Estimation of Genetic Parameters of Reproductive Traits in Holstein Cattle from Southern China
Wenjie Li, Shuwen Xia, Yanming Quan, Yangyang Shen, Weining Li, Kunlin Chen, Zhenjiang An, Yingying Jiang, Zengxiang Pan, Huili Wang

TL;DR
This study estimates genetic parameters for reproductive traits in Holstein cattle from southern China, finding that heifers have higher genetic potential for these traits than multiparous cows.
Contribution
The study provides region-specific genetic insights for Holstein cattle in southern China, emphasizing the importance of management over genetics for multiparous cows.
Findings
Heifers showed higher genetic potential for reproductive traits compared to multiparous cows.
Reproductive traits in multiparous cows have low heritability, indicating strong environmental influences.
Strong genetic correlations were observed among functionally related reproductive traits in multiparous cows.
Abstract
Reproductive performance directly affects the productivity and profit of dairy farms. To support region-specific breeding, this study estimated the genetic parameters for key reproductive traits in Holstein cattle from southern China. Analysis of over 117,000 records from 44,861 cows showed that heifers had better reproductive efficiency and higher genetic potential for these traits than multiparous cows. In multiparous cows, reproductive traits were found to have very low genetic heritability, meaning they are influenced more by farm management and environment than by genetics. This provides clear guidance: genetic selection for reproductive efficiency should be prioritized in heifer breeding programs, while for multiparous cows, investment should focus on optimizing nutritional, health, and environmental management to improve reproductive outcomes. Reproductive performance is a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
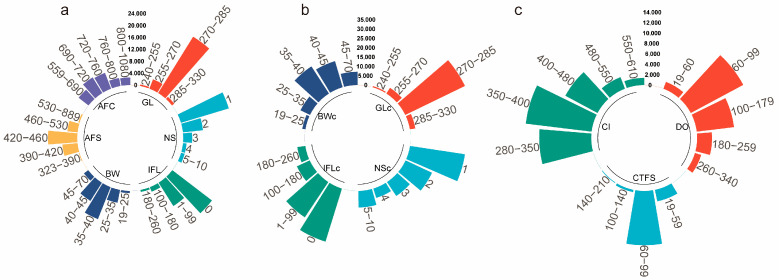 Figure 1
Figure 1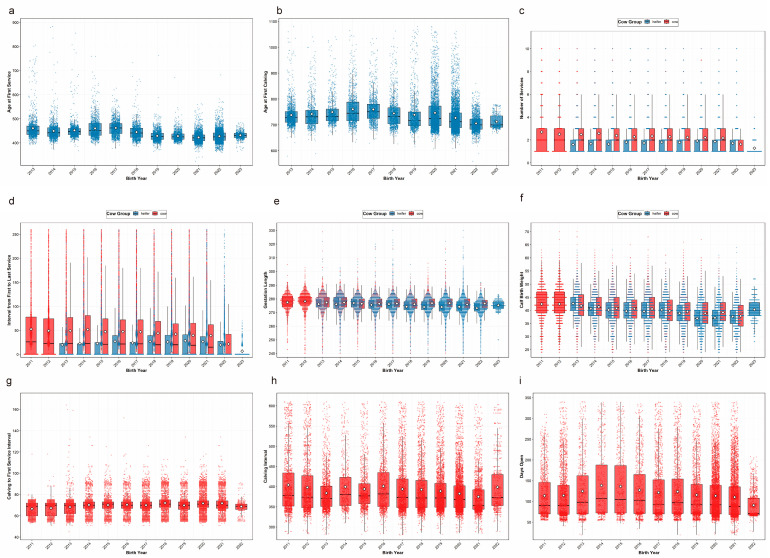 Figure 2
Figure 2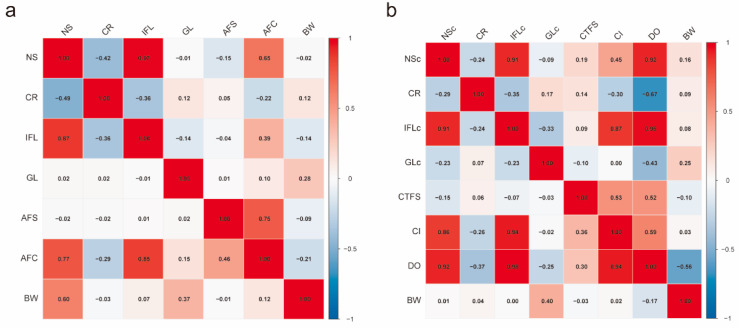 Figure 3
Figure 3- —Biological Breeding-Major Projects in National Science and Technology
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and phenotypic traits in livestock · Reproductive Physiology in Livestock · Effects of Environmental Stressors on Livestock
1. Introduction
Reproductive traits are critical functional characteristics in dairy cattle breeding [1], directly impacting calving intervals, lactation cycles, and herd renewal efficiency, thereby influencing farm productivity and profitability [2,3]. Among the key indicators used to assess reproductive development and efficiency in heifers are Age at First Service (AFS) and Age at First Calving (AFC), which reflect heifer maturation synchronization [4,5]. AFC is a defining moment that describes an important event in the cow’s life, as it marks the beginning of the dairy cow’s productive career [6,7]. Number of Services (NS) typically refers to the total instances of artificial insemination performed within a single estrous cycle or until conception is achieved. It serves as a key operational indicator for assessing both individual cow reproductive efficiency and overall herd reproductive management. NS is highly negatively correlated with Conception Rate at First Service (CR) and positively correlated with both Interval from First to Last Service (IFL) and Calving Interval (CI) [8,9,10]. Consequently, NS functions as a core variable that comprehensively reflects the efficiency of the entire reproductive management chain, from estrus detection to successful conception. These traits are influenced by both genetic and environmental factors [11], making genetic evaluation essential for developing effective breeding and management strategies [12].
The estimation of genetic parameters is fundamental to modern dairy cattle breeding [13,14]. Heritability and genetic correlations among traits provide key insights into the genetic architecture of reproduction and guide selection decisions [14,15,16]. Previous studies have primarily focused on estimating genetic parameters for Holstein cattle [17,18,19]. Schwarz et al. [20] reported heritability estimates ranging from 0.014 to 0.211 in German Holsteins, while corresponding values in Korean dairy cattle were between 0.10 and 0.17 [16]. In northern China, Guo et al. [21] estimated heritability ranges for reproductive traits in Beijing Holsteins to be between 0.034 and 0.100, and Chen et al. [22] reported heritability estimates ranging from 0.231 to 0.483 for Holstein heifers in Ningxia. However, studies on genetic parameters for Holstein populations in southern China remain relatively scarce [23]. China exhibits substantial north–south differences in climate, management practices, feed resources [24], and breeding history [25]. These environmental factors likely interact with genetic backgrounds [26], causing significant divergence in the phenotypic expression of reproductive traits [27]. Consequently, genetic parameters estimated from northern herds may not be directly applicable to southern populations.
Reproductive performance differs substantially between heifers and multiparous cows due to difference in physiological stage, disorder susceptibility, and genetic expression patterns [14,28]. Combining data from both groups may obscure trait-specific genetic patterns. Internationally, heifers and cows are commonly assessed separately in genetic evaluations to improve selection accuracy [10,14,28]. Physiologically, heifers are in a stage of ongoing growth and gradual maturation of reproductive function. Their growth capacity and ability to maintain pregnancy exhibit considerable genetic potential [22]. They generally outperform multiparous cows in reproductive metrics [14,29], warranting separate genetic analysis.
This study aims to estimate genetic parameters for key reproductive traits in southern Chinese Holstein cattle, with classified analyses for heifers and multiparous cows. Objectives included estimating variance components, heritability, and genetic correlations among traits, and evaluating genetic relationships across parities. This study can establish a genetic basis for tailored breeding strategies across regions and physiological stages, thereby facilitating the genetic improvement of low-heritability traits, and ultimately enhance the productivity and sustainability of dairy systems in southern China.
2. Materials and Methods
2.1. Phenotypic Data
Reproductive data from 2010 to 2024 were collected from eight large-scale dairy farms in southern China (Shanghai, Jiangsu, Zhejiang, Anhui, Fujian, and Yunnan). The study area encompasses a broad longitudinal gradient from the eastern coastal region to the southwestern inland region, covering longitudes from approximately 97° E to 123° E and latitudes from 21° N to 35° N. The dataset comprises 117,514 records from 44,861 Holstein cows, including insemination, pregnancy checks, and calving events, with 39,207 records from heifers and 78,307 records from multiparous cows. Cattle were housed in stall barns under an intensive confinement system. Cooling measures (fans and sprinklers) were implemented during summer. All animals were fed a total mixed ration (TMR) formulated according to the Chinese Feeding Standard for Dairy Cattle, based on corn silage, alfalfa hay, corn, soybean meal, and mineral supplements.
The reproductive traits analyzed in the study are shown in Table 1, along with their corresponding definitions. Given the distinct reproductive characteristics between heifers and multiparous cows, the reproductive data for these two groups were analyzed separately.
For the dichotomous trait CR, outcomes were coded as 0 (no confirmed pregnancy) or 1 (confirmed pregnancy). Records prior to the first calving were classified as parity 0 for heifers, while records for the first, second, and third calvings in multiparous cows were coded as parities 1, 2, and 3, respectively [30].
2.2. Quality Control of Reproductive Data
Data quality control was performed according to following criteria [31]: AFS: 270–900 days; NS/NSc: 1–10; IFL/IFLc: 0–260 days; GL: 240–330 days; CTFS: 19–210 days; AFC: 510–1080 days; CI: 280–610 days; DO: 19–340 days; BW/BWc: 24–70 kg. After quality control, the dataset was reduced from 117,514 records to 116,954 records.
2.3. Pedigree Information
Pedigree information was obtained for 80,794 Holstein cattle, comprising 2695 bulls and 78,099 cows. Among the 44,861 phenotyped individuals, 44,126 (98.4%) had parental records, and 43,982 (98.0%) had grandparental records. Overall, the pedigree completeness reached 85%. Offspring birth dates were cross-checked with parental birth dates, and unreasonable pedigree entries were either removed or corrected to meet the requirements of DMU [32] software (Version 6, release 5.6) for genetic evaluation. Missing parent or grandparent information was replaced with “0”.
2.4. Analysis of Fixed Effects
Reproductive traits in Holstein cattle are influenced by numerous non-genetic factors, including herd (H), birth year (BY), year of first service (FSY), birth season (BS), season of first service (FSS), calving season (CSE), parity (P), and inseminator (BRE). The classification levels for these non-genetic factors are shown in Table 2. In this study, factors potentially affecting the traits were first treated as fixed effects for analysis of variance, and only those found to be statistically significant were retained in the final genetic analysis model. The Generalized Linear Mixed Model (GLMM) in R (v 4.4.0) was employed to test the significance of fixed effects. The model is as follows:
where is the observed phenotypic value of the reproductive traits for the i-th herd, the j-th birth year, the k-th year of first service, the l-th birth season, the m-th season of first service, the n-th calving season, the o-th parity, and the p-th breeder. denotes the herd effect; denotes the birth year effect; denotes the year of first service effect; denotes the birth season effect; denotes the season of first service effect; denotes the calving season effect; denotes the parity effect; denotes the breeder effect; and represents the random residual error. A significance threshold of p < 0.05 was applied to determine which fixed effects should be included in the final model.
2.5. Genetic Evaluation Models
2.5.1. Single-Trait Animal Model
A single-trait animal model was employed to estimate the genetic parameters for reproductive traits in heifers (AFS, AFC, IFL, NS, GL, BW, and CR). The model is defined as follows:
where is the vector of observed phenotypic values for the heifer traits; is the vector of fixed effects; is the vector of additive genetic effects; is the vector of random residual effects; and and are the corresponding design matrices for and , respectively.
2.5.2. Single-Trait Repeatability Model
A single-trait repeatability model was used to estimate the genetic components for reproductive traits in multiparous cows (DO, CTFS, CI, IFLc, NSc, GLc, BWc, and CRc). This model is appropriate for traits with repeated measurements and is defined as:
where is the vector of individual phenotypic values; is the design matrix for fixed effects; is the vector of fixed effects; is the vector of additive genetic effects; is its corresponding design matrix; is the vector of permanent environmental effects; is the corresponding design matrix; and is the vector of residual effects.
2.5.3. Bivariate Models
To estimate trait correlations separately for heifers and multiparous cows, bivariate models were applied in two sequential analyses.
Bivariate animal model for heifer traits:
Bivariate repeatability animal model for multiparous cow traits:
In these expressions, represents the vector of phenotypic values for the i-th trait; is the vector of fixed effects for the i-th trait; is the vector of additive genetic effects for the i-th trait; is the vector of permanent environmental effects for the i-th trait; is the vector of random residual effects for the i-th trait; and , and are the corresponding design matrices.
2.5.4. Multi-Trait Models
To analyze reproductive traits across multiple parities, multi-trait models were employed, in which the same trait recorded in different parities was treated as multiple distinct traits.
Four-trait model for traits with records in both heifers and multiparous cows, including NS, CR, IFL, GL and BW:
Here, subscript 0, 1, 2, and 3 represent the parity groups (0 for heifers, and 1, 2, and 3 for first, second, and third parity in multiparous cows, respectively).
Three-trait model for traits with records only in multiparous cows including CTFS, CI, and DO:
In this model, subscript 1, 2, and 3 represent the first, second, and third parity in multiparous cows.
In the expressions above, represents the phenotypic value vector for the i-th trait; is the vector of additive genetic effects for the i-th trait; is the vector of random residual effects for the i-th trait; and and are the corresponding design matrices.
2.6. Estimation of Genetic Parameter
Based on the variance and covariance components estimated by the DMU software (Version 6, release 5.6) [32], the heritability ( ), repeatability ( ), genetic correlation ( ), and phenotypic correlation ( ) for each trait were calculated with the following formulas:
Heritability:
where is the additive genetic variance for the trait, and is the phenotypic variance. For heifers: . For multiparous cows: . Here, is the residual variance, and is the permanent environmental variance.
Repeatability:
Genetic correlation:
where is the additive genetic covariance between trait 1 and trait 2; and are the additive genetic variances of trait 1 and trait 2, respectively.
Phenotypic correlation:
where is the phenotypic covariance between trait 1 and trait 2; and are the phenotypic variances of trait 1 and trait 2, respectively.
3. Results
3.1. Descriptive Statistics of Reproductive Traits
Phenotypic data for heifers and multiparous cows for most reproductive traits followed a normal distribution, excluding the NS/NSc and IFL/IFLc traits (Figure 1). Summary statistics are presented in Table 3. Heifers showed relatively small variability, as indicated by their low coefficients of variation (CV). GL showed the smallest CV among all traits, while NS was primarily 1–2 services (91.5%), and IFL was mainly 0–99 d (89.7%).
In contrast, multiparous cows exhibited greater phenotypic variability for several traits, as evidenced by higher CV values. Although GLc remained within 270–285 d, NSc frequently extended to a third service (18.7% of records vs. 4.2% in heifers). BWc shifted toward 35–45 kg (78.9% of records), with a higher proportion exceeding 45 kg (11.3% vs. 3.6% in heifers). DO ranged from 60 to 179 d (76.4%), CTFS was 60–99 d, and CI spanned 280–480 d (89.2%).
Figure 2 illustrates the temporal distribution of key reproductive phenotypes in heifers (born 2013–2023) and multiparous cows (born 2011–2022) across southern China. Phenotypic distributions differ markedly between heifers and multiparous cows in key reproductive traits (Figure 2c–f).
In heifers, AFS, AFC and IFL showed a steady downward trend over the study period. GL stayed stable within the 270–280 d range throughout the period, and BW stabilized at a moderate 35–40 kg from 2018 onward, reflecting consistent reproductive performance optimization. For multiparous cows, notable positive shifts were evident: NSc, IFLc, CI and DO exhibited a downward trend. Similar to heifers, GLc maintained stability at 270–280 d. CTFS remained consistent at around 70 d, and BWc stabilized near 38 kg after 2013.
3.2. Genetic Parameters and Variance Components of Reproductive Traits
Heritability estimates for reproductive traits in heifers ranged from 0.04 to 0.47 (Table 4). AFS showed the highest heritability (0.47 ± 0.02), followed by GL (0.19 ± 0.03) and BW (0.19 ± 0.03). NS (0.14 ± 0.01) and AFC (0.12 ± 0.02) displayed moderate heritability. CR (0.08 ± 0.01) and IFL (0.04 ± 0.01) were low-heritability traits. Based on these estimates, AFS was classified as a high-heritability trait (h^2^ > 0.30), whereas NS, GL, AFC, and BW were of medium heritability (0.10 < h^2^ ≤ 0.30). CR and IFL were considered low-heritability traits (h^2^ ≤ 0.10).
For multiparous cows, heritability estimates ranged from 0.03 to 0.14, with repeatability coefficients between 0.05 and 0.36 (Table 5). GLc (0.14 ± 0.03) and BWc (0.12 ± 0.03) exhibited medium heritability, whereas CRc, NSc, IFLc, CTFS, CI, and DO were low-heritability traits (h^2^ ≤ 0.10). Repeatability was highest for CRc (0.36) and lowest for CTFS and DO (both 0.05).
3.3. Genetic and Phenotypic Correlations of Reproductive Traits
Genetic and phenotypic correlations among reproductive traits in heifers and multiparous cows are summarized in Figure 3. In heifers, genetic correlations ranged from −0.42 to 0.97, and phenotypic correlations from −0.49 to 0.87 (Figure 3a). Notably, high genetic correlations existed between NS and IFL, NS and AFC, as well as between AFC and AFS, with absolute values ranging from 0.65 to 0.97. Phenotypically, NS was strongly correlated with IFL, AFC, and calf birth weight (BW), with absolute values reaching 0.87. CR exhibited a moderate negative phenotypic correlation with NS (−0.49).
For multiparous cows, genetic correlations ranged from −0.67 to 0.96 (Figure 3b). High positive genetic correlations were observed between NSc and IFLc, NSc and DO, IFLc and CI, and IFLc and DO, with values between 0.87 and 0.96. Phenotypic correlations spanned −0.37 to 0.98, with IFLc showing particularly high phenotypic associations with NSc (0.91), CI (0.94), and DO (0.98).
Regarding phenotypic correlations, the range was −0.37 to 0.98. IFLc showed relatively high phenotypic correlations with NSc, CI, and DO, ranging from 0.91 to 0.98.
3.4. Correlation Coefficients Between Different Parity Levels for the Same Trait Based on a Multi-Trait Model
Genetic and phenotypic correlations between different parity levels for the same trait were estimated using a multi-trait model and are presented in Table 6. Phenotypic correlations were generally low (0.00–0.40) between heifers and multiparous cows, as well as among parities within multiparous cows.
Genetic correlations were strongly positive for GL between heifers and all three parities in multiparous cows (0.99) and among parities within multiparous cows (0.99). For other reproductive traits, genetic correlations between heifers and multiparous cows were moderate, ranging from 0.13 (between heifer IFL and multiparous cows at third parity) to 0.85 (between heifer GL and multiparous cows GLc across all parities). However, higher genetic correlations were observed among different parity levels within multiparous cows, ranging from 0.40 to 0.99 for most traits, except for CTFS, CI, and DO, where genetic correlations between the first parity and the other two parities were below 0.40. Notably, genetic correlations between heifers and first-parity cows were generally higher than those between heifers and later-parity cows. Negative phenotypic correlations were observed for traits such as NS, IFL, and DO across different parity levels.
4. Discussion
4.1. Overview of Reproductive Performance in Holstein Cattle from Southern China
This study investigated reproductive traits of Holstein cattle across multiple provinces in southern China. Heifers generally exhibited superior reproductive efficiency compared to multiparous cows, as reflected by a shorter IFL, lower NS, and lower BW. These findings align with previous studies [8,33,34], indicating better conception potential in heifers. Since heifers have not yet undergone lactational stress or postpartum metabolic challenges, their reproductive systems remain in an optimal physiological state. The observed differences between heifers and multiparous cows may be attributed to physiological changes in the reproductive system after first calving and the negative energy balance associated with high milk production [17,30,35].
Heifers exhibited earlier AFS and AFC in this study compared to previous studies in Beijing [21], Ningxia [22] and the southern part of China [23]. This may be attributed to earlier sexual maturity in southern China, as well as management strategies on farms that emphasize early breeding and calving in heifer rearing [36,37]. For multiparous cows, the DO (118 days) was consistent with reports from Beijing [21] and Shandong [13] in China, and from Denmark [38], but shorter than previous findings in southern China [23] (approximately 157 days). Given that heat stress has been shown to prolong DO [39], this improvement likely reflects enhanced farm management (e.g., cooling facilities and nutritional adjustments) mitigating summer heat stress in the region [40,41].
The mean CI was 392.10 ± 63.84 d, which is similar to previous studies [42,43], such as those from Beijing (397.60 ± 60.0 d) [21] and Canada (395.4–398.2 d), but shorter than reported for Jiangsu (421.14 ± 80.07 d) [44] and an earlier study in southern China (429.19 ± 100.05 d) [23], indicating optimized postpartum reproductive recovery. In this study, the mean GL was 274 d for heifers and 276 d for multiparous cows, consistent with global and domestic benchmarks for Holstein cattle [14,17,23].
4.2. Heritability of Reproductive Traits
In this study, the estimated heritability for reproductive traits in Holstein cattle from southern China ranged from low to moderate. Except for AFS (0.47) in heifers, the heritability for other reproductive traits is at a low-to-moderate level (0.03–0.19), which is consistent with findings from other studies [33,45].
Heifer AFS exhibited a notably high heritability (0.47), exceeding estimates from Canadian (0.12–0.13) [46], German [47], and other southern Chinese Holstein populations (0.06) [23]. This discrepancy may be attributed to the strictly controlled rearing environment for heifers on the studied farms, where standardized nutrition, disease management, and housing conditions likely reduce the environmental variance component for AFS [48]. Additionally, differences in statistical methods and management strategies may also contribute to variations in heritability estimates [49,50].
In contrast, the heritability of traits in multiparous cows is generally lower than those for heifers, consistent with the previous studies [46,51]. For instance, heritability for NS (0.14) and CR (0.08) was higher in heifers than in multiparous cows (NSc: 0.03; CRc: 0.07), which aligns with findings in German [52] and northern Chinese studies [51]. Our estimates for CR/CRc (0.08 and 0.07) were moderately higher than those from Nordic [14] and Dutch populations [53]. These variations may arise from differences in genetic variation between populations, the statistical models employed for analysis, or genotype–environment interactions [54,55]. Notably, GL and BW both exhibit moderate heritability (0.14–0.19) in both heifers and multiparous cows, suggesting appreciable genetic influence and potential for genetic improvement.
For traits exclusive to multiparous cows, the heritability for CTFS and DO was 0.08 and 0.05, respectively. Similar results have been reported for Iranian [56,57] and South African Holsteins [29], but our results are higher than estimates from Tunisian [58] and northern Chinese Holsteins [21]. The heritability for DO (0.05) and CI (0.07) was close but not identical, whereas Iranian results reported very similar values of 0.069 and 0.070 for DO and CI, respectively [57].
These results reveal significant differences in the genetic background of reproductive traits between heifers and multiparous cows, supporting the need for group-specific breeding strategies. Heifers possess greater genetic potential for reproductive improvement; selection for moderate heritability in heifers, such as AFS, AFC, NS, GL, and BW, could enhance early reproductive efficiency. Physiologically, heifers, having not yet undergone lactational stress or postpartum metabolic disorders, maintain an optimal reproductive state [35]. In contrast, most reproductive traits in multiparous cows generally exhibit low heritability and are highly susceptible to environmental influences [19,29,38]. Differences in rumen microbiome [59,60], metabolism [35,40], and management [39,41] across parities may explain the heritability differences between heifers and multiparous cows. For example, heifers exhibit a more stable rumen microbial structure [61], whereas multiparous cows undergo microbiota disruption due to lactation and metabolic shifts [60], indirectly impairing reproductive efficiency [59]. However, direct evidence linking rumen microbiome composition to reproductive trait variation across parities in Holstein cows in southern China remains limited, necessitating further research to confirm this mechanism.
4.3. Genetic and Phenotypic Correlations of Reproductive Traits
High genetic correlations among different reproductive traits reveal a similar genetic background, enabling indirect selection for correlated traits [29]. In this study, a high genetic correlation (0.75 ± 0.05) was observed between AFC and AFS in heifers. This result aligns with findings from Jagusiak and Zarnecki [62] and Brzáková et al. [63]. As both AFS and AFC are traits specific to heifers and collectively reflect their reproductive performance level [23], they may share a common physiological basis. Furthermore, the high positive genetic correlation between IFL and NS is likely due to the delayed timing of the final service caused by an increased number of inseminations, which is also reflected in the high phenotypic correlation between these two traits.
For multiparous cows, DO exhibited high genetic correlations (0.45–0.96) with several traits, including NSc, IFLc, and CI. The highest genetic correlation was observed between DO and IFLc (0.96), suggesting that DO could potentially substitute for IFLc in genetic evaluations when farm insemination records are incomplete [13,21]. Furthermore, high genetic correlations were found among traits specific to multiparous cows, reflecting their ability to conceive, maintain pregnancy, and return to estrus postpartum. The genetic correlations were 0.53 between CTFS and CI, 0.52 between CTFS and DO, and 0.59 between CI and DO. These estimates align with reports in Chinese [21,30] and New Zealand [64] Holstein populations. Phenotypic correlations among CTFS, DO, and CI were also strong (0.30–0.94), supporting the close biological linkage among these intervals, which aligns with previous studies [21,29,65]. In practice, using CTFS as a proxy for DO can minimize data errors, as DO excludes cows culled for infertility, and CTFS is available earlier. Shorter CTFS and DO offer a shorter CI, which can prolong the lactation period, thereby improving productivity and herd profitability [66].
Biologically phenotypic correlations were observed, and their patterns were like those of the genetic correlations for all traits. Very strong positive relationships of 0.98 and 0.94 were identified between DO and IFLc, and between DO and CI, respectively, meaning that when one of these traits decreases, the other tends to decrease as well. A negative phenotypic relationship (−0.37) was found between CR and DO, which is similar to the estimate found in Korean dairy cattle (−0.66) [8] and Chinese dairy cattle (−0.75) [13], reflecting the expected trade-off between reproductive and postpartum anestrus.
4.4. Genetic Relationships Across Parities Provide a Basis for Early Selection
In general, moderate genetic correlations were observed between heifer traits and their corresponding traits in multiparous cows, ranging from 0.13 (between heifer IFL and third-parity IFLc) to 0.85 (between heifer GL and GLc in the first, second, and third parity). This finding is consistent with reports by Pryce et al. [42], Norman et al. [45], and Gredler and Schnyder [67].
Although heifer reproductive traits are phenotypically distinct from those of multiparous cows, they are genetically correlated to a certain extent, a pattern also noted in Nordic Holsteins and red cattle [14]. For most traits, correlations among parities 1–3 ranged from 0.40 to 0.99, except for CTFS, CI, and DO, which showed lower correlations (<0.40) between first and later parity. Notably, genetic correlations were typically higher between heifers and first-parity cows than between heifers and higher-parity cows.
CR/CRc exhibited genetic correlations of 0.50 to 0.91 across parities, which is similar to Liu et al.’s estimate of 0.6 between specific parities [51] but lower than Nordic estimate of 0.90–0.97 [14], reflecting population or modeling differences. Meanwhile, gestation length (GL/GLc) showed near-unity genetic correlations (up to 0.99) across all parities, indicating it can be treated as the same trait genetically regardless of parity. In contrast, genetic correlation between NS and CR was low (0.03–0.47) between heifers and multiparous cows, indicating that these traits may be regulated by parity-specific genes and should be treated differently in genetic evaluation. These findings hold significant implications for genomic selection and early breeding: reproductive traits in heifers can serve as early indicators for predicting future reproductive performance [47]. Particularly for traits with high genetic correlations, such as GL and BW, where early selection in heifers can effectively enhance lifetime reproductive efficiency. Conversely, traits with low cross-parity correlations may require parity-specific breeding objectives or evaluation models.
4.5. Influence of Environment and Management on Reproductive Traits and Breeding Implications
The hot and humid climate of southern China constitutes a major environmental challenge to dairy cow reproductive performance. The low heritability estimates for most reproductive traits in multiparous cows observed highlight the substantial contribution of environmental variance, with heat stress being a particularly key factor [68,69]. Heat stress can extend DO and CI through multiple pathways, including suppression of estrus expression, impairment of oocyte competence [70,71], and compromised embryo survival [72]. Consistent with this, previous studies indicate that each unit increase in the temperature–humidity index can prolong Days Open by 5–10 days [39]. Therefore, genetic selection for improved reproductive in southern China should be integrated with active environmental management strategies.
To bridge the research outcomes with practical breeding applications, a differentiated selection strategy is proposed for heifers and multiparous cows. For heifer breeding programs, prioritize genetic selection for traits with moderate-to-high heritability (e.g., AFS) by incorporating estimated breeding values (EBVs) into selection indices. In contrast, reproductive traits in multiparous cows typically demonstrate low heritability (<0.10), leading to limited annual genetic gain. However, substantial genetic variance in multiparous cows indicates considerable genetic potential remains. In practice, for multiparous cows, management interventions, including optimized formulas and environment control, should be prioritized to minimize the impact of non-genetic factors on their reproductive performance. Practical measures such as implementing cooling systems [73,74] (e.g., spray-and-fan ventilation), scheduling breeding to avoid peak heat periods [69], and formulating diets that enhance thermoregulation and mitigate heat stress are essential to support genetic potential and optimize reproductive outcomes.
5. Conclusions
This study presents a comprehensive genetic analysis of key reproductive traits in Holstein cattle from southern China, with separate evaluations for heifers and multiparous cows. The main findings demonstrate that reproductive performance in heifers is generally superior to that in multiparous cows, as evidenced by shorter breeding intervals and fewer services required. Genetic parameter estimates revealed low-to-moderate heritability for most traits, indicating substantial environmental influence. Notably, significant genetic differences were observed between heifers and multiparous cows for the same traits, supporting their treatment as distinct traits in genetic evaluations. Strong genetic correlations were identified among functionally related traits, particularly between CI and DO in multiparous cows. These findings have important implications for breeding and management strategies in southern China. Given the low heritability of reproductive traits, genetic improvement should emphasize multi-trait selection indices that incorporate reproductive traits with other economically important characteristics. Simultaneously, management practices, particularly those addressing heat stress mitigation in the region’s challenging climate, are essential to optimize reproductive performance and enhance overall herd productivity.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Miglior F. Muir B.L. Van Doormaal B.J. Selection indices in Holstein cattle of various countries J. Dairy Sci.2005881255126310.3168/jds.S 0022-0302(05)72792-215738259 · doi ↗ · pubmed ↗
- 2Giordano J.O. Kalantari A.S. Fricke P.M. Wiltbank M.C. Cabrera V.E. A daily herd Markov-chain model to study the reproductive and economic impact of reproductive programs combining timed artificial insemination and estrus detection J. Dairy Sci.2012955442546010.3168/jds.2011-497222916951 · doi ↗ · pubmed ↗
- 3Sitko E.M. Di Croce F.A. Mc Neel A.K. Weigel D.J. Giordano J.O. Effect of reproductive management programs that prioritized artificial insemination at detected estrus or timed artificial insemination on the economic performance of primiparous Holstein cows of different genetic merit for fertility J. Dairy Sci.20231066495651410.3168/jds.2022-2267437474372 · doi ↗ · pubmed ↗
- 4Hutchison J.L. Van Raden P.M. Null D.J. Cole J.B. Bickhart D.M. Genomic evaluation of age at first calving J. Dairy Sci.20171006853686110.3168/jds.2016-1206028624286 · doi ↗ · pubmed ↗
- 5Márton J. Bene S.A. SzabóF. Heritability Estimates of Age at First Calving and Correlation Analysis in Angus Cows Bred in Hungary Animals 202414371510.3390/ani 1424371539765619 PMC 11672652 · doi ↗ · pubmed ↗
- 6Hare E. Norman H.D. Wright J.R. Trends in calving ages and calving intervals for dairy cattle breeds in the United States J. Dairy Sci.20068936537010.3168/jds.S 0022-0302(06)72102-616357301 · doi ↗ · pubmed ↗
- 7Bene S.A. Kőrösi Z.J. Bognár L. Polgár J.P. SzabóF. Relationship Between Age at First Calving and 305-Day Milk Yield in Hungarian Holstein-Friesian Cows: Trends and Genetic Parameters Animals 202515364810.3390/ani 1524364841463932 PMC 12729427 · doi ↗ · pubmed ↗
- 8Lee J.G. Seo J. Alam M. Song H. Lee S. Cho J. Dang C.G. Lee J. Estimation of genetic parameters for reproductive traits in Korean dairy cattle Anim. Biosci.202538334010.5713/ab.24.045539483025 PMC 11725725 · doi ↗ · pubmed ↗