Molecular, Functional, and Ecological Characterization of Antarctic Penguin Orthoavulaviruses (AVV17–19)
Gabriela Muñoz, César Echeverría, Raúl Alegria, Rafael A. Medina, Cristian Torres, Sergio A. Bucarey, Marcelo González-Aravena, Víctor Neira

TL;DR
This study investigates three avian avulaviruses in Antarctic penguins, finding limited replication and environmental spread.
Contribution
The study provides novel insights into the replication dynamics and ecological spread of Antarctic penguin avulaviruses.
Findings
AVV18 showed productive replication in REM 134 cells and embryonated chicken eggs.
AVV17 and AVV19 did not replicate efficiently in tested cell lines or eggs.
Field surveillance detected low-level circulation of AVV17–19 only in penguin fecal samples, not in environmental samples.
Abstract
In Antarctic ecosystems, avian avulaviruses (AVVs) contribute to viral diversity in bird populations, particularly penguins. However, little is known about their biological properties and circulation patterns. This study analyzed three historical penguin avulavirus strains (AVV17, AVV18, and AVV19) to better understand their replication and distribution. Using molecular techniques, the viruses were identified and tested for replication ability in cell cultures and embryonated chicken eggs. Only for the AVV18 productive replication in cells was evidenced, while AVV17 and AVV19 showed no efficient replication. Field surveillance in the South Shetland Islands revealed low-level circulation of AVV17, AVV18, and AVV19, detectable only in penguin fecal samples, with no viral presence in environmental samples. These findings provide valuable insight into the replication dynamics and limited…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
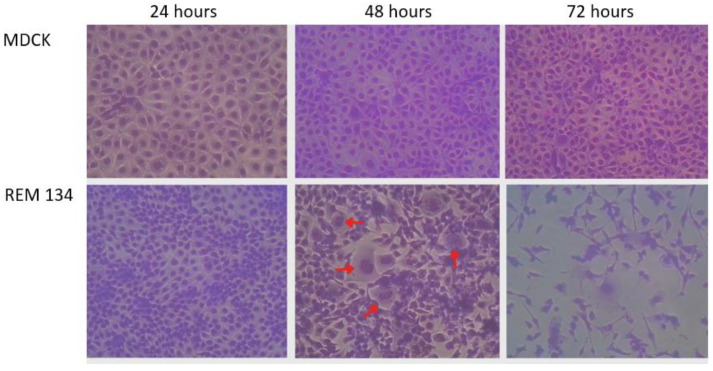 Figure 1
Figure 1- —Animal Virology Laboratory, Faculty of Veterinary and Animal Sciences, Universidad de Chile
- —ANID National Doctoral Scholarship
- —ANID Anillo
- —Center for Research in Influenza Pathogenesis (CRIP)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVirology and Viral Diseases · Respiratory viral infections research · Viral gastroenteritis research and epidemiology
1. Introduction
Avian avulaviruses (AVVs; family Paramyxoviridae, genus Orthoavulavirus) are enveloped, negative-sense, single-stranded RNA viruses that infect a broad range of avian hosts and exhibit diverse biological and pathogenic phenotypes [1]. The best-studied member is Avian Orthoavulavirus 1 (AOaV-1), also known as Newcastle disease virus (NDV), due to its significant impact on poultry health and international trade [2,3]. Beyond poultry systems, AOaV-1 and other avulaviruses circulate widely in wild birds, where infection outcomes range from subclinical to severe disease depending on viral genotype, host species, and ecological context. Understanding the diversity, host range, and maintenance of avulaviruses in natural systems is therefore central to both avian health and broader One Health-relevant surveillance [4], particularly in wildlife reservoirs where viral persistence occurs in the absence of apparent disease.
Polar ecosystems have long been considered remote, yet mounting evidence indicates that they are neither virologically isolated nor static. Early studies in Antarctic birds documented serological evidence of exposure to NDV/AOaV-1 and to Avulaviruses, which at the time could not be assigned to a defined species, in penguin colonies, supporting historical contact between them [5]. Subsequent work also reported evidence of orthomyxo- and paramyxoviruses in Antarctic fauna, including reports consistent with the detection of AVVs in penguins from Antarctic regions [6]. More recently, serological evidence further supports the exposure of Antarctic and sub-Antarctic birds to NDV/AOaV-1, as demonstrated by the detection of specific antibodies in penguins and other seabirds from Antarctica and the Magallanes region [7]. Together, these observations indicate that Avulaviruses, including NDV/AOaV-1, have a history of circulation across polar and subpolar avian communities, spanning multiple decades and host species, raising questions about viral introduction routes, persistence, and host-specific constraints in extreme environments.
In parallel, the discovery of novel penguin-associated AVVs has reshaped our understanding of Antarctic avulavirus diversity. In 2017, three genetically distinct avulaviruses were isolated from Gentoo penguins (Pygoscelis papua) in Antarctica and characterized as novel species (later corresponding to AVV17, AVV18, and AVV19), with complete genome sequencing supporting their distinctness [8]. Notably, these viruses correspond to viral species previously detected or inferred in Antarctic birds but could not be taxonomically resolved at the time of earlier serological and virological surveys. Later, Wille et al. (2019) expanded the landscape of Antarctic penguin AVVs by demonstrating that Antarctic penguins act as reservoirs of avulavirus diversity, with evidence of multiple avulavirus lineages and patterns of shedding dependent on host and sampling context [9]. Complementary to molecular studies, Olivares et al. (2019) provided serological evidence that AVV17-19 exposure is widely distributed across penguin species and locations along the Antarctic Peninsula, indicating broad-scale circulation over space and time [10], and suggesting long-term maintenance of these viruses within Antarctic penguin populations.
Despite these advances in recent years, major gaps remain. Much of the available information on Antarctic AVVs is derived from molecular detection and/or serology, approaches that are essential for surveillance but do not directly resolve whether distinct AVVs differ in replicative capacity, cellular permissiveness, or propagation potential under standardized laboratory conditions. In particular, few studies have leveraged molecularly validated historical isolates to explore lineage-specific phenotypic differences. Moreover, while earlier work established the presence and distribution of AVV17-19, less is known about their current circulation in recent Antarctic seasons and whether viral RNA can be detected outside the host (e.g., environmental substrates), which would inform shedding intensity and environmental persistence in polar settings.
Accordingly, in this study, we aimed to integrate molecular, functional, and ecological approaches to refine the understanding of Antarctic penguin avian AVVs. First, we performed species-level identification of historical egg-isolated AVV17, AVV18, and AVV19 stocks using a previously validated, lineage-specific L-gene RT-qPCR assay. Second, we conducted a standardized replication screening in mammalian cell lines and embryonated chicken eggs to explore lineage-specific differences in permissiveness and propagation capacity under the experimental systems tested. Finally, we conducted contemporary field surveillance during the 2024–2025 Antarctic season to evaluate the current circulation of these AVVs in penguin populations and their detection in environmental samples. By linking phenotypic information derived from historical isolates with present-day circulation patterns, spanning more than a decade of Antarctic surveillance, this study provides a framework for understanding the biological behavior and ecological maintenance of Antarctic Avulaviruses.
2. Materials and Methods
2.1. Molecular Identification and Classification of Historical Penguin Orthoavulavirus Isolates
Historical avian virus isolates analyzed in this study originated from Antarctic avian influenza surveillance expeditions conducted during the austral summers of 2014 and 2018. During these campaigns, cloacal and fecal samples were collected from Gentoo (Pygoscelis papua), Adélie (P. adeliae), and chinstrap penguins (P. antarcticus) at multiple locations in Antarctica. Samples were initially processed as part of routine influenza A virus (IAV) surveillance. Processing included centrifugation and inoculation into specific-pathogen-free (SPF) embryonated chicken eggs, following standard virological protocols [11]. Allantoic fluids harvested from inoculated eggs were screened by hemagglutination (HA) assays [12]. Several isolates produced positive HA reactions, with titers ranging from 4 to 128 hemagglutinating units. These HA-positive samples were subsequently tested for IAV by real-time RT-PCR following World Health Organization (WHO) protocols and were consistently negative [13,14]. At the time of isolation, the HA-positive, IAV-negative viruses were therefore considered putative avian paramyxoviruses; however, due to limited molecular tools and incomplete taxonomic frameworks available at that time, most isolates could not be assigned to a defined avulavirus species. Then, archived egg-derived viral stocks were preserved at −20 °C in the Animal Virology Laboratory, Universidad de Chile. With the subsequent description and taxonomic classification of novel penguin-associated avulaviruses, including avian avulavirus 17, 18, and 19, these historical isolates were reanalyzed in the present study to enable their molecular identification and classification prior to functional characterization.
For PCR amplification, total RNA was first extracted using TRIzol™ (Invitrogen™, Carlsbad, CA, USA) according to the manufacturer’s instructions and eluted in 50 µL of nuclease-free water. After extraction, RNA samples were stored at −20 °C until analysis. Lineage-specific real-time RT-qPCR assays targeting the L gene were performed using an Applied Biosystems thermocycler and the OneStep RT-PCR Kit in a final reaction volume of 20 µL. AVV. Primer sets were those described and validated by Wille et al. (2019) for AVV17, AVV18, and AVV19 detection and classification (Table 1) [9]. Cycling conditions included reverse transcription at 45 °C for 10 min, initial denaturation at 95 °C for 10 min, followed by 45 cycles of 95 °C for 15 s and 60 °C for 45 s [9].
2.2. In Vitro Replication Assays and Phenotypic Characterization in Mammalian Cell Lines
A subset of historical avulavirus isolates and derived subcultures (n = 35) exhibiting the lowest Ct values in lineage-specific RT-qPCR assays, representing all detected AVV lineages, was selected for in vitro time-course screening for replication indicators. These isolates were used to evidence replication phenotypes in mammalian cell lines and to identify lineage-specific differences in cellular permissiveness and propagation capacity.
Cell cultures: The REM 134 and MDCK cell lines were used for viral growth assays. Cells were maintained in high-glucose Dulbecco’s modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% antibiotic–antimycotic solution (Corning^®^, Corning, Manassas, VA, USA). Cultures were incubated at 37 °C in a humidified atmosphere containing 5% CO_2_. REM 134 and MDCK cells were used as pragmatic screening platforms to evaluate the permissiveness of archived isolates under standardized conditions; embryonated chicken eggs were included as an avian-permissive propagation system.
Evaluation of cytopathic effects in REM 134 and MDCK cells: For infection assays, REM 134 and MDCK cells were trypsinized, seeded into 6-well plates, and incubated at 37 °C with 5% CO_2_ until reaching 90–100% confluence. Monolayers were inoculated with 200 µL of each avulavirus isolate and incubated for 1 h at 37 °C to allow viral adsorption. Following adsorption, the inoculum was removed and replaced with viral maintenance medium consisting of DMEM supplemented with 2% bovine serum albumin (BSA), TPCK-treated trypsin (1 µg/mL), and 1% antibiotic–antimycotic solution (penicillin 100 U/mL, streptomycin 100 µg/mL). Negative controls of uninfected cells were included.
Cells were monitored for cytopathic effects (CPE) at 24, 48, and 72 h post-infection using an inverted microscope (Nikon Eclipse TS100, Tokyo, Japan). Morphological changes, including syncytium formation, cytoplasmic vacuolization, and loss of cell adherence, were recorded. In addition, Diff-Quick cytological staining was performed to further evaluate virus-induced cellular alterations following standard methanol fixation and sequential eosinophilic and basophilic staining procedures. Isolates exhibiting reproducible cytopathic effects and/or evidence of viral replication indicators, as determined by longitudinal RT-qPCR analyses using the L-gene protocol described above and showing a decrease in Ct values over time relative to the original inoculum, were subsequently selected for propagation in embryonated chicken eggs to further assess productive infection under this permissive system and to allow comparison with cell culture-based replication outcomes. Briefly, SPF embryonated chicken eggs were inoculated with the selected AVV18 isolate following standard procedures for avian virus propagation. Eggs were incubated under controlled conditions and monitored daily. Allantoic fluid was aseptically harvested and screened by lineage-specific L-gene RT-qPCR as described above. To further evaluate replication-associated activity, a subset of RT-qPCR–positive allantoic fluids was re-inoculated onto REM 134 cell monolayers under the same conditions described for the cell-culture assays, and cells were monitored for cytopathic effects (CPE). In parallel, hemagglutination (HA) activity of allantoic fluid was assessed using a standard red blood cell (RBC) agglutination assay. The HA testing was performed using two-fold serial dilutions of allantoic fluid in PBS in V-bottom microtiter plates, followed by the addition of an equal volume of 0.5% chicken RBCs. Plates were gently mixed and incubated at room temperature until hemagglutination patterns were readable. HA titers were recorded as the reciprocal of the highest dilution showing complete hemagglutination and expressed as hemagglutinating units (HAU). Appropriate negative controls were included in all assays.
2.3. Molecular Surveillance During the 2024–2025 Antarctic Season
Between December 2024 and February 2025, molecular surveillance for avian avulaviruses was conducted at selected Antarctic sites within the South Shetland Islands and the Antarctic Peninsula. Sampling locations were chosen based on logistical feasibility, accessibility during the austral summer, and the presence of established seabird colonies. Cloacal swabs were collected directly from individual birds using sterile swabs, while environmental samples were obtained from substrates within the immediate surroundings of bird colonies, including areas with visible fecal deposition. Sample numbers, host species, and sample type by site are now reported in Table 2. All samples were collected from adult individuals, as this was the only age class authorized under our sampling permit. The study was approved by the University of Chile Institutional Animal Care and Use Committee (IACUC) (protocol 22603-VET-UCH).
All samples were placed in viral transport medium and stored at −20 °C until processing. Sampling sites included Ardley Island (62°12′ S, 58°55′ W), Lions Rump on King George Island (62°11′ S, 58°59′ W), the western shore of the Fildes Peninsula (62°12′ S, 58°57′ W), Isabel Riquelme Islet (63°19′ S, 57°54′ W), and Rozas Island (63°18′ S, 57°54′ W). These locations support high densities of wildlife, including breeding colonies of Gentoo (Pygoscelis papua), chinstrap (P. antarcticus), and Adélie (P. adeliae) penguins, as well as southern giant petrels (Macronectes giganteus).
Wildlife populations at each site were systematically monitored during sampling activities. Clinical observations were performed by trained veterinarians, with particular attention to general condition and the presence of overt signs of disease. All field activities were conducted in accordance with the environmental protection and animal welfare guidelines established under the Antarctic Treaty System.
Detection of avian Avulaviruses in cloacal and environmental samples was performed using lineage-specific real-time RT-PCR assays targeting the L gene, as described above [10].
3. Results
3.1. Molecular Classification of Historical Avian Avulavirus Samples
Lineage-specific RT-qPCR screening of historical avian samples revealed the presence of multiple avian avulavirus lineages in penguins sampled during the 2014 and 2018 Antarctic seasons (Table 1). Among samples collected from P. papua at Kopaitic Island in 2014, AVV17, AVV18, and AVV19 were detected. Twelve out of the 13 isolates were classified. AVV17 was identified in five Kopaitic Island isolates, with Ct values ranging from 13.81 to 28.58; AVV18 was detected in three samples (Ct 21.74–34.19), and AVV19 was detected in four samples (Ct 17.96–32.39).
In samples collected in 2018, AVV19 was detected in a P. papua sample from Ardley Island (Ct 24.13), whereas no avulavirus RNA was detected in the other five samples. Among samples collected at the Avian Island in 2018, only AVV17 RNA was detected; four P. adeliae samples showed late amplification signals (Ct 34.32–39.14), while one sample from this site remained negative. Overall, these results confirm the circulation of multiple avian avulavirus lineages in Antarctic penguins across different locations and sampling years, while also highlighting substantial heterogeneity in detection frequency and viral RNA levels among samples.
3.2. Transient Replication and Syncytium Formation of AVV18 in REM 134 Cells
Replication indicator assays in mammalian cell lines were attempted using 10 distinct avian avulavirus isolates, selected based on their lower Ct values (<30) in the lineage-specific RT-qPCR screening (Table 2). Owing to multiple propagation steps and parallel passages, these experiments comprised 35 independent culture attempts derived from the original 10 isolates. A total of 25 AVV19-positive samples and 7 AVV17-positive samples were included in the initial screening, with Ct values ranging from 17 to 22. However, no cytopathic effects (CPE) or consistent decreases in Ct values suggestive of productive replication were observed in either REM 134 or MDCK cells. Accordingly, AVV17 and AVV19 isolates did not evidence productive replication in mammalian cell culture and were not further propagated in embryonated chicken eggs.
In contrast, an AVV18 isolate (Avv18/P. papua/Kopaitic/Neira-88/2014) originally derived from embryonated chicken eggs showed evidence of initial replication in REM 134 cells, as indicated by a reduction in Ct values from 22 at 24 h post-infection (hpi) to 18 at 48 hpi, accompanied by syncytium formation and progressive monolayer destruction (Figure 1). However, subsequent passages from REM 134 to REM 134 did not sustain evidence compatible with transient replication/limited amplification under these conditions, nor was evidence found in MDCK cells, where Ct values increased from 26 to 28. Consequently, the AVV18 isolate was re-propagated in embryonated chicken eggs to confirm productive viral replication under permissive conditions. Thus, 32 SPF embryonated chicken eggs were inoculated. Viral amplification was observed in 10 eggs, as indicated by reduced RT-qPCR Ct values (Ct range: 16–18). Allantoic fluid from these RT-qPCR–positive eggs produced replication-associated signals after re-inoculation into REM 134 cells. Hemagglutination activity was detected in 6/7 tested eggs, with titers ranging from 128 to 1024 HAU.
Representative micrographs of MDCK and REM 134 cells at control, 24 h, and 48 h post-infection. Syncytia (red arrows) and progressive monolayer destruction are evident in REM 134, whereas MDCK cells show only mild detachment at 48 hpi. Images were captured at 10× magnification.
3.3. Limited Detection of Avian Avulaviruses in Antarctic Birds During the 2024–2025 Season
During the surveillance period, a total of 716 samples were collected across five Antarctic sites: Ardley Island, Lions Rump, the western shore of the Fildes Peninsula, Isabel Riquelme Islet, and Rozas Island. At Ardley Island, 535 samples were obtained from P. papua (432 environmental and 103 cloacal swabs), of which three cloacal samples were positive for AVV17 (0.6% of total P. papua samples; 2.9% of cloacal samples). Among P. antarcticus (46 cloacal samples), one sample was positive for AVV17 (2.2%), and two were positive for AVV18 (4.3%), all detected in cloacal swabs. In P. adeliae (34 samples; three environmental and 31 cloacal), two cloacal samples were positive for AVV18 (5.9%) and one for AVV19 (2.9%). All other bird species sampled at Ardley Island (Sterna vittata, Eudyptes chrysolophus, Larus dominicanus, and Stercorarius antarcticus) tested negative for all avulavirus lineages.
At Lions Rump, all seven environmental samples collected from Larus dominicanus (n = 3) and Stercorarius antarcticus (n = 4) were negative for avian avulaviruses. On the western shore of the Fildes Peninsula, three of ten cloacal samples from P. papua were positive for AVV18 (30%), and one sample was positive for AVV19 (10%). Additionally, three of thirteen cloacal samples from P. antarcticus were positive for AVV18 (23.1%). Samples from P. adeliae (one cloacal), Stercorarius antarcticus (two environmental), Macronectes giganteus (seven environmental), and Larus bransfieldensis (one environmental) were all negative.
At Isabel Riquelme Islet, all samples collected tested negative for avian avulaviruses. Similarly, at Rozas Island, all 19 samples collected from P. antarcticus (12 environmental and seven cloacal) were negative. Overall, avian avulavirus detection was primarily associated with species of the genus Pygoscelis, with AVV18 being the most frequently detected lineage (10/716 samples, 1.4%), followed by AVV17 (4/716, 0.6%) and AVV19 (2/716, 0.3%). All positive detections were obtained from cloacal swabs, whereas no avulavirus RNA was detected in environmental samples, indicating limited circulation and shedding in the surveyed Antarctic bird populations. These results are summarized in Table 3. Finally, because the number of positive detections was very small, formal statistical comparisons and inferential analyses were not performed.
4. Discussion
Antarctic avian avulaviruses represent an emerging component of polar viromes, yet their biological properties, replication constraints, and contemporary circulation patterns remain incompletely defined. In this study, we combined molecular classification of archived egg-derived isolates with functional assays in mammalian cell lines and embryonated eggs, and complemented these experiments with molecular surveillance during the 2024–2025 Antarctic season. Overall, our findings indicate that Antarctic AVVs circulate at low levels in penguin populations, while exhibiting lineage-specific variation in their capacity to initiate replication-associated signals under standardized laboratory conditions.
Importantly, we provide a retrospective molecular classification of historical hemagglutinating, IAV-negative isolates recovered during influenza surveillance that could not be assigned to a defined species at the time of collection. By applying lineage-specific RT-qPCR assays targeting the L gene, we were able to re-identify and classify a substantial proportion of archived egg-derived stocks as AVV17, AVV18, or AVV19, thereby directly linking historical isolation efforts with the taxonomic framework later established for penguin-associated avulaviruses [8,9]. Importantly, isolates that remained unclassified are most plausibly explained by very low viral RNA levels at the time of testing, compounded by prolonged storage over several years, which may have led to partial RNA degradation despite frozen preservation. This highlights both the value and the inherent limitations of retrospective analyses of archived viral material. Nevertheless, this molecular “bridge” between historical samples and modern diagnostic tools enables functional and phenotypic characterization of viruses that would otherwise persist only as undocumented historical observations. Importantly, this retrospective classification provides species-level identification of archived isolates, but it does not establish strain-level relatedness or direct genetic continuity with contemporary detections; such linkage would require sequencing-based analyses.
In cell culture, AVV18 showed evidence of replication in REM 134 cells, including syncytium formation and a decrease in Ct values between 24 and 48 hpi. However, replication was not sustained upon subsequent passages in REM 134, and no productive replication was observed in MDCK cells. This pattern is consistent with transient replication, where an initial round of infection produces measurable cytopathic effects and increased viral RNA levels, but the infection fails to amplify efficiently across serial passages. Such limited propagation may reflect an inoculum effect [15]; insufficient production of infectious progeny, or cell line-specific constraints that prevent sustained spread. Alternatively, monoculture conditions in REM 134 may lack essential determinants of multicycle replication, such as adequate receptor density [16], or additional entry/egress factors and intercellular interactions that support efficient cell-to-cell transmission [17,18]. Accordingly, we interpret these findings as replication-associated signals under the specific in vitro conditions tested, rather than definitive evidence of sustained multicycle productive infection or a generalizable lineage property. Therefore, infectivity-based assays (e.g., TCID_50_ or plaque assays) will be required in future work.
For paramyxoviruses, cell entry and multicycle spread are strongly shaped by host protease availability and proteolytic activation of the fusion (F) glycoprotein, which is a key determinant of tropism, spread, and pathogenicity [19]. In particular, differences in F cleavage requirements can restrict productive replication in vitro even when viral attachment and initial entry occur, potentially explaining a phenotype compatible with early fusogenic CPE followed by failure to sustain replication across passages [20,21]. Although TPCK-treated trypsin was included to support F activation, lineage-specific compatibility with the available protease(s) and/or efficiency of fusion activation may still limit amplification beyond the first replication cycle [19]. Importantly, Ct values alone cannot demonstrate production of infectious progeny, because RNA detection does not distinguish between infectious and non-infectious virus and can reflect restricted replication or residual input RNA; therefore, infectivity-based titration (e.g., TCID_50_ or plaque assay) is needed to determine whether AVV18 produces low-level infectious progeny in REM 134 cells or whether the observed Ct decrease reflects a limited, non-sustained replication event.
In contrast, AVV17 and AVV19 remained detectable by RT-qPCR in historical stocks but did not produce CPE or consistent Ct decreases suggestive of replication in either REM 134 or MDCK cells. This lack of replication could be explained by reduced viability of archived material, lineage-specific host range restrictions, or the requirement for alternative permissive systems not captured by the two mammalian cell lines used here. Differences in cytopathic outcomes among cell lines highlight the importance of host factors [22]. MDCK cells did not support evidence of productive replication of AVV17, AVV18, or AVV19, potentially due to intact type I interferon signaling restricting infection and/or limited availability of compatible sialylated receptors [23]. Recent work shows that disrupting IFNAR1 signaling in MDCK cells can markedly enhance replication of influenza vaccine viruses [24], underscoring interferon-mediated restriction as a major determinant of permissiveness. REM 134 cells, derived from a mammary carcinoma, may exhibit altered interferon signaling pathways [25], which could partially explain why AVV18 initiated replication in this system. Nevertheless, the inability to sustain replication in REM 134 suggests that permissiveness is incomplete. In previous work, MDBK cells exhibited more pronounced cytopathic effects than Vero cells following infection with penguin-associated avulaviruses, although viral replication could not be sustained across passages [8]. In contrast, Vero cells, despite lacking type I interferon production, showed more limited cytopathic responses [8]. Together, these observations highlight that in vitro phenotypes depend not only on innate immune status but also on cell-specific factors, and that cell line selection remains a decisive and context-dependent factor for the phenotypic characterization of Antarctic avulaviruses. Because avian-derived cell systems were not available for this study, we refrain from inferring avian tropism from mammalian cell-culture results and highlight avian models as a priority for future work. Importantly, the embryonated egg system provided a permissive benchmark to support evidence of productive replication when cell culture outcomes were limited. AVV18 amplification in a subset of inoculated eggs, evidenced by Ct decreases, supports the conclusion that this lineage can replicate efficiently under appropriate conditions even when multicycle replication in mammalian monolayers is constrained. This reinforces the value of using eggs as a “reference propagation system” for avian viruses and provides a practical route for generating standardized virus stocks for downstream studies (e.g., sequencing, antigenic characterization, or receptor-binding assays).
From an ecological perspective, our 2024–2025 surveillance revealed low-level circulation of AVV17–19 across sites, with detections limited to cloacal swabs and no viral RNA detected in environmental samples. Several factors may explain the lack of environmental positivity, including PCR inhibitors in fecal matrices [26], rapid viral inactivation under environmental conditions [27], and low or intermittent shedding. Environmental degradation of RNA and the fragility of lentogenic paramyxoviruses outside the host have been documented [28], and may be accentuated in Antarctic settings due to freeze–thaw cycles, UV exposure, and physicochemical conditions that reduce stability. We did not perform dedicated inhibition spike-in controls, which is a limitation; therefore, negative environmental RT-qPCR results should be interpreted cautiously and do not exclude low-level shedding at the surveyed sites. Additionally, non-penguin bird species were primarily represented by environmental samples, which may have reduced detection sensitivity and limited inference about their potential role in AVV dissemination. The restriction of positive detections to cloacal swabs also provides insight into shedding patterns and surveillance strategy. While Wille et al. (2019) reported that oropharyngeal shedding can be prominent (in experimentally inoculated chickens) and more prevalent in penguin chicks, our study, which focused on adult birds and incorporated extensive environmental sampling, suggests that cloacal detection may occur at low levels and may be more readily captured in targeted swabbing than in environmental substrates [9]. This underscores the importance of aligning sampling design (age class, sample type, timing) with the expected biology of each lineage when interpreting apparent prevalence and comparing across studies.
Overall, the observed positivity was low and host-associated, with detections primarily in Pygoscelis penguins. The absence of overt clinical disease is consistent with predominantly low pathogenic phenotypes described for avulaviruses in wild birds [8,9,10]. Compared with prior studies, we observed a higher proportion of AVV18 detections, whereas AVV17 and AVV19 predominated in earlier reports [9,10]. However, Olivares et al. used serology [10], which reflects past exposure rather than active infection [29], limiting direct comparability with RT-qPCR. Moreover, differences in host age, colony structure, sampling site, and interannual viral circulation likely contribute to variability across seasons. Therefore, differences in lineage frequency across studies should be interpreted cautiously and cannot be taken as evidence of long-term replacement or persistence of specific strains without sequencing across seasons.
Finally, our results should be interpreted considering several limitations. First, the number of cloacal swabs was constrained by field logistics and environmental restrictions, which may reduce sensitivity to detect rare events. Second, testing additional avian-relevant systems, such as primary avian epithelial cells, avian cell lines, or ex vivo organ culture, may better capture the natural host tropism of Antarctic AVVs than mammalian monolayers. Addressing these points in future work will be important for defining the mechanisms governing lineage-specific replication, host restriction, and persistence in Antarctic ecosystems.
5. Conclusions
Antarctic avian avulaviruses (AVVs) represent an emerging component of the polar virome, though their biological characteristics, replication constraints, and current circulation patterns remain not fully understood. In this study, we combined species-level molecular classification of archived egg-derived isolates with functional assays in mammalian cell lines and embryonated eggs, alongside molecular surveillance during the 2024–2025 Antarctic season. Our findings show that AVVs circulate at low levels in penguin populations, with lineage-specific differences in replication-associated capabilities under controlled laboratory conditions.
A significant contribution of this work is the retrospective molecular classification of historical avulavirus isolates that were originally recovered during influenza surveillance but were not assigned to a specific species at the time. By using lineage-specific RT-qPCR assays, we successfully re-identified and classified these isolates as AVV17, AVV18, and AVV19, bridging historical isolation efforts with modern diagnostic tools. Additionally, we observed that AVV18 exhibited transient replication in REM 134 cells, but evidence of replication could not be sustained across passages, suggesting potential limitations in the cell culture system. Furthermore, our 2024–2025 field surveillance revealed low-level circulation of AVVs in penguins, with viral RNA detected primarily in cloacal swabs and absent from environmental samples. Overall, these results highlight the complex dynamics of AVV replication and circulation in the Antarctic ecosystem, offering insights into their ecological distribution and the factors affecting their persistence in such extreme environments.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Klink A.C. Rula O. Sushko M. Bezymennyi M. Mezinov O. Gaidash O. Bai X. Stegniy A. Sapachova M. Datsenko R. Discovery of Avian Paramyxoviruses APMV-1 and APMV-6 in Shorebirds and Waterfowl in Southern Ukraine Viruses 20231569910.3390/v 1503069936992408 PMC 10058161 · doi ↗ · pubmed ↗
- 2Hu Z. He X. Deng J. Hu J. Liu X. Current Situation and Future Direction of Newcastle Disease Vaccines Vet. Res.2022539910.1186/s 13567-022-01118-w 36435802 PMC 9701384 · doi ↗ · pubmed ↗
- 3Moreira G. Rodrigues S. Gomes-Gonçalves S. Silva G. Amorim I. Silva E. Carmezim S. Soeiro V. Mesquita J.R. Moreira G. Highly Virulent Newcastle Disease Virus in Eurasian Collared Doves in the North of Portugal Animals 202515356310.3390/ani 1524356341463847 PMC 12729603 · doi ↗ · pubmed ↗
- 4Maes P. Amarasinghe G.K. Ayllón M.A. Basler C.F. Bavari S. Blasdell K.R. Briese T. Brown P.A. Bukreyev A. Balkema-Buschmann A. Taxonomy of the Order Mononegavirales: Second Update 2018 Arch. Virol.20191641233124410.1007/s 00705-018-04126-430663023 PMC 6460460 · doi ↗ · pubmed ↗
- 5Morgan I.R. Westbury H.A. Virological Studies of Adelie Penguins (Pygoscelis adeliae) in Antarctica Avian Dis.1981251019102610.2307/15900777039589 · doi ↗ · pubmed ↗
- 6Austin F.J. Webster R.G. Evidence of Ortho- and Paramyxoviruses in Fauna from Antarctica J. Wildl. Dis.19932956857110.7589/0090-3558-29.4.5688258855 · doi ↗ · pubmed ↗
- 7Ariyama N. Tapia R. Godoy C. Agüero B. Valdés V. Berrios F. García Borboroglu P. Pütz K. Alegria R. Barriga G.P. Avian Orthoavulavirus 1 (Newcastle Disease Virus) Antibodies in Five Penguin Species, Antarctic Peninsula and Southern Patagonia Transbound. Emerg. Dis.2021683096310210.1111/tbed.1403733587778 PMC 8684044 · doi ↗ · pubmed ↗
- 8Emeny J.M. Morgan M.J. Regulation of the interferon system: Evidence that Vero cells have a genetic defect in interferon production J. Gen. Virol.19794324725210.1099/0022-1317-43-1-247113494 · doi ↗ · pubmed ↗