Antifungal Susceptibility Trends Among Filamentous Fungi: An Epidemiological Evaluation on Aspergillus spp., Fusarium spp., and Scedosporium spp. from Southern Italy
Maddalena Calvo, Marta Caccamo, Dalila Maria Cammarata, Laura Trovato

TL;DR
This study examines antifungal resistance trends in filamentous fungi from Southern Italy, revealing rising resistance to key drugs.
Contribution
The study provides new epidemiological data on antifungal susceptibility trends in Aspergillus, Fusarium, and Scedosporium species in Southern Italy.
Findings
Aspergillus fumigatus showed increased resistance to amphotericin B with 9.1% reduced susceptibility.
Lomentospora prolificans and Scedosporium apiospermum demonstrated multidrug resistance across tested antifungals.
Fusarium species exhibited variable susceptibility with higher MICs for azoles and amphotericin B.
Abstract
Background/Objectives: Antifungal resistance among filamentous fungi is an increasing global concern with significant implications for clinical management. Herein, we propose a study aiming to investigate in vitro susceptibility patterns and epidemiology of filamentous fungi in Southern Italy, focusing on MIC distributions and resistance trends. Methods: We reported susceptibility results from Aspergillus spp., Fusarium spp., and Scedosporium/Lomentospora spp. clinical isolates, which underwent azoles, echinocandins, and amphotericin B in vitro testing. Results: Aspergillus fumigatus was the most frequently isolated species, showing an alarming increase in reduced susceptibility to amphotericin B (9.1%). The highest MIC ranges for this antifungal drug emerged in the case of A. fumigatus (1–4 mg/L) and A. terreus (2–8 mg/L), while A. flavus (0.5–4 mg/L) and A. niger (0.25–4 mg/L) showed…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
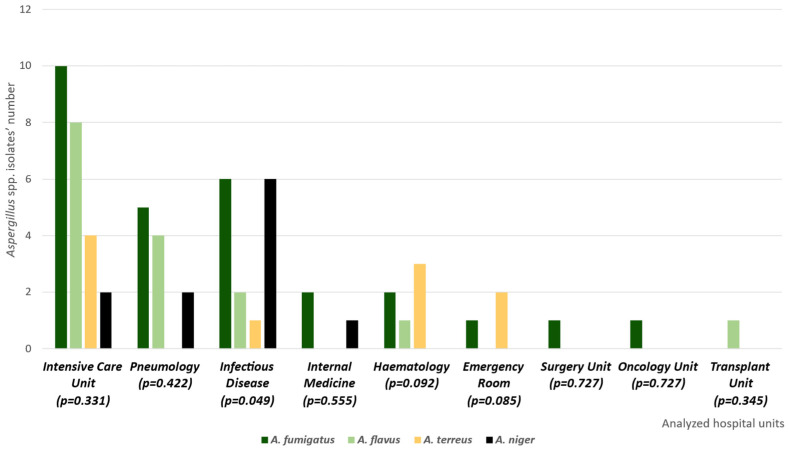 Figure 1
Figure 1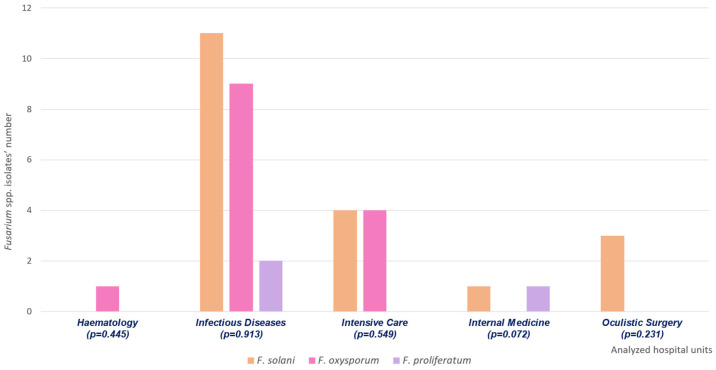 Figure 2
Figure 2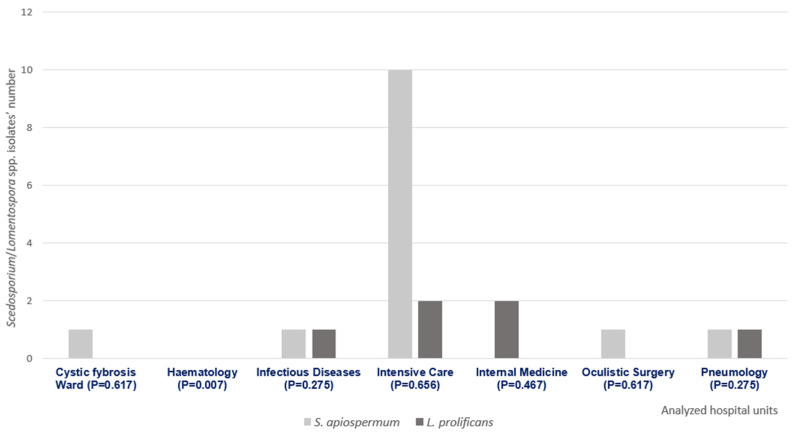 Figure 3
Figure 3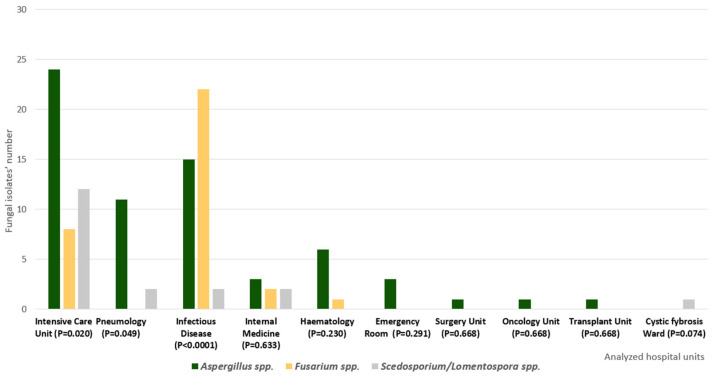 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Ocular Infections and Treatments · Nail Diseases and Treatments
1. Introduction
Invasive mould infections affect blood vessels, deep tissues, and organs after conidia inhalation or traumatic insertion. Respiratory transmission and cut or wound contaminations through fungal fragments correlate with mould infections [1]. Aspergillosis is one of the most common invasive mould infections, expressing several clinical forms, such as pulmonary aspergillosis, cerebral aspergillosis, and osteomyelitis, along with rare invasive conditions, such as endocarditis and gastrointestinal aspergillosis [2]. The World Health Organization’s latest report on alert isolates described Aspergillus fumigatus as the main clinically relevant species, counting it within the critical group of the priority list [3]. However, Aspergillus flavus, Aspergillus terreus, and Aspergillus niger significantly contribute to human pathogenesis, especially regarding respiratory infections [4].
Invasive pulmonary aspergillosis (IPA) expresses a consistent impact on patients reporting specific risk factors such as neutropenia, solid organ or stem-cell transplants, immunological impairments, corticosteroids therapies, mechanical ventilation, and chronic respiratory diseases [2,4]. As regards other severe fungal infections, invasive Fusarium spp. infections affect either immunocompetent or immunocompromised patients. Specifically, patients with leukemia or undergoing bone marrow transplants are possibly affected by dissemination episodes. On the other hand, immunocompetent patients may be injured by localized diseases such as keratitis [5]. The Fusarium solani complex (F. solani sensu stricto, F. keratoplasticum, F. falciforme) includes the most important human fusariosis aetiological agents, causing both bloodstream infections and keratitis [5,6,7]. Furthermore, Fusarium oxysporum has been isolated from keratitis and is invasive, while Fusarium proliferatum is an uncommon human pathogen rarely isolated in the case of respiratory infections [5,6,7,8]. According to these assumptions, the WHO integrated Fusarium spp. within the high-priority fungal pathogens [3]. Scedosporium spp. integrate the panel of possible rare mould aetiologies, including Scedosporium apiospermum complex (Scedosporium apiospermum sensu strictu, Scedosporium boydii) and Lomentospora prolificans [9,10]. All of these species colonize human respiratory airways, occasionally causing invasive infections among immunocompromised patients [10]. Moreover, Scedosporium spp. have been indicated as a frequent colonizing agent of cystic fibrosis patients’ respiratory tract [11]. The WHO guidelines included both Scedosporium apiospermum and Lomentospora prolificans among medium-priority fungal pathogens [3].
Despite difficulties in filamentous fungi management within the laboratory diagnostic routine, several methodologies have been applied for in vitro susceptibility testing. Clinical microbiology workflows mainly include gradient tests and broth microdilution [12,13]. The Clinical and Laboratory Standards Institute (CLSI) standardized performance instructions about filamentous fungi susceptibility testing [14], reporting clinical breakpoints and/or epidemiological cut-off values only for certain antifungal drugs against Aspergillus spp. [15]. The European Committee on Antimicrobial Susceptibility Testing (EUCAST) established comparable guidelines for Aspergillus spp. [16,17]. Unfortunately, all the above-mentioned documents do not include reference for Fusarium spp. or Scedosporium spp., except for quality control CLSI references [14,15].
Experimental studies highlighted some specific susceptibility patterns for the clinically relevant filamentous fungi. Aspergillus spp. reported azole resistance due to cyp51A gene mutations, which may alter the targeted 14-α-sterol demethylases. Membrane permeability variations and efflux pump overexpression contribute to the same resistance profile, which has been extensively related to environmental isolates and their exposure to fungicide azole agriculture usage. Echinocandin resistance may be related to 1,3-β-d-glucan synthetase gene mutations, even if these molecules are occasionally used because of their fungistatic activity against three spp. [18].
Fusarium spp. frequently exhibit intrinsic resistance to azoles, partly attributable to the reduced binding affinity of 14-α-sterol demethylase. By contrast, amphotericin B resistance is linked to changes in ergosterol content or composition of the fungal cell membrane and enhanced stress response mechanisms.
F. solani shows the most concerning susceptibility pattern, often including high amphotericin B MIC values and intrinsic azole resistance. Furthermore, echinocandins are not an appropriate treatment option in the case of fusariosis, due to the capability of Fusarium spp. to express 1,3-β-d-glucan synthetase gene mutations [19]. A multi-drug intrinsic resistance to azoles and amphotericin B characterizes Scedosporium spp. and L. prolificans. Scedosporium spp. reveal ergosterol modifications or efflux pump overexpression, while L. prolificans also produces hydrolytic enzymes able to inactivate antifungal drug structure. The previously cited echinocandin resistance mechanisms are constitutively present within Scedosporium spp. and L. prolificans [20]. Since several published studies have documented an emerging resistance phenomenon for filamentous fungi [21,22], it is essential to enrich the scientific literature about their susceptibility profiles. Herein we present an epidemiological analysis of Aspergillus spp., Fusarium spp., and Scedosporium spp., aiming to describe their in vitro antifungal susceptibility trends in Southern Italy.
2. Results
A total of 65 Aspergillus spp. isolates were collected during the study period. Among the 28 A. fumigatus strains, 13 emerged from bronchoalveolar lavage fluids (BALs), 5 from bronchial aspirates, and 10 from sputum samples. Eleven A. flavus samples were derived from bronchial aspirates, four from sputum samples, and one from BAL, globally accounting for sixteen strains. The analysis also included 11 A. niger isolates, isolated from BAL (6) and bronchial aspirates (5). We isolated 10 A. terreus samples from bronchial aspirates (4), BAL (3), and sputum samples (3). These respiratory isolates allowed an infection classification (probable pulmonary aspergillosis according to the European Organization for Research and Treatment of Cancer and the Mycoses Study Group Education and Research Consortium (EORTC/MSGERC) guidelines) [23,24]. Figure 1 illustrates the Aspergillus spp. isolate distribution within the included hospital units, along with a statistical analysis of this diffusion. This analysis demonstrates the statistical significance of the Aspergillus spp. distribution within the infectious disease unit. Furthermore, Table 1 summarizes antifungal susceptibility details for the same strains.
The study included 36 Fusarium spp. isolates, including 19 F. solani, 14 F. oxysporum, and 3 F. proliferatum. Figure 2 shows their distributions within different hospital units, along with a statistical analysis of the eventual significance of species distributions in specific wards.
Regarding F. solani, eleven isolates emerged from sputum samples, four from bronchial aspirates, three from corneal scrapings, and one from paranasal sinus biopsy. Nine F. oxysporum isolates were derived from sputum samples, two from bronchial aspirates, two from blood, and one from bronchoalveolar lavage fluids (BALs). Finally, all the F. proliferatum samples were from sputum. Corneal scraping and blood culture isolates were obtained from proven invasive fungal infection episodes resulting from deep localization and bloodstream invasion [23]. Otherwise, the respiratory isolates allowed an infection classification according to the EORTC/MSGERC guidelines [23]. These criteria define invasive respiratory infection based on clinical evidence (i.e., a reverse halo sign), patient rick factors (i.e., neutropenia or immunological impairments), and mycological evidence, such as Fusarium spp. isolation from any respiratory sample (sputum, bronchial aspirate, or BAL). Since our patients reported at least one clinical sign, one risk factor, and mycological culture isolation, their cases were all categorized as probable invasive respiratory mould infections. Table 2 summarizes MIC ranges, MIC50, and MIC90 of the tested antifungal drugs.
We collected 16 S. apiospermum and 4 L. prolificans. Figure 3 illustrates their distribution within the involved hospital wards and the corresponding statistical analysis. This analysis illustrates the statistical significance of the absence of Scedosporium/Lomentospora spp. isolates in the hematology unit.
As regards S. apiospermum, we isolated seven strains from BAL, four from bronchial aspirates, four from sputum samples, and one from corneal scrapings. All the L. prolificans strains emerged from respiratory samples (two from sputum, one from BAL, and one from bronchial aspirate). The above-mentioned criteria [23] allowed infection classification into a proven case, based on the positive result of corneal scraping, and several probable invasive respiratory infections, based on respiratory sample cultures. Table 3 reports the MIC ranges, MIC90, and MIC50 of the tested antifungal drugs against Scedosporium spp. isolates. Finally, Figure 4 illustrates the distribution of Aspergillus spp., Fusarium spp., and Scedosporium/Lomentospora spp. within the different hospital units in cases of invasive respiratory infection, along with a corresponding statistical analysis, which emphasizes the significance (p < 0.05) of intensive care and infectious disease units.
Table 4 and Table 5 summarize detailed MIC values for Fusarium proliferatum and Lomentospora prolificans, whose isolate numbers were not sufficient to establish MIC50 or MIC90.
3. Discussion
The present manuscript was designed to contribute additional data to the current literature on antifungal susceptibility trends among moulds, emphasizing the epidemiological scenario of Southern Italy and the MIC value distribution. We focused our attention on Aspergillus spp., Fusarium spp., Scedosporium apiospermum, and Lomentospora prolificans due to relevant documentation of their intrinsic susceptibility profiles against azoles and/or echinocandins [18,19,20]. Furthermore, all of these fungal pathogens have been included within latest WHO priority list, both for antifungal resistance rates and diffusion capability within critically ill patients [3].
This focus was motivated by the increasing frequency of antifungal resistance reported in filamentous fungi, especially involving Aspergillus fumigatus and resistance to triazoles. Several multicentre studies and epidemiological surveillance programmes have documented a progressive rise in triazole-resistant A. fumigatus isolates, often associated with prior antifungal exposure or the extensive use of azole compounds in agricultural settings [25]. Considering the first-line role of triazoles in the treatment of invasive aspergillosis, the clinical implications of this phenomenon are substantial.
As a consequence, amphotericin B has been proposed as a potential alternative therapeutic option in geographical areas characterized by a high prevalence of azole resistance [26,27]. In agreement with previous reports, A. fumigatus represented the most frequently isolated species among Aspergillus spp. in our collection.
Notably, our findings indicated a proportion of Aspergillus spp. isolates showing reduced susceptibility to amphotericin B (9.1%). Although amphotericin B has historically retained good activity against Aspergillus spp., this observation may suggest the presence of slight changes in local susceptibility patterns when compared to historical epidemiological data. Nevertheless, such findings should be interpreted with caution, as amphotericin B resistance in Aspergillus species remains incompletely understood and is considered a multifactorial phenomenon. Importantly, the MIC values recorded in the present study are largely consistent with those previously reported for Aspergillus spp. isolates originating from the same geographical area [28,29,30]. In this context, our results may provide a rationale for further investigations, including in vitro combination or synergy studies involving amphotericin B and other antifungal agents, which remain relatively underexplored for Aspergillus spp. isolates [28,29,30].
Fusarium spp. are extensively known for their elevated MIC values for azoles and amphotericin B, along with a demonstrated intrinsic resistance to echinocandins. Unfortunately, these moulds are often related to negative clinical outcomes due to difficult therapeutic management [31]. However, the genus includes heterogeneous species, and our data enforced this assumption. Specifically, amphotericin B and triazoles (voriconazole, posaconazole) revealed high MIC values for all the collected F. solani strains. On the other hand, F. oxysporum and F. proliferatum showed lower MIC values for the same antifungal drugs. Scientific and clinical case reports have highlighted the critical conditions related to L. prolificans and S. apiospermum infections [32]. These species frequently exhibit antifungal resistance, and our data confirmed the same criticism, revealing high MIC values for both of the above-mentioned species in the case of echinocandins, azoles, and amphotericin B.
Our study certainly has several limitations. Unfortunately, invasive fungal infections caused by filamentous fungi are significantly underestimated due to limited sensitivity rates within diagnostic protocols. Moreover Aspergillus spp., Scedosporium spp., and Scedosporium/Lomentospora spp. isolates are difficult-to-manipulate pathogens in most laboratory routines. They are often related to intense conidia aerosolization, technical expertise requirements, and dedicated specialized personnel. Consequently, culture isolation, identification, and antifungal susceptibility testing are highly complicated to perform. According to these assumptions, our study included a few isolates for each analyzed species. More studies with higher sample sizes will be necessary to further investigate in vitro mould susceptibility trends. Furthermore, molecular characterization of certain resistance mechanisms may clarify specific antifungal resistance trends.
Finally, susceptibility testing profiles, including ultimate antifungal drugs (e.g., isavuconazole and rezafungin), may be ideal to enforce epidemiological and clinical considerations.
In our opinion, our collected data highlight the necessity of continuously updating antifungal susceptibility trends for filamentous fungi. Regrettably, in vitro methodologies have not been sufficiently optimized for this purpose, and extended investigations should be conducted to achieve their official standardization in mould testing.
4. Materials and Methods
This study included a four-year (2021–2024) epidemiological analysis of all filamentous fungi isolated from clinical samples of patients, recovered at the University Hospital Policlinico, Catania (Italy). Specifically, the analysis included moulds from respiratory samples (bronchoalveolar lavage fluid, sputum, bronchial aspirate) and corneal scrapings. According to the routinary laboratory procedure, we inoculated these biological samples into Sabouraud Dextrose Agar with 2% of glucose, incubating the plates at 30 °C for 7 days. The MALDI Biotyper^®^ Sirius System mass spectrometry technology (Bruker, Billerica, MA, USA) identified the eventual filamentous fungi colonies. Specifically, fragments from grown colonies underwent a chemical extraction step involving ethanol, formic acid, and acetonitrile. Extracted fungal material subsequently went through centrifugation processes, collecting a supernatant. An aliquot of 1 µL was inoculated into the MALDI plate and covered with 1 µL of supplementary formic acid and α-cyano-4-hydroxycinnamic acid (HCCA) matrix. Regarding fungal species excluded from the MALDI identification panel, we identified L. prolificans based on microscopic and macroscopic morphological features, supporting our evidence with the absence of any growth in Sabouraud Dextrose agar with 2% of glucose and cycloheximide subcultures.
In vitro antifungal susceptibility was defined using Sensititre Yeast-One (Thermo Fisher Scientific, Waltham, MA, USA), with interpretation following the manufacturer’s instructions (https://documents.thermofisher.com/TFS-Assets/MBD/Package-Inserts/018-_PIYSTIVD-US-V3.2_CID9833.pdf, accessed on 4 January 2021). The inoculated plates were incubated at 37 °C for 48 h before verifying the antifungal susceptibility profile. This method has been clinically validated for yeast susceptibility testing, but it has also been extensively investigated for Aspergillus spp., Fusarium spp., and Scedosporium spp. in specific laboratory-validated contexts [33,34,35,36]. Additionally, previously published studies have highlighted the possibility of applying the Sensititre method to diagnostic workflows due to its elevated agreement with standardized broth microdilution [37,38,39,40,41].
We categorized the gathered MIC values according to filamentous fungi CLSI guidelines, reporting some clinical breakpoints (CBPs) for Aspergillus spp. isolates [15]. MICs higher than epidemiological cut-off (E-COFF) values were attributed to hypothetically resistant isolates in the absence of clinical breakpoints. For echinocandins tested against Aspergillus spp., minimum effective concentrations (MECs) were determined based on the presence of abnormal, short, and highly branched hyphal growth, rather than conventional MIC endpoints. All the resistance MIC values or the MIC values higher than the E-COFF were confirmed through the CLSI standardized broth microdilution method. In the cases of Fusarium spp., S. apiospermum, and L. prolificans, we did not classify the reported MIC values due to the absence of CBP and E-COFF.
Moreover, we reported MIC ranges for all the analyzed species, while MIC50 and MIC90 were calculated only for species with isolate numbers equal to or higher than 10 (Aspergillus fumigatus, Aspergillus niger, Aspergillus flavus, Aspergillus terreus, Fusarium solani, Fusarium oxysporum, and Scedosporium apiospermum). We followed these minimum conditions to guarantee statistically balanced isolate populations. The occurrence of trailing growth was mainly observed with azole antifungals and led to MICs, defined as the lowest drug concentration causing a significant reduction in growth compared to the drug-free growth control.
We performed a statistical analysis of Aspergillus spp., Fusarium spp., and Scedosporium/Lomentospora spp. within the included hospital units, using MedCalc Statistical Software version 17.9.2 (MedCalc Software bvba, Ostend, Belgium; http://www.medcalc.org; 2017; accessed on 9 January 2026) and reporting the corresponding p values. Specifically, χ^2^ and Fisher’s exact tests were applied to establish the categorical variables as percentages. The same statistical analysis finally assessed the distribution of Aspergillus spp., Fusarium spp., and Scedosporium/Lomentospora spp. genera within the different hospital units in cases of invasive respiratory infection.
5. Conclusions
The present study confirmed a concerning increase in filamentous fungi resistance percentages, particularly highlighting reduced susceptibility in A. fumigatus, F. solani, and L. prolificans. Our results emphasize the importance of enforcing epidemiological surveys, optimizing susceptibility testing, and performing precise species identification in the laboratory diagnosis of invasive fungal infection.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Available online: https://www.cdc.gov/fungal/about/about-invasive-mold-infections.html#:~:text=Overview,mold%20spores%20in%20the%20air(accessed on 31 October 2025)
- 2Fosses Vuong M. Hollingshead C.M. Waymack J.R. Aspergillosis Stat Pearls [Internet]Stat Pearls Publishing Treasure Island, FL, USA 2025 Available online: https://www.ncbi.nlm.nih.gov/books/NBK 482241/(accessed on 22 February 2023)29489184 · pubmed ↗
- 3WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action World Health Organization Geneva, Switzerland 2022
- 4Franconi I. Rizzato C. Ghelardi E. Lupetti A. Hospital distribution, seasonality, time trends and antifungal susceptibility profiles of all Aspergillus species isolated from clinical samples from 2015 to 2022 in a tertiary care hospital BMC Microbiol.20242411110.1186/s 12866-024-03267-838570761 PMC 10988875 · doi ↗ · pubmed ↗
- 5Nucci M. Anaissie E. Invasive fusariosis Clin. Microbiol. Rev.202336 e 00159-2210.1128/cmr.00159-2237937988 PMC 10732078 · doi ↗ · pubmed ↗
- 6Demonchy J. Biard L. Clere-Jehl R. Wallet F. Mokart D. Moreau A.-S. Argaud L. Verlhac C. Pène F. Lautrette A. Multicenter Retrospective Study of Invasive Fusariosis in Intensive Care Units, France Emerg. Infect. Dis.20243021522410.3201/eid 3002.23122138270146 PMC 10826781 · doi ↗ · pubmed ↗
- 7Ferreira da Cunha Neto J. da Silva Rocha W.P. Makris G. Sandoval-Denis M. Hagen F. Crous P.W. Chaves G.M. Fusarioid keratitis and other superficial infections: A 10-years prospective study from Northeastern Brazil P Lo S Negl. Trop. Dis.202418 e 001224710.1371/journal.pntd.001224738885283 PMC 11213298 · doi ↗ · pubmed ↗
- 8Benish M. Elitzur S. Arad-Cohen N. Barg A.A. Ben-Harosh M. Bielorai B. Fischer S. Gilad G. Levy I. Rosenfeld-Keidar H. Invasive Fusariosis in Pediatric Hematology/Oncology and Stem Cell Transplant Patients: A Report from the Israeli Society of Pediatric Hematology-Oncology J. Fungi 2022838710.3390/jof 8040387 PMC 903096335448618 · doi ↗ · pubmed ↗