of the Genus Nanorana Günther, 1896 (Anura: Dicroglossidae) from Hengduan Mountains of China Asian Herpetol. Res.20231430031810.3724/ahr.2095-0357.2023.0025 · doi ↗
- 8Wang K. Che J. Lin S. Deepak V. Aniruddha D.R. Jiang K. Jin J. Chen H. Siler C.D. Multilocus phylogeny and revised classification for mountain dragons of the genus Japalura (Reptilia: Agamidae: Draconinae)Zool. J. Linn. Soc.201918524626710.1093/zoolinnean/zly 034 · doi ↗