Genomic Characterisation of Antibiotic-Resistant Escherichia coli from an Intensive Poultry Production System in the uMgungundlovu District, KwaZulu-Natal, South Africa: A Snapshot
Nelisiwe S. Gumede, Joshua Mbanga, Charles Hunter, Melissa Ramtahal, Sabiha Y. Essack, Linda A. Bester

TL;DR
This study characterizes antibiotic-resistant Escherichia coli from a poultry farm in South Africa, highlighting the risk of resistance spreading through food and the environment.
Contribution
The study provides genomic insights into multidrug-resistant E. coli in a South African poultry system and their global phylogenetic links.
Findings
74% of confirmed E. coli isolates were multidrug-resistant, with high resistance to tetracycline, ampicillin, and trimethoprim–sulfamethoxazole.
Resistance genes were frequently associated with mobile genetic elements like plasmids and integrons.
Predominant sequence types showed phylogenetic relatedness to poultry and environmental isolates from other African countries.
Abstract
Background: Intensive poultry production systems can act as reservoirs for antibiotic-resistant and multidrug-resistant (MDR) Escherichia coli, posing a public health risk through food and environmental transmission. Methods: This study investigated the genomic characteristics of antibiotic-resistant E. coli isolated from an intensive poultry production system in the uMgungundlovu District, KwaZulu-Natal, South Africa. Chicken litter, wastewater, and floor swab samples were collected over three consecutive production cycles. Putative E. coli isolates were detected using the Colilert-18 system, cultured on eosin methylene blue agar, and genomically confirmed by quantitative PCR (q-PCR) targeting the uidA gene. Whole genome sequencing was performed using the Illumina MiSeq platform, followed by bioinformatic analyses to assess resistance genes, mobile genetic elements, and phylogenetic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
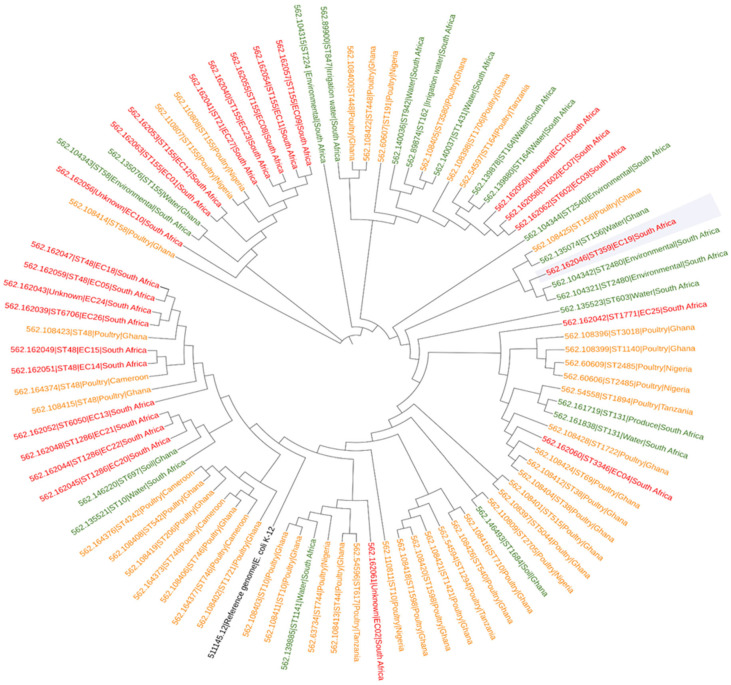 Figure 1
Figure 1- —South African Research Chairs Initiative of the Department of Science and Technology and National Research Foundation of South Africa
- —South African Medical Research Council (SAMRC) Self-Initiated Research Grant, and Welcome Trust
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Pharmaceutical and Antibiotic Environmental Impacts · Antimicrobial agents and applications
1. Introduction
Escherichia coli (E. coli) is a commensal bacterium commonly found in the gastrointestinal tract of warm-blooded animals [1]. It is widely used as an indicator organism because it is well characterised and relatively easy to isolate and study [2]. The detection of E. coli in food products and water is considered an indication of faecal contamination, which supports its use in monitoring hygiene and water quality [3]. Although E. coli is typically harmless, certain strains can become pathogenic under favourable conditions, causing disease that may require antibiotic treatment [4]. The excessive and inappropriate use of antibiotics contributes to the emergence of antibiotic-resistant bacteria, potentially rendering these treatments ineffective in the future [5].
A recent scoping review reported an increasing prevalence of antibiotic-resistant E. coli across Africa in both clinical and non-clinical settings, with prevalence rates reaching up to 98.3% [6]. High levels of resistance were observed against commonly used antibiotics, including ampicillin, tetracycline, and trimethoprim–sulfamethoxazole, across several African countries [6]. Similarly, a review by Ramtahal et al. [7] documented widespread resistance within the poultry industry to ampicillin, tetracycline, and sulfonamides, as well as to fluoroquinolones and third-generation cephalosporins. In the poultry industry, antibiotics are used for growth promotion, disease prophylaxis, and metaphylaxis and are typically administered orally at varying doses depending on their purpose [8].
The use of antibiotics in the intensive poultry production industry significantly enhances chicken growth rates, reduces disease prevalence, and provides benefits to both breeders and their flocks [9]. However, administering low doses of antibiotics to chickens for growth promotion creates selective pressure, which leads to the development of antibiotic-resistant bacteria [10]. The transmission of antibiotic resistance genes is facilitated by mobile genetic elements (MGEs), including integrons, insertion sequences, plasmids, and transposons [11]. Antibiotic-resistant E. coli has become a significant concern in intensive poultry farming, as it is frequently linked to animal disease, food safety risks, and public health implications [12]. Pathogenic strains such as avian pathogenic E. coli (APEC) are responsible for colibacillosis are found associated with poultry, and this leads to increased mortality in poultry and significant economic losses within the poultry industry [1].
The loss in profit puts a strain on the economy of low- and middle-income countries like South Africa, since the poultry industry is among the major contributors to agricultural revenue, accounting for 18% of the total agricultural gross value [10]. Recent data from the Department of Health, South Africa, reported a marked increase in antibiotic importation for animal use, reaching 2,488,754 tons and nearly doubling the amount used for human disease [13]. The spread of antibiotic-resistant E. coli is facilitated by such use, with potential dissemination through both animal products and the environment.
Antibiotic-resistant bacterial strains potentially spread to humans through occupational exposure or via the food chain [8,14]. Additionally, the transfer of antibiotic-resistant bacteria to soil and aquatic environments has been demonstrated through runoff of chicken litter used as fertilisers [15]. To gain a deeper understanding of E. coli’s genetic makeup and mechanisms of antibiotic resistance, next-generation sequencing (NGS) is essential. Whole genome sequencing (WGS) is particularly valuable as it provides detailed information on the bacterium’s genome, including pathogenicity and virulence factors [4].
To assess the prevalence and dissemination of antibiotic-resistant E. coli in an intensive poultry production system, E. coli genomes from chicken litter, wastewater, and floor swabs were characterised using whole genome sequencing and bioinformatics analysis to delineate the resistome, mobilome, and phylogeny.
2. Results
2.1. Prevalence of E. coli and Antibiotic Susceptibility Profiling
Out of 54 samples collected, 150 bacterial isolates were randomly selected from chicken litter and wastewater samples to constitute the final sample size, and 47% (n = 70) were confirmed as E. coli through molecular screening. Moreover, E. coli was not isolated from any of the floor swab samples. All confirmed E. coli isolates exhibited resistance to at least one antibiotic. Notably, all isolates were resistant to tetracycline (TET). Additionally, 94% (n = 66) showed resistance to ampicillin (AMP), while 76% (n = 53) were resistant to trimethoprim–sulfamethoxazole (SXT). The lowest resistance rate was observed against ciprofloxacin (CIP), in only 3% (n = 2) of isolates. No resistance was detected to gentamicin (GEN), meropenem (MEM), cefotaxime (CTX), ceftriaxone (CRO), ceftazidime (CAZ), or cefepime (FEP) (Table 1). The resistant E. coli isolates consistently exhibited similar resistance profiles across all sampling cycles, except for cycle 2 (December), during which two isolates showed resistance to ciprofloxacin (Table 1). Ciprofloxacin also had the highest proportion of intermediate susceptibility among the antibiotics tested, with 23% (n = 16) of isolates falling into this category. Multidrug resistance (MDR) was defined as, i.e., microbes, resistant to at least one antibiotic in three or more classes [16]. A total of 74% (n = 52) isolates were classified as MDR. The most common antibiogram pattern, AMP-SXT-TET, accounted for 71% (n = 50). AMP-TET-SXT-CIP and AMP-TET-CIP accounted for 1.4% (n = 1) each, as shown in Table 1.
2.2. Escherichia coli Antibiotic Resistance Genes
The whole genome analysis (Table 2) demonstrated that a higher number of isolates, 68% (n = 17), carried fluoroquinolone resistance genes in the genotypic analysis compared to the phenotypic AST results. The most frequently detected fluoroquinolone resistance gene was qnrS1, followed by oqxA and oqxB. One isolate (EC04) carried qnrB19, which is linked to plasmid-mediated quinolone resistance. Out of 17 isolates carrying fluoroquinolone resistance genes, eight isolates represented chicken litter, and the remaining nine represented wastewater. A total of 72% (n = 18) isolates carried the disinfectant gene sitABC, and 52% (n = 13) of sitABC genes were carried on isolates harbouring fluoroquinolone resistance.
Extended-spectrum β-lactamase (ESBL) genes were also identified, with multiple bla_TEM_ genes detected. Bla_TEM1B_ was the most common gene, followed by bla_TEM_-135 and the bla_LAP-1_ gene. A total of 36% (n = 9) of isolates carried ESBL-resistant genes, with a distribution of four isolates from chicken litter and five from wastewater. These results correspond with the AST results. Other resistance genes identified included those conferring resistance to aminoglycosides (aadA1, aadA2, aadA5, aph(3′)-Ia, aph(3″)-Ib, aph(6)-Id), bleomycin (bleo), chloramphenicol (cmlA1), fosfomycin (fosA3, fosA4), sulfamethoxazole (sul2, sul3), tetracycline (tetA, tetB, tetM), and trimethoprim (dfrA5, dfrA12, dfrA14, dfrA15, dfrA17). Isolates were further investigated for mutations in quinolone resistance determinant regions (QRDRs). The QRDRs investigated consist of the DNA gyrase (gyrA and gyrB) as well as the DNA topoisomerase IV (parC and parE) genes. In our study, only gyrA (S83L, D87N, D678E*, A828S*, P872S*) and parC genes (S80I, D475E*, T718A* K665R*) were identified. A total of eight isolates harboured both mutations in the QRDR, while one isolate harboured the gyrA mutation gene only. Among the identified QRDR mutations, there were known mutations with phenotypic effects known to cause resistance to nalidixic acid and ciprofloxacin (S83L, D87N). Those marked with an asterisk are novel mutations (Table 2).
2.3. Sequence Types and Plasmid Replicons
MLST analysis (Table 2) revealed a variety of sequence types (STs), with nine hits from litter and 12 hits from wastewater. These included ST155, 24% (n = 6), ST48, 16% (n = 4), ST1286, 12% (n = 3), and ST602, 8% (n = 2), as well as the following singletons: ST3346, 4% (n = 1), ST6050, 4% (n = 1), ST359, 4% (n = 1), ST1771, 4% (n = 1), ST6706, 4% (n = 1), and ST21, 4% (n = 1). The remaining 16% (n = 4) of the isolates had unknown STs. All isolates carried plasmid replicons that included IncF, IncFIB, IncFIC, IncFII, IncI2, p0111, IncI1-I(Alpha), IncX1, IncY, Col(pHAD28), IncFIA(HI1), IncFIB(AP001918), IncFIC(FII), IncHI1A, IncHI1B(R27), IncFIA, IncX4, IncQ1, IncB/O/K/Z, IncI(Gamma), Col440I, (pLF82-PhagePlasmid), and ColpVC. The distribution of plasmid replicons among chicken litter and wastewater isolates was even, with Inc being the most common plasmid replicon identified.
2.4. Mobile Genetic Elements Associated with ARGs in the Analysed E. coli Genomes
The isolates were further analysed for ARGs and MGEs on NCBI, and two isolates (EC03 and EC11) fell off due to large genomic size, resulting in a total of 23 isolates (Table 3). The main findings from the synteny analysis (Table 3) illustrated the distribution of MGEs across genomes. Almost all ARGs were associated with insertion sequences (ISs), except for EC10; others were associated with transposons, except for EC04, EC05, EC07, EC10, EC19, EC25, and EC26, and almost half of the ARGs were carried on integron class 1. A diversity of ISs was evident within isolates; two types of transposons, Tn3 and TnAs1, were observed, whereas integrons were represented by intl class 1 only.
2.5. Phylogenetic Tree Analysis
The phylogenetic tree provides insight into the genetic relatedness of E. coli isolates from South Africa and various African countries (Figure 1). The E. coli genomes from our study showed close genetic relation with poultry-derived E. coli from Cameroon, Ghana, Nigeria, and Tanzania; E. coli was isolated from the environmental samples, which included unspecified environment isolates, isolates in South Africa, unspecified water from Ghana and South Africa, irrigation water from South Africa, soil from Ghana, and produce from South Africa. These comparisons are based on genomes reported between 2015 and 2024. No data was found for 2025. The isolates appear to have originated from a common ancestor shared with the comparative strains but somehow branched out of the family tree to form new nodes. Multilocus sequence typing (MLST) analysis discovered that a lot of the isolates from our study clustered towards poultry isolates; only a few clustered towards the environmental isolates. Most of the E. coli isolates clustered into several sequence types, with ST67, ST48, ST155, and ST164 emerging as the dominant lineages within the intensive poultry production system. A small group of isolates (EC19, EC25, EC04, and EC02) clustered independently from the key poultry-associated E. coli cluster in the phylogenetic analysis, highlighting a greater genetic divergence.
2.6. Discussion
The study investigated the phenotypic and genomic characteristics of antibiotic-resistant E. coli from poultry litter and wastewater from an intensive poultry production system. No presumptive E. coli was isolated on all floor swabs sampled after disinfection, indicating an effective sanitation method and less risk of transfer of antibiotic-resistant E. coli strains from one poultry flock to the next production cycle.
2.6.1. Antibiotic Susceptibility Profile
To better understand resistance trends in poultry-associated E. coli, we examined the patterns of antibiotic resistance observed across multiple sampling cycles. The resistance patterns remained largely consistent across the cycles, with all E. coli isolates exhibiting relatively high levels of resistance to three antibiotics, i.e., tetracycline (highest prevalence), followed by ampicillin and trimethoprim–sulfamethoxazole. Among the December samples, two isolates (EC02 and EC03) exhibited resistance to ciprofloxacin (Table 1). These findings are consistent with a study by Phiri et al. (10) conducted in Zambia. In that study, broiler litter, cloacal swabs, and carcass swabs were collected from poultry farms, abattoirs, and open markets across seven districts to determine the prevalence and antimicrobial resistance profiles of Salmonella and E. coli. The E. coli isolates demonstrated high resistance to ampicillin (68%), tetracycline (8%), and trimethoprim–sulfamethoxazole (65%), while resistance to ciprofloxacin was comparatively lower (21%). These results are in contrast to the results found by McIver et al. [17] in a study conducted in a similar region to our study, on a commercial poultry farm in South Africa. The study assessed the antibiotic-resistant E. coli obtained from litter, faeces, wastewater from the chicken house, truck, crates used in transportation, and final meat products for consumption. The AST results demonstrated a low level of antibiotic resistance to AMP, TET, and STX. In comparison, in this study, the low level of fluoroquinolone resistance observed on ASTs was only evident in two isolates (EC02 and EC03) that were phenotypically resistant to CIP, even though ciprofloxacin ASTs did not correlate with the ARGs identified from the WGS results, where 68% of isolates carried fluoroquinolone-resistant genes. This could have resulted from the fact that under laboratory conditions, resistance genes may be present but not expressed; the presence of a resistant gene in a genome does not always indicate its expression [18].
2.6.2. Antibiotic Resistance and Mobile Genetic Elements
Table 3 reveals that above 98% of isolates were positive for the qnrS1 gene, which is a plasmid-mediated quinolone resistance (PMQR) gene. Almost all qnrS1 genes were carried on plasmid, except for EC12 (contig 41), which was carried on chromosome strain 67 (accession number CP128443.1). Antibiotic-resistant genes and MGEs from the E. coli were closely related to the target sequence found in a Genbank database (National Library of Medicine, National Center for Biotechnology Information, USA), with the most hits being plasmids (Table 3). Overall, 36% of the isolates carrying the qnrS1 gene also possessed mutations within the quinolone resistance-determining regions (QRDRs) of the DNA gyrase (gyrA) or topoisomerase IV (parC) genes. These mutations are known to confer resistance to fluoroquinolones, thereby reducing the effectiveness of this class of antibiotics. Isolates EC03 and EC07 were odd as they exhibited phenotypic resistance that did not correspond with genomic resistance. Isolate EC03 had an antibiogram of CIP-TET-SXT-AMP, and EC07 had TET-SXT-AMP, but both harboured the fosA3 resistance gene and had a fluoroquinolone point mutation gyrA S83L. Isolate EC25 exhibited TET resistance phenotypically, and aph(6)-Id, aph(3″)-Ib, tet(B) genomically, but it had a fluoroquinolone point mutation gyrA (S83L) and a novel parC (K665R).
The isolates’ resistant profile to fluoroquinolones had no correlation between the phenotypic and genomic results, which might have a novel mechanism or carry resistance genes that have not yet been annotated in the database [18]. A large number (68%) of isolates conferring resistance to fluoroquinolones was alarming, as it might render ciprofloxacin ineffective in the future. A review study conducted by [19] revealed that antibiotic-resistant E. coli within the poultry industry has increased above 60% globally since 2011 to 2024, with a multidrug resistance surpassing 50%, which is concerning.
Table 3 also demonstrated a wide range of ARGs associated with MGEs, indicating active and varied horizontal gene transfer among E. coli strains within the poultry farm. The MGEs identified included class 1 integrons (intI1), transposons (Tn3, TnAs1), and ISs (IS, ISKpn19). Most ARGs were carried; some were associated with one or more MGE. Five ARGs, dfrA, CmlA1, ANT(3″)-Ia, AadA5, and sul3, were carried on Intl1. Most dfrA- and AadA-resistant genes carried on intl1 were also associated with TnAs1, whereas ANT(3″)-Ia was either carried on intl1 alone or in association with TnAs1. This was observed on EC01, EC08, EC10, EC12, EC13, EC24, and EC27. The DfrA gene from EC21 and EC22 was carried on intl1 only. CmlA1 and sul3 were carried on intl1 and associated with IS256; refer to EC05, EC18, and EC20. Among the resistant genes harboured by EC02, EC05, and EC20, QacL was also identified. This gene encodes proteins found in the Qac (quaternary ammonium compound) efflux pump family, which contributes to antibiotic resistance. EC05 and EC18 also harboured the esX (ESAT-6 secretion system)-resistant gene, which confers resistance to macrolides and is known as a virulence factor in Mycobacterium tuberculosis [20]. The esX-resistant gene is also associated with the opportunistic M. avium complex (MAC), which causes a chronic infection of avian tuberculosis in birds. Even though modern husbandry has decreased the incidence of avian tuberculosis within the commercial poultry industry, there have been sporadic outbreaks reported in commercial poultry [21]. Another study conducted by Ogundare et al. [22] in South Africa, Gauteng and Limpopo provinces, aimed to determine the virulence profiles and AMR genes of zoonotic APEC, focusing on foodborne EHEC isolated from close human contacts, poultry, swine, and environmental water samples collected from abattoirs and poultry farms, and it also identified the esX gene on their isolates. This might indicate horizontal gene transfer.
2.6.3. Genetic Relatedness
The phylogenomic analysis from our study revealed that the majority of isolates in this study showed closer genetic similarity to poultry-derived E. coli than to environmental isolates. The isolates were closely related to poultry E. coli isolates found in Cameroon, Ghana, Nigeria, and Tanzania and environmental E. coli strains previously identified in South Africa and Ghana (Figure 1). A high number of isolates were closely related to poultry isolates from Ghana and Nigeria, whereas a few exhibited genetic relatedness to isolates circulating in Tanzania. Isolates from our study, such as EC15 (562.162049), EC18 (562.162047), had MLST (ST48), were similar to poultry isolates found in Ghana (562.108415) (562.108423) and Cameroon (562.164374), whereas ST155 was shared by isolates EC08 (562.162055) and EC23 (562.162040), as well as isolates from Nigeria poultry (562.110807 and 562.110808). ST48 is commonly associated with foodborne diseases, mostly prevalent in poultry meat [23]. ST155 is also prevalent in poultry; strains carrying ST155 often harbour different antibiotic resistance, like tetracycline, sulfonamides, and β-lactams [24]. ST155 (24%) was the most common sequence type in our study, followed by ST1286 (12%) and ST602 (8%). All ST155 isolates carried the tet(A)-resistant gene, which aligns with findings by Davies et al. [25], who similarly reported a high prevalence of ST155 and the associated presence of the tet(A)-resistant gene in E. coli from poultry in Bangladesh. The genetic relatedness observed among poultry E. coli suggests possible transmission.
3. Materials and Methods
3.1. Ethical Clearance
Ethical approval was obtained from the University of KwaZulu-Natal Animal Research Ethical Committee (AREC) (reference number AREC00002891/2021). Additionally, a Section 20A permit for researching animals was secured from the Department of Agriculture, Land Reform and Rural Development (DALRRD) (reference number 12/11/1/5 (2283AC)).
3.2. Study Site and Population
The study was conducted at an intensive poultry production system located in the uMgungundlovu District of KwaZulu-Natal, South Africa. The farm comprised 12 chicken houses, each housing over 25,000 broiler chickens. One chicken house was randomly chosen as the study site. Samples were collected at the end of each production cycle over three consecutive months: November 2023 for cycle one, December 2023 for cycle two, and January 2024 for cycle three.
Each cycle consisted of 35 days of chicken growth, followed by two days of chicken litter removal (samples of the chicken litter were collected before its removal), one day of chicken house washing (wastewater samples collected on the day of the wash), and one day for disinfecting (floor swab samples were collected 24 h after disinfection).
3.3. Sample Collection
A composite sampling strategy was carried out at the end of five weeks, after reaching a full growth production cycle. The chicken house was divided into three rows, and five chicken litter samples were collected from each row at approximately 5 m intervals, starting from the front and moving to the back of the house. This resulted in 15 chicken litter samples. To collect the faecal samples, a sterile disposable spatula and sterile zip-lock bags were used. The samples were stored on ice and transported to the Antimicrobial Resistance Unit (ARU), University of KwaZulu-Natal, Westville campus, for processing.
After two days, based on the hygiene maintenance program of the production, wastewater from the first wash was collected from the drain that allows water flow from inside the chicken house, before mixing with wastewater from the rest of the farm. Two wastewater samples were collected in a 2 L sterile bottle at 30 min time intervals each. Following 24 h of disinfecting the chicken house, the floor was swabbed using sterile swabs, which included swabbing the corners and the drains inside the house. Each sampling point was separated by five steps; the floor swab samples resulted in 75 swabs, which were then placed in one sterile bottle containing sterile distilled water (dH_2_O), making one sample per sampling cycle. Both wastewater and floor swabs followed the same storage and transportation procedure as previously described.
3.4. Isolation and Identification of E. coli
Escherichia coli was isolated and identified using a method previously described by Mbang et al. [26] with modifications [27].
For chicken litter, 1 g of solid litter was added to 9 mL sterile dH_2_O, briefly vortexed, and filtered. Wastewater was thoroughly mixed and filtered prior to use, and floor swab samples were placed on a shaker at 125 rpm for 2 h before further processing. A one in 100-fold dilution was conducted, where 1 mL of each sample was diluted into 100 mL of sterile distilled water. Subsequently, the putative presence of E. coli was determined using the Colilert system (IDEXX Ltd., Westbrook, ME, USA) according to the manufacturer’s instructions, with additional modification (1 g of chicken litter was mixed with 9 mL dH_2_O, filtered before using it in the Colilert system). A single Colilert reagent was added to each sample and mixed well before pouring the mixture onto a Colilert tray that was sealed and incubated at 37 °C for 18–24 h. After incubation, the trays were viewed under a UV light visualiser (Sigma-Aldrich, Steinheim, Germany) at 260 nm. The wells with blue fluorescence indicated putative E. coli presence. A random selection of wells that fluoresced from each tray was streaked onto a selective medium, Eosin Methylene Blue agar (EMB) (Merck, Darmstadt, Germany), for putative phenotypic identification and incubated at 37 °C for 18–24 h for each sample type. After incubation, two isolates were sub-cultured from EMB onto Nutrient agar (Oxoid, Hampshire, UK) to confirm culture purity and stored in Tryptic Soy Broth (Oxoid, Hampshire, UK) with 20% glycerol at −80 °C for further analysis. Each cycle provided 30 isolates from chicken litter, resulting in a total of 90 isolates for three sampling cycles. For wastewater, each sampling cycle resulted in 2 × 2 L samples that provided 20 isolates, resulting in 60 isolates for three consecutive sampling cycles. A total of 150 presumptive E. coli from chicken litter and wastewater were subjected to q-PCR for confirmation. No E. coli growth was isolated from floor swab samples. E. coli ATCC 25922 (Oxoid, Hampshire, UK) was used as a positive control.
3.5. Molecular Confirmation of Isolates
DNA was extracted using the heat lysis method as previously described by Abrar et al., [28]. The extracted DNA was utilised to confirm E. coli using the uidA gene on a QuantiStudio 5 RealTime PCR System (ThermoFisher Scientific, Waltham, MA, USA) [29] using the forward primer 5′-AAAACGGCAAGAAAAAGCAG-3′ and the reverse primer 5′-ACGCGTGGTTAACAGTCTTGC-3′, which targeted the uidA gene. All primers were acquired from Inqaba Biotechnical Industries (Pty) Ltd., Pretoria, South Africa. Optimised thermal cycling conditions for uidA (β-D-glucuronidase) included initial uracil–DNA glycosylase (UDG) activation at 50 °C for 2 min, activation of Dual-LockTM polymerase at 95 °C for 2 min, denaturation set at 95 °C for 15 s, annealing set at 60 °C for 15 s, and extension at 72 °C for 10 s for 35 cycles, followed by a final extension at 72 °C for 5 min. A high-resolution melting curve was generated by ramping up the temperature from 65 °C to 95 °C at a continuous rate of 0.15 °C/s. Each reaction included a positive control, E. coli ATCC 25922, as well as a negative template control, which consisted of nuclease-free water replacing the DNA template.
3.6. Antibiotic Susceptibility Testing (AST)
Confirmed E. coli isolates (n = 70), 47% were tested against a panel of ten antibiotics using the Kirby–Bauer disc diffusion method on Mueller–Hinton Agar (Oxoid, Hampshire, UK), following the Clinical and Laboratory Standards Institute (CLSI) guidelines [30]. The selection of ten antibiotics aimed to determine whether E. coli from the sampling site showed resistance to 3rd and 4th generation cephalosporins, carbapenems, and quinolones, classified as “shared class of antibiotics by animals and humans and also assess the state of antibiotics commonly used by the farm, e.g., ampicillin, trimethoprim-sulfamethoxazole, tetracycline [31]. The antibiotics tested were ampicillin (AMP, 10 μg), ceftriaxone (CRO, 30 µg), cefotaxime (CTX, 30 µg), cefepime (FEP, 30 µg), ceftazidime (CAZ, 30 µg) (CIA), ciprofloxacin (CIP, 5 µg), gentamicin (GEN, 10 µg), meropenem (MEM, 10 µg), tetracycline (TET, 30 µg), and trimethoprim–sulfamethoxazole (SXT, 25 µg). The diameters of the zones of inhibition were measured and interpreted using the breakpoint criteria provided by the CLSI (2020) [30]. E. coli ATCC 25922 was used as a positive control.
Out of the 70 confirmed E. coli isolates, 25 were selected for whole genome sequencing based on unique antibiograms and MDR profiles, along with two additional isolates that exhibited resistance to tetracycline only. The selection process accounted for all sampling site representation and the three sampling cycles.
3.7. Whole Genome Sequencing Analysis and Bioinformatic Analysis
Whole genome sequencing was conducted at the National Institute for Communicable Diseases (NICD), Johannesburg, South Africa. Genomic DNA from bacterial samples was extracted using the GenElute Bacterial Genomic DNA Kit (Sigma Aldrich, St. Louis, MO, USA) according to the manufacturer’s instructions. DNA concentration and purity were assessed at a 260/280 nm absorbance ratio using a Nanodrop 8000 (Thermo Scientific, Waltham, MA, USA). Subsequently, libraries were prepared using the Nextera XT DNA Library Preparation Kit (Illumina, San Diego, CA, USA) and subjected to whole genome sequencing on an Illumina MiSeq platform (Illumina, San Diego, CA, USA). Raw sequence reads were quality-trimmed using Sickle v1.33 (https://github.com/najoshi/sickle, accessed on 15 November 2024) and assembled using the SPAdes v3.6.2 genome assembler. The resulting genomes were submitted to GenBank and assigned accession numbers under BioProject PRJNA1183844. Genome annotation and analysis included identification of antibiotic-resistant genes (ARGs) and disinfectant genes using ResFinder v4.6.0 (https://cge.food.dtu.dk/services/ResFinder, accessed on 16 November 2024). Plasmid replicon typing was obtained via PlasmidFinder v2.1 (https://cge.food.dtu.dk/services/PlasmidFinder/ accessed on 16 November 2024). Multilocus sequence typing (MLST) was performed using the MLST 2.0 database (https://cge.cbs.dtu.dk/services/MLST/; accessed on 17 November 2024.
Mutations conferring resistance to fluoroquinolones and plasmid/chromosomal sequences with the closest nucleotide homology were analysed using BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE_TYPE=BlastSearch; accessed on 29 July 2025). The DNA gyrase (GyrA and GyrB) as well as the DNA topoisomerase IV (parC and parE) genes were analysed on BLAST using the E. coli ATCC 25922 strain as a reference. Finally, genome synteny and mobile genetic elements (MGEs) were examined on the National Center for Biotechnology Information (NCBI) platform (https://www.ncbi.nlm.nih.gov/, accessed on 6 January 2025).
3.8. Phylogenomic Analyses
A phylogenetic analysis was performed to investigate the relationships among E. coli genomes from this study and those reported in South Africa and other African countries from 2015 to 2024. The genomes were downloaded, annotated, and analysed from the Bacterial and Viral Bioinformatics Resource Centre (BV-BRC) (https://www.bv-brc.org/, accessed on 21 September 2025). The comparison included genomes derived from both poultry litter (n = 11) and wastewater (n = 12). The phylogenetic tree was constructed using the maximum likelihood method on BV-BRC. E. coli K-12 (accession: 511145.12) served as a genome reference. Tree annotation, editing, and visualisation were carried out using the Interactive Tree of Life (iTOL) platform (https://itol.embl.de/; accessed on 5 March 2025).
4. Conclusions
This study provides genomic insight into antibiotic-resistant Escherichia coli circulating within an intensive poultry production system in the uMgungundlovu District, KwaZulu-Natal, South Africa. Antibiotic-resistant E. coli were detected in samples from different sources, including faecal matter and wastewater, but they were not recovered from floor surfaces following disinfection. This suggests that the sanitation practices used were effective in reducing E. coli contamination on floors.
Findings from faecal matter and wastewater samples revealed a high prevalence of antibiotic-resistant poultry-associated E. coli. The isolates harboured multiple antibiotic resistance genes (ARGs) conferring resistance to commonly used classes of antibiotics. The detection of plasmid-mediated resistance genes, together with mutations in quinolone resistance-determining regions, indicates that both acquired and chromosomal mechanisms contribute to the observed resistance profiles.
Furthermore, the presence of diverse mobile genetic elements in poultry-derived E. coli highlights the potential for horizontal gene transfer within the poultry production environment. Overall, the results demonstrate that E. coli from intensive poultry systems are multidrug-resistant and carry numerous transmissible ARGs. The release of these resistant bacteria into the environment through faecal waste and wastewater following production cycles represents a significant public health concern.
5. Study Limitations
This study provides valuable genomic insight into antibiotic-resistant Escherichia coli within an intensive poultry production system; however, several limitations should be acknowledged. First, sampling was restricted to a single chicken house within one production system, which may limit the generalisability of the findings to other poultry farms, production systems, or geographic regions. Second, the study represents a snapshot in time, and the absence of time-resolved sampling limits the ability to assess temporal dynamics of antimicrobial resistance emergence, persistence, and dissemination across production cycles.
In addition, while isolates were collected from multiple sample types, detailed spatial metadata within the production environment was limited. This constrains robust inference regarding transmission pathways and the directionality of bacterial spread between poultry, waste streams, and the surrounding environment. Finally, although genomic analyses identified resistance-associated mutations, including a potentially novel mutation, functional validation was not performed, and, therefore, the phenotypic contribution of these mutations to antimicrobial resistance remains to be confirmed.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kathayat D. Lokesh D. Ranjit S. Rajashekara G. Avian Pathogenic Escherichia coli (APEC): An Overview of Virulence and Pathogenesis Factors, Zoonotic Potential, and Control Strategies Pathogens 20211046710.3390/pathogens 1004046733921518 PMC 8069529 · doi ↗ · pubmed ↗
- 2Ruiz N. Silhavy T.J. How Escherichia coli Became the Flagship Bacterium of Molecular Biology J. Bacteriol.2022204 e 00230-0022210.1128/jb.00230-2235916528 PMC 9487582 · doi ↗ · pubmed ↗
- 3Jang J. Hur H.G. Sadowsky M.J. Byappanahalli M.N. Yan T. Ishii S. Environmental Escherichia coli: Ecology and public health implications—A review J. Appl. Microbiol.201712357058110.1111/jam.1346828383815 · doi ↗ · pubmed ↗
- 4Mudau K.L. Ntobeng L.R. Kalu C.M. Tekere M. Pathogenicity and virulence factors of Escherichia coli discovered using next generation sequencing technologies and proteomics Front. Bacteriol.20254167777510.3389/fbrio.2025.1677775 · doi ↗
- 5Samreen Ahmad I. Malak H.A. Abulreesh H.H. Environmental antimicrobial resistance and its drivers: A potential threat to public health J. Glob. Antimicrob. Resist.20212710111110.1016/j.jgar.2021.08.00134454098 · doi ↗ · pubmed ↗
- 6Nnah E.P. Asante J. Amoako D.G. Abia A.L.K. Essack S.Y. Antibiotic-resistant Escherichia coli (E. coli) at one health interfaces in Africa: A scoping review Sci. Total Environ.202595817758010.1016/j.scitotenv.2024.17758039642619 · doi ↗ · pubmed ↗
- 7Ramtahal M.A. Amoako D.G. Akebe A.L.K. Somboro A.M. Bester L.A. Essack S.Y. A Public Health Insight into Salmonella in Poultry in Africa: A Review of the Past Decade: 2010–2020 Microb. Drug Resist.20222871073310.1089/mdr.2021.038435696336 · doi ↗ · pubmed ↗
- 8Burow E. Grobbel M. Tenhagen B.-A. Simoneit C. SzabóI. Wendt D. Kürbis C. Ladwig-Wiegard M. Banneke S. Käsbohrer A. Antibiotic Resistance in Escherichia coli from Broiler Chickens After Amoxicillin Treatment in an Experimental Environment Microb. Drug Resist.2020261098110710.1089/mdr.2019.044232915693 PMC 7482129 · doi ↗ · pubmed ↗