Alimentary Tract Anatomy and Morphology in Early Adult Mediterranean Killifish Aphanius fasciatus (Valenciennes, 1821)
Maria Cristina Guerrera, Lidia Pansera, Marialuisa Aragona, Kamel Mhalhel, Mauro Cavallaro, Maria Levanti, Rosaria Laurà, Giuseppe Montalbano, Francesco Abbate, Antonino Germanà

TL;DR
This paper describes the anatomy of the digestive system in Mediterranean killifish, revealing a specialized intestinal chamber and sexual dimorphism in the anal region.
Contribution
The first detailed anatomical study of the digestive tract in Aphanius fasciatus, including a novel description of a food-processing intestinal chamber and sex-based structural differences.
Findings
A specialized anterior intestinal chamber functions for food storage and preparation without gastric glands.
Sexual dimorphism is observed in the anal region, with males having a more toned and folded structure compared to females.
The oropharyngeal tract and esophagus lead to a dilated intestinal region with mucosal folds and mucus-secreting epithelium.
Abstract
The Mediterranean killifish, Aphanius fasciatus, is a small fish that is able to survive in variable salinity and temperature, making it an important bioindicator in coastal ecosystems. Despite its ecological relevance, little is known about its anatomy. This study provides the first detailed examination of its digestive tract, revealing a specialized anterior intestinal chamber that stores and processes food before digestion, and differences between males and females in the anal region. These findings offer valuable insight into the biology of this species and support its use as a reliable experimental model for studies in anatomy, physiology, and environmental monitoring. Background/Aims: The Mediterranean killifish, Aphanius fasciatus (Valenciennes, 1821), is a small euryhaline and eurytherm cyprinodont. While its ecology and role as a bioindicator are well known, its anatomy…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1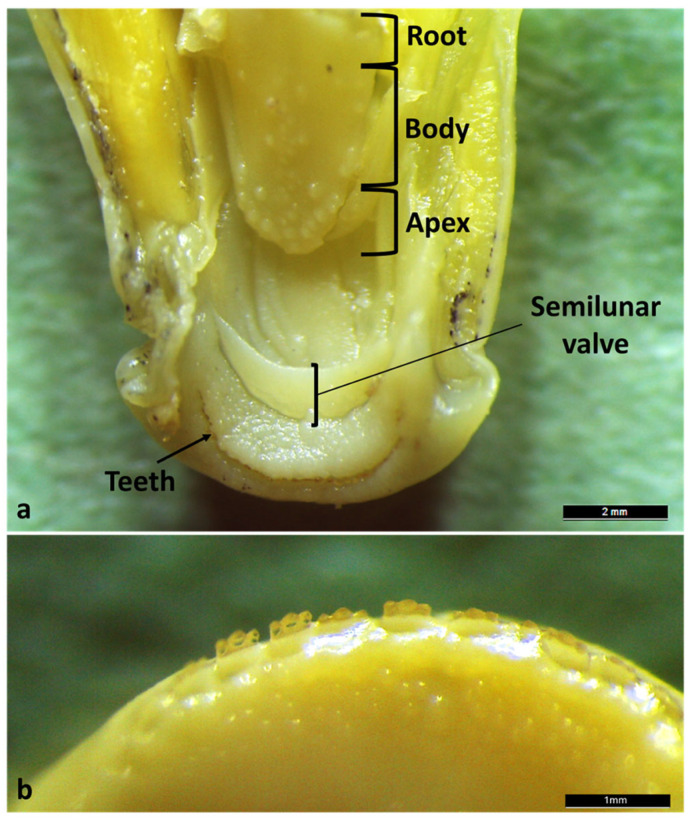 Figure 2
Figure 2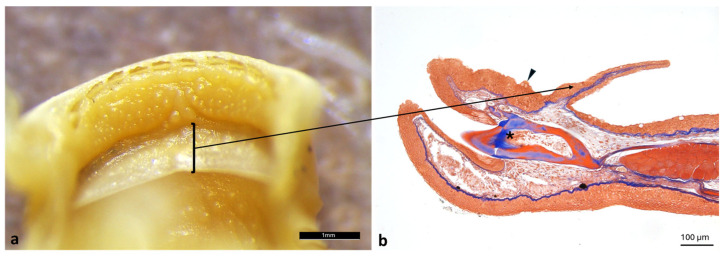 Figure 3
Figure 3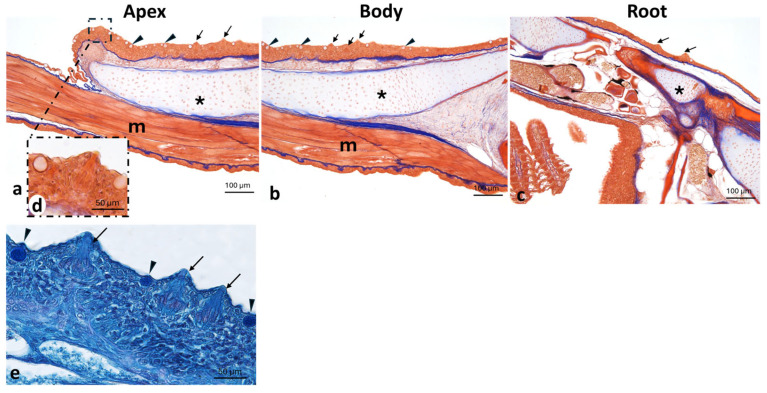 Figure 4
Figure 4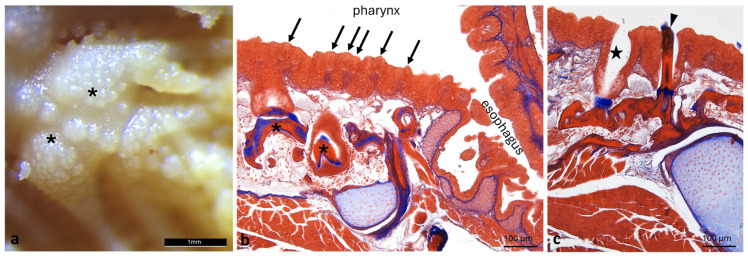 Figure 5
Figure 5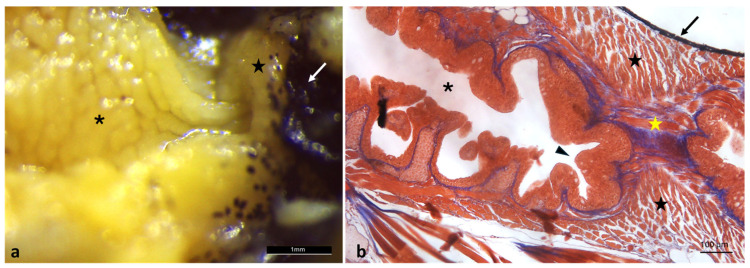 Figure 6
Figure 6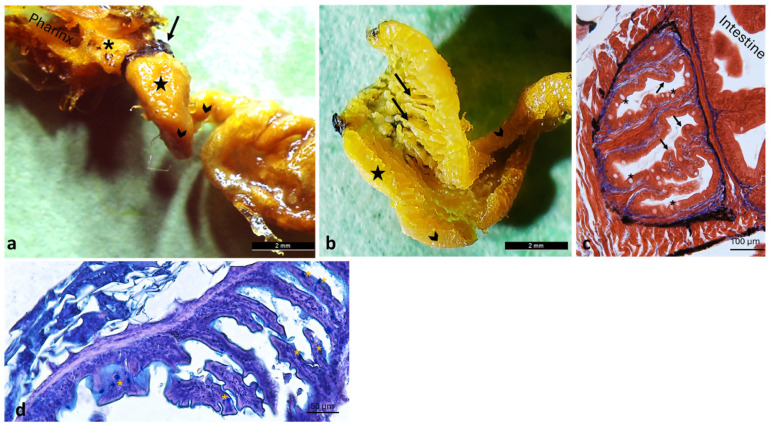 Figure 7
Figure 7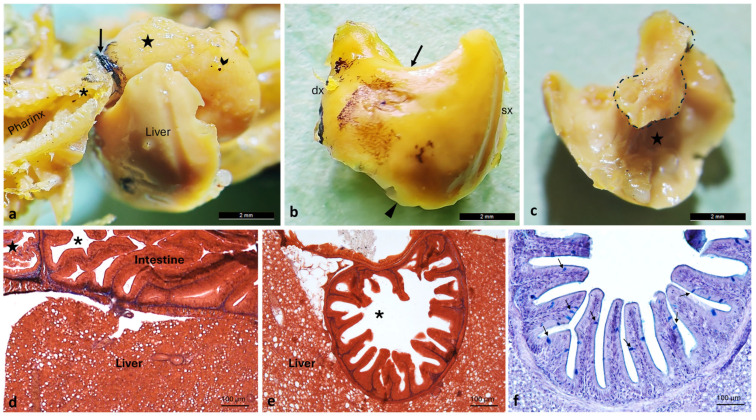 Figure 8
Figure 8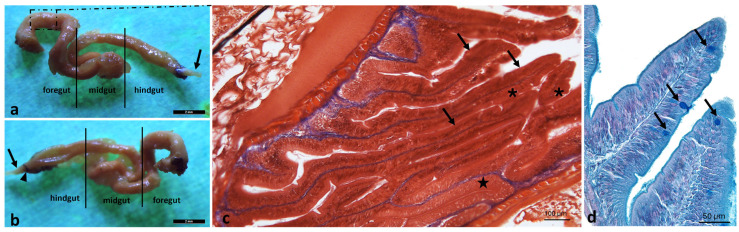 Figure 9
Figure 9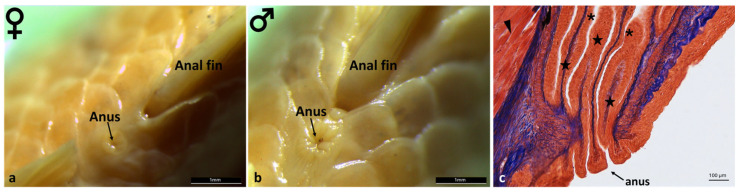 Figure 10
Figure 10- —PNRR—Missione 4, Componente 2, Investimento 1.1—Bando Prin 2022 PNRR—Decreto Direttoriale
- —Progetto Finanziato dalla Unione Europea-Next Generation EU_ PRIN_PNRR_ 2022_missione 4 investimento 1.1.
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsComparative Animal Anatomy Studies · Fish biology, ecology, and behavior · Fish Biology and Ecology Studies
1. Introduction
Aphanius fasciatus (Valenciennes, 1821), commonly known as the Mediterranean killifish, is a small teleost fish belonging to the order Cyprinodontiformes (Actinopterygii: Cyprinodontiformes). As a euryhaline and eurythermic species, it tolerates a wide range of physicochemical conditions, including temperature (5–39 °C) and salinity (0–180 g/L), allowing it to inhabit transitional environments such as estuaries and coastal lagoons [1,2,3,4]—shallow systems located between land and sea—along the Mediterranean region [5,6,7,8]. These habitats experience marked daily and seasonal fluctuations in physical and chemical properties, driven by tidal dynamics and solar heating [6,7,9]. A. fasciatus is typically associated with brackish or hypersaline water bodies, particularly salt marshes and lagoons, where environmental parameters show substantial variation [10,11,12]. The species completes its entire life cycle under these variable environmental conditions, reaching 2–9 cm in total length, reaching sexual maturity within the first year, with a lifespan of up to seven years [4,12,13,14,15]. Reproduction is characterized by strong male–male competition for access to females [16] and pronounced sexual dimorphism [16]. Females are generally larger than males but display less developed and less colorful caudal, anal, and dorsal fins [17]. In contrast, males exhibit bright body coloration and iridescent yellow pigmentation on the dorsal and anal fins, traits involved in intrasexual aggressive interactions [16,18]. The species exhibits rapid population dynamics due to its short generation time and high reproductive rate [8,14,16,18]. It is gregarious and largely sedentary, with limited dispersal capacity throughout the year. Seasonal movements are restricted, with populations likely shifting to deeper areas during winter and dispersing to nearby sites only during heavy rainfall events that flood coastal lagoons [12]. Consequently, gene flow among populations is limited, often resulting in marked genetic isolation even among geographically close groups [4,7,11,19]. Owing to these ecological characteristics, A. fasciatus has been widely employed as an indicator species for environmental quality in transitional waters, using molecular, cellular, and physiological biomarkers to assess contaminant effects [1,5,6,7,9,16,20,21,22]. In Italian transitional environments, only a limited number of fish species complete their entire life cycle within these habitats, among which the Mediterranean killifish A. fasciatus represents a key example [23]. Accordingly, Italian legislation recognizes this species as an indicator of complex ecosystems requiring specific conservation measures, in accordance with Council Directive 92/43/EEC. Despite the extensive ecological knowledge and its established role as a sentinel species, a comprehensive anatomical and morphological description of the gastrointestinal tract of A. fasciatus is still lacking. The present work addresses this gap, also revealing functional adaptations of the digestive system related to feeding habits and environmental variability. These fundings provide novel morphological data that support functional interpretations and comparisons with other teleost models used in biomedical and ecotoxicological research.
2. Materials and Methods
2.1. Sample Collection
A detailed macroscopic and microscopic morphological investigation was conducted on Aphanius fasciatus specimens in order to describe the species’ distinctive anatomical features and to highlight potential similarities with other experimental teleost models. Ten early adult A. fasciatus (five males and five females, with a body size ranging between 30 and 40 mm; see Figure 1) were collected in the Vendicari Nature Reserve (Siracusa, southeastern Sicily, Italy; 36.7907° N, 15.0896° E).
At the time of collection, ambient air temperature ranged approximately from 18 to 22 °C, relative humidity was moderate to high, and weather conditions were mild with no significant rainfall. The measured water physico-chemical parameters (Table 1) indicate a highly saline marine–brackish environment, well oxygenated and characterized by moderate turbidity, conditions consistent with habitats typically occupied by Aphanius fasciatus.
2.2. Light Microscopy
The specimens were fixed in Bouin solution for 12–18 h [24]; after removal of the excess fixative, they were maintained in ethanol 70° [25].
Macroscopic investigations were performed using a Leica stereomicroscope (M205C) equipped with a Leica IC80 HD digital camera (Leica, Milan, Italy) [25]. For histological purposes, the sample were dehydrated through a graded ethanol series (70–100°), cleared in xylene, and embedded in paraffin (Bio-Optica Milano S.p.A., Milan, Italy; code 08-7910) [26,27,28]. Paraffin blocks were sectioned into 7 μm serial slices using a Leica RM2135 microtome, mounted on gelatin-coated slides, and dried for 24 h [29,30,31,32]. The sections were subsequently deparaffinized, rehydrated, and stained with Masson’s trichrome using aniline blue (Cat.# 04-010802, Bio-Optica S.p.A., Milan, Italy) [33], and Alcian Blue pH 2,5-PAS (Bio-optica Milano S.p.A, Milan, Italy, CAT. # 04–163802) staining [27,34]. Finally, stained preparations were examined, and photomicrographs acquired with Leica Application Suite LAS v4.7 software using a Leica DMRB light microscope equipped with a Leica DFC7000 T camera (Leica Microsystems GmbH, Wetzlar, Germany) [35].
3. Results
The study examined the complete digestive tract of Aphanius fasciatus using longitudinal and transverse sections obtained from the entire specimen. To expose the tongue, the jaws were disarticulated. The tongue appears as a thickening of the floor of the mouth although it offers three regions for description: apex, body and root. Only the apex appears free (Figure 2a). On both jaws, the presence of a semilunar valve (Figure 2a) and a single row of small, tricuspid, incisive-like teeth were observed (Figure 2b).
Histological analysis of the semilunar valve revealed a thin connective core lined on both sides by a simple squamous epithelium interspersed with mucus-secreting goblet cells. The connective tissue is mainly composed of dense collagen fibers (Figure 3).
Both the apex (Figure 4a) and the body (Figure 4b) are covered by mucosa lined with a non-keratinized stratified squamous epithelium containing abundant goblet cells and scattered taste buds, which progressively becomes thinner toward the root (Figure 4c). The submucosa, initially dense, progressively becomes looser and overlies a well-developed cushion of hyaline cartilage, which is continuous with the proper floor of the mouth, characterized by a prominent layer of striated muscle tissue (Figure 4).
Under the stereomicroscope, the surface of the pharynx does not appear smooth but features dense, irregularly distributed small elevations or protuberances (Figure 5a), which under the light microscope are identified as taste buds (Figure 5b). In the submucosa, pharyngeal teeth at different stages of eruption are present (Figure 5b,c). Some of them reach the surface (Figure 5c), and some dental alveoli are empty (Figure 5c).
The pharyngeal region continues posteriorly into the esophagus without a distinct structural boundary, leading to the cardiac sphincter. This area is marked by a distinctly pigmented ring (Figure 6a), while histological sections show circular muscle fibers indicative of sphincteric activity. The esophagus exhibits a mucosa raised in folds and lined by stratified squamous epithelium (Figure 6).
Beyond the sphincter lies a balloon-shaped dilated portion, structurally adapted to facilitate the passage of food toward the intestine. This dilated region, referred to as the “pre-intestinal chamber”, exhibits a mucosa organized into tall folds and a simple columnar epithelium interspersed with mucus-secreting goblet cells (Figure 7).
The liver appears large, unilobed, and compact, its cranial margin is convex, whereas the dorsal margin is concave due to the impression made by the dilated intestinal portion (Figure 8). The liver partially surrounds the intestine through a small papillary process extending from the right margin, which curves caudomedially to partially encircle the proximal intestinal segment (Figure 8).
The intestinal length is approximately four times the body length. It has a tubular shape and consists of the foregut, midgut, and hindgut; however, it lacks distinct regional segmentations, reflecting an omnivorous diet. The mucosa forms internal longitudinal folds that increase the absorptive surface area. The height and number of these folds gradually decrease in the caudal direction. A simple columnar epithelium interspersed with mucus-secreting goblet cells was observed (Figure 9).
The morphological analysis of the anal region of A. fasciatus revealed marked sexual dimorphism. In females, the anus appears relaxed, with mucosal folds that are faint or barely discernible (Figure 10a). In males, by contrast, the anus exhibits a tonic morphology, with tall and well-defined mucosal folds (Figure 10b). Microscopic analysis of the rectal tract shows the mucosa raised into folds lined by stratified squamous epithelium, supported by a dense connective tissue framework. These folds extend into the anus. In addition, a well-developed skeletal muscle layer provides the sphincteric function (Figure 10c).
4. Discussion
Several studies have demonstrated the suitability of the Mediterranean killifish (Aphanius fasciatus) as a bioindicator species for environmental contamination in transitional waters, based on its molecular, cellular, and physiological responses to pollutants in estuaries and lagoons ecosystems [1,5,7,20]. Overall, these studies confirm the species’ sensitivity to environmental stressors and support its relevance for ecotoxicological monitoring. Within this framework, the present study provides the first comprehensive anatomical and morphological characterization of the gastrointestinal tract of A. fasciatus, from the oral cavity to the anal region, thereby establishing a structural framework for future functional, comparative, and toxicological investigations.
In the oral cavity, the presence of semilunar valves delimiting a depression identifiable as a gustatory fossa was observed. This structure, characterized by a high density of taste buds, likely functions as a pre-taste chamber. Histologically, the valves are composed of a simple squamous epithelium supported by connective tissue rich in dense collagen fibers, suggesting a structure capable of providing both stability and flexibility during feeding. Similar semilunar valves have been described in other teleosts, including Danio rerio, where upper and lower valves are prominent and lined by stratified epithelium containing numerous mucous and rodlet cells, indicating a conserved morphological feature involved in oral cavity partitioning and food manipulation [36]. Variations in valve morphology among teleost species are generally associated with different feeding strategies, such as suction feeding or substrate grazing [37,38]. In annual killifishes (Nothobranchius spp.), the oral region exhibits a simplified organization consistent with rapid life-history traits and opportunistic feeding, with reduced specialization compared with euryhaline species such as A. fasciatus [39]. In contrast, basal teleosts, including Heterotis niloticus, retain more archaic oropharyngeal structures, linked to omnivorous–detritivorous diets [25]. Compared with these species, A. fasciatus exhibits a more generalized oral configuration, supporting dietary flexibility in variable brackish environments.
The presence of small tricuspid, incisor-like teeth on both jaws represents a further distinctive feature of the anterior oral region of A. fasciatus. These teeth are continuously replaced, reflecting the polyphyodont condition typical of many teleosts [40]. Their morphology suggests a functional role in grasping and fragmenting food prior to swallowing. Comparable tricuspid dentitions have been reported in herbivorous and omnivorous teleosts, including Ctenopharyngodon idella, in which specialized pharyngeal teeth and a keratinized chewing pad are involved in processing of plant-derived material [41,42]. In contrast, carnivorous teleosts typically possess large conical or caniniform teeth adapted for prey capture and retention [43]. In A. fasciatus, the tricuspid morphology increases the functional surface area of the dentition, enhancing efficiency in cutting or grinding food prior to swallowing and indicating functional convergence with other euryhaline teleosts, including Poecilia spp. [44,45]. Conversely, D. rerio lacks oral teeth and relies exclusively on pharyngeal dentition located on the fifth ceratobranchials for suction-based feeding and crushing [40,46,47,48]. H. niloticus exhibits small conical (villiform) teeth arranged around a rigid, bone-supported “bony tongue,” forming a triturating surface adapted to omnivorous–detritivorous diets [25,49,50]. The tongue of A. fasciatus appears as a thickening of the floor of the mouth and is organized into three regions: the apex (partially free), the body, and the root. This tripartite organization parallels that described in Oncorhynchus mykiss and Salmo salar, where the tongue is associated with fungiform-like papillae, taste buds, and cartilaginous elements involved in food manipulation [35,51]. Similar epithelial features have been described in other teleosts, although the density and distribution of mucous cells and taste buds vary according to dietary habits [52,53,54]. Similarly, grass carp exhibits abundant taste buds on palatal and lingual folds supported by connective and muscular cores, reflecting adaptation to herbivorous feeding [39]. Compared with Nothobranchius furzeri, A. fasciatus shows a more differentiated lingual organization, likely related to its opportunistic feeding strategy in highly variable brackish environments [55,56]. In contrast, H. niloticus has a thick, cartilaginous “bony tongue” associated with tubular organs rich in mucous cells and taste buds, supporting extensive intraoral processing [25]. In adult D. rerio, the tongue is poorly regionalized, with a thinner epithelium and fewer mucus cells, reflecting suction-based feeding [57,58]. In this species, taste buds are widely distributed throughout the oral cavity, reducing the need for lingual specialization [57,59,60]. In A. fasciatus, the presence of scattered taste buds and goblet cells suggests an active role of the tongue in both sensory evaluation and lubrication of food. The hyaline cartilage pad underlying the lingual mucosa provides structural support that is absent in D. rerio, enhancing mobility during swallowing [58]. Accordingly, food transport in zebrafish relies mainly on hydrodynamic forces and branchial mechanisms rather than on active lingual movements [61,62,63,64,65]. The lingual mucosa of A. fasciatus, composed of non-keratinized stratified squamous epithelium with goblet cells and taste buds at an intermediate density, is consistent with an omnivorous feeding habit when compared with the higher densities reported in Poecilia spp. [44,45]. This organization, supported by a hyaline cartilage pad continuous with the striated musculature of the oral floor, parallels that described in grass carp and Oncorhynchus spp. [35,66,67], emphasizing functional versatility handling over specialized grazing in Poecilia spp. [13,15,44,45].
In line with the sensory and mechanical role of the oral structures, the pharyngeal region represents the functional continuation of food evaluation and processing. The pharyngeal tract of A. fasciatus is short and lined by stratified squamous epithelium with interspersed taste buds and well-developed pharyngeal teeth at different stages of eruption. The coexistence of sensory and masticatory components suggests a dual functional role, combining gustatory detection with mechanical food processing. Similar arrangements have been reported in other omnivorous teleosts, in which pharyngeal dentition contributes to crushing or grinding, while taste buds support food selection [37,68]. The presence of both oral and pharyngeal incisor-like teeth in A. fasciatus reflects adaptation to a mixed diet including algae, invertebrates, and larvae. Comparable patterns have been described in O. niloticus and Ctenopharyngodon idella [39,41,69], whereas strictly carnivorous species such as Oncorhynchus spp. possess larger conical pharyngeal teeth primarily adapted for prey retention rather than grinding [35]. In D. rerio, pharyngeal teeth are smaller and less robust, reflecting a diet dominated by microinvertebrates and detritus, while taste buds are widely distributed for chemical detection [58]. Nothobranchius spp. exhibit short pharyngeal tracts with simple dentition suited to rapid prey ingestion in ephemeral habitats [70]. Species of Poecilia spp. display moderate pharyngeal dentition adapted to both invertebrate- and plant-based diets [71], whereas H. niloticus shows robust pharyngeal teeth and a muscular tract associated with filter-feeding and food processing [25,49,50,72]. Overall, the integration of oral teeth, pharyngeal dentition, and taste buds enables A. fasciatus to exploit a wide range of trophic resources, linking its pharyngeal morphology to omnivorous feeding strategies across teleosts, from microprey specialists to dietary generalists and herbivores [73,74]. Once food processing within the oropharyngeal tract is completed, the digestive system transitions into the esophagus, a key segment responsible for controlled transport toward the intestinal tract.
Macroscopically, the cardiac region of the esophagus in A. fasciatus appears as a distinctly pigmented ring. Histological analysis identified this structure as composed of circular muscle fibers consistent with sphincteric function, regulating food transit from the oropharyngeal tract toward the digestive tract and preventing regurgitation. Well-developed esophageal sphincters are commonly reported in teleost fishes [75,76,77]. Distal to this region, the esophagus expands into a dilated balloon-shaped segment that likely serves as a temporary storage chamber prior to intestinal transit. Comparable esophageal dilations have been described in omnivorous species such as O. niloticus and Ctenopharyngodon idella, where they accommodate variable food volumes and facilitate peristalsis [39,75,76,78,79]. In contrast, D. rerio exhibits limited cardiac region dilation [80,81], while Nothobranchius spp. possess short esophagi with simple sphincters [82,83]. In Poecilia spp., sphincters are modest and dilation is reduced [84,85,86], whereas H. niloticus shows prominent sphincters and a markedly dilated proximal esophagus supporting bulk storage associated with filter-feeding [25,49,50,87]. The pronounced pigmentation of the cardiac ring in A. fasciatus may contribute to protection against mechanical stress or ultraviolet exposure, as suggested in other teleosts [25,75,88,89]. Together, the presence of both a functional sphincter and esophageal dilation represents an adaptive feature typical of omnivorous fishes, with structural variation reflecting different ecological strategies [76,90,91,92,93]. The anterior region of the digestive tract of A. fasciatus is characterized by a dilated chamber with tall mucosal folds lined by a simple columnar epithelium rich in mucin-secreting goblet cells, substantially increasing the absorptive surface area and facilitating pre-intestinal processing of ingested material [94,95,96,97,98]. The absence of true gastric glands indicates that this region functions as a predigestive tract, promoting mixing and partial absorption rather than acid or peptic digestion [99,100,101]. Agastric conditions represent a convergent evolutionary strategy among several teleost lineages [102,103]. In this context, the structure observed in A. fasciatus, here referred to as a “pre-intestinal chamber,” appears to optimize nutrient processing prior to enzymatic digestion. Comparable anterior intestinal enlargements, commonly described as “intestinal bulbs,” are reported in stomachless species such as D. rerio and other cypriniforms, where they represent major sites of enzyme activity and absorption [99,104,105,106]. Similar functional adaptations occur in Nothobranchius spp. [70,82,107,108] and Poecilia spp. [109,110,111], whereas H. niloticus, despite possessing a true stomach, exhibits proximal digestive segments showing functional similarities with predigestive chambers [25,87,112,113].
The functional efficiency of this predigestive region is closely supported by the associated hepatic system, which plays a key role in digestion and metabolic regulation.
Within the coelomic cavity, A. fasciatus present a large, compact, and unilobed liver. The organ displays a dorsal concavity produced by the impression of the proximal intestine, which is partially enveloped by a small hepatic process extending caudomedially from the right margin. This anatomical configuration highlights the central role of the liver in coordinating digestion and metabolism downstream of the predigestive tract [68,114,115]. Similar arrangements occur in Nothobranchius spp., in which the liver is closely associated with the short intestine, supporting rapid metabolic processing in ephemeral habitats [82,107,116]. In omnivorous Poecilia spp., the liver supports the metabolism of both animal and plant-derived nutrients [117,118,119]. H. niloticus features a large, lobed liver that supports bulk food storage and metabolic processing [25,87,112]. In agastric D. rerio, the liver is closely associated with the anterior intestinal bulb, contributing to bile secretion and metabolic regulation in the absence of a stomach [80,104,120]. Thus, A. fasciatus relies on an integrated hepatic system to support digestion through bile production and metabolic regulation, paralleling teleosts with agastric or partially agastric digestive tracts. Following hepatic secretion and metabolic processing, the digestive tract continues into the intestine, which represents the main site of digestion and nutrient absorption in this agastric species.
The intestine in A. fasciatus is tubular and lacks distinct regional segmentation, consistent with an omnivorous feeding regime. The mucosa forms longitudinal folds lined by a simple columnar epithelium with interspersed mucin-secreting goblet cells [13,80,102,114,115,121,122]. In the absence of a stomach, enteric digestion, supported by early pancreatic and biliary secretions, replaces gastric processing [106,114,123]. The relative length of the intestine exceeds that typically observed in predatory species, supporting the digestion of a mixed diet that includes plant material, detritus, and invertebrates [92,124]. Folds enhance digestive efficiency without increasing organ volume, paralleling simplified digestive models such as those of D. rerio and Poecilia reticulata, which rely on decentralized digestive functions [118]. Poecilia spp. shows long, folded intestine compensating for absent stomachs, processing plant and animal matter [85,125]. In contrast, Nothobranchius spp. exhibit shorter intestinal tracts adapted for rapid ingestion of invertebrates [82,107,108,126]. H. niloticus exhibits a long, highly folded intestine integrating digestive secretions to support bulky omnivory [25,49,50,72]. Thus, A. fasciatus relies on extended intestinal absorption and digestive secretions distributed along the intestinal tract, consistent with omnivory in agastric teleosts.
Finally, the morphological analysis of the anal region in A. fasciatus revealed marked sexual dimorphism. Females exhibited a lax anus with barely evident or absent mucosal folds, reflecting reduced sphincter tonicity that likely facilitates egg passage during oviposition in externally fertilizing species [15,16,127,128]. In contrast, males showed a tonic anus with prominent mucosal folds, indicating enhanced sphincter musculature development that allows controlled milt release, enhancing reproductive efficiency [15,128,129]. Comparable sexual dimorphism is observed in other externally fertilizing teleosts. In D. rerio, males display distinctive anal fin pigmentation and genital pore morphology associated with spawning behavior [80,130,131,132,133]. In P. reticulata, males possess a gonopodium derived from the anal fin, enabling directed sperm transfer, whereas females retain a more flexible oviposition region [134,135,136]. Males of Nothobranchius spp. exhibit hypertrophied anal musculature adapted for rapid milt expulsion, while females maintain compliant anal regions suitable for brief spawning events [82]. In H. niloticus, sexual dimorphism of the anal region is less pronounced but still reflects differential sphincter development linked to gamete control and egg passage [25,136]. Overall, this dimorphism in A. fasciatus underscores the specialization of the anal region for sex-specific reproductive roles, correlating mucosal architecture and sphincter tonicity with fertilization efficiency across teleosts.
5. Conclusions
This study provides the first comprehensive anatomical and histological description of the digestive tract of Aphanius fasciatus, offering a detailed overview of its structural organization and adaptive features.
The integration of macroscopic and microscopic observations reveals a digestive system well suited to a broad range of environmental conditions, consistent with the species’ remarkable euryhaline nature. The structural traits observed suggest functional specializations that enhance digestive efficiency and physiological plasticity under fluctuating salinity and nutrient availability. From an ecological perspective, these findings shed light on how A. fasciatus maintains digestive performance in habitats exposed to anthropogenic stressors, thus reinforcing its recognized resilience in transitional and brackish environments. The morphological data contribute to comparative frameworks within teleost digestive biology, highlighting both conserved features shared with cyprinodontiforms and distinctive traits that reflect the adaptive evolution of this taxon. Moreover, the detailed dataset provided here offers a solid foundation for future functional, molecular, and ecotoxicological investigations, enabling meaningful comparisons with other teleost model species and helping to distinguish conserved digestive traits from lineage- and habitat-specific adaptations. From a translational viewpoint, the integration of ecological, anatomical, and ecotoxicological information in A. fasciatus supports its role as a bridge species between environmental risk assessment and experimental pathophysiology, particularly for investigating pollutant-induced alterations of digestive structure and function.
Given its tolerance to environmental stress and its ease of maintenance under laboratory conditions, this species represents a suitable model for evaluating the effects of contaminants on digestive physiology, as well as for comparative anatomical studies among euryhaline teleosts. Collectively, the evidence presented confirms the scientific relevance of A. fasciatus as both a bioindicator species and as an experimental model organism in biology research. Moreover, this study focused on early adult subjects, so the anatomical patterns reported should be interpreted within this context. Considering that data from other developmental stages are currently missing in bibliography, these results lay the groundwork for future comparable studies on the developmental changes in the gastrointestinal tract in A. fasciatus.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lionetto M.G. Zonno V. Schiavone R. Giordano M.E. Barca A. Belmonte G. Verri T. The Mediterranean Killifish Aphanius fasciatus (Valenciennes, 1821) (Teleostei: Cyprinodontidae) as a Sentinel Species for Protection of the Quality of Transitional Water Environments: Literature, Insights, and Perspectives Water 202315272110.3390/w 15152721 · doi ↗
- 2Valdesalici S. Langeneck J. Barbieri M. Castelli A. Maltagliati F. Distribution of natural populations of the killifish Aphanius fasciatus (Valenciennes, 1821) (Teleostei: Cyprinodontidae) in Italy: Past and current status, and future trends Ital. J. Zool.20158221222310.1080/11250003.2014.1003418 · doi ↗
- 3Edwards S.L. Marshall W.S. 1—Principles and Patterns of Osmoregulation and Euryhalinity in Fishes Fish Physiology Mc Cormick S.D. Farrell A.P. Brauner C.J. Academic Press London, UK 2012 Volume 32144
- 4Pappalardo A.M. Calogero G.S. Šanda R. Giuga M. Ferrito V. Evidence for Selection on Mitochondrial OXPHOS Genes in the Mediterranean Killifish Aphanius fasciatus Valenciennes, 1821 Biology 20241321210.3390/biology 1304021238666824 PMC 11048645 · doi ↗ · pubmed ↗
- 5Abbassi A. Kessabi K. Casado M. Pérez-Jiménez A. Trenzado C.E. Rufino-Palomares E.E. Guerbej H. Piña B. Messaoudi I. Adverse effects of environmentally relevant concentration of microplastics on gill epithelium permeability in the euryhaline Mediterranean killifish Aphanius fasciatus Chemosphere 202436814378310.1016/j.chemosphere.2024.14378339577801 · doi ↗ · pubmed ↗
- 6Kessabi K. Annabi A. Hassine A.I.H. Bazin I. Mnif W. Said K. Messaoudi I. Possible chemical causes of skeletal deformities in natural populations of Aphanius fasciatus collected from the Tunisian coast Chemosphere 2013902683268910.1016/j.chemosphere.2012.11.04723260252 · doi ↗ · pubmed ↗
- 7Kessabi K. Navarro A. Casado M. Saïd K. Messaoudi I. Piña B. Evaluation of environmental impact on natural populations of the Mediterranean killifish Aphanius fasciatus by quantitative RNA biomarkers Mar. Environ. Res.20107032733310.1016/j.marenvres.2010.06.00520638715 · doi ↗ · pubmed ↗
- 8Pappalardo A.M. Gonzalez E.G. Tigano C. Doadrio I. Ferrito V. Comparative pattern of genetic structure in two Mediterranean killifishes Aphanius fasciatus and Aphanius iberus inferred from both mitochondrial and nuclear data J. Fish Biol.201587698710.1111/jfb.1269326010329 · doi ↗ · pubmed ↗