ALDH1L2 orchestrates redox–growth coupling in renal carcinoma: pan-cancer evidence and mechanistic validation of the ROS–Akt/mTOR/S6K axis
Chao Jiang, Songsong Liu, Liwen Zhang, Shiji Li, Jinyou Wang, Yi Wang

TL;DR
This study explores the role of ALDH1L2 in different cancers, showing it affects tumor growth and survival, especially in kidney cancer.
Contribution
The first systematic analysis of ALDH1L2's divergent roles in kidney, bladder, and prostate cancers with pan-cancer and experimental validation.
Findings
ALDH1L2 expression is linked to survival outcomes in kidney renal clear cell carcinoma.
ALDH1L2 knockdown suppresses KIRC cell proliferation and migration while increasing ROS levels.
ALDH1L2 shows tissue-specific effects, promoting growth in bladder and prostate cancers but inhibiting it in kidney cancer.
Abstract
Aldehyde dehydrogenase family 1 member L2 (ALDH1L2) has been relatively understudied in cancer. We aimed to systematically characterize its expression patterns, clinical significance, and potential functions across cancers and to validate its biological roles in urologic tumors. Leveraging The Cancer Genome Atlas pan-cancer resource, we profiled ALDH1L2 across tumor types with respect to expression patterns, clinical outcomes, genomic features, immune contexture, epigenetic associations, and indices of stemness and heterogeneity. Protein-level differences were examined by immunohistochemistry in bladder cancer (BLCA), prostate adenocarcinoma (PRAD), and kidney renal clear cell carcinoma (KIRC) tissues. To functionally interrogate ALDH1L2, we performed siRNA-mediated knockdown in relevant cell models and evaluated proliferation and motility-related phenotypes using wound-healing,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
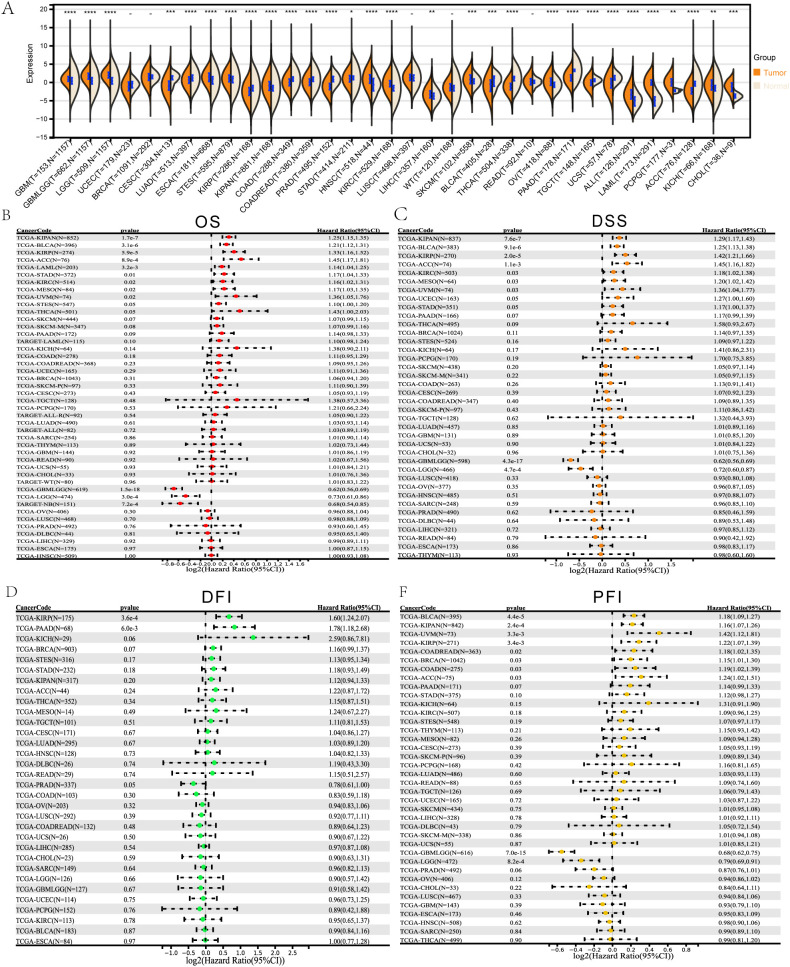 Figure 1
Figure 1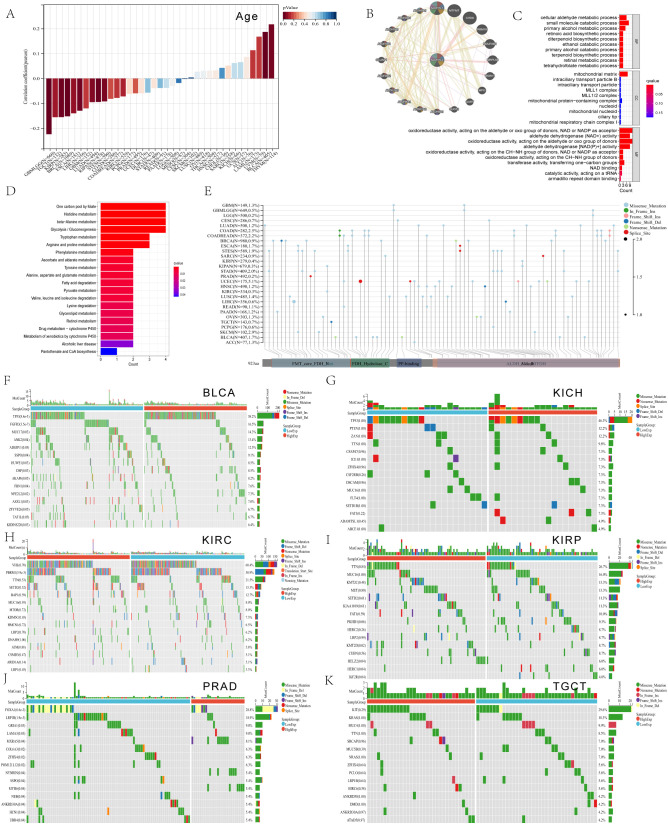 Figure 2
Figure 2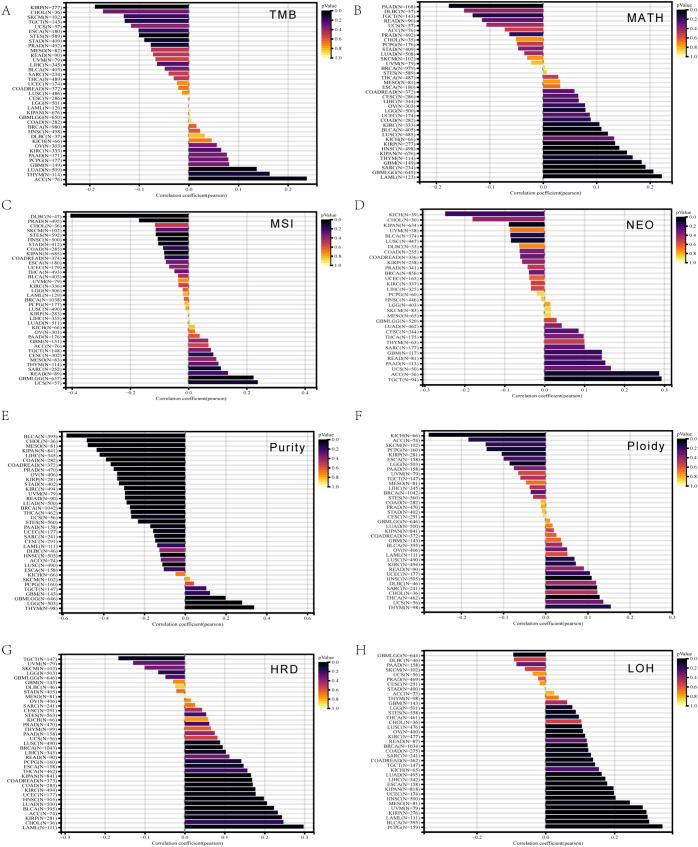 Figure 3
Figure 3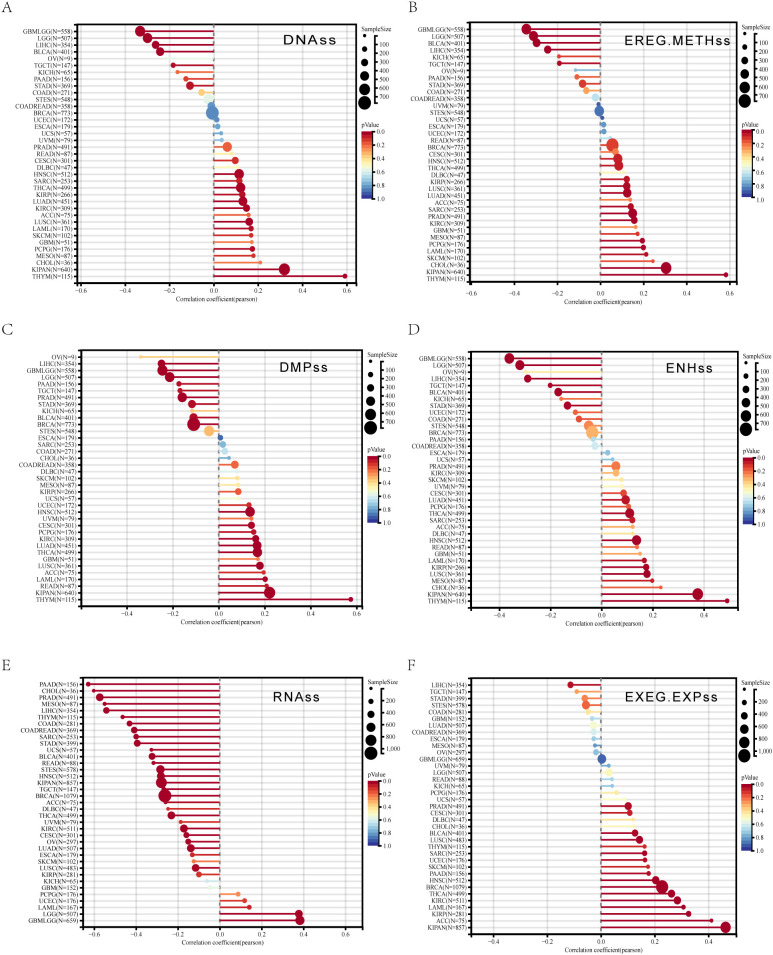 Figure 4
Figure 4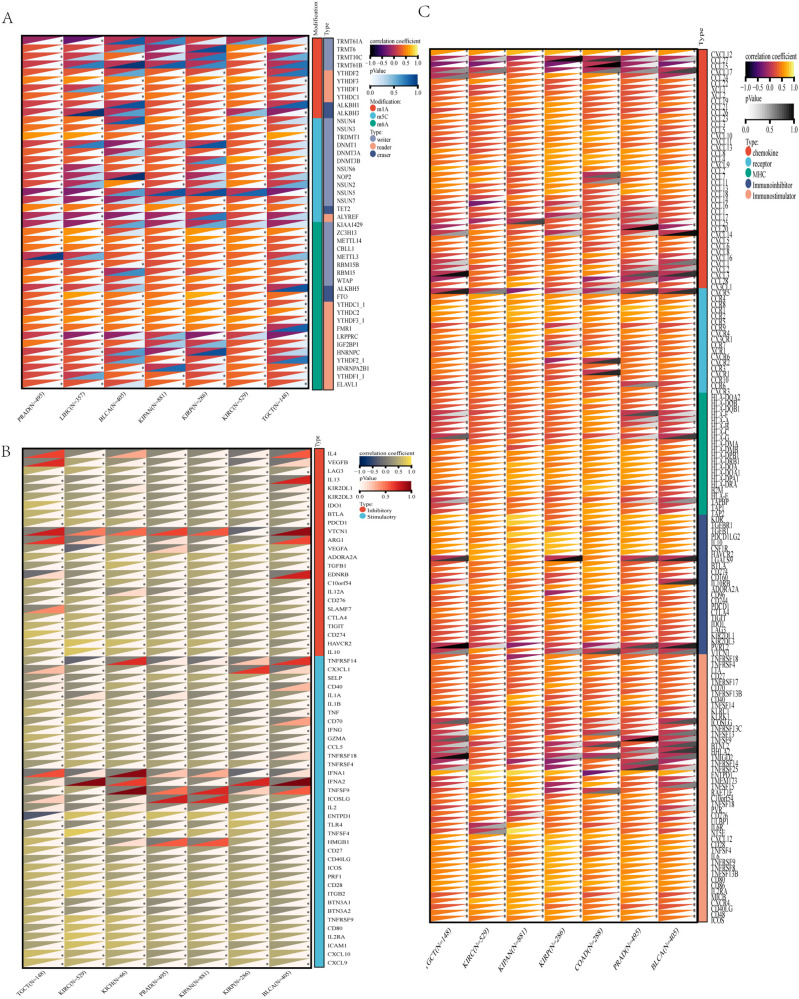 Figure 5
Figure 5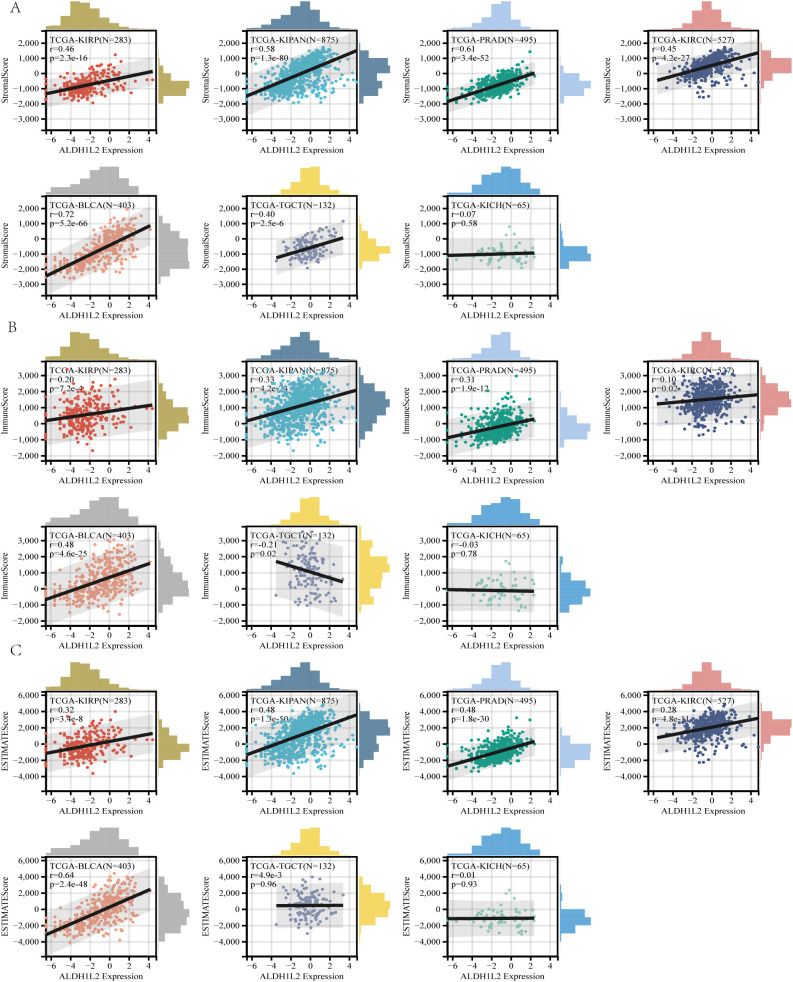 Figure 6
Figure 6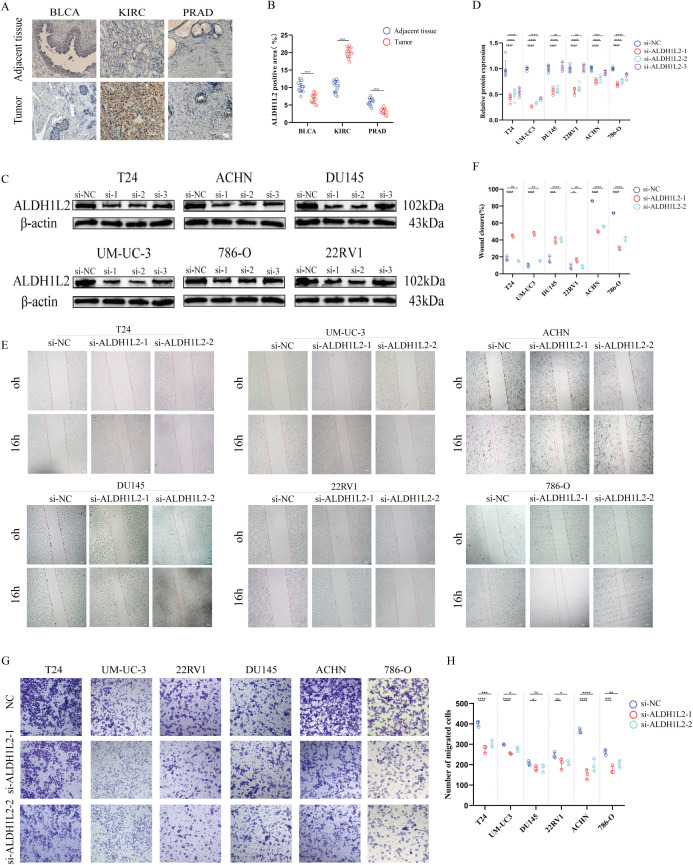 Figure 7
Figure 7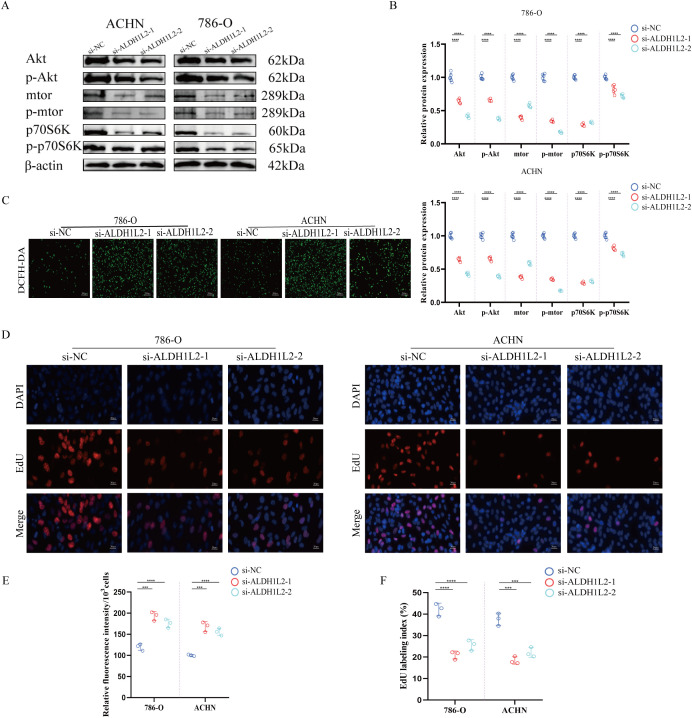 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer Cells and Metastasis · Cancer, Hypoxia, and Metabolism · Ferroptosis and cancer prognosis
Introduction
ALDH1L2 is a mitochondrial folate-dependent enzyme encoded at chromosome 12q23.3 (1, 2). First identified in 2010, ALDH1L2 participates in the folate cycle by catalyzing the conversion of 10-formyltetrahydrofolate to CO2 and tetrahydrofolate, generating mitochondrial NADPH in the process, which is crucial for maintaining cellular redox balance and antioxidant defense (3). Beyond folate metabolism, emerging evidence suggests that ALDH1L2 may influence lipid metabolism and energy homeostasis (4). For instance, ALDH1L2 knock-out mice exhibit lipid droplet accumulation in mitochondria and reduced ATP levels, potentially linked to impaired coenzyme A biosynthesis (4). These findings indicate that ALDH1L2 is not merely a metabolic enzyme but may also function as a regulatory factor in cellular homeostasis.
The involvement of ALDH1L2 in tumor biology has only recently begun to be appreciated (5–7). Previous studies reported that knocking down ALDH1L2 could inhibit distant metastasis of melanoma cells by suppressing the folate pathway (5). In clinical cohorts, elevated ALDH1L2 expression correlated with poorer overall survival in patients with colorectal cancer and lung adenocarcinoma (1). Aging-related metabolic remodeling and redox imbalance are increasingly recognized as shared drivers of chronic diseases, including cancer, providing a rationale to interrogate mitochondrial metabolic enzymes in tumor progression (8). Given the close links between one-carbon metabolism and aging as well as circadian rhythms—both known risk factors for various malignancies (9–12)—ALDH1L2 might represent a key metabolic regulator connecting these biological processes to tumorigenesis.
The role of ALDH1L2 in pan-cancer remains poorly defined, which is attributed to its inconsistent functions across different cancer types. High ALDH1L2 expression correlates with poor prognosis in pancreatic ductal adenocarcinoma (13). This enzyme is also upregulated in human colorectal tumor tissues relative to normal tissues (14), and patients with low ALDH1L2 expression exhibit radioresistance (1). Knockdown of ALDH1L2 has been demonstrated to suppress distant metastasis in human melanoma cells (1). Conversely, ALDH1L2 loss can promote metastatic progression in breast cancer cells by increasing the production of formate and formylmethionine (fMet) (7). Additionally, it has been reported that ALDH1L2 promotes hepatocellular carcinoma (HCC) progression through tumor-associated macrophage polarization, and ALDH1L2 knockdown enhances the anti-HCC efficacy of sorafenib (15). However, a systematic pan-cancer analysis of ALDH1L2 is currently lacking, and its context-dependent roles across different tumor types are poorly understood. Furthermore, most existing studies rely on bioinformatics analyses, lacking direct experimental validation of ALDH1L2 function in cancer cells. Recent global analyses show that the rising burden of urinary tract tumor disease and increasing cross-border inequalities highlight the urgent need for clinically viable biomarkers and targetable pathways for urological cancers (16).
To address these gaps, we performed an integrated analysis of ALDH1L2 across multiple human cancers using TCGA dataset. We focused on KIRC, BLCA, and PRAD, where ALDH1L2 exhibited differential expression and prognostic significance. In addition to informatics analysis, we conducted functional experiments in cancer cell lines—including wound healing, Transwell assays, Western blotting, EdU assay, and ROS detection—to experimentally validate the role of ALDH1L2. Our findings provide new insights into the tumor type-specific functions of ALDH1L2, particularly revealing its regulatory role in the Akt/mTOR signaling pathway and ROS generation in KIRC, thereby highlighting its potential as a prognostic biomarker and therapeutic target.
Methods
Identification and prognostic analysis
Transcriptome profiles and matched clinical annotations for TCGA pan-cancer cohorts were obtained from the UCSC Xena browser (17–19), and ALDH1L2 expression values were extracted for each sample. Samples from primary solid tumors, normal solid tissues, primary blood-derived cancers (TCGA-LAML), and metastatic lesions (TCGA-SKCM) were included at the data acquisition stage. To improve data quality, cases with ALDH1L2 expression equal to zero or follow-up shorter than 30 days were removed before analysis. Expression values were then converted to log_2_(x + 0.001). Cancer types with fewer than ten eligible samples were removed, yielding 38 tumor entities with available ALDH1L2 expression and survival information. Associations between ALDH1L2 expression and survival endpoints were examined using Cox proportional hazards models, and statistical significance was assessed with log-rank tests. For tumor–normal comparisons, we considered primary tumor samples together with normal solid tissues and, where available, blood-derived normal samples. Cancer types with fewer than three normal samples, or with zero expression in all normal tissues, were excluded, resulting in 18 tumor types with usable tumor–normal information. Differences in ALDH1L2 expression between tumor and normal tissues were evaluated using the Wilcoxon rank-sum test or the Wilcoxon signed-rank test as appropriate. Official TCGA abbreviations for each cohort included in this study are summarized in Additional File 1: Table S1.
Tumor stemness, heterogeneity, and mutational landscape
We next explored whether ALDH1L2 is linked to stem-like features in tumors. Specifically, Spearman’s rank correlation was used to relate ALDH1L2 mRNA abundance to multiple established stemness indices spanning methylation and transcriptome-derived scores (DMPss, DNAss, ENHss, EREG.EXPss, EREG-METHss, and RNAss) (20). In parallel, a panel of heterogeneity and genome-instability measures—HRD (homologous recombination deficiency), LOH (loss of heterozygosity), NEO, tumor ploidy, tumor purity, MATH (mutant-allele tumor heterogeneity), MSI (microsatellite instability), and TMB (tumor mutational burden)—was assembled, where TMB was computed from MuTect2 mutation calls and processed with “maftools” (21, 22). After integrating mutation and expression matrices, samples carrying only synonymous alterations were excluded. Within each tumor entity, cases were dichotomized by the median ALDH1L2 expression, and group-wise differences in mutation prevalence were evaluated using a Chi-square framework.
RNA modification-related genes and tumor immune microenvironment
To explore potential links between ALDH1L2 and epitranscriptomic regulation, we assessed Spearman correlations between ALDH1L2 and 44 genes involved in RNA modifications, including writers, readers, and erasers for m1A, m5C, and m6A. Correlations between ALDH1L2 mRNA expression and 36 inhibitory checkpoints (23), 22 stimulatory checkpoints (24), and 68 immunomodulator genes (chemokines, receptors, MHC molecules, immunoinhibitors, immunostimulators) (25) were also investigated. The TIMER and ESTIMATE algorithms were used via the R package “IOBR” (26) to evaluate the TME. Relationships between ALDH1L2 and DNA methylation of its own locus, its mRNA expression, and tumor-infiltrating lymphocytes (TILs) (27) were further explored using the TISIDB database.
Single-cell validation using TISCH2
To further validate the cellular distribution of ALDH1L2 within the tumor microenvironment, we interrogated publicly available single-cell RNA-seq datasets using the Tumor Immune Single Cell Hub 2 (TISCH2) database. The “Gene” module was used to visualize ALDH1L2 expression across annotated cell populations in urological tumor–related datasets, including BLCA, PRAD, and kidney cancer cohorts. Average expression levels were summarized by cell type as provided by the TISCH2 standardized annotation pipeline and presented as log(TPM/10 + 1).
Tumor purity–adjusted immune correlation analysis using TIMER3
To examine whether the association between ALDH1L2 and immune infiltration was confounded by tumor purity, we performed purity-adjusted correlation analyses using TIMER3. The “Immune–Gene” module was applied to evaluate the association between macrophage subset expression and estimated infiltration levels and CD8^+^ T cell signatures in the TCGA cohort ALDH1L2 including BLCA, KICH, KIRC, KIRP, PRD, and TGCT. Immunoinfiltration was inferred by multiple deconvolution methods such as TIMER, EPIC, xCell, CIBERSORT/CIBERSORT-ABS, quanTIseq, MCP-counter, and Consensus-TME, and partial Spearman correlations were reported and tumor purity adjusted. A two-sided P < 0.05 was considered statistically significant.
Immunohistochemistry and scoring
This study utilized tissue microarrays (containing bladder cancer, prostate adenocarcinoma, clear cell renal cell carcinoma tumor tissues, and corresponding adjacent tissues; purchased from Shanghai Zhuoli Biotech Company), which had passed ethical review. After routine dewaxing and rehydration of 4 μm FFPE sections, antigen retrieval was performed using heat-mediated method with citrate buffer (pH 6.0). Endogenous peroxidase activity was blocked with 3% H_2_O_2_. The sections were incubated with the primary antibody anti-ALDH1L2 (Proteintech, Rabbit, 21391-1-AP, 1:1500) at 4 °C overnight or at 37 °C for 60 mins. After washing, an HRP-conjugated secondary antibody was applied, DAB was used as the chromogen, and nuclei were counterstained with hematoxylin before dehydration and mounting. The HRP-conjugated secondary antibody and DAB chromogen were supplied in the immunohistochemistry kit (zsbio, PV-6000). The primary outcome measure was the percentage of ALDH1L2-positive area (% positive area), calculated as (DAB-positive pixel area/ROI tissue area) × 100%.
Cell culture and transfection
Human cell models of KIRC, BLCA, and PRAD were maintained in DMEM containing 10% fetal bovine serum in a humidified incubator (37 °C, 5% CO_2_). ALDH1L2 was knocked down using targeted siRNA. Transfection efficiency was verified by Western blot (WB).
Wound healing assay
Cells were seeded in 6-well plates and cultured to ~90% confluence. A straight scratch was generated using a sterile 200-µL pipette tip held perpendicular to the plate surface. Detached cells were removed by gently washing twice with PBS, followed by incubation in fresh medium (with reduced serum when indicated). Images were acquired at 0 h and 24 h under identical microscope settings, and the same wound area was recorded by referencing pre-marked positions on the plate underside. Wound closure was quantified using ImageJ by measuring the wound area at each time point. The migration rate was calculated as: Wound closure (%) = (A0 − At)/A0 × 100, where A0 and At represent wound area at 0 h and time t, respectively.
Transwell assay
Cell migration and invasion abilities were detected. Cells were fixed, stained, and counted.
Western blotting
Cell lysates were prepared in RIPA buffer supplemented with protease and phosphatase inhibitors. Protein concentrations were quantified using a BCA assay, and 20–40 μg of total protein per sample was resolved by SDS–PAGE before transfer onto PVDF membranes. Membranes were blocked in 5% BSA (for phospho-proteins) or 5% non-fat milk and then incubated at 4 °C overnight with primary antibodies against ALDH1L2 (Proteintech, 21391-1-AP, 1:1500), Akt (CST, 9272, 1:1000), p-Akt(Ser473)(CST, 9271, 1:1000), mTOR (CST, 2972, 1:1000), p-mTOR (Ser2448)(CST, 2971, 1:1000), p70S6K(Thr389) (Affinity, AF6226, 1:1000), p-p70S6K (Affinity, AF3228, 1:1000) and β-actin (Affinity, AF7018, 1:10000). After washing, membranes were incubated with HRP-linked secondary antibodies for 1 h at room temperature, and signals were developed using an ECL substrate. Band intensities were quantified in ImageJ; phosphorylated proteins were normalized to their corresponding total proteins and to the loading control. All assays were performed in ≥3 independent biological replicates.
ROS detection
Intracellular ROS were assessed using DCFH-DA (Beyotime, S0033). After transfection, cells were incubated with 10 μM DCFH-DA in serum-free medium at 37 °C for 20 min in the dark, followed by three washes with PBS to remove excess probe. Fluorescence images were captured using the same exposure settings. ROS levels were quantified by ImageJ as mean fluorescence intensity and normalized to cell number, reported as relative fluorescence intensity (RFI) per 10³ cells. At least five random fields were analyzed per condition, and experiments were repeated in three independent biological replicates.
EdU assay
DNA synthesis and proliferative activity were assessed using an EdU incorporation kit (Beyotime, ST067).Cells were exposed to EdU for a defined labeling period, fixed, permeabilized, and subjected to a click-chemistry reaction to fluorescently label incorporated EdU, while nuclei were counterstained with DAPI. Images were acquired using fluorescence microscopy. The EdU labeling index was quantified using ImageJ as the percentage of EdU-positive nuclei among total DAPI-stained nuclei:EdU labeling index (%) = (EdU^+^nuclei/total nuclei) × 100. At least five random fields were quantified per condition, and experiments were repeated in three independent biological replicates.
Statistical analysis
All statistical analyses were performed using R software (version 3.6.4) and appropriate R packages.
Non-parametric tests, including the unpaired Wilcoxon rank-sum test and the Wilcoxon signed-rank test, were used for two-group comparisons, while the Kruskal–Wallis test was applied for multiple-group comparisons. Unless otherwise specified, all p values were two-sided, and p < 0.05 was considered statistically significant. Significance levels are indicated as follows: *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
Results
Differential expression and prognostic analysis
Compared with matched normal tissues, ALDH1L2 mRNA levels were significantly dysregulated in multiple TCGA cancer types, with clear upregulation in a subset of tumors and downregulation in others (Figure 1A). Among urologic malignancies, ALDH1L2 expression was markedly higher in KIRC than in adjacent kidney tissue, whereas BLCA and PRAD showed reduced expression relative to their corresponding normal controls.
Identification of ALDH1L2 expression and prognostic analysis. (A) Pan-cancer analysis of ALDH1L2 for differential expression between tumor and normal tissues. (B) Pan-cancer analysis of ALDH1L2 for Overall Survival (OS). (C) Pan-cancer analysis of ALDH1L2 for Disease-Specific Survival (DSS). (D) Pan-cancer analysis of ALDH1L2 for Disease-Free Interval (DFI). (E) Pan-cancer analysis of ALDH1L2 for Progression-Free Interval (PFI). Overall Survival, OS; Disease-Specific Survival, DSS; Progression-Free Interval, PFI; Disease-Free Interval, DFI.
Survival analyses across the pan-cancer cohort demonstrated that elevated ALDH1L2 expression was associated with unfavorable overall survival (OS) in several entities, including KIPAN, BLCA, KIRP, adrenocortical carcinoma (ACC), KIRC, acute myeloid leukemia (LAML), and stomach adenocarcinoma (STAD), whereas low ALDH1L2 expression predicted poor OS in some brain tumors (Figure 1B). Consistent patterns were observed for disease-specific survival (DSS), where high ALDH1L2 expression indicated worse DSS in KIPAN, BLCA, KIRP, ACC, KIRC, LAML, and STAD, while reduced expression correlated with poorer DSS in glioma-related cohorts (Figure 1C). For DFI and PFI, ALDH1L2 also showed tumor-type-dependent prognostic value: high expression was linked to shorter DFI in pancreatic adenocarcinoma (PAAD) and KIRP and to inferior PFI in BLCA, KIPAN, ACC, uveal melanoma (UVM), breast cancer (BRCA), colon adenocarcinoma (COAD), colorectal cancer (COADREAD), and KIRP, whereas low ALDH1L2 expression was associated with unfavorable PFI in certain glioma subgroups (Figures 1D, E).
Clinically, ALDH1L2 levels were significantly correlated with sex, TNM stage, pathological grade, and overall clinical stage in multiple tumor types, including urologic cancers (Supplementary Figures S1B–G). For example, ALDH1L2 expression was positively related to T stage and higher pathological grade in KIRC and KIPAN, while age-stratified analyses showed a positive correlation between ALDH1L2 and age in BLCA but negative correlations in KIPAN, KIRC, and PRAD, suggesting that ALDH1L2 expression patterns may be influenced by both tumor biology and host factors(Supplementary Figures S1B–G).
The cancers consistently identified through both differential expression and prognostic analyses were KIPAN and BLCA for DSS and PFI. ALDH1L2 expression also varied significantly across age groups (Figure 2A), correlating positively in BLCA and negatively in KIPAN, KIRC, and PRAD.
Analysis of ALDH1L2 in relation to age, biological function, and mutational landscape. (A) Pan-cancer analysis of ALDH1L2 expression by age. (B) Correlation analysis between ALDH1L2 and the ALDH family as well as common pathogenic gene mutations. (C) Gene Ontology (GO) terms enriched among genes associated with ALDH1L2. (D) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways significantly enriched for ALDH1L2-related genes. (E–H) Somatic mutation profiles after stratifying patients by the median ALDH1L2 expression: the 15 most frequently mutated genes are shown for (E) BLCA, (F) KIRC, (G) KIRP, and (H) PRAD. Key mutated genes include TP53, FGFR3, and MUC17 in BLCA; PBRM1 and KDM5C in KIRC; KMT2D and IGF2R in KIRP; and FOXA1 and LRP1B in PRAD.
Tumor heterogeneity, stemness, mutational landscape, RNA modifications, and immune checkpoint genes
Regarding tumor heterogeneity, in PRAD, the mRNA expression of ALDH1L2 was negatively correlated with MSI (R = -0.17) and tumor purity (R = -0.35) (Figures 3A–H). In BLCA, ALDH1L2 mRNA expression showed positive correlations with HRD (R = 0.22) and LOH (R = 0.30), but was negatively correlated with tumor purity (R = -0.58) (Figures 3A–H). For TGCT, ALDH1L2 mRNA levels showed an inverse association with HRD (R = −0.17). Within the pan-kidney cohort (KIPAN), higher ALDH1L2 expression aligned with increased MATH (R = 0.15), HRD (R = 0.17), and LOH (R = 0.20), while it tracked with lower tumor purity (R = −0.43), reduced neoantigen load (NEO; R = −0.09), and decreased MSI (R = −0.08).
Pan-cancer spearman analysis of tumor heterogeneity and ALDH1L2 expression. (A) Correlation between TMB and ALDH1L2 levels. (B) Correlation between MATH and ALDH1L2 levels. (C) Correlation between MSI and ALDH1L2 levels. (D) Correlation between NEO load and ALDH1L2 levels. (E) Correlation between Tumor Purity and ALDH1L2 levels. (F) Correlation between Ploidy and ALDH1L2 levels. (G) Correlation between HRD and ALDH1L2 levels. (H) Correlation between LOH and ALDH1L2 levels. TMB, tumor mutational burden; MATH, mutant-allele tumor heterogeneity; MSI, microsatellite instability; NEO, neoantigen; HRD, homologous recombination deficiency; LOH, loss of heterozygosity.
Spearman analyses further linked ALDH1L2 to tumor stemness in a cancer-type-specific manner
In KIRC and KIPAN, ALDH1L2 was positively associated with DNAss, EREG-METHss, DMPss, and EREG.EXPss, but showed an opposite trend for RNAss (Figures 4A–F).
Pan-cancer spearman analysis of tumor stemness and ALDH1L2 expression. (A) Correlation between DNAss and ALDH1L2 levels. (B) Correlation between EREG-METHss and ALDH1L2 levels. (C) Correlation between DMPss and ALDH1L2 levels. (D) Correlation between ENHss and ALDH1L2 levels. (E) Correlation RNAss and ALDH1L2 levels. (F) Correlation between EREG-METHss and ALDH1L2 levels.
In BLCA, ALDH1L2 displayed concordant positive relationships with DNAss, EREG-METHss, DMPss, and ENHss (Figures 4A–F). By contrast, PRAD exhibited a mixed pattern: ALDH1L2 was negatively related to RNAss and EREG.EXPss, yet remained positively associated with EREG-METHss (Figures 4A–F).
Age, biological function, and genetic mutation landscape
The correlation of ALDH1L2 with the ALDH family and commonly mutated genes is presented in Figure 2B. Functional annotation of ALDH1L2-associated genes was then performed using GO and KEGG analyses. GO terms highlighted enrichment in biological processes (BP) and molecular functions (MF), whereas KEGG results mainly pointed to pathways involved in one-carbon unit metabolism in folate-related reactions and amino-acid metabolic programs (Figures 2C, D).
The somatic alteration landscape of ALDH1L2 is presented in Figure 2E. For downstream comparisons, patients were stratified into ALDH1L2-high and ALDH1L2-low groups according to the median expression within each tumor type. In BLCA, mutation frequencies differed between expression strata for TP53, FGFR3, MUC17, ANK2, SSPO, HUWE1, DSP, FBN1, NFE2L2, ASXL1, and KIDINS220 (Figure 2F). In KIRC, group-wise differences were most notable for PBRM1, KDM5C, and LRP1 (Figure 2H). In KIRP, the ALDH1L2-defined strata showed distinct mutation patterns involving KMT2D, HELZ2, HERC1, and IGF2R (Figure 2I). In PRAD, differential mutation prevalence was observed for FOXA1, LRP1B, GRIA1, LAMA3, MXRA5, COL6A3, ZFHX4, POM121L12, NYNRIN, SSPO, MYH6, NEB, ANKRD30A, HCN1, and UBR4 (Figure 2J). By contrast, no clear mutation-frequency differences were detected between ALDH1L2-high and -low groups in KICH (Figure 2G) or TGCT (Figure 2K).
RNA modification-associated genes, immune checkpoints, and immunomodulatory genes
Regarding RNA modifications, we found that the expression level of ALDH1L2 in KICH, PRAD, BLCA, KIPAN, KIRP, KIRC, and TGCT was correlated with writer, reader, and eraser genes involved in m1A, m5C, and m6A RNA modifications. Multiple immune checkpoint genes (Figure 5B) and immunomodulatory genes (Figure 5C) showed associations with ALDH1L2 expression levels across all seven cancer types.
Relationships between ALDH1L2 and RNA modification machinery, immune checkpoints, and immunoregulatory factors. Spearman’s rank correlation was used to evaluate the association between ALDH1L2 expression and: (A) genes involved in RNA modifications, (B) immune checkpoint molecules, and (C) immunomodulatory genes.
Multi-level analyses link ALDH1L2 to the tumor microenvironment, validated by TISCH2 and TIMER3
Across urologic and related tumor types, ALDH1L2 expression displayed distinct microenvironmental associations as quantified by ESTIMATE (Figures 6A–C). In TGCT, ALDH1L2 was aligned with a higher stromal component (stromal score, R = 0.40) but a reduced immune component (immune score, R = −0.21). In KIRP, increasing ALDH1L2 levels tracked with higher stromal, immune, and composite ESTIMATE scores (R = 0.46, 0.20, and 0.32, respectively). Similar concordant patterns were observed in KIPAN (stromal, immune, and ESTIMATE scores: R = 0.58, 0.33, and 0.48) and PRAD (R = 0.61, 0.31, and 0.48). In KIRC, ALDH1L2 showed more modest but still positive relationships with stromal, immune, and ESTIMATE scores (R = 0.45, 0.10, and 0.28), whereas BLCA exhibited comparatively stronger concordance across these metrics (R = 0.72, 0.48, and 0.64).
Association between ALDH1L2 expression and tumor immune microenvironment. (A) Correlation between ALDH1L2 expression and Stromal Score. (B) Correlation between ALDH1L2 expression and Immune Score. (C) Correlation between ALDH1L2 expression and ESTIMATE Score.
Immune deconvolution further indicated that ALDH1L2 expression generally increased with immune cell infiltration (Supplementary Figure S1A). In PRAD, KIRP, KIPAN, and KIRC, higher ALDH1L2 was associated with greater inferred abundance of B cells, CD4^+^ T cells, CD8^+^ T cells, neutrophils, macrophages, and dendritic cells. KICH showed positive associations limited to B cells, CD8^+^ T cells, macrophages, and dendritic cells. In BLCA, ALDH1L2 correlated with CD4^+^ T cells, CD8^+^ T cells, neutrophils, macrophages, and dendritic cells. To strengthen the immune-related findings derived from bulk TCGA analyses, we performed external validation using single-cell and purity-adjusted resources. In the TISCH2 single-cell atlas (Supplementary Figure S2A), ALDH1L2 expression was generally low across major lymphocyte compartments, whereas detectable signals were preferentially observed in stromal/mesenchymal subsets (e.g., fibroblasts and myofibroblasts) and plasma cell populations in several BLCA/PRAD/kidney cancer datasets, supporting a microenvironment-associated expression pattern. In parallel, TIMER3 analyses demonstrated that the correlations between ALDH1L2 expression and immune infiltration estimates persisted after tumor purity adjustment, with macrophage-related signatures showing broadly positive associations across multiple deconvolution algorithms (Supplementary Figure S2B). Notably, the association with CD8^+^ T-cell–related signatures appeared more context-dependent across tumor types, consistent with the heterogeneous immune correlations observed in our pan-cancer TME analyses.
ALDH1L2 knockdown was verified and regulated migration and invasion in bladder, prostate, and renal cancer cells
We found that the expression of ALDH1L2 was slightly lower in BLCA compared to normal bladder tissue; its expression was significantly higher in KIRC than in normal kidney tissue; while in PRAD, ALDH1L2 expression was also slightly lower than in normal prostate tissue (Figures 7A, B). The interference efficiency of siRNA targeting ALDH1L2 was validated by qPCR and WB. Subsequently, si-*ALDH1L2–*1 and si-*ALDH1L2–*2 were selected for further experiments. Wound healing and Transwell assays were performed using two bladder cancer cell lines (T24 and UMUC3), two renal cancer cell lines (ACHN and 786-O), and two prostate cancer cell lines (22RV1 and DU145). The results indicated that si-ALDH1L2 reduced the migratory ability of ACHN and 786-O cells but exhibited a certain enhancing effect on the migration of bladder and prostate cancer cells (Figures 7C–H).
*ALDH1L2 shows distinct expression patterns in urological cancers and differentially regulates cell migration and invasion. (A) Representative immunohistochemistry (IHC) images of ALDH1L2 in bladder urothelial carcinoma (BLCA), kidney renal clear cell carcinoma (KIRC), and prostate adenocarcinoma (PRAD) tissues and matched adjacent normal tissues (n = 15 paired samples per cancer type). (B) Quantification of ALDH1L2 staining in (A) using ALDH1L2-positive protein area (%). (C) Western blot validation of ALDH1L2 knockdown efficiency using three siRNAs (si-1, si-2, si-3) in BLCA (T24, UM-UC-3), KIRC (ACHN, 786-O), and PRAD (DU145, 22RV1) cell lines; β-actin served as the loading control. (D) Densitometric quantification of ALDH1L2 protein levels in (C), normalized to β-actin and expressed relative to the negative control (si-NC). (E) Representative images of wound-healing assays at 0 h and 16 h after scratching in the indicated cell lines transfected with si-NC or two selected siRNAs (si-ALDH1L2–1 and si-ALDH1L2-2). (F) Quantification of wound closure in (E), calculated based on wound area measurements. (G) Representative images of Transwell migration assays following ALDH1L2 knockdown in the indicated cell lines. (H) Quantification of migrated cells in (G), presented as the number of migrated cells per field. (*p<0.05; **p<0.01; ***p<0.001; ***p<0.0001)
ALDH1L2 knockdown enhances ROS generation, suppresses the Akt/mTOR/S6K signaling pathway, and inhibits the proliferation of renal carcinoma cells
Western blot analysis revealed that the activity of the Akt/mTOR/S6K signaling pathway was markedly suppressed following ALDH1L2 knockdown (Figures 8A, B). Since ALDH1L2 showed a good prognostic correlation and a consistent tumor-promoting phenotype in KIRC, we prioritized the kidney cancer model to explore the mechanistic nature of the ROS–Akt/mTOR axis, while ACHN and 786-O were more classic in the cell model of kidney cancer. We performed ROS generation assays in two renal carcinoma cell lines, ACHN and 786-O. The results demonstrated that knockdown of ALDH1L2 significantly promoted ROS production (Figures 8D, F). Furthermore, 5-ethynyl-2’-deoxyuridine (EdU) incorporation assays indicated that ALDH1L2 knockdown effectively inhibited the proliferation of both ACHN and 786-O cells (Figures 8C, E).
*ALDH1L2 knockdown suppresses the Akt/mTOR/S6K signaling pathway, enhances ROS generation, and inhibits renal cancer cell proliferation. (A, B)ALDH1L2 knockdown in renal cancer cells suppresses the Akt/mTOR/S6K signaling pathway and quantification results. (C) Representative fluorescence images of intracellular reactive oxygen species (ROS) detected by DCFH-DA staining in ACHN and 786-O cells transfected with si-NC or si-ALDH1L2. (D) Representative images of EdU incorporation assays in ACHN and 786-O cells following ALDH1L2 knockdown; nuclei were counterstained with DAPI. (E) Quantification of ROS levels corresponding to (C), expressed as relative fluorescence intensity (RFI) per 10³ cells. (F) Quantification of proliferative activity corresponding to (D), presented as the EdU labeling index (%) = EdU+ nuclei/total DAPI nuclei × 100. (***p<0.001; ***p<0.0001)
Discussion
This study represents the first systematic effort to elucidate the expression patterns and biological roles of ALDH1L2 across multiple cancer types by integrating pan-cancer analysis, immunohistochemical validation, and functional experiments. Pan-cancer analysis revealed that ALDH1L2 is differentially expressed in clear cell renal cell carcinoma (KIRC), bladder urothelial carcinoma (BLCA), and prostate adenocarcinoma (PRAD), with its expression levels significantly correlating with patient prognosis. Immunohistochemical staining further confirmed the downregulation of ALDH1L2 expression at the tissue level. Notably, functional experiments demonstrated that ALDH1L2 knock down exerted opposing effects depending on the cancer type: it suppressed cell migration and proliferation in KIRC, whereas it promoted these processes in BLCA and PRAD. These findings indicate that ALDH1L2 can play a context-dependent, dual role in tumorigenesis, which is shaped by tissue origin and metabolic background.
Previous studies have established that ALDH1L2 plays a critical role in mitochondrial one-carbon metabolism, NADPH production, and lipid homeostasis (4, 5, 7, 28). Loss of ALDH1L2 function leads to reduced antioxidant capacity, metabolic reprogramming, and the subsequent accumulation of lipids and ROS (29, 30). In hormone-dependent cancers such as breast and prostate cancer, the excessive utilization of lipids and cholesterol is a key driver of cell proliferation (31–36), which may explain the tumor-promoting effect of ALDH1L2 observed in BLCA and PRAD. Cellular senescence represents a context-dependent program that can restrain proliferation yet also remodel the tumor ecosystem, thereby shaping therapy responsiveness and disease evolution (37). KIRC cells exhibit high dependence on mitochondrial metabolism; therefore, ALDH1L2 deficiency is more likely to disrupt NADPH balance, induce ROS accumulation, and activate the Akt/mTOR pathway, thereby enhancing proliferative capacity. Such cancer type-specific effects underscore the complexity of ALDH1L2 function and untangle its unique position at the intersection of metabolic and signaling networks.
There is now uncertainty in the treatment of the tumor microenvironment, as immune infiltration does not necessarily translate into effective anti-tumor immunity, as bone marrow-driven immunosuppression and CD8+ T cell depletion can decouple immune abundance from immune capacity (38). And advancements in tumor immunotherapy targeting one-carbon metabolism have demonstrated significant progress. This metabolic network influences immunotherapy outcomes via several interconnected pathways, notably the folate cycle, the methionine cycle, and the transsulfuration pathway (39, 40). One-carbon metabolism plays an important role in circadian rhythms, which can influence the tumor microenvironment by affecting immune components, metabolic status, and treatment vulnerability (41). Consequently, key enzymes within one-carbon metabolism—including serine hydroxymethyltransferase (SHMT), methylenetetrahydrofolate dehydrogenase (MTHFD), thymidylate synthase (TYMS), and dihydrofolate reductase (DHFR)—have become a major focus of investigation. Inhibitors of SHMT have been shown to suppress the growth of pancreatic tumor xenografts and demonstrate anticancer activity in vivo (42). They also exert potent antitumor effects through mechanisms such as inducing G1–S cell cycle arrest and inhibiting breast cancer growth (43). Several MTHFD inhibitors have exhibited favorable in vivo antitumor activity following oral administration (44, 45). Meanwhile, TS and DHF reductase inhibitors have already been applied in clinical settings (46–49), with multiple compounds demonstrating substantial antitumor effects in clinical trials (50, 51). The regulated cell death process is closely linked to redox homeostasis and anti-cancer immunity, supporting the translational theory for ROS-related vulnerability combined with existing signal inhibitors (52). Hypoxia is a common metabolic feature of solid tumors, which may weaken the activity of natural killer (NK) cells, thereby promoting immune escape (53). Emerging evidence suggests that microbiota–immune crosstalk may critically shape the urinary tumor immune milieu and influence disease behavior as well as treatment responses, highlighting potential confounders and opportunities for stratification (54). Therefore, targeting ALDH1L2 holds promising therapeutic potential, although further exploration is required for the development of antitumor therapies based on one-carbon metabolism.
Mechanistic studies indicated that ALDH1L2 knock down activates the Akt/mTOR pathway and promotes ROS generation. The Akt/mTOR signaling axis plays a central role in metabolic reprogramming and growth regulation, while moderate levels of ROS can act as signaling molecules to further drive tumor proliferation. EdU assay confirmed that ALDH1L2 deletion enhances cell proliferative capacity. These results suggest the existence of an “ALDH1L2–NADPH–ROS–Akt/mTOR–Proliferation” functional axis, providing a new perspective for understanding the tumor-suppressive role of ALDH1L2 in KIRC. Future studies utilizing Akt/mTOR inhibitors or ROS scavengers may help validate this causal relationship and offer novel strategies for the treatment of renal cell carcinoma.
Clinically, ALDH1L2 expression holds potential as a prognostic biomarker for KIRC patients, demonstrating considerable predictive value when combined with immune infiltration and molecular subtyping analyses. To minimize the impact of tumor purity on our findings, we additionally analyzed these associations using TIMER2.0 alongside purity-adjustment methods, and the results remained consistent. In addition to host-intrinsic determinants, the intratumoral microbiota has been implicated in shaping immunotherapy responsiveness, suggesting that microenvironmental context may modulate biomarker–immune associations across cohorts (55). However, the specific mechanisms underlying its tumor-promoting role in BLCA and PRAD require further investigation, particularly regarding its relationship with cholesterol homeostasis, androgen signaling, and the tumor microenvironment. Against the backdrop of the increasing global burden of urothelial malignancies (16), accumulating evidence indicates intersections between age-related metabolic remodeling—such as hypoxia, myeloid suppression (8), and microbiota immune contextualization (38, 53–55)—and drug sensitivity governed by redox regulation (52). This provides a strong rationale for exploring ALDH1L2 as a context-dependent metabolic−immune hub in urological tumors.
Despite providing multi-layered evidence, this study has several limitations. Our experimental validation was primarily conducted in ex vivo cellular models, lacking support from animal studies or clinical samples. Moreover, the direct causal relationship between ALDH1L2 and immune−cell infiltration remains unclear. Systematic investigations into the expression differences of ALDH1L2 across diverse populations are still lacking. Our detection methods were unable to distinguish between the potential catalytic effects of ALDH1L2 and its non−enzymatic or structural functions. Although TCGA provides large−scale evidence, residual batch effects and cross−cohort clinical heterogeneity may influence the pan−cancer correlations. Future studies should focus on validating the proposed mechanisms in animal models, further dissecting the interactions between ALDH1L2 and lipid metabolism or immune microenvironment factors, and evaluating its clinical feasibility as a diagnostic or therapeutic target.
In summary, this study uncovers the differential expression and clinical significance of ALDH1L2 in various cancers, and confirms its heterogeneous roles in urological tumors through immunohistochemistry and functional experiments. Further mechanistic studies suggest that ALDH1L2 influences KIRC cell proliferation by regulating the Akt/mTOR pathway and ROS dynamics. Collectively, these results position ALDH1L2 as a candidate marker with potential translational relevance and motivate further studies on how mitochondrial one-carbon/redox metabolism interfaces with growth signaling in distinct tumor settings.
Conclusion
In this work, we integrate pan-cancer profiling with tissue and cell-based validation to define ALDH1L2 as a context-dependent regulator linking mitochondrial redox control to growth signaling in urologic malignancies. In KIRC, loss of ALDH1L2 drives ROS accumulation and Akt/mTOR activation, thereby promoting cell proliferation and migration; conversely, it exhibits an opposing effect in BLCA and PRAD. These observations support the utility of ALDH1L2 in molecular stratification and suggest that therapeutic strategies combining mTOR pathway inhibition with redox regulation may be worthy of evaluation in appropriately screened patient subgroups. Given that relevant therapeutic agents have already entered clinical use, future validation of ALDH1L2-based stratification and corresponding targeting strategies across different cancer types holds clear translational promise.
With the increasing burden of urinary tract malignancies worldwide [xx], more and more evidence highlights the convergence of age-related metabolic remodeling [xx], such as hypoxia, myeloid suppression and microbiota immune contextualization [xx–xx], and redox-modulated druggable susceptibility [xx], providing a strong basis for exploring ALDH1L2 as context-dependent metabolic immune junctions in urologic tumors.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yu L Guo Q Luo Z Wang Y Weng J Chen Y . TXN inhibitor impedes radioresistance of colorectal cancer cells with decreased ALDH 1L 2 expression via TXN/NF-κB signaling pathway. Br J Cancer. (2022) 127:637–48. doi: 10.1038/s 41416-022-01835-1, PMID: 35597868 PMC 9381770 · doi ↗ · pubmed ↗
- 2Ma Q Hao S Hong W Tergaonkar V Sethi G Tian Y . Versatile function of NF-ĸB in inflammation and cancer. Exp Hematol Oncol. (2024) 13:68. doi: 10.1186/s 40164-024-00529-z, PMID: 39014491 PMC 11251119 · doi ↗ · pubmed ↗
- 3Krupenko NI Dubard ME Strickland KC Moxley KM Oleinik NV Krupenko SA . ALDH 1L 2 is the mitochondrial homolog of 10-formyltetrahydrofolate dehydrogenase. J Biol Chem. (2010) 285:23056–63. doi: 10.1074/jbc.M 110.128843, PMID: 20498374 PMC 2906299 · doi ↗ · pubmed ↗
- 4Krupenko NI Sharma J Pediaditakis P Helke KL Hall MS Du X . ALDH 1L 2 knockout mouse metabolomics links the loss of the mitochondrial folate enzyme to deregulation of a lipid metabolism observed in rare human disorder. Hum Genomics. (2020) 14:41. doi: 10.1186/s 40246-020-00291-3, PMID: 33168096 PMC 7654619 · doi ↗ · pubmed ↗
- 5Piskounova E Agathocleous M Murphy MM Hu Z Huddlestun SE Zhao Z . Oxidative stress inhibits distant metastasis by human melanoma cells. Nature. (2015) 527:186–91. doi: 10.1038/nature 15726, PMID: 26466563 PMC 4644103 · doi ↗ · pubmed ↗
- 6Chen L Zhang Z Hoshino A Zheng HD Morley M Arany Z . NADPH production by the oxidative pentose-phosphate pathway supports folate metabolism. Nat Metab. (2019) 1:404–15. doi: 10.1038/s 42255-019-0043-x PMC 648912531058257 · doi ↗ · pubmed ↗
- 7Hennequart M Pilley SE Labuschagne CF Coomes J Mervant L Driscoll PC . ALDH 1L 2 regulation of formate, formyl-methionine, and ROS controls cancer cell migration and metastasis. Cell Rep. (2023) 42:112562. doi: 10.1016/j.celrep.2023.112562, PMID: 37245210 · doi ↗ · pubmed ↗
- 8Chmielewski PP Data K Strzelec B Farzaneh M Anbiyaiee A Zaheer U . Human aging and age-related diseases: from underlying mechanisms to pro-longevity interventions. Aging Dis. (2024) 16:1853–77. doi: 10.14336/AD.2024.0280, PMID: 38913049 PMC 12221391 · doi ↗ · pubmed ↗