Occallatibacter bavaricus sp. nov., a new representative of the Acidobacteriota isolated from fen soils, reclassification of Terracidiphilus gabretensis as Occallatibacter gabretensis comb. nov. and emended description of the genus Occallatibacter
Katharina J. Huber, János Papendorf, Carolin Pilke, Petra Büsing, Boyke Bunk, Cathrin Spröer, Sarah Kirstein, Jacqueline Wolf, Meina Neumann-Schaal, Manfred Rohde, Michael Pester

TL;DR
A new soil bacterium, Occallatibacter bavaricus, was discovered in Germany and reclassified another species into the same genus.
Contribution
The paper introduces a new bacterial species and reclassifies an existing one based on genetic and phenotypic data.
Findings
Occallatibacter bavaricus was isolated from fen soil and has unique growth and lipid characteristics.
Terracidiphilus gabretensis was reclassified as Occallatibacter gabretensis based on phylogenetic and chemotaxonomic analysis.
The new species shares 96.1–96.7% 16S rRNA gene sequence similarity with related acidobacterial strains.
Abstract
The acidobacterial strain JP12T affiliated with the Terriglobales has been isolated from fen soil sampled at Schlöppnerbrunnen II near Bayreuth, Germany. The strain was Gram-stain-negative, non-motile, displayed non-spore-forming rods that divide by binary fission and segregate exopolysaccharide-like structures. JP12T grew at temperatures of 4–40 °C (best between 24 and 30 °C), at pH values of 3.7–6.0 (best between 4.1 and 5.6) and at NaCl concentrations of 0–0.5% (best 0.25%, w/v). MK-8 was identified as the major respiratory quinone. The major fatty acids of the strain JP12T were iso-C15:0, C16:0, iso-C17:1 ω7c, iso-C17:0 and iso-diabolic acid. Phosphatidylglycerol, phosphatidylethanolamine and ornithine-containing lipids were the major polar lipids. Lysophosphatidylethanolamine, phosphatidylinositol, diphosphatidylglycerol and high-mass intact polar lipids occurred in small amounts.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
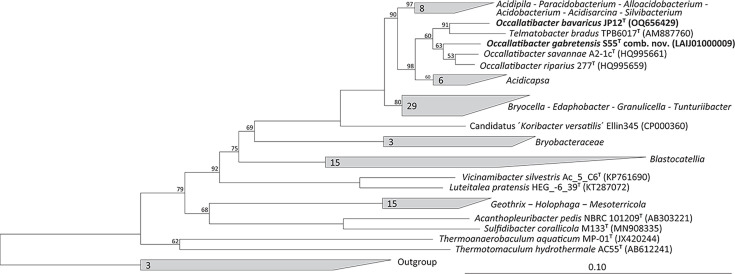 Fig. 1
Fig. 1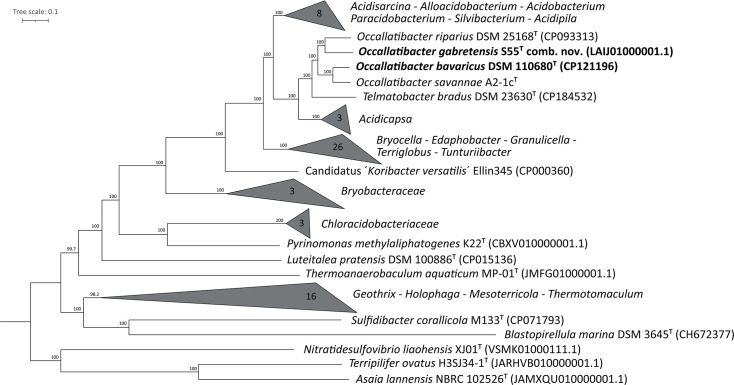 Fig. 2
Fig. 2 Fig. 3
Fig. 3| Characteristic | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
|
| |||||
|
|
|
|
|
|
|
|
|
| 100 | 98.29 | 97.71 | 96.09 |
|
|
| 98.29 | 100 | 97.00 | 95.31 |
|
|
| 97.71 | 97.00 | 100 | 95.27 |
|
|
| 96.09 | 95.31 | 95.27 | 100 |
|
| |||||
|
|
|
|
|
|
|
|
|
| 100/97.45 | 72.71/72.85 | 72.68/72.72 | 71.68/71.85 |
|
|
| 72.71/72.85 | 100 | 72.13 | 71.11 |
|
|
| 72.68/72.72 | 72.13 | 100 | 71.29 |
|
|
| 71.68/71.85 | 71.11 | 71.29 | 100 |
|
| |||||
|
|
|
|
|
|
|
|
|
| 100/93.00 | 13.40/13.40 | 13.30/13.20 | 13.10/13.00 |
|
|
| 13.40/13.40 | 100 | 13.00 | 12.90 |
|
|
| 13.30/13.20 | 13.00 | 100 | 12.90 |
|
|
| 13.10/13.00 | 12.90 | 12.90 | 100 |
|
| |||||
|
|
|
|
|
|
|
|
|
| 100/98.33 | 69.80/69.76 | 69.88/69.82 | 67.54/67.56 |
|
|
| 69.80/69.76 | 100 | 69.03 | 67.77 |
|
|
| 69.88/69.82 | 69.03 | 100 | 67.24 |
|
|
| 67.54/67.56 | 67.77 | 67.24 | 100 |
|
| |||||
|
|
|
|
|
|
|
|
|
| 100/92.47 | 63.99/63.64 | 56.34/56.11 | 45.42/44.25 |
|
|
| 63.99/63.64 | 100 | 54.12 | 45.25 |
|
|
| 56.34/56.11 | 54.12 | 100 | 48.44 |
|
|
| 45.42/44.25 | 45.25 | 48.44 | 100 |
| Fatty acid/strain | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
|
| – | – | – | – | – |
|
| – | – | – | 0.5 | – |
|
| 0.3 | – | 0.2 | 0.1 | – |
|
| 0.9 | – | – | 0.1 | – |
|
| 0.1 | 0.1 | 0.2 | 0.2 | – |
|
|
|
|
|
|
|
|
| 0.3 | 0.6 | 0.2 | 0.2 | 0.2 |
|
| 0.1 | – | – | – | – |
|
| 0.5 | 0.2 | 0.2 | 1.3 | 0.2 |
|
| – | – | – | 0.3 | – |
|
| – | – | – | – | – |
|
| 0.6 | 0.4 | 0.6 | 0.7 | 0.7 |
|
| 2.2 | 1.1 | 1.4 | 1.4 | 3.2 |
|
|
|
|
|
|
|
|
| 0.2 | 0.7 | 0.7 | 0.6 | 0.4 |
|
|
|
|
|
|
|
|
| 0.9 | 0.5 | 1.2 | 0.7 | 0.6 |
|
| 0.1 | – | – | – | – |
|
| – | – | 0.3 | – | 0.1 |
|
| 0.1 | – | – | – | – |
|
| 0.3 | 0.4 | 0.4 | 0.5 | 0.8 |
|
| 0.1 | – | – | 0.2 | – |
|
| – | – | – | – |
|
|
|
|
|
|
|
|
| Characteristic | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| Source | Fen soil | River bank soil | Savannah soil | Coniferous soil | Peatland soil |
| Cell shape | Rod | Rod | Rod | Rod | Rod |
| Cell length/width (µm) | 1.2×0.45 | 1.0–2.0 | 1.0–2.5 | 0.6–1.2 | 2–10 |
| Cell division | BF | BF | BF |
| BF |
| Motility | − | (+) | (+) | + | + |
| Pigmentation | Yellow-to-pale pink | Cream pink | Bright pink | White | White beige |
| Spores | − | − | − | − | − |
| Capsule/EPS formation | + | + | + | + | − |
| Oxidase | − | − | − | − | − |
| Catalase | (+) | + | + | − | (+) |
| Quinone | MK-8 | MK-8 | MK-8, MK-7 | MK-8 | MK-8 |
| Metabolism | AC | AC | AC | AC | AnC |
| NaCl tolerance (%, w/v) | 0–0.5 | 0–1.0 | 0–0.5 | 0 | 0–0.1 |
| Temperature range/optimum (°C) | 4-40 | 11-40 | 11-40 | 12-30 | 4-35 |
| pH range/optimum | 3.7–6.0 | 3.5–8.5 | 3.5–6.5 | 3–6 | 3.0–7.5 |
| DNA G+C content (mol%) | 56.4 | 59.6 | 58.5 | 57.3 | 57.6 |
| Lipids |
- —http://dx.doi.org/10.13039/501100001659 Deutsche Forschungsgemeinschaft
- —http://dx.doi.org/10.13039/501100001659 Deutsche Forschungsgemeinschaft
- —http://dx.doi.org/10.13039/501100001659 Deutsche Forschungsgemeinschaft
- —http://dx.doi.org/10.13039/501100001659 Deutsche Forschungsgemeinschaft
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Anaerobic Digestion and Biogas Production · Microbial Community Ecology and Physiology
Data Summary
All supporting data have been provided within the article or through supplementary data files.
Introduction
Although the Acidobacteriota represent one of the most abundant fractions of the active bacterial community in different soil environments [13], comparably low numbers of described strains are available. Due to improved cultivation attempts over the past years, the number of described species has been increased to 81. A high percentage of the known and described acidobacterial species belongs to the genera Granulicella [4], Telmatobacter [5], Edaphobacter [6], Tunturiibacter [7] and Terriglobus [8] within the order Terriglobales [9]. While the genera Edaphobacter, Tunturiibacter, Granulicella and Terriglobus harbour at least four species, only one or two species are described for the genera Occallatibacter [10], Telmatobacter [5] and Terracidiphilus [11] so far. Interestingly, Terriglobales dominate in soils with low pH (~3.7–5.0), such as fen and peatland soils [1215], and display different metabolic potentials, such as polysaccharide hydrolysis, sugar utilization, aerobic respiration, several fermentative capabilities and hydrogen oxidation [1517], which were proven by physiological characteristics of isolated species or indicated by genomic analyses.
Terriglobales strains were mainly isolated by the use of specialized media imitating nutrient content, pH value, oxygen relation and temperature of the original habitat. However, soil bacteria are subjected to varying environmental conditions, such as water availability, temperature changes and oxygen concentrations caused by seasonal changes and weather impacts. Fen soils, which are characterized by low and stable pH values, high carbon content and high water saturation, are also directly influenced by seasonal changes and precipitation values. Therefore, varying conditions during isolation experiments, i.e. oxygen availability, would better imitate soil conditions and could enable the isolation of previously unculturable acidobacterial strains.
In the present study, we describe one new representative of the Acidobacteriota isolated from fen soil, representing a new species within the genus Occallatibacter, within the order Terriglobales, isolated by the use of varying oxygen conditions during the cultivation process.
Origin and isolation
Strain JP12^T^ was isolated from a soil sample that was collected from the Schlöppnerbrunnen II fen (50° 8′ 8.27″ N 11° 52′ 48.3″ E) close to Bayreuth, Germany, in December 2018, and was subsequently stored at 4 °C. In the laboratory, 5 g of fen soil (pH 4–5) was mixed with 49.5 ml of MES buffer (10 mM, pH 5.0), serially diluted and inoculated in SSE/HD 1:10 medium (DSMZ medium 1426 – https://www.dsmz.de/microorganisms/medium/pdf/DSMZ_Medium1426.pdf; Leibniz-Institute DSMZ - German Collection of Microorganisms and Cell Cultures GmbH). The SSE/HD 1:10 medium contained soil solution equivalent (SSE) [18], was supplemented with a 10-vitamin solution [19] and a trace element solution SL-10 [20], and was solidified with purified agar (Oxoid, Basingstoke, UK), if indicated. 180 µl SSE/HD 1:10 medium (pH 5.0) were mixed with 20 µl of the respective soil solution suspensions in 96-well plates and incubated at 20 °C in the dark under changing oxygen conditions – 1 week under oxic conditions (normal laboratory conditions; 21% O_2_) and 1 week under a microaerobic atmosphere (candle jar technique; 3–5% O_2_) – without shaking. Additionally, 100 µl of the serial dilution step 10^-3^ were equally spread on SSE/HD 1:10 medium plates (pH 5.0) and incubated as described above. After 4 weeks of incubation, 26 turbid wells of the liquid-medium approach and 42 colonies of the solid-medium approach were screened for the presence of Acidobacteriota by PCR employing the primer pair 27f and 1492r [21] (see Supplementary Material for details) and subsequent Sanger sequencing. Thirteen Acidobacteriota-positive cultures were purified by subsequent restreaking on SSE/HD 1:10 agar plates until strain JP12^T^ was isolated, which was checked by colony PCR and Sanger sequencing.
16S rRNA gene phylogeny
The 16S rRNA gene sequence of JP12^T^ was amplified by colony PCR employing the primer pair 8f [22] and 1492r [21] and subsequent Sanger sequencing. blast (National Center for Biotechnology Information) analysis placed JP12^T^ in the vicinity of the type strains of Occallatibacter savannae A2-1c^T^ (16S rRNA sequence identity value of 96.7%), Telmatobacter bradus TPB6017^T^ (96.4%), Occallatibacter riparius 277^T^ (96.3%) and Terracidiphilus gabretensis S55^T^ (96.1%).
The 16S rRNA gene sequence of JP12^T^ was added to the small subunit ribosomal RNA non-redundant reference database silva version 138.1 (https://www.arb-silva.de/) via the programme package arb for phylogenetic tree calculations. The alignment was corrected manually based on the secondary-structure information after the implementation by the automated alignment. All three different algorithms, neighbour-joining, maximum-parsimony and maximum-likelihood algorithms (termini filter; 1,376 valid columns between position 63 and 1,439 of the Escherichia coli 16S rRNA reference gene; 1,000 bootstrap resamplings), placed JP12^T^ within the Terriglobales, in the vicinity of the strains T. bradus TPB6017^T^, O. savannae A2-1c^T^, O. riparius 277^T^ and T. gabretensis S55^T^ (Figs 1, S1 and S2, available in the online Supplementary Material).
Rooted neighbour-joining phylogenetic tree (Felsenstein correction) based on almost full-length 16S rRNA gene sequences showing the relationship of strain JP12T and related type strains. Bootstrap values are expressed as percentages of 1,000 replicates and are indicated at the respective branching points. The following sequences were used as outgroups: Novipirellula rosea LHWP3T (JF748734), Blastopirellula marina DSM 3645T (X62912) and Pirellula staleyi DSM 6068T (CP001848). The bar indicates 10% nucleotide divergence.
Ecological distribution
To examine the distribution of strain JP12^T^ in the environment, the 16S rRNA gene sequence of JP12^T^ was compared via blast (https://blast.ncbi.nlm.nih.gov/Blast.cgi) against acidobacterial sequences from cultured and uncultured environmental samples. Interestingly, all 16S rRNA gene sequence hits with identity values above 99.0% derived from soil samples from German forest soils [23], polychlorinated biphenyl-polluted soil [24], oak forest soil [25], pristine soil [26] and Australian pasture soils [27], indicating an adaptation of JP12^T^ and closely related sequence types to the soil environment.
Phylogenomic placement
The DSMZ sequencing service department sequenced the genome of JP12^T^ on the PacBio Sequel IIe instrument (Pacific Biosciences, Menlo Park, CA, USA; details of the methods used are given in the Supplementary Material). The raw sequence data were quality checked, filtered and resulted in 80,563 sequence reads with a mean subread length of 4,324 bp. The HGAP version 4 protocol included in the SMRT Link analysis software assembled the genome of JP12^T^, which finally displayed a length of 5,987,865 bp and a G+C content of 56.4 mol%. As high sequencing coverage values could be obtained (487 mean coverage), a correction of the genome by short-read sequencing was not necessary. After the assembly, the genome was automatically oriented after oriC and subsequently analysed with the Prokka pipeline, resulting in 5,040 coding sequence regions, 3 rRNAs and 76 tRNAs (Table S1). CheckM analysis (v1.2.2) provided by NCBI (National Center for Biotechnology Information) confirmed 97.22% completeness and 2.4% contamination of the genome of JP12^T^.
To confirm the phylogenetically closest relatives of JP12^T^, a phylogenomic tree was calculated based on 120 single-copy marker genes of all available acidobacterial type strain genomes, which were extracted and aligned with GTDB-Tk v2.4.0 [28]. The tree calculation was performed with IQ-TREE 2, multicore version 2.4.0, using ultrafast bootstraps (n=1,000) after automatic substitution model selection, selecting LG+F+R9 as the best-fit model [29]. The resulting maximum-likelihood tree identified Occallatibacter savannae A2-1c^T^, O. riparius DSM 25168^T^ and Terracidiphilus gabretensis S55^T^ as the phylogenetically closest relatives of JP12^T^ (Fig. 2).
Phylogenomic reconstruction of the Acidobacteriota based on a concatenated alignment of 120 single-copy bacterial marker proteins. Blastopirellula marina DSM 3645T, Nitratidesulfovibrio liaohensis XJ01T, Terripilifer ovatus H3SJ34-1T and Asaia lannensis NBRC 102526T were used as outgroups. The bar indicates 10% sequence divergence.
Whole-genome similarity indices
Genome similarity indices of JP12^T^ and its phylogenetically closest relatives were used for species and genus demarcation. Average nucleotide identity (ANI), average amino acid identity (AAI), digital DNA–DNA hybridization (dDDH) and percentage of conserved proteins (POCPs) were calculated employing the OrthoANIu algorithm via the EzBioCloud web service webpage (https://www.ezbiocloud.net/tools/ani) [30], the EzAAI tool [31], the Genome-to-Genome Distance Calculator version 3.0 (https://ggdc.dsmz.de/ggdc.php) and the POCP-nf pipeline (https://github.com/hoelzer/pocp?tab=readme-ov-file#run) [3235], respectively.
ANI values of 71.8%, 73.9%, 71.2% and 71.2%, as well as dDDH values of 20.7%, 19.5%, 19.3% and 20.6%, of strain JP12^T^ compared with Occallatibacter riparius 277^T^ and 307, O. savannae A2-1c^T^, Terracidiphilus gabretensis S55^T^ and Telmatobacter bradus DSM 23630^T^ (Table 1) clearly showed that JP12^T^ represents a new species within the Telmatobacter–Occallatibacter–Terracidiphilus clade.
AAI values of 70.0% and 76.8% for strain JP12^T^ compared with O. riparius 277^T^ and O. savannae A2-1c^T^, respectively, exceeded the suggested genus demarcation threshold of 68% [3637]. In addition, an AAI value of 69.22% between JP12^T^ and T. gabretensis S55^T^ slightly exceeded the genus demarcation threshold. Interestingly, an AAI value of 69.9% between O. riparius 277^T^ and T. gabretensis S55^T^ and an AAI value of 69.0% between O. savannae A2-1c^T^ and T. gabretensis S55^T^ would suggest that T. gabretensis S55^T^ also belongs to the genus Occallatibacter. Therefore, strain JP12^T^ represents a new species within the Occallatibacter genus.
An AAI value of 67.5% between O. riparius 277^T^ and T. bradus DSM 23630^T^ and an AAI value of 67.8% between O. savannae A2-1c^T^ and T. bradus DSM 23630^T^ fell slightly below the suggested genus demarcation value of 68%, confirming that T. bradus DSM 23630^T^ represents a separate genus within the Occallatibacter–Telmatobacter–Terracidiphilus clade, whereas an AAI value of 68.08% between JP12^T^ and T. bradus DSM 23630^T^ slightly exceeded the suggested threshold of 68% for genus demarcation (Table 1).
To support the findings of the AAI values, POCP values were additionally calculated. Strain JP12^T^, the three Occallatibacter strains and T. gabretensis S55^T^ displayed POCP values of 54.12–92.47% among each other, while T. bradus DSM 23630^T^ showed POCP values of 45.25–48.44% compared with JP12^T^, the three Occallatibacter strains and T. gabretensis S55^T^ (Table 1). The suggested POCP demarcation value for the definition of a genus is 50% [35]. Therefore, the calculated POCP values confirmed that strains JP12^T^ and T. gabretensis S55^T^ would represent species of the Occallatibacter genus and that T. bradus DSM 23630^T^ would further represent a separate genus within the Terriglobales.
Genomic features
The annotation and analysis platform implemented on the Bacterial and Viral Bioinformatics Resource Center (BV-BRC) webpage assigned 22% of the genome sequence of JP12^T^ to known subsystem categories. The groups with the most assigned genes were ‘protein synthesis’ (113 genes), ‘cofactors, vitamins, prosthetic groups’ (81), ‘amino acids and derivatives’ (111) and ‘stress response, defence and virulence’ (94). While most of the known and strongly represented subsystem categories contained genes for the ‘normal’ metabolism, higher gene numbers of ‘stress response, defence and virulence’ would explain the broad temperature and pH growth and tolerance ranges detected for JP12^T^ (see Section Growth ranges). As fen soils, such as Schlöppnerbrunnen II, from which JP12^T^ was isolated, are characterized by high carbon content, high water saturation and underlie seasonal fluctuations in temperature and water availability, strain JP12^T^ had to be adapted to changing temperature values and also pH values because of changes in the water capacity and oxygen availability of the fen soils.
Twenty-nine genes of JP12^T^ were identified as antibiotic resistance genes, suggesting a role in the survival of Acidobacteriota strains despite their low biomass production and a potential growth advantage against fast-growing bacteria of the Pseudomonadota, Actinomycetota and Bacteroidota. The genome of JP12^T^ was analysed via the antiSMASH pipeline (https://antismash.secondarymetabolites.org) [38]. The options ‘relaxed detection strictness’ and ‘all possible extra features’ detected ten secondary metabolite regions in strain JP12^T^, encoding either terpene (or terpene-precursor), hydrogen-cyanide RiPP-like (ribosomally synthesized and post-translationally modified peptides), T3PKS (type III polyketide synthases), NRPS-like (nonribosomal peptide synthetase) or T1PKS (type I polyketide synthases) compounds (Table S2). The presence of cbb3-type cytochrome-c-oxidase subunit I and putative bifunctional cbb3-type cytochrome-c-oxidase subunit II/cytochrome-c explains why strain JP12^T^ was able to survive under microaerophilic conditions in the fen soil, during the isolation process and under microaerophilic conditions in the laboratory. Genes for the degradation of complex carbon compounds, such as xylan (endo-1,4-β-xylanase, EC 3.2.1.8) and laminarin (laminarinase, EC 3.2.1.39), as detected in the genome of JP12^T^, confirm that strain JP12^T^ is adapted to the harsh conditions of the soil environment and is able to use complex hemicellulose as growth substrates. However, the ability to degrade complex substrates could not be proven under laboratory conditions. Interestingly, in the genome of strain JP12^T^, genes for gamma-hexachlorocyclohexane degradation, benzoate degradation via hydroxylation, bisphenol A degradation and fluorobenzoate degradation could be determined and could hint for a potential role of the strain in bio-restoration of contaminated soils.
Morphological and physiological characterization
Strain JP12^T^ grew as pink, silk-mat-like, round colonies with 0.5–1.0 mm in diameter on SSE/HD 1:10 agar plates after 3 weeks of incubation at 28 °C. In liquid medium, JP12^T^ showed the formation of yellow-to-pale pink aggregates, which were too stable to be dissolved by normal shaking. However, during longer incubation periods in liquid culture, strain JP12^T^ seemed to lose this characteristic, and nearly no flocks were determined in the cultures anymore.
JP12^T^ was stained according to the Gram, India ink and malachite green methods, respectively, and subsequently examined by light microscopy (Zeiss Axio Scope A.1 equipped with an AxioCam MRc camera; Carl Zeiss, Jena, Germany) to determine cell wall structures, cell size, presence of capsules and endospores, as described before [39]. Strain JP12^T^ formed non-motile rods (0.45±0.10×1.20±0.35 µm in width and length, respectively; n=10 cells) that divided by binary fission (Fig. 3a). While a Gram-negative cell wall structure and the presence of capsules could be confirmed for JP12^T^, as for the two closely related Occallatibacter strains [10] and Terracidiphilus gabretensis S55^T^ [11], no endospores were detected, comparable to all other known Acidobacteriota strains. Additionally, the formation of aggregates, as seen in liquid cultures, could be confirmed by the presence of exopolysaccharide (EPS)-like structures in the scanning (Fig. 3b) and transmission electron micrographs (Fig. 3c).
Phase-contrast photomicrograph (a), scanning electron micrograph (b) and transmission electron micrograph (c) of strain JP12T. Scale bars, 1 µm.
Anaerobic growth of strain JP12^T^ tested negative in duplicates on SSE/HD 1:10 medium (pH 5.0) under anoxic conditions by the application of the Oxoid™ AnaeroGen™ Compact System for 6 weeks at 28 °C. Incubation of strain JP12^T^ for 4 weeks at 28 °C in a candle jar confirmed that strain JP12^T^ could grow under microaerophilic conditions and also showed a biomass production capacity comparable to that under oxic conditions. For better comparability with other strains of the Occallatibacter–Terracidiphilus clade, the physiological tests were performed under oxic conditions.
Growth ranges
Liquid SSE/HD 1:10 medium was used to test the growth ranges and optima of JP12^T^ regarding temperature (4, 10, 15, 20, 22, 24, 27, 30, 36, 40 and 45 °C) and pH values (2.5–10). The pH of the medium for the temperature test was buffered at pH 5.0, whereas the pH of the medium for the pH tolerance test was buffered with the substrates MES (used for pH values 2.5–6.5), HEPES (used for pH values 7.0–8.0), HEPPS (used for pH value 8.5) and CHES (used for pH values 9.0–10.0). All buffers displayed a final concentration of 10 mM. NaCl concentrations of 0%, 0.25%, 0.50%, 1.0%, 3.0%, 5.0%, 7.5% and 10% (w/v) were adjusted in liquid HD 1:10 medium (DSMZ medium 1124; https://www.dsmz.de/microorganisms/medium/pdf/DSMZ_Medium1124.pdf) to determine the salinity tolerance. JP12^T^ showed growth at temperatures between 4 and 40 °C (best between 24 and 30 °C), grew at pH values between 3.7 and 6.0 and showed best growth at pH values of 4.1–5.6. JP12^T^ could only endure minor amounts of NaCl (0–0.5%, best at 0.25% NaCl, w/v).
Nutritional tests and enzymatic activities
For the substrate utilization test of strain JP12^T^ (including different substrates such as sugars, keto acids, organic acids, amino acids, alcohols, casamino acids, laminarin, peptone, yeast extract, casein hydrolysate and Tween 80), triplicates of liquid 1x SSE solution [18] were adjusted at pH 5.0, supplemented with 1 ml l^−1^ vitamin solution, 1 ml l^−1^ trace element SL-10 solution and the respective test substrates at concentrations between 0.5 and 10 mM or rather 0.001% and 0.05% (w/v; Table S3). After 8 weeks of incubation in the dark at 20 °C, weak growth or rather growth was positively evaluated if the OD_660nm_ mean values (optical density) of the substrates exceeded the control values (culture without substrate) by a factor of 1.2–1.5 or by a factor larger than 1.5, respectively. JP12^T^ grew on various organic acids and amino acids (Table S4; further details are given in the species description). Interestingly, JP12^T^ grew well on DSMZ medium 1426 containing glucose, yeast extract and peptone, but did not grow on any of these substrates tested separately. Other acidobacterial strains, such as Blastocatella fastidiosa A2-16^T^, Brevitalea aridisoli Ac_11_E3^T^, Arenimicrobium luteum Ac_12_G8^T^ and Stenotrophobacter terrae Ac_28_D10^T^, also seemed to prefer a combination of substrates for growth (growth in DSMZ medium 1426) rather than the use of certain single substrate compounds [39].
After 4 weeks of incubation of strain JP12^T^ on solidified SSE medium supplemented with 1 ml l^−1^ vitamin solution, 1 ml l^−1^ trace element SL-10 solution, 0.005% (w/v) yeast extract and complex substrates such as starch, cellulose, pectin, colloidal and non-colloidal chitin, polygalacturonic acid and xylan (0.5 g l^−1^ final concentration), the degradation potential was detected by staining the agar with the respective staining solutions and observing the appearance of clearing zones as reported earlier [40]. No clearing zones, and therefore no degradation potential, could be confirmed for the different tested complex substrates. Additionally, API^®^ZYM and API^®^20NE galleries (bioMérieux, Marcy-l'Étoile, France) were employed to determine further exoenzyme activity potential, the formation of indole, aesculin degradation, urease activity, reduction of nitrogen compounds and the assimilation capacity of different carbon compounds (Table S5). Catalase and cytochrome-c-oxidase activities were tested by standard protocols [4142]. JP12^T^ displayed only minor catalase activity reactions and no reaction of cytochrome-c-oxidase.
Chemotaxonomic characterization
For fatty acids, polar lipids and respiratory quinone analyses, cells of JP12^T^, Occallatibacter riparius DSM 25168^T^/DSM 25169, O. savannae DSM 25170^T^ and Terracidiphilus gabretensis DSM 100509^T^ were grown in SSE/HD 1:10 medium (pH 5.5) at 25 °C for 14 days. Only Telmatobacter bradus DSM 23630^T^ was grown in half-strength R2A liquid medium (https://bacmedia.dsmz.de/medium/830) at 25 °C for 14 days, as the strain could not be grown in SSE/HD 1:10 medium at all.
To extract membrane-spanning lipids such as iso-diabolic acid, a method adapted from Sinninghe Damsté et al. [43] was employed. A total of ∼20 mg of wet biomass was treated with 1,900 µl methanol and 100 µl of 1M HCl solution in a sealed reaction tube and heated at 65 °C for 3 h. Following this, the pH value was adjusted to 7 using a KOH solution (2 M KOH:methanol, 1:1, v/v). The sample was then evaporated to dryness. Methylation was carried out by adding 1 ml methanol and 50 µl concentrated sulphuric acid, followed by overnight incubation at 50 °C. Subsequently, 500 µl of saturated NaCl solution was added, and then the mixture was extracted twice with 1 ml hexane:dichloromethane (4:1, v/v). The combined organic phases were evaporated and redissolved in methyl tert-butyl ether for analysis via GC with flame ionization detection (on an Agilent Technologies 6890B).
Equivalent chain length values were calculated analogously to the Sherlock Microbial Identification System to provide peak naming according to the TSBA6 database. Identification of fatty acids was performed via mass spectrometry [44]. The position of double bonds of monosaturated fatty acids was determined by derivatization with dimethyl sulphide [45].
The fatty acid profiles of JP12^T^ showed very similar composition patterns, with iso-C_15:0_ (26.5%), iso-C_17:1_ ω7c (11.3%), iso-C_17:0_ (6.3%) and iso-diabolic acid (49.2%) as the major fatty acids, compared with its closest phylogenetic relatives – O. riparius DSM 25168^T^/DSM 25169, O. savannae DSM 25170^T^, T. gabretensis DSM 100509^T^ and T. bradus DSM 23630^T^ (Table 2). Only the presence of an iso-diabolic acid likely isomer (16.9%) in T. gabretensis DSM 100509^T^ and a threefold higher amount of iso-C_15:1_ ω7c (3.0%) in T. bradus DSM 23630^T^ differed the strains within the Occallatibacter–Telmatobacter–Terracidiphilus clade from each other (Table 2).
Respiratory quinones were extracted from 5 mg of wet biomass via solid-phase extraction and analysed via reversed-phase HPLC coupled to a diode array detector [44]. MK-8 was detected as the sole respiratory quinone of JP12^T^.
Polar lipids of strain JP12^T^ were extracted using a modified Bligh and Dyer extraction [46] and analysed via HPLC-MS as described previously [47]. Phosphatidylglycerol, phosphatidylethanolamine and ornithine-containing lipids were identified as major polar lipids. In addition, lysophosphatidylethanolamine, phosphatidylinositol, diphosphatidylglycerol and high-mass intact polar lipids occurred in small amounts.
Taxonomic conclusion
Although the 16S rRNA phylogenetic trees identified Telmatobacter bradus TPB6017^T^ as the phylogenetically closest relative of the strain JP12^T^, the results of the fatty acid analysis, phylogenomic tree and genome comparison results (POCP and AAI values) suggest that JP12^T^ would represent a new species within the Occallatibacter genus. In addition, JP12^T^ and T. bradus TPB6017^T^ differed in physiological traits: T. bradus TPB6017^T^ does not form any aggregates or EPS-like structures and is a facultatively anaerobic, chemo-organotrophic organism, unlike JP12^T^. While all Occallatibacter strains and Terracidiphilus gabretensis DSM 100509^T^ were able to grow on SSE/HD 1:10, T. bradus DSM 26360^T^ was not able to grow on this medium at all.
Moreover, the genome comparison results, i.e. AAI and POCP values, and the phylogenomic tree analysis of the Occallatibacter–Telmatobacter–Terracidiphilus clade underlined our findings that T. gabretensis S55^T^ would represent a species within the Occallatibacter genus and that T. bradus TPB6017^T^ should not be merged with the Occallatibacter genus.
Therefore, based on the morphological, physiological and phylogenetic characteristics and whole-genome similarity indices, we propose JP12^T^ as a new species within the acidobacterial genus Occallatibacter–Occallatibacter bavaricus sp. nov. (Table 3) and the reclassification of Terracidiphilus gabretensis as Occallatibacter gabretensis comb. nov.
Description of Occallatibacter bavaricus sp. nov.
Occallatibacter bavaricus (ba.va’ri.cus. N.L. masc. adj. bavaricus, pertaining to Bavaria, to indicate the original habitat of the type strain of the species, which was isolated from fen soil close to Bayreuth, Germany).
Cells are Gram-negative, non-motile rods with 0.45 µm in width and 1.20 µm in length and divide by binary fission. Cytochrome-c-oxidase activity is negative, and catalase activity is weakly positive. Neither motility nor spores are observed, but EPS-like structures are present. Capsules are present. In liquid cultures, strong yellow-to-pale pink aggregates are formed, which cannot be dissolved by shaking. Long-term incubations and several transfers to fresh media reduce this effect. On agar plates, pink, silk-mat-like, round colonies with 0.5–1.0 mm in diameter are formed.
Grows occurs at temperature values of 4–40 °C (best between 24 and 30 °C) and at pH values of 3.7–6.0 (best between 4.1 and 5.6). Small amounts of NaCl (0–0.5%; best 0.25% NaCl, w/v) support the growth.
The major fatty acids are iso-C_15:0_, C_16:0_, iso-C_17:1_ ω7c, iso-C_17:0_ and iso-diabolic acid. MK-8 is the major respiratory quinone. Phosphatidylglycerol, phosphatidylethanolamine and ornithine-containing lipids are the major polar lipids. In addition, glycophospholipid, lysine lipid and high-mass intact polar lipid occur in small amounts. Utilizes arabinose, N-acetylgalactosamine, maleic acid, 1,2-butandiole, 2,3-butandiole, 1,2-propandiole, alanine, arginine, asparagine, isoleucine, ornithine, proline, acetate, butyrate, β-hydroxybutyrate, γ-hydroxybutyrate, isobutyrate, tyrosine, serine, phenylalanine, glycine, leucine, histidine, valine, methionine, threonine, succinate, nicotinic acid, Tween 80, adipate, protocatechuate, shikimate, aspartate, glutamate, laminarin, malate, citrate, tartrate, fermented rumen extract, isovaleric acid, heptanoic acid, fumarate, trimethoxybenzoate, crotonate, 2-oxoglutarate, sodium pyruvate, acetoin, ascorbate, glyoxylate, 2-oxovalerate and 2-oxogluconate. Grows weakly on caproate, caprylate, dulcitol, ethylene glycol, erythrose, erythrulose, α-hydroxybutyrate, isocitrate, levulinate, 2-oxogluconate, cysteine, glutamine, benzoate, tryptophan and formate. No growth is observed on glucose, lactose, fructose, cellobiose, galactose, mannose, melezitose, raffinose, fucose, sorbose, lyxose, maltose, rhamnose, sucrose, trehalose, xylose, adonitol, arabitol, mannitol, myo-inositol, sorbitol, xylitol, l-lysin-HCl, hydroxy-l-proline, casamino acids, casein hydrolysate, yeast extract, glycolate, malonate, propionate, oxaloacetate, lactate, butanol, ethanol, glycerin, methanol, propanol, peptone, N-acetylglucosamine, glucosamine, gluconate, glucuronate, lyxitol, crotonate, 2-oxoglutarate, sodium pyruvate, acetoin, ascorbate or glyoxylate.
Positive for leucine arylamidase, acid phosphatase, naphthol-AS-BI-phosphohydrolase, β-galactosidase (API^®^ZYM and API^®^20NE), β-glucuronidase, α-glucosidase, β-glucosidase (API^®^ZYM and API^®^20NE), N-acetylglucosaminidase and α-fucosidase. Weakly positive for esterase C4, esterase lipase C8, valine arylamidase and α-galactosidase.
The type strain is JP12^T^ (=DSM 110680^T^=CECT 30267^T^) and was isolated from fen soil collected at Schlöppnerbrunnen II in Germany. The DNA G+C content of the type strain is 56.4 mol%, and the GenBank/EMBL/DDBJ accession numbers for the 16S rRNA gene sequence and the genome sequence are OQ656429 and CP121196, respectively.
Description of Occallatibacter gabretensis comb. nov.
Occallatibacter gabretensis (ga.bret.en’sis. N.L. masc. adj. gabretensis, pertaining to Gabreta, the Celtic name of the Bohemian Forest, the mountain range in central Europe where the type strain was isolated).
Basonym: Terracidiphilus gabretensis Garcia-Fraile et al. 2016 [11].
The description of the species is provided by García-Fraile et al. [11]. According to phylogenetic and phylogenomic data, Terracidiphilus gabretensis was transferred into Occallatibacter.
The type strain of the species is S55^T^ (=CECT 8791^T^=NBRC 111238^T^).
The DNA G+C content of the type strain is 57.3 mol%, and the GenBank/EMBL/DDBJ accession numbers for the 16S rRNA gene sequence and the genome sequence are KP120762 and LAIJ01000000, respectively.
Emended description of the genus Occallatibacter Foesel et al. 2016
The description is as given by Foesel et al., 2016 [10], with the following modifications: mostly catalase-positive but oxidase-negative. Tumbling events as motility and polysaccharide degradation might occur. The DNA G+C content ranges from 56.4 to 59.6 mol%.
Supplementary material
10.1099/ijsem.0.007086Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Janssen PH Identifying the dominant soil bacterial taxa in libraries of 16S r RNA and 16S r RNA genes Appl Environ Microbiol 2006721719172810.1128/AEM.72.3.1719-1728.200616517615 PMC 1393246 · doi ↗ · pubmed ↗
- 2Jones RT Robeson MS Lauber CL Hamady M Knight R et al A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses ISME J 2009344245310.1038/ismej.2008.12719129864 PMC 2997719 · doi ↗ · pubmed ↗
- 3Lauber CL Hamady M Knight R Fierer N Pyrosequencing-based assessment of soil p H as a predictor of soil bacterial community structure at the continental scale Appl Environ Microbiol 2009755111512010.1128/AEM.00335-0919502440 PMC 2725504 · doi ↗ · pubmed ↗
- 4Pankratov TA Dedysh SN Granulicella paludicola gen. nov., sp. nov., Granulicella pectinivorans sp. nov., Granulicella aggregans sp. nov. and Granulicella rosea sp. nov., acidophilic, polymer-degrading acidobacteria from Sphagnum peat bogs Int J Syst Evol Microbiol 2010602951295910.1099/ijs.0.021824-020118293 · doi ↗ · pubmed ↗
- 5Pankratov TA Kirsanova LA Kaparullina EN Kevbrin VV Dedysh SN Telmatobacter bradus gen. nov., sp. nov., a cellulolytic facultative anaerobe from subdivision 1 of the Acidobacteria, and emended description of Acidobacterium capsulatum Kishimoto et al. 1991 Int J Syst Evol Microbiol 20126243043710.1099/ijs.0.029629-021460138 · doi ↗ · pubmed ↗
- 6Koch IH Gich F Dunfield PF Overmann J Edaphobacter modestus gen. nov., sp. nov., and Edaphobacter aggregans sp. nov., acidobacteria isolated from alpine and forest soils Int J Syst Evol Microbiol 2008581114112210.1099/ijs.0.65303-018450699 · doi ↗ · pubmed ↗
- 7Messyasz A MännistöMK Kerkhof LJ Häggblom MM Genome analysis and description of Tunturibacter gen. nov. expands the diversity of Terriglobia in tundra soils Environ Microbiol 202426 e 1664010.1111/1462-2920.1664038775217 · doi ↗ · pubmed ↗
- 8Eichorst SA Breznak JA Schmidt TM Isolation and characterization of soil bacteria that define Terriglobus gen. nov., in the phylum Acidobacteria Appl Environ Microbiol 2007732708271710.1128/AEM.02140-0617293520 PMC 1855589 · doi ↗ · pubmed ↗