Extracellular stimulation and ephaptic coupling of neurons in a fully coupled finite element-based Extracellular—Membrane—Intracellular (EMI) model
Karoline Horgmo Jæger, Aslak Tveito

TL;DR
This paper studies how neurons interact through extracellular fields using a detailed computational model, revealing how external stimulation and ephaptic coupling affect neural activity.
Contribution
A fully coupled finite element-based EMI model is used to study extracellular stimulation and ephaptic coupling in detailed neuron models.
Findings
Weak sinusoidal stimulation induces subthreshold membrane oscillations matching the stimulus frequency.
Extracellular stimulation modulates spike rates and timing depending on strength and distance.
Purkinje neurons synchronize ephaptically based on distance and extracellular conductivity.
Abstract
The extracellular potential surrounding neurons is of great importance: it is measured to interpret neural activity, it underpins ephaptic coupling between neighboring cells, and it forms the basis for external stimulation of neural tissue. These phenomena have been studied for decades, both experimentally and computationally. In computational models, variants of the classical cable equation for membrane dynamics and an electrostatic equation for the extracellular field are the most common approaches. Such formulations however, typically decouple the governing equations and therefore neglect the bidirectional coupling between the extracellular (E) space, the cell membrane (M), and the intracellular (I) space. We use a finite element-based Extracellular-Membrane-Intracellular (EMI) approach that solves a fully coupled system to study extracellular stimulation and ephaptic coupling in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
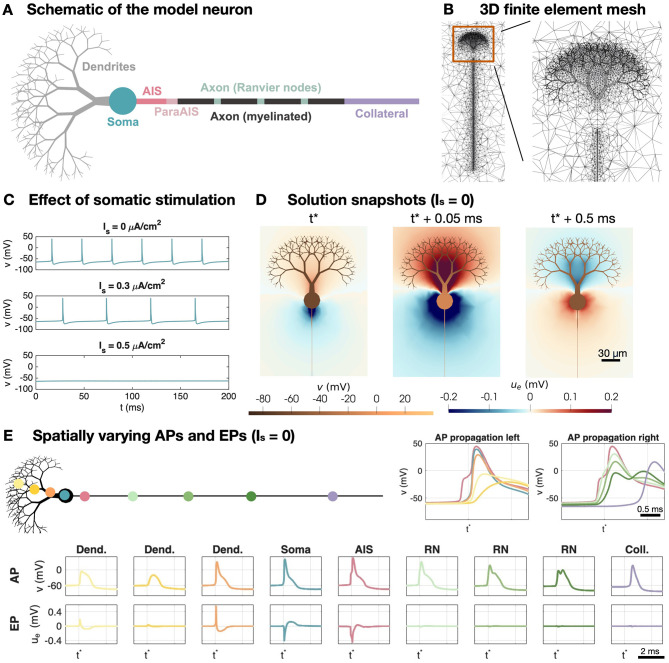 Figure 1
Figure 1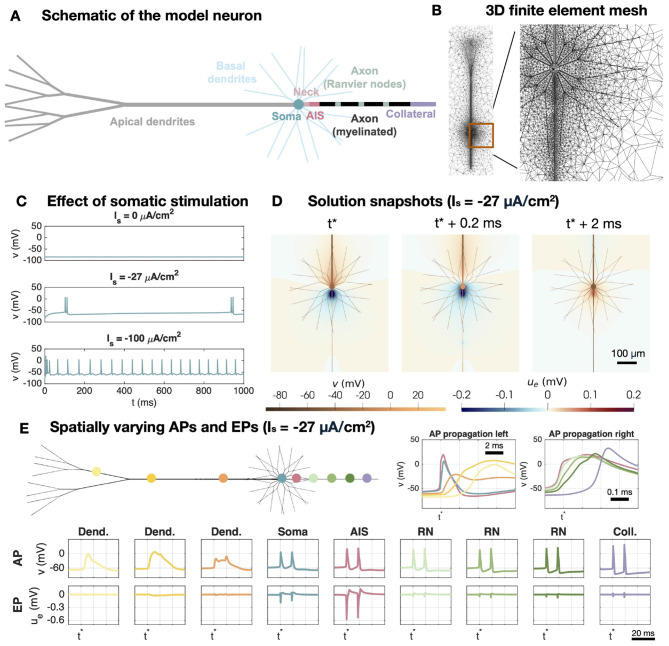 Figure 2
Figure 2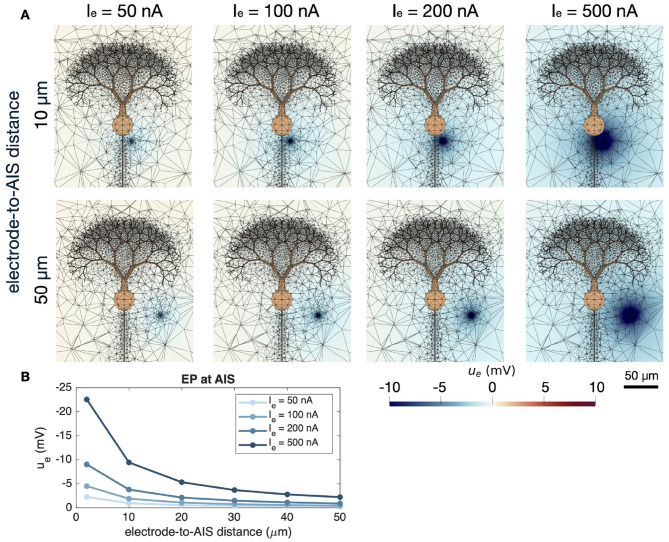 Figure 3
Figure 3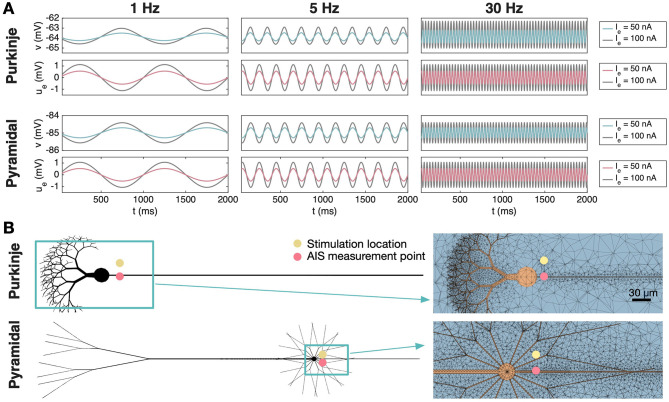 Figure 4
Figure 4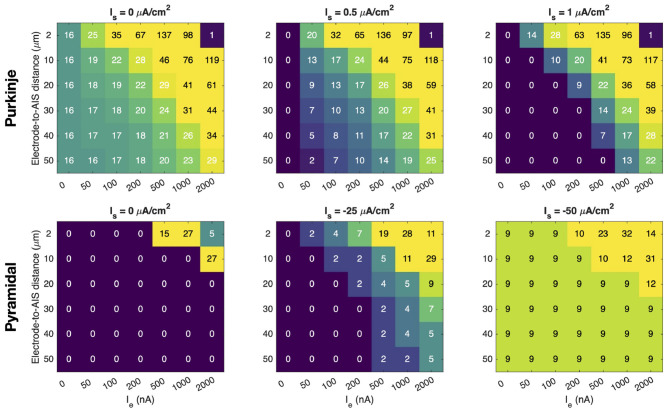 Figure 5
Figure 5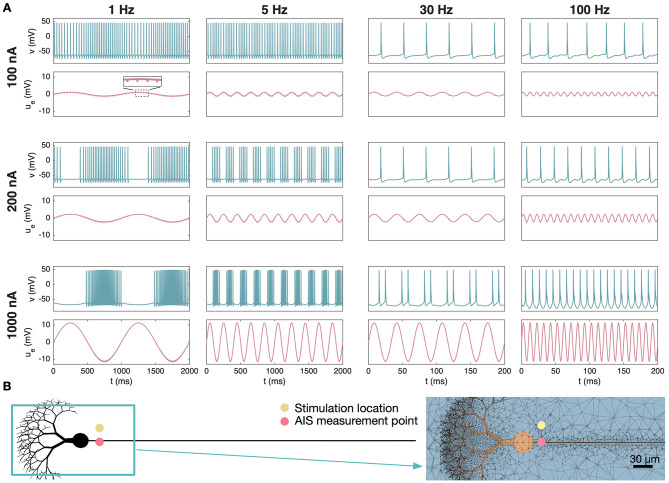 Figure 6
Figure 6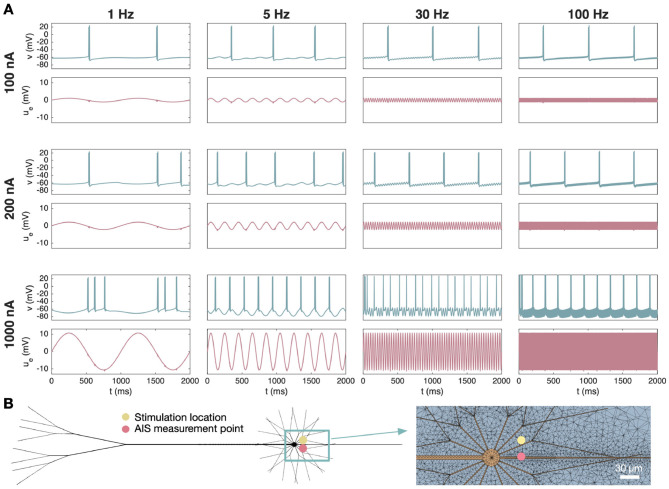 Figure 7
Figure 7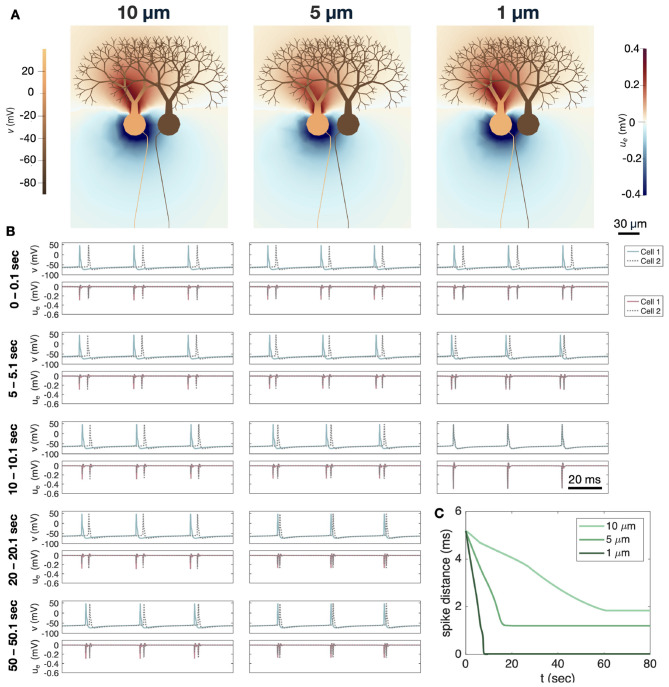 Figure 8
Figure 8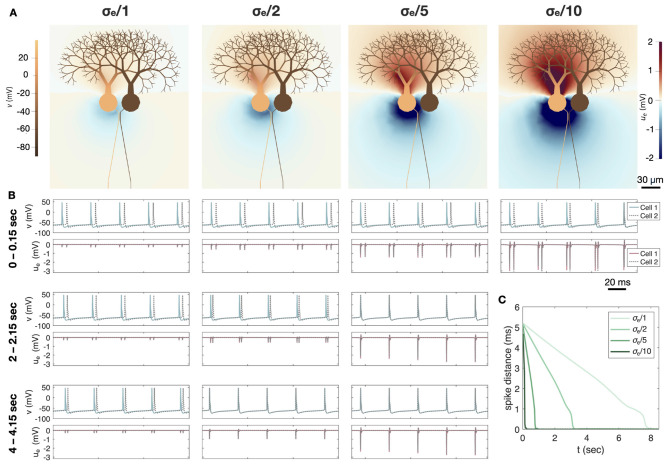 Figure 9
Figure 9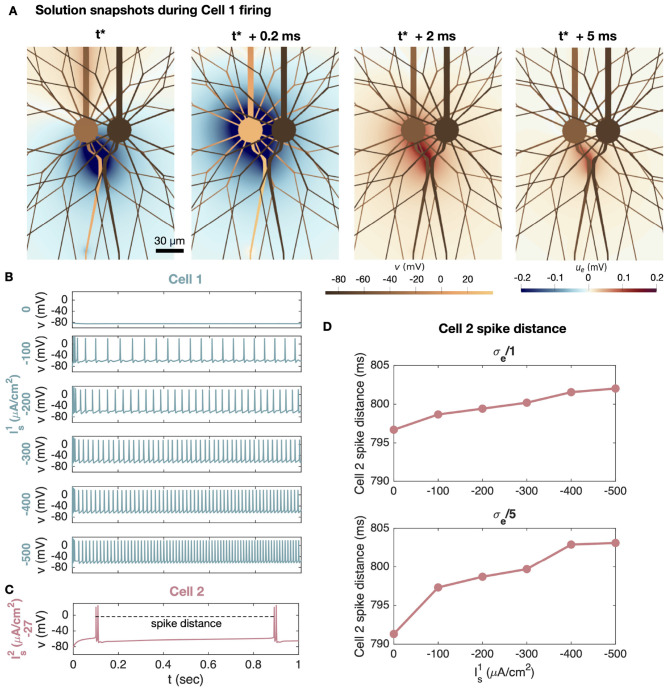 Figure 10
Figure 10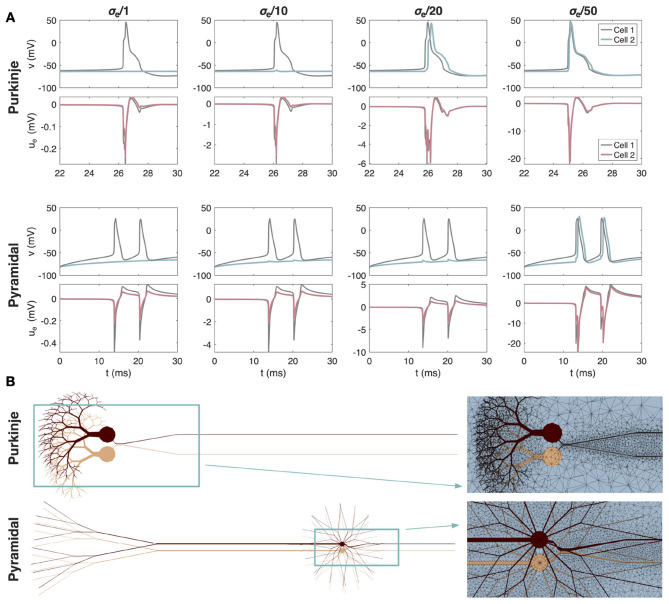 Figure 11
Figure 11Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVestibular and auditory disorders · Hearing, Cochlea, Tinnitus, Genetics · Neural dynamics and brain function
Introduction
1
The extracellular potential (EP) is of fundamental importance for investigating the properties and dynamics of neural tissue (Buzsáki et al., 2012; Hong and Lieber, 2019). It is also essential for understanding the biophysical mechanisms underlying externally applied fields, such as deep brain stimulation (DBS) (Ng et al., 2024; Lozano et al., 2019). The DBS technique has achieved substantial clinical success in treating movement disorders such as Parkinson's disease, dystonia, and essential tremor, and is increasingly applied to epilepsy, chronic pain, obsessive-compulsive disorder, Tourette syndrome, depression, and other neuropsychiatric conditions, with emerging trials targeting circuits involved in Alzheimer's disease, addiction, and post-traumatic stress disorder (Krauss et al., 2021; Zhang et al., 2024). Moreover, the EP is thought to play a role in the interaction of neighboring neurons that are not directly coupled through gap junctions or chemical synapses (Anastassiou et al., 2011; Stacey et al., 2015; Han et al., 2018). For instance, ephaptic interactions have been shown to contribute to inhibition (Blot and Barbour, 2014) and synchronization (Han et al., 2018) of Purkinje neurons, and to guide neocortical network activity (Fröhlich and McCormick, 2010). In addition, modeling studies have demonstrated that endogenous fields perturb spike thresholds and timing and can entrain spiking (Goldwyn and Rinzel, 2016; Moreno Cunha et al., 2024; Jæger and Tveito, 2025b).
In order to study these phenomena computationally, accurate and fully coupled models of the action potential (AP) and the associated EP is of considerable importance. Here, we apply a detailed computational model to quantify how external electrical stimulation modulates neuronal activity and to investigate how ephaptic coupling contributes to the interaction and synchronization of neighboring neurons.
Computational models of the EP surrounding neurons are most often based on a post-hoc, or open-loop, approach, in which the membrane potential and the EP are computed separately—and in that order (Holt and Koch, 1999; Shifman and Lewis, 2019). Solving the membrane equations under the assumption of a constant EP offers a substantial reduction in computational complexity. The EP can then be estimated from the transmembrane currents using analytical expressions based on line-source or point-source approximations (Rall, 1962; Pettersen et al., 2006; Telenczuk et al., 2020). These approximations provide useful estimates of local field potentials but neglect the bidirectional coupling between the cell membrane and the extracellular domain, which becomes important when cells are closely spaced, when strong external fields are applied, or when the aim is to understand ephaptic interactions.
In this study, we consider two neuron types: cerebellar Purkinje neurons and neocortical layer 5 pyramidal neurons. The membrane dynamics of these cells are described by models based on Masoli et al. (2024) and Hay et al. (2011), respectively. The Purkinje neurons exhibit autonomous pacemaking activity, whereas the layer 5 pyramidal neurons remain quiescent in the absence of external input. Together, the two cell types span a broad range of electrophysiological properties, providing complementary cases for studying endogenous field interactions and externally driven responses.
In the presentation of the models, we show how the two neuron types respond to electrical stimulation applied to the cell membrane of the soma. The Purkinje neuron is subjected to a positive stimulation current, where increasing current strength reduces the number of APs per second. Conversely, the layer 5 pyramidal neuron is subjected to a negative stimulation current, which induces increasingly frequent APs as the current strength increases. Next, we study how the Purkinje and pyramidal neurons are modulated by an external current source as the distance to the source, the current amplitude, and the stimulation frequency are varied. In addition, we investigate ephaptic interactions between neighboring cells. First, we examine how rapidly two neighboring Purkinje neurons synchronize through ephaptic coupling as the cell distance is increased or as the extracellular conductivity is reduced. Moreover, we examine how the firing pattern of one layer 5 pyramidal neuron affects the firing pattern of a neigboring neuron through ephaptic coupling, and whether a neuron can directly trigger excitation in a neighboring neuron through ephaptic coupling.
Our aim is to contribute to the understanding of external stimulation of neurons, as used in DBS, and of ephaptic interactions in neural tissue, at the level of individual cells. Clearly, DBS acts on large populations of neurons (Ng et al., 2024; Lozano et al., 2019), but the basic mechanism is already present at the single-cell level. Similarly, neural interactions and synchronization through ephaptic coupling is typically studied in large neuronal populations, yet it remains important to understand the underlying mechanism between just two neurons (Anastassiou et al., 2011; Stacey et al., 2015; Han et al., 2018; Moreno Cunha et al., 2024; Lee et al., 2024). This is the type of mechanistic understanding we aim to provide in the present study using the fully coupled EMI model.
In terms of modeling, we follow the strategy introduced in (Agudelo-Toro and Neef 2013) and further developed in Tveito et al. (2017b); Buccino et al. (2019); Jæger and Tveito (2025b), in which the dynamics of the extracellular (E) space, the cell membrane (M), and the intracellular (I) space are directly coupled through a system of partial differential equations (the EMI system). In Tveito et al. (2017b), the EMI model was used to evaluate the accuracy of the classical cable model combined with an analytical volume-conductor approximation for the EP, using a simple cuboid neuron geometry. The effect of neural probes (or microelectrode arrays) on the EP was addressed using the EMI model in Buccino et al. (2019). In Jæger and Tveito (2025b), the EMI model was used to demonstrate that a cell may exhibit a full action potential while producing a negligible EP in certain configurations. More recently, the EMI model was used to study the effect of myelination in axons (Gatti et al., 2026), and analytical solutions were derived to assess the numerical accuracy of EMI solvers in Ballesteros et al. (2025). In addition to neural applications, the EMI system has also been applied to study the dynamics of small collections of cardiomyocytes (Jæger et al., 2019, 2021a, 2024; Telle et al., 2023). The present paper builds on these developments by applying the fully coupled EMI formulation to detailed Purkinje and layer 5 pyramidal neuron geometries to study extracellular stimulation and ephaptic coupling across stimulation strength, distance, frequency, and extracellular conductivity.
Methods
2
The Extracellular-Membrane- Intracellular (EMI) model
2.1
We represent the electrical potential of neurons and their surrounding extracellular space using the Extracellular-Membrane-Intracellular (EMI) model (Agudelo-Toro and Neef, 2013; Tveito et al., 2017b; Jæger and Tveito, 2020).
The EMI model is given by the following system of equations:
Here, ui and ue (in mV) are the intracellular and extracellular potentials, defined in the intracellular and extracellular domains, Ω_i_ and Ω_e, respectively. Moreover, v = ui−ue_ is the membrane potential given on the membrane, Γ, which is defined as the interface between Ω_i_ and Ω_e. In addition, σi_ and σ_e_ (in mS/cm) are the intracellular and extracellular conductivities, respectively, ni and ne are the outward pointing normal vectors of Ω_i_ and Ω_e, respectively, Cm_ (in μF/cm^2^) is the specific membrane capacitance, and Is (in μ A/cm^2^) is a stimulation current density. Unless otherwise stated, Is = 0. Time, t, is given in milliseconds (ms). Furthermore, Iion (in μ A/cm^2^) is the current density through different types of ion channels on the cell membrane, s are gating variables and ionic concentrations involved in modeling these current densities, and F(t, v, s) model the dynamics of these additional state variables.
For the model neurons considered in this study, the density of the different types of ion channels vary for different locations of the neuron, like the soma, the dendrites and the axon initial segment (AIS). This results in a spatially varying expression for Iion, which is described in more detail in the following subsections and in Supplementary material.
In addition, we consider axons partially covered by myelin. In the myelin covered part of the membrane, both Iion and Cm are set to zero, resulting in the no-flux boundary conditions
on the myelinated membrane.
In our simulations, we let the neurons be surrounded by approximately 500 μm of extracellular space in spatial each direction. On the outer boundary of the extracellular domain, we apply the Dirichlet boundary condition
Cerebellar Purkinje neuron model
2.2
We model cerebellar Purkinje neurons using the EMI modeling setup described in Jæger and Tveito (2025b), based on the cable model formulation from Masoli et al. (2024). The Purkinje neuron is divided into seven different morphological parts, the dendrites, the soma, the axon initial segment (AIS), a ParaAIS, three Ranvier nodes separated by myelinated membrane sections and an axon collateral (see Figure 1A). The ion channel densities for each of these sections are described in Supplementary material. The three-dimensional (3D) finite element mesh of a neuron and the surrounding extracellular space is illustrated in Figure 1B.
Setup of the cerebellar Purkinje neuron model. (A) Schematic of the different geometrical parts of the model neuron. The membrane model differs between these parts as described in Supplementary material. (B) Illustration of the 3D finite element mesh used in the EMI model simulations. (C) Effect of somatic stimulation (Is) on the somatic membrane potential. For Is = 0, the cell fires APs spontaneously. For Is = 0.3 μA/cm2, the firing frequency is reduced and for Is = 0.5 μA/cm2, the AP firing ceases. (D) Solution snapshots of the membrane potential and EP in the case of no somatic stimulation. (E) APs and EPs at different locations of the neuron. The AP is initiated in the AIS and propagates to the left through the soma and dendrites and to the right through the axon.
In the absence of stimulation, the Purkinje neuron fires spontaneous APs. These are initiated in the AIS and propagate left through the soma and dendrites and right through the axon (see Figure 1E). In the soma and AIS clear negative EPs are generated during AP firing, whereas positive EPs are generated near the dendrites. Figure 1D provide spatial solution snapshots of the membrane potential and the surrounding EP at three points in time during AP firing.
If a positive stimulation current, Is, is applied in the soma of the Purkinje neuron model, the frequency of the AP firing is reduced, and if the stimulation is sufficiently strong (e.g., Is = 0.5 μ A/cm^2^), the AP firing is completely suppressed (see Figure 1C).
Neocortical layer 5 pyramidal neuron model
2.3
The model for neocortical layer 5 pyramidal neurons is set up in a similar manner as the model for cerebellar Purkinje neurons (see Figure 2). The model for the membrane dynamics is based on Hay et al. (2011), and the specific parameter values are specified in Supplementary material.
Setup of the neocortical layer 5 pyramidal neuron model. (A) Schematic of the different geometrical parts of the model neuron. The membrane model differs between these parts as described in Supplementary material. (B) Illustration of the 3D finite element mesh used in the EMI model simulations. (C) Effect of somatic stimulation (Is) on the somatic membrane potential. For Is = 0, the cell do not fire APs, but for negative somatic stimulation, APs are fired. (D) Solution snapshots of the membrane potential and EP in the case of Is = −27 μA/cm2. (E) APs and EPs at different locations of the neuron in the case of Is = −27 μA/cm2. The AP is initiated in the AIS and propagates to the left through the soma and dendrites and to the right through the axon.
The pyramidal neuron morphology consists of eight different parts, the apical dendrites, the basal dendrites, the soma, the axon neck between the soma and the AIS, the AIS, three Ranvier nodes separated by myelinated membrane sections and an axon collateral (see Figure 2A). The channel densities and geometry for each of these different sections are specified in Supplementary material. In Figure 2B, the 3D finite element mesh used to represent a pyramidal neuron and its surrounding extracellular space is illustrated.
In the absence of stimulation, the pyramidal neuron do not fire APs, but when the soma is stimulated by a sufficiently strong negative stimulation current, Is, APs are fired (see Figure 2C). When the strength of Is is further increased, the frequency of the AP firing increases.
In response to a somatic stimulation (Is = −27 μA/cm^2^), the AP is first initiated in the AIS and propagates left through the axon neck, soma and dendrites, and right through the axon, as depicted in Figure 2E. In the apical dendrite, the AP lasts considerably longer than in the soma and axon. More specifically, two AP spikes occur in the soma and axon during one spike in the apical dendrite for this somatic stimulation strength. During AP firing there are clear negative EP spikes near the soma and AIS, and the largest spikes are generated near the AIS, consistent with experimental observations in Bakkum et al. (2019). Figure 2D shows spatial solution snapshots of the membrane potential of the neuron and the surrounding EP near the soma and AIS during AP firing.
Stimulation protocols
2.4
In our simulations, we consider three different types of stimulation.
I: Constant somatic stimulation
The first considered type of stimulation is a constant membrane current density, Is (in μA/cm^2^), applied to the somatic part of the membrane. This membrane current density is included in the EMI model as described in (Equation 1). A positive Is reduces the neuron's excitability, whereas a negative Is increases the excitability of the neuron (see Figures 1C, 2C).
II: Constant extracellular stimulation
A second type of considered stimulation is an extracellular point source, representing extracellular stimulation through an electrode. This point source is represented by a sphere cut out of the mesh of the extracellular domain. On the surface of this sphere, a Neumann boundary condition of the form
is applied, where Ie (in μA) is the stimulation strength, Ae (in cm^2^) is the surface area of the sphere, and ne is the outward pointing normal vector of the extracellular domain (i.e., pointing toward the center of the stimulation sphere). For a positive Ie, a negative EP is emanating from the stimulation sphere. The stimulation sphere is located in the vicinity of the AIS and the distance will be reported for each simulation. The EP surrounding the Purkinje neuron at rest as a result of a constant extracellular stimulation is illustrated for a few choices of Ie in Figure 3A. In addition, the EP at an AIS membrane point as a function of the electrode-to-AIS distance is provided in Figure 3B in a similar manner as in Figure 1B in Lee et al. (2024) and Figure 1C in Anastassiou et al. (2011).
Extracellular potential generated by a constant extracellular stimulation source near a cerebellar Purkinje neuron. (A) Extracellular potential in a sheet in the center of the domain in the z-direction. In addition, the finite element mesh is indicated by black lines and the neuron is shown in orange. In the upper panel, the distance between the stimulation source and the AIS is 10 μm, and in the lower panel the distance is 50 μm. (B) Extracellular potential measured at an AIS membrane point of the Purkinje neuron as a function of the distance to the electrode.
III: Sinusoidal extracellular stimulation
The third considered stimulation type is a sinusoidal extracellular stimulation changing strength and sign with time during the simulation. This stimulation is implemented in the same manner as the constant extracellular stimulation, except that the boundary condition on the stimulation sphere is given by
where Ie (in μA) is the stimulation strength, Ae (in cm^2^) is the surface area of the stimulation sphere, f (in Hz) is the stimulation frequency, and t (in seconds) is time.
Numerical methods
2.5
We solve the EMI system of equations numerically using the spatial and temporal operator splitting technique introduced in Jæger et al. (2021b) with a global time step of Δt = 0.01 ms. We apply the MFEM finite element library (Anderson et al., 2021; MFEM, 2026) with first-order elements and a mesh generated using Gmsh (Geuzaine and Remacle, 2009). For the non-linear membrane model dynamics, we use the first order Rush Larsen method (Rush and Larsen, 1978) with code generated using the Gotran software (Hake et al., 2020) and a time step of Δt = 0.001 ms for the Purkinje neuron and Δt = 0.01 ms for the pyramidal neuron. The numerical code is available on Zenodo (https://doi.org/10.5281/zenodo.18183929; Jæger, 2026).
Results
3
In this section, we report results of our EMI model simulations of extracellular stimulation and ephaptic interactions of neurons. We begin by considering a single neuron in the vicinity of an extracellular stimulation source. We consider either a single cerebellar Purkinje neuron or a single neocortical layer 5 pyramidal neuron. The purpose of these simulations is to investigate how alterations in the EP (caused by, e.g., extracellular stimulation or by other firing neurons) can affect single-neuron dynamics. We start by considering subthreshold effects on the neuronal membrane potential. Then we move on to investigate how extracellular stimulation can modulate the number of AP spikes and the timing of AP spikes. Next, we focus on two neigbouring neurons that are not connected by synapses or gap junctions and investigate ephaptic interactions between the two cells.
Subthreshold membrane potential effects induced by weak extracellular stimulation
3.1
As a first investigation into the effects of extracellular stimulation on neurons, we consider a non-firing cerebellar Purkinje neuron and a non-firing neocortical layer 5 pyramidal neuron located near a sinusoidal extracellular stimulation source with relatively weak stimulation amplitudes. The Purkinje neuron is made quiescent by a somatic stimulation current of Is = 1 μA/cm^2^, while no somatic stimulation is applied for the pyramidal neuron. The extracellular stimulation source is located 20μm from the AIS of the neurons. Figure 4 shows the membrane potential and the EP in an AIS membrane point of the two neurons. We observe that the extracellular stimulation yields an EP of up to about ± 0.5 mV and ± 1 mV for the stimulation amplitudes of Ie = 50 nA and Ie = 100 nA, respectively, at the AIS membrane. This results in a similarly sized alteration in the AIS membrane potential of the two cells. These subthreshold membrane potential fluctuations are present for different applied stimulation frequencies (1 Hz, 5 Hz, and 30 Hz), consistent with previous experimental findings (Anastassiou et al., 2011; Lee et al., 2024).
Subthreshold effects at the AIS of a cerebellar Purkinje neuron and a neocortical layer 5 pyramidal neuron under sinusoidal extracellular stimulation. For the pyramidal neuron, no somatic stimulation (Is) is applied, whereas for the Purkinje neuron, Is = 1 μA/cm2 is used to maintain a stable resting membrane potential. (A) Membrane potential, v, and extracellular potential, ue, shown at an AIS membrane point for three different stimulation frequencies (1 Hz, 5 Hz, 30 Hz) and two different stimulation strengths (Ie = 50 nA and Ie = 100 nA). (B) Illustration of the simulation setup and a part of the 3D mesh used in the simulations. The AIS measurement point is indicated by a pink circle and the stimulation electrode is illustrated by a yellow circle. The distance between the stimulation electrode and the AIS is 20 μm.
Extracellular stimulation modulates neuronal spikes
3.2
Next, we investigate how extracellular stimulation modulates the dynamics of firing neurons.
Constant extracellular stimulation modulates the number of AP spikes
3.2.1
In Figure 5, we examine how a constant extracellular stimulation affects the number of AP spikes during a 500 ms simulation. Moreover, we investigate how this number is influenced by the extracellular stimulation strength, Ie, and the distance between the stimulation source and the neuron AIS.
Effect of constant extracellular stimulation on the number of spikes for a cerebellar Purkinje neuron and a neocortical layer 5 pyramidal neuron. For each extracellular stimulation strength, Ie, and electrode-to-AIS distance, the number of somatic spikes during a 500 ms simulation is reported. Three different levels of somatic stimulation, Is, are considered for each cell type.
In the upper panel of Figure 5, extracellular stimulation is applied near a Purkinje neuron, and in the left panel, no somatic stimulation is applied. If no extracellular stimulation is applied, the neuron fires 16 spikes during the 500 ms simulation. However, when a sufficiently strong extracellular stimulation is applied sufficiently close to the neuron, the spike frequency is increased. The number of spikes increases further as the extracellular stimulation strength is increased and as the distance between the stimulation source and the neuron decreases. Yet, when a very strong stimulation is applied close to the neuron (upper right corner), the stimulation is so strong that the neuron fails to repolarize, so there is no rapid AP firing. Moreover, for a distance of 2 μm and Ie = 1, 000 nA, the neuron approaches the no-repolarization state causing a reduced number of spikes compared to the case of Ie = 500 nA.
In the middle and right upper panels of Figure 5, a positive stimulation current, Is, is included across the membrane of the soma, making the cell quiescent in the absence of extracellular stimulation. However, if a sufficiently strong extracellular stimulation is applied sufficiently near the neuron, AP firing is initiated and, like in the left panel, the firing frequency increases as the extracellular stimulation strength is increased or the distance between the stimulation source and the neuron AIS is reduced. We also observe that, as expected, a stronger extracellular stimulation is required to initiate AP firing when the positive somatic stimulation is increased because this makes the cell less excitable (compare middle and right upper panels of Figure 5).
In the lower panel of Figure 5, we consider a layer 5 pyramidal neuron near the stimulation source. In the absence of extracellular stimulation, this neuron does not fire APs if there is no somatic stimulation or if a relatively weak somatic stimulation of Is = −25 μA/cm^2^ is applied. Yet, AP firing may be initiated if a sufficiently strong extracellular stimulation is applied sufficiently near the neuron (see lower left and middle panels of Figure 5). As expected, AP firing is more easily initiated if some negative somatic stimulation is present (middle panel) compared to the case of no somatic stimulation (left panel). In the right panel, the somatic stimulation is strong enough to initiate AP firing in the absence of extracellular stimulation, but the cell fires more APs if a sufficiently strong extracellular stimulation is applied sufficiently near the neuron AIS. Nevertheless, the layer 5 pyramidal neuron generally appears to require a stronger extracellular stimulation to affect AP firing than what is required to modulate the firing of the Purkinje neuron.
Sinusoidal extracellular stimulation modulates the timing of AP spikes
3.2.2
After considering a constant extracellular stimulation source, we now focus on AP spikes modulated by extracellular stimulation of a sinusoidal shape. Figure 6A shows the membrane potential and EP of a cerebellar Purkinje neuron near an extracellular stimulation source, and Figure 6B shows the simulation setup. We consider four different stimulation frequencies (1 Hz, 5 Hz, 30 Hz and 100 Hz) and three different stimulation strengths (Ie = 100 nA, Ie = 200 nA, and Ie = 1, 000 nA) applied 20 μm from the AIS. No somatic stimulation is applied. In the left panel of Figure 6A, a 1 Hz stimulation frequency is considered. We observe that this results in an EP of sinusoidal shape at the AIS with small extracellular spikes during AP firing (see the boxed inset in the upper left panel). For a relatively weak stimulation (100 nA), the AP spikes appear to occur more frequently when the EP is negative and less frequently when the EP is positive. When the amplitude of the extracellular stimulation is increased this tendency is even more pronounced, with no AP firing during the most positive values of the EP. For 5 Hz stimulation, similar behavior is observed. The 30 Hz stimulation, on the other hand, is more similar to the inherent firing frequency of the Purkinje neuron, and the cell appears to simply fire once each time the EP is negative. For the 100 Hz frequency, the stimulation effect seem to be limited for the low stimulation amplitude, whereas for the higher amplitudes the cell fires every (Ie = 1, 000 nA) or every other (Ie = 200 nA) time the EP is in a negative phase.
Effect of sinusoidal extracellular stimulation on the spike timing of a cerebellar Purkinje neuron. (A) The membrane potential, v, and the extracellular potential, ue, at an AIS membrane point for four different stimulation frequencies and three different stimulation strengths. In the extracellular potential traces, we observe small perturbations caused by the AP firing, which are magnified in the boxed inset for the 1 Hz, 100 nA case. Note that the scaling of the x-axis is different between the two cases of high frequency and the two cases of low frequency. (B) Illustration of the simulation setup and a part of the 3D mesh used in the simulations. The distance between the stimulation electrode and the AIS is 20 μm and no somatic stimulation, Is, is applied.
Figure 7 shows the results of similar simulations of a layer 5 pyramidal neuron in the vicinity of extracellular stimulation. In this case, we apply a somatic stimulation current of −27 μA/cm^2^, which is strong enough to trigger AP firing of the neuron, but only by a narrow margin. In this case, we observe that the frequency and strength of the extracellular stimulation has an effect on the timing and number of AP spikes, but the connection between extracellular stimulation and AP firing appears a bit less straightforward than for the Purkinje neuron.
Effect of sinusoidal extracellular stimulation on the spike timing of a neocortical layer 5 pyramidal neuron. (A) The membrane potential, v, and the extracellular potential, ue, at an AIS membrane point for four different stimulation frequencies and three different stimulation strengths. (B) Illustration of the simulation setup and a part of the 3D mesh used in the simulations. The distance between the stimulation electrode and the AIS is 20 μm, and the somatic stimulation, Is, is set to −27 μA/cm2.
Synchronization of two cerebellar Purkinje neurons
3.3
The next stage in our investigations is the study of ephaptic effects between two neurons. As a first example of such effects, we consider ephaptic synchronization of two firing cerebellar Purkinje neurons following up our preliminary analysis of this topic in Jæger and Tveito (2025b).
Synchronization of spiking Purkinje neurons depends on cell distance
3.3.1
In Figure 8, we explore potential synchronization of two firing Purkinje neurons for three different cell distances. From left to right, the minimal distance between the AIS of the two neurons is 10 μm, 5 μm and 1 μm. The axon of the neurons are shaped to bend toward each other in order to achieve these relatively small cell distances (see Figure 8A). No somatic or extracellular stimulation is applied, and the cells beat spontaneously. In order to study synchronization, we manipulate the initial part of the simulation such that the left neuron (denoted as Cell 1) fires APs approximately 5 ms before the right neuron (denoted as Cell 2). The cells are not connected by gap junctions or synapses, but the EP generated when one cell fires subtly alters the membrane potential of the other cell. In Figure 8, we observe that these subtle effects eventually cause the two neurons to start firing APs in synchrony if they are located sufficiently close. This was also observed in Jæger and Tveito (2025b).
Synchronization of two cerebellar Purkinje neurons for different cell distances. The shortest distance between the AISs of the two neurons is 10 μm, 5 μm and 1 μm in the left, middle, and right panels, respectively. (A) Solution snapshots of the extracellular potential (ue) and the membrane potential (v, color shown inside neuron) during AP firing of the left cell (Cell 1). (B) Membrane potential and extracellular potential in an AIS membrane point of each cell. Each panel row focuses on a different time interval during the simulation, and the columns correspond to the same three cell distances as in (A). Note that for a cell distance of 1 μm, the cells are completely synchronized after 10 s. Therefore, the plots for last time interval is omitted for this cell distance. (C) Temporal distance between AP spikes for the two cells as a function of time for the three different cell distances. Synchronization is considerably faster and the synchronized spike difference is considerably smaller for a short cell distance.
Figure 8A shows snapshots of the EP and membrane potential solutions during AP firing of Cell 1 for the three different cell distances. We observe that a negative EP emanates from the AIS and soma of Cell 1, causing a negative EP at the membrane of Cell 2. Note that during AP firing of Cell 2 a similar EP is generated from that cell. Figure 8B shows the membrane potential and EP at an AIS membrane point of each cell. The panel columns correspond to the three different considered cell distances and the rows represent different time intervals during a long simulation. In the upper row, we consider the solutions at the start of the simulation. In this case, Cell 2 fires APs approximately 5 ms after Cell 1. In the next row, the cells have been firing APs next to each other for 5 s. For a 10 μm cell distance, the solutions appear quite similar as those recorded in the start of the simulation, but for the smaller cell distances, the distance between the AP spikes of the two cells have clearly been reduced. In the third row, the cells have been firing APs next to each other for 10 s. Now, there is no longer any visible distance between the AP spikes for a 1 μm cell distance, implying that the cells are now synchronized. Note also that the amplitude of the EP spikes approximately doubles in this case because the EPs generated by the two cells add up. After 20 s of simulation, the distance between spikes is about 1 ms for the 5 μm cell distance. However, after 50 s, the spike distance appears to be approximately the same, indicating that the cells have reached a stable state of synchrony.
In Figure 8C, the results of Panel B are summarized. Here, the temporal distance between the spikes of the two cells are plotted as a function of time for the three considered cell distances. We observe that once the cells have reached a stable state of synchrony, the distance between spikes do not start to increase again; the cells remain synchronized. In addition, synchronization occurs considerably faster as the cell distance is reduced and the stable distance between spikes is shorter as the cell distance is reduced. This is consistent with the experimental observation of improved cerebellar Purkinje neuron synchronization for reduced AIS distances in Han et al. (2018).
Rapid synchronization of Purkinje neurons in presence of high extracellular resistivity
3.3.2
In Figure 9, we fix the AIS distance between the two cells at 1 μm and consider the case of increased extracellular resistivity. Since resistivity is the reciprocal of conductivity, this corresponds to a reduced extracellular conductivity, σ_e. We consider the default value of σe_ = 3 mS/cm (Einevoll et al., 2013) as well as three levels of reduction: σ_e/2, σe/5, and σe_/10. Like in Figure 8, the initial part of the simulation is manipulated such that Cell 1 (left neuron) initially fires APs approximately 5 ms before Cell 2 (right neuron).
Synchronization of two cerebellar Purkinje neurons in the presence of reduced extracellular conductivity, σe. The AISs of the two neurons are separated by a minimum of 1 μm of extracellular space. (A) Solution snapshots of the extracellular potential (ue) and the membrane potential (v) during AP firing of the left cell (Cell 1) for four different values of σe. The default physiological value of σe = 3 mS/cm is divided by 1, 2, 5, or 10 to represent different degrees of assumed σe reduction. (B) Membrane potential and extracellular potential in an AIS membrane point of each cell at different time intervals during the simulation. We consider the same four values of σe as in (A). Note that for σe/10, the cells are almost completely synchronized after only five spikes. Therefore, the plots for later time intervals are omitted for this value of σe. (C) Temporal distance between AP spikes for the two cells as a function of time for the four different values of σe. Synchronization is considerably faster for reduced values of σe.
Figure 9A shows snapshots of the membrane potential and EP during Cell 1 firing. We observe that the magnitude of the EP is considerably larger for a reduced extracellular conductivity. Note again, that a similar EP emerge from Cell 2 when that cell fires APs. Figure 9B shows the membrane potential and EP in an AIS membrane point of the two cells at a few different time intervals. When σ_e_ is reduced by a factor of 10, we observe that the two cells are almost completely synchronized already after only five AP spikes. For the remaining values of σ_e, synchronization takes a bit longer, but it occurs faster for small values of σe. In Figure 9C, these results are summarized by plotting the distance between spikes for the two cells as a function of time. We observe that for the default value of σe, synchronization takes about 8 s, for σe/2, it takes a little more than 3 s, and for σe_/5 it takes less than a second.
Ephaptic interactions between two neocortical layer 5 pyramidal neurons modulate spike timing
3.4
We now consider the case of ephaptic interactions between two neocortical layer 5 pyramidal neurons. In this case, we examine how the AP firing of a cell receiving substantial somatic stimulation modulates the AP firing of a nearby cell receiving weak somatic stimulation. Figure 10 shows such ephaptic interactions between two spiking pyramidal neurons. Like for the Purkinje neurons, we let the axon of the pyramidal neurons bend toward each other such that the minimum distance between the AIS of the two cells is 1 μm (see Figure 10A). The two cells are not connected by gap junctions or synapses. In Figure 10A, we show solution snapshots for a few time points during AP firing of the left cell (Cell 1). First, a negative EP is generated close to the AIS, and later toward the soma. Then, after a few milliseconds, a positive EP is present close to the soma and the AIS.
Ephaptic interactions between two neocortical layer 5 pyramidal neurons. The AISs of the two neurons are separated by 1 μm of extracellular space. (A) Solution snapshots of the extracellular potential (ue) and the membrane potential (v) during AP firing of the left cell (Cell 1). We zoom in on a part of the domain close to the soma and AIS. In this simulation, Cell 1 is stimulated by a somatic stimulation current of Is1=-200 μA/cm2. (B) Membrane potential in an AIS point of Cell 1 for different values of somatic stimulation, Is1, ranging from 0 to −500 μA/cm2. (C) Membrane potential in an AIS point of Cell 2. The somatic stimulation of Cell 2 is kept fixed at Is2=-27 μA/cm2 in all simulations. To quantify the ephaptic effect of Cell 1 stimulation on Cell 2 firing, we measure the distance between the first of the two double spikes of Cell 2 as indicated by the dashed line. In the simulation plotted in (C), Is1=0, and in (A–C) the default value of σe = 3 mS/cm is applied. (D) Cell 2 spike distance as a function of the somatic stimulation strengths of Cell 1 (Is1). We consider the default value of σe in addition to the default value reduced by a factor of 5 to represent assumed σe reduction. The spike distance is defined in (C).
In Figure 10B, we illustrate the membrane potential at the AIS of Cell 1 for a number of different values of somatic stimulation ( ) causing AP firing of different frequencies. As we vary the somatic stimulation of Cell 1, the somatic stimulation of Cell 2 is kept constant at A/cm^2^. The resulting Cell 2 AIS membrane potential is plotted in Figure 10C. In that simulation . However, we want to investigate whether another choice of somatic Cell 1 stimulation, , might alter the spike timing of Cell 2 even though the stimulation of that cell is kept the same. To that end, we define the spike distance for Cell 2 as illustrated in Figure 10C.
In Figure 10D, we report the spike distance of Cell 2 for different Cell 1 stimulation levels. We observe that as the stimulation strength of Cell 1 is increased and Cell 1 fires APs more frequently, the distance between spikes for Cell 2 increases by several milliseconds. We consider the default value of σ_e_ in addition to a reduced value of σ_e/5. The increase in Cell 2 spike distance as a result of rapid Cell 1 firing is more prominent for the reduced value of σe_. Since the stimulation of Cell 2 is kept constant and the two cells are not connected by gap junctions or synapses, these spike time alterations must be caused by ephaptic interactions between the neurons.
AP triggering through ephaptic coupling in the presence of high extracellular resistivity
3.5
As a final set of examples of ephaptic interactions between neighboring neurons, we consider the case of an AP being triggered in a neuron due to the EP generated from a nearby firing neuron. We consider the default value of σ_e_ = 3 mS/cm in addition to three cases of reduced conductivities. The setup used in the simulations is illustrated in Figure 11B.
Potential AP triggering through ephaptic coupling between two cerebellar Purkinje neurons and between two neocortical layer 5 pyramidal neurons. (A) Membrane potential (v) and extracellular potential (ue) in an AIS membrane point plotted for each cell and four different values of the extracellular conductivity, σe. The default physiological value of σe = 3 mS/cm is divided by 1, 10, 20, or 50 to represent different degrees of extensive σe reduction. Note that the scaling of the y-axis for ue differs for each value of σe. (B) Illustration of the simulation setup and a part of the 3D mesh used in the simulations. The AISs of the two neurons are separated by 1 μm of extracellular space. For the Purkinje neurons, Cell 1 is not stimulated and fires spontaneous APs. Cell 2 is made quiescent by a somatic stimulation current of Is2=0.7μA/cm2. For the pyramidal neuron, Cell 1 is stimulated by a somatic stimulation current of Is1=-50μA/cm2, causing AP firing, while Cell 2 is stimulated by a weaker somatic stimulation current of Is2=-25μA/cm2, which is not enough in itself to trigger AP firing. For both cell types, the extracellular potential generated during Cell 1 firing is strong enough to trigger AP firing in Cell 2 when σe is very small.
For the pyramidal neurons, we let Cell 1 fire APs as a result of a somatic stimulation current of A/cm^2^. Cell 2 is given a weak somatic stimulation current of A/cm^2^, which is not strong enough to trigger AP firing in itself. In Figure 11A, we plot the membrane potential and EP in an AIS membrane point of each cell. We observe that for the default value of σ_e, APs are fired in Cell 1, but not in Cell 2. The EP generated during Cell 1 firing reaches a value of about −0.5 mV at the AIS of Cell 1 and about −0.2 mV at the AIS of Cell 2. The corresponding perturbation of the membrane potential, v = ui−ue, of Cell 2 is not large enough to cause AP firing. As the value of σe_ is reduced, the amplitude of the EP is increased, and we see clear perturbations of the membrane potential of Cell 2. Yet, even for σ_e_ reduced by a factor of 20, this perturbation is not large enough to trigger an AP. However, when σ_e_ is reduced by a factor of 50, the membrane potential perturbation of Cell 2 caused by the EP generated during Cell 1 firing is large enough to trigger AP firing at Cell 2 as well.
For the Purkinje neurons, Cell 1 does not receive any somatic stimulation current and fires spontaneous APs, while Cell 2 is given a positive somatic current of A/cm^2^, which makes the cell quiescent on its own. However, if the extracellular conductivity is very low (default σ_e_ divided by a factor of 20 or 50), the negative EP generated from Cell 1 firing is enough to cause firing of Cell 2 as well (see Figure 11A).
Discussion
4
This work examines how neurons respond to extracellular stimulation and how local EPs can transmit signals ephaptically from one cell to adjacent cells. Our study is based on a finite element implementation of the EMI model (Tveito et al., 2017b; Jæger and Tveito, 2025b; Agudelo-Toro and Neef, 2013), developed using the MFEM finite element software package (Anderson et al., 2021), which allows us to study both endogenous field interactions and responses to imposed stimulation within the same modeling framework. We examine two contrasting cell types—autonomously firing Purkinje neurons and quiescent layer 5 pyramidal neurons. The membrane dynamics of the two neuron types follow Masoli et al. (2024) and Hay et al. (2011), respectively.
A fully coupled EMI formulation is particularly useful when (i) micrometer-scale extracellular structure is central to the question (tight cell–cell appositions, narrow clefts, or strong geometric confinement), (ii) σ_e_ is reduced relative to bulk values (or is spatially heterogeneous), and/or (iii) one studies long-duration stimulation/entrainment where small spike-timing differences can accumulate over time. In these regimes, the EP is not merely an output quantity but can contribute non-negligibly to the effective electrical load seen by the membrane. To illustrate this point, Supplementary Figure S1 compares EMI to an open-loop MI+E formulation and shows that the discrepancy grows substantially as σ_e_ is decreased; when σ_e_ is reduced by a factor 10, MI+E predicts markedly delayed firing relative to EMI in otherwise identical setups. In the two-cell setting, the same low-σ_e_ regime is also where EMI predicts ephaptic synchronization in our configuration, while the open-loop MI+E workflow does not capture this behavior.
Ephaptic effects are not expected to be uniform over the cell surface. They are strongest where (i) the extracellular space is most confined and (ii) membrane excitability is highest. In the present models, this naturally points to the axon initial segment (AIS), where a high sodium channel density can make the membrane particularly susceptible to field-induced modulation. A practical advantage of the EMI framework in this context is that the EP is computed with the correct local geometry and coupled back to the membrane dynamics at the same spatial location, without assuming that the local extracellular field in confined microdomains can be represented adequately by a bulk or far-field approximation, or that its effect can be captured accurately by post-processing. An approach for computing the EP is often based on an analytical solution that relies on assumptions that clearly do not hold here (an infinite, uniform domain). Furthermore, even when the qualitative behavior is unchanged, the closed-loop formulation can yield systematic quantitative differences in spike timing and entrainment in geometrically confined or low-σ_e_ regimes (Supplementary Figure S1).
The potential decays as 1/r from a current source
4.1
A first step in examining how neurons can influence each other ephaptically is to understand how a single neuron responds to an external current source. This is also of independent interest, as it forms the basis for electrical stimulation techniques used in a range of experimental and clinical settings. The impact of an extracellular current source on a neuron has been investigated experimentally, see, e.g., Anastassiou et al. (2011) and Lee et al. (2024), and the spatial decay of the potential is known analytically from the solution of the electrostatic equation Holt and Koch (1999). If r denotes the distance in the extracellular space to a point current source, the resulting potential decays as 1/r, a pattern illustrated in Anastassiou et al. (2011) and Lee et al. (2024). The lower panel of Figure 3 follows this decay profile.
The subthreshold membrane potential follows the stimulus frequency
4.2
In Figure 4, a weak extracellular current source modulates the subthreshold membrane potential of a nearby neuron, and the resulting oscillation follows the sinusoidal frequency of the stimulus current. The amplitude of the membrane response increases in proportion to the strength of the applied current source. This type of subthreshold frequency-following, with an approximately linear dependence on stimulus amplitude, is consistent with experimental and modeling studies of neurons exposed to weak oscillatory electric fields (Anastassiou et al., 2011; Lee et al., 2024; Deans et al., 2007).
Spike frequency is modulated by the strength and distance of a constant current source
4.3
In Figure 5 we show how the spike frequency is affected by the strength of an external stimulation source and the distance to this source. For the Purkinje neuron, we first consider the case without somatic stimulation, i.e., Is ≡ 0, see (Equation 1). In this setting, the number of spikes during a 500 ms simulation increases as the extracellular current becomes stronger and as the distance to the source decreases. However, for sufficiently strong stimulation at sufficiently close distances, the AP fails to repolarize properly, and the number of spikes is reduced to a single spike.
Next, we add a positive somatic membrane current to the Purkinje neuron, thereby reducing its excitability. This results in fewer spikes, while the dependence on the strength of the extracellular current source and the distance to the source remains similar to the case without added membrane current. Doubling the additional membrane current continues this trend and yields substantially fewer spikes.
The neocortical layer 5 pyramidal neuron is quiescent in the absence of membrane stimulation (i.e., Is ≡ 0). In the lower part of Figure 5, we note that for this cell type the extracellular stimulus must be both close and strong in order to initiate spikes. When a sufficiently strong negative Is is applied to the membrane, spikes are generated, and still more appear when the extracellular stimulus is close and strong.
Spike frequencies are modulated by the frequency of a sinusoidal current source
4.4
In Figures 6, 7 we show how a sinusoidal extracellular current source affects the membrane potential and EP of the two neuronal models. For the Purkinje neuron, a weak low-frequency stimulus has limited impact on the membrane potential, but as the stimulation strength increases the timing of the AP firing is influenced in a phase-dependent manner by the sinusoidal drive. When the phase of the stimulation current produces a sufficiently positive perturbation in the EP, the membrane potential becomes silent and no spikes are generated, whereas the spiking frequency is increased in the opposite phase. These effects are also clear for a 5 Hz stimulation frequency. When the frequency is increased to 30 Hz or 100 Hz, low stimulation amplitudes again have limited impact on the spikes, whereas strong stimulation suppresses spiking during phases of strongly positive EP and activate spiking during phases of negative EP.
In Figure 7, the neocortical layer 5 pyramidal neuron is subjected to a weak somatic membrane stimulation in addition to the sinusoidal extracellular stimulation. We observe that the sinusoidal stimulation induces a combination of occasional spikes and subthreshold modulation rather than regular firing at the drive frequency. Nevertheless, spikes seem to preferentially be generated when the EP is negative corresponding to a negative phase of the stimulation current.
These findings are consistent with experimental studies showing that weak oscillatory electric fields modulate spike timing and entrainment in a cell-type-dependent manner (Lee et al., 2024) and that transcranial alternating current stimulation (tACS) can phase-lock single-neuron firing in vivo (Krause et al., 2019). They are also consistent with modeling work based on the Pinsky–Rinzel neuron model (Pinsky and Rinzel, 1994), which demonstrates changes in firing sensitivity and activity patterns under AC-induced electric fields (Yuan et al., 2025).
Ephaptic interactions between neighboring neurons
4.5
In both cardiac electrophysiology and neuroscience, the possibility that one cell can excite its neighbor without direct coupling via gap junctions or chemical synapses has been extensively discussed, see, e.g., Han et al. (2018); Blot and Barbour (2014); Anastassiou and Koch (2015); Bokil et al. (2001) for neural tissue and Veeraraghavan et al. (2014); Gourdie (2019); Jæger et al. (2023) for cardiac tissue. For both organs, this question goes to the core of how tissue organizes excitation waves and is therefore of considerable interest. Here, we assess ephaptic coupling for the two neurons under consideration.
In Figures 8, 9 we follow up the analysis presented in Jæger and Tveito (2025b), where we showed that two adjacent cerebellar Purkinje neurons synchronize using the modeling framework applied here. In Figure 8 we illustrate how the APs of two neighboring Purkinje neurons synchronize and how this process depends on the distance between the cells. When the cells are close (1 μm measured at the AIS), the synchronization is rapid and the two APs become completely synchronized. However, when the distance increases to 5 μm, the APs synchronize only partially and reach a stable state in which the spike firing of the two neurons is separated by about 1.2 ms. When the distance is increased to 10 μm, a similar behavior is observed, but the converged distance is about 1.8 ms. The emergence of near-synchronous firing between neighboring Purkinje cells in our simulations is consistent with in vivo recordings showing sub-millisecond, tightly synchronized activity of nearby Purkinje cells and population-wide spike alignment (Han et al., 2018; Sedaghat-Nejad et al., 2022), and with modeling studies demonstrating that decreasing inter-neuronal spacing or increasing packing density strengthens ephaptic interactions and promotes spike-time synchrony (Stacey et al., 2015; Goldwyn and Rinzel, 2016).
In Figure 9 we show how the speed of synchronization depends on the value of the extracellular conductivity, σ_e. Panel C demonstrates that synchronization is strongly affected by σe_ in the sense that reducing σ_e_ markedly increases the speed of synchronization. This is consistent with the observation in Tveito et al. (2017b) that the ephaptic current scales as O(1/σ_e). A similar sensitivity of ephaptic effects to the properties of the extracellular space has been reported in computational studies: in networks of ephaptically coupled neurons, stronger ephaptic potentials and reduced inter-neuronal spacing promote spike synchronization and microcluster formation (Stacey et al., 2015), in axon bundles, higher extracellular resistivity (equivalently, lower conductivity) together with increased fibre density enhance ephaptic coupling and facilitate near-synchronous spike volleys (Schmidt and R. Knösche, 2022), while earlier EMI simulations show that the ephaptic current scales approximately as O(1/σe_) and thus becomes negligible for very large extracellular conductivities (Tveito et al., 2017b).
In Figure 10 we investigate ephaptic interactions between two neocortical layer 5 pyramidal neurons separated by 1 μm of extracellular space at the AIS. Here, Cell 2 is given a constant weak somatic stimulation current, while the somatic current of Cell 1 is stronger and varied between experiments. Panel A illustrates how an AP firing in Cell 1 perturbs the EP in the narrow cleft between the cells, potentially affecting the dynamics of Cell 2. Indeed, in Panel D, this effect is quantified by reporting the distance between two spikes of Cell 2 as a function of the somatic stimulation of Cell 1. The spike distance is observed to change by several milliseconds depending on the stimulation and resulting firing frequency of Cell 1. Furthermore, the effect increases when σ_e_ is reduced.
Can a neuron excite its neighbor ephaptically?
4.6
As mentioned above, a long-standing question in both cardiac electrophysiology and neuroscience is whether a cell can directly excite its neighbor through the extracellular space alone. We have seen above that one cell can influence another, but can this interaction trigger a full AP? A classical reference is Weingart and Maurer (1988), who examined impulse transfer between isolated myocyte pairs. When two single myocytes were gently pushed together in a side-to-side configuration, they observed no electrotonic interaction and no impulse transfer, and concluded that ephaptic transmission was unlikely. This experiment was revisited in Waaben et al. (2024), but with the cells aligned end-to-end rather than side-by-side. In this geometry, the high-density sodium channel regions at the intercalated disc of the myocytes are brought into proximity, and AP transmission was observed even when gap junctional coupling was absent. Thus, whether signal transfer occurs depends strongly on the cellular configuration.
In Figure 11 we explore this question for our two neuron types placed one μm apart. For the EP generated by one cell to be strong enough to elicit a full AP in its neighbor, the extracellular conductivity σ_e_ must be sufficiently small. This is consistent with the ephaptic current scaling as O(1/σ_e). Whether such small σe_ values occur locally in neural tissue remains uncertain, although geometric confinement and narrow extracellular clefts could, in principle, create domains with reduced effective conductivity. However, in Figure 11, we find that for the EP generated by one cell to directly trigger an AP in a neighboring neuron separated by one μm, the extracellular conductivity, σ_e_, must be reduced by roughly a factor of twenty to fifty relative to the standard value of 3 mS/cm. We repeated the simulations with the distance between the neurons reduced by one half, but this did not change the conclusion regarding the requirements for direct excitation. This places the direct triggering regime outside the range of physiological conductivities. In healthy neural tissue, bulk extracellular conductivity is typically in the range of 3–6 mS/cm (Logothetis et al., 2007).
A notable exception to these bulk values is the Pinceau structure surrounding the Purkinje cell AIS, which forms a locally high-resistivity compartment and supports strong ephaptic inhibition (Blot and Barbour, 2014). However, this structure is anatomically and functionally specialized, and its ephaptic effect is inhibitory rather than excitatory. Thus, while Figure 11 shows that direct ephaptic triggering is theoretically possible in the extreme limit of conductivity, it is unlikely to operate as a mechanism for excitatory propagation in typical cortical or cerebellar tissue. Instead, ephaptic effects are more plausibly involved in modulating spike timing and supporting synchronization, as observed at physiological conductivities in Figures 9, 10.
Numerical implementation
4.7
Several numerical implementations of the EMI system (1) have been proposed. In Tveito et al. (2017b), a straightforward finite difference scheme was used. The complete domain was represented by a finite difference grid in which intra- and extracellular nodes overlap at the membrane, allowing all unknowns to be fully coupled. A finite element approach was presented in Tveito et al. (2017a) and re-used in Buccino et al. (2019). Furthermore, a boundary element approach was used in de Souza et al. (2024), and a cut finite element method was used in Berre et al. (2024).
The implementation used in the present project is described in detail in Jæger et al. (2021b). The underlying idea is to use operator splitting to formulate the problem in terms of standard components in scientific computing: Poisson/Laplace problems in the intracellular and extracellular domains, coupled to ordinary differential equations at the membrane nodes. This decomposition enables straightforward use of well-tested solvers from standard numerical libraries. In this work we use the MFEM finite element library, but any standard finite element library will work well, since the spatial subproblems are standard Poisson/Laplace equations; the key point is to reduce the coupled EMI system to these standard components, as described in Jæger et al. (2021b). The applied numerical code is available on Zenodo (Jæger, 2026).
Experimental validation
4.8
For both cardiomyocytes, see, e.g., Weingart and Maurer (1988); Waaben et al. (2024); Veeraraghavan et al. (2014); Gourdie (2019), and neurons, see, e.g., Anastassiou et al. (2011); Anastassiou and Koch (2015); Han et al. (2018), there has been a lengthy debate on whether ephaptic coupling is of physiological significance. One reason for this debate is that ephaptic effects are expected to be strongest in strictly confined extracellular spaces, where direct measurements of voltages are technically challenging. Computational models have therefore been commonly applied (Sperelakis and Mann, 1977; Schmidt and R. Knösche, 2022; Jæger et al., 2023), but these studies have often lacked accompanying experimental validation.
For the present study, partial validation of the overall EMI framework could be obtained by comparing stimulation–response curves to experiment. For example, the spike counts presented in Figure 5 for both the Purkinje neuron and the neocortical layer 5 pyramidal neuron are strongly modulated by the distance to the external current source and by the stimulation strength. Such dependencies are directly measurable in vitro (or ex vivo) using controlled extracellular stimulation, and could be compared to model predictions in terms of spike count, spike timing shifts, and threshold currents as functions of stimulation amplitude and distance.
Limitations
4.9
Although the present computational scheme based on the EMI model allows for a more detailed representation of neurons than commonly used cable-based models, there are several important limitations. First, the 3D neuron geometries were generated in Gmsh as idealized morphologies and were not constrained by anatomical reconstructions. Second, the membrane models contain a long list of parameters that are guided by experimental measurements, but these parameters are associated with substantial uncertainty and non-uniqueness, in the sense that quite different parameter sets may reproduce similar activity patterns (Marder and Taylor, 2011; Prinz et al., 2004). Third, we have assumed that ionic concentrations are constant in space and time within both the intracellular and extracellular domains. These concentrations could have been allowed to vary by, e.g., applying the KNP-EMI model, which combines the EMI framework with an electroneutral electrodiffusion description and allows ion concentrations and reversal potentials to evolve dynamically (Ellingsrud et al., 2021, 2020). Moreover, at nanometer scales near membranes and in narrow extracellular clefts, the homogenized description used in KNP-EMI breaks down, and electrodiffusive effects, Debye layers and steep local concentration gradients become important. Such effects can be captured only by nanoscale Poisson-Nernst-Planck models, see, e.g., Pods (2017); Pods et al. (2013); Jæger and Tveito (2025a), and are not included in the EMI model formulation applied here.
Conclusion
5
In this study we used a fully coupled EMI formulation to examine how extracellular stimulation and local field effects influence cerebellar Purkinje neurons and neocortical layer 5 pyramidal neurons. The simulations reproduce the expected 1/r decay of potentials from a current source, show that weak fields induce subthreshold oscillations that follow the stimulus frequency, and quantify how constant and sinusoidal stimulation modulate spike rates and timing. For Purkinje neurons, ephaptic interactions synchronize neighboring cells in a distance- and conductivity-dependent manner. In addition, ephaptic interactions between layer 5 pyramidal neurons cause spike time alterations. Together, these results clarify how single-cell biophysics shapes field-mediated interactions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agudelo-Toro A. Neef A. (2013). Computationally efficient simulation of electrical activity at cell membranes interacting with self-generated and externally imposed electric fields. J. Neural Eng. 10:026019. doi: 10.1088/1741-2560/10/2/02601923503026 · doi ↗ · pubmed ↗
- 2Anastassiou C. A. Koch C. (2015). Ephaptic coupling to endogenous electric field activity: why bother? Curr. Opin. Neurobiol. 31, 95–103. doi: 10.1016/j.conb.2014.09.00225265066 · doi ↗ · pubmed ↗
- 3Anastassiou C. A. Perin R. Markram H. Koch C. (2011). Ephaptic coupling of cortical neurons. Nat. Neurosci. 14, 217–223. doi: 10.1038/nn.272721240273 · doi ↗ · pubmed ↗
- 4Anderson R. Andrej J. Barker A. Bramwell J. Camier J.-S. Cerveny J. . (2021). MFEM: a modular finite element methods library. Comput. Math. Applic. 81, 42–74. doi: 10.1016/j.camwa.2020.06.009 · doi ↗
- 5Bakkum D. J. Obien M. E. J. Radivojevic M. Jäckel D. Frey U. Takahashi H. . (2019). The axon initial segment is the dominant contributor to the neuron's extracellular electrical potential landscape. Adv. Biosyst. 3:1800308. doi: 10.1002/adbi.20180030830882024 PMC 6420080 · doi ↗ · pubmed ↗
- 6Ballesteros C. Cheviakov A. Spiteri R. J. (2025). Analytical solutions for the cardiac extracellular-membrane-intracellular model. Math. Methods Appl. Sci. doi: 10.1002/mma.70409 · doi ↗
- 7Berre N. Rognes M. E. Massing A. (2024). Cut finite element discretizations of cell-by-cell EMI electrophysiology models. SIAM J. Sci. Comput. 46, B 527–B 553. doi: 10.1137/23M 1580632 · doi ↗
- 8Blot A. Barbour B. (2014). Ultra-rapid axon-axon ephaptic inhibition of cerebellar Purkinje cells by the pinceau. Nat. Neurosci. 17, 289–295. doi: 10.1038/nn.362424413696 · doi ↗ · pubmed ↗