Hot Mitochondria and the Second Law of Thermodynamics
Alexei Tkachenko, Belem Yoval-Sánchez, Alexander Galkin

TL;DR
This paper examines whether mitochondria can create large temperature differences inside cells and concludes that such differences are unlikely due to thermodynamic limits.
Contribution
The study provides a model-independent thermodynamic analysis using the Second Law to show that mitochondria cannot sustain large temperature differences.
Findings
The maximum temperature difference across the mitochondrial membrane is limited to a fraction of a degree.
Nonequilibrium processes like proton pumping cannot create significant local temperature increases.
Fluorescence thermometry claims of large temperature hotspots are inconsistent with biophysics.
Abstract
Mitochondria are central hubs of cellular bioenergetics, converting chemical free energy into ATP while inevitably releasing heat during respiration. Fluorescence-based thermometry has been interpreted to show intracellular “hot spots” more than 10 °C above the bulk physiological temperature, implying that mitochondria might operate far outside conventional thermal bounds. Such claims, however, appear inconsistent with basic biophysics: the small size of mitochondria, their aqueous and highly conductive environment, and their limited power output all argue against large steady-state temperature gradients. This discrepancy has prompted renewed scrutiny of both the physical limits of intracellular heat transfer and the biological interpretation of nanoscale thermal measurements. A key open question is whether nonequilibrium biochemical processes, such as respiration-driven proton pumping,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
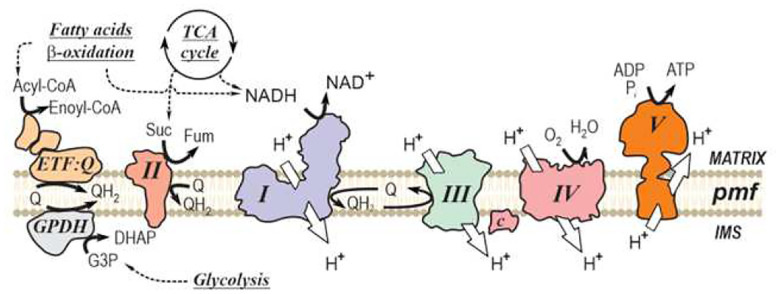 Figure 1
Figure 1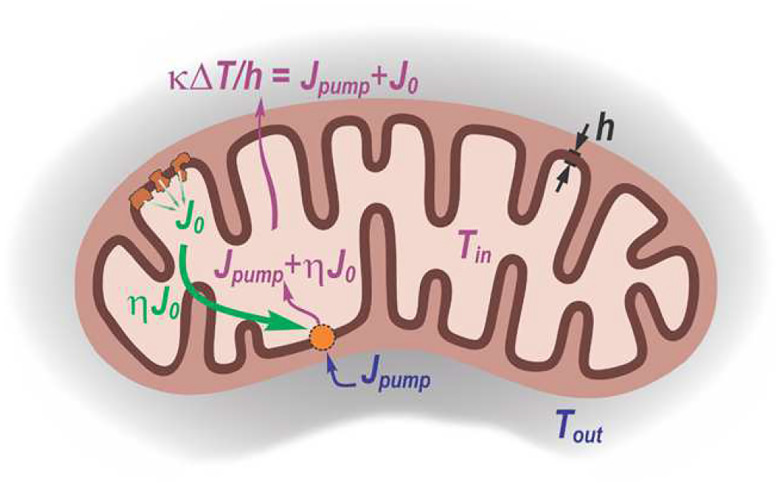 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMitochondrial Function and Pathology · ATP Synthase and ATPases Research · thermodynamics and calorimetric analyses
INTRODUCTION
I.
Cellular aerobic catabolism relies on the breakdown of sugars, lipids, and proteins into smaller molecules, followed by their oxidation in mitochondria through a series of exergonic reactions. A fraction of the released free energy is conserved and stored in the form of adenosine triphosphate (ATP), the universal energy currency of all cellular life forms. This process was termed oxidative phosphorylation by W. Engelhardt in the late 1930s [1, 2], with its components located at the inner mitochondrial membrane (Fig. 1). During the oxidative phase, several respiratory substrates (NADH, succinate, glycerol 3-phosphate, acyl-coenzyme A, and others) are oxidized by membrane-bound dehydrogenases. The derived electrons are transferred to the membrane-soluble carrier ubiquinone and subsequently passed to Complex III. From there, electrons are shuttled via cytochrome c to Complex IV, where molecular oxygen serves as the terminal electron acceptor, producing water. The free energy released during these redox reactions drives proton translocation by Complexes I (NADH dehydrogenase), III, and IV across the inner membrane, from the matrix to the intermembrane space, generating the proton-motive force (pmf) (Fig. 1). During the phosphorylation phase, the backflow of protons from the intermembrane space to the matrix drives the synthesis of ATP from ADP and inorganic phosphate at Complex V or ATP synthase. In aerobic cells, the majority of ATP is produced in mitochondria via oxidative phosphorylation.
The thermodynamic characteristics of ATP production during oxidative phosphorylation have been examined by thousands of researchers over the past century. The thermodynamic efficiency of the ATP formation process, i.e., the percentage of the total free energy change that is stored in the form of ATP, is often regarded as being high (~ 60–70%) [3]. When the rates of substrate oxidation and ATP synthesis are taken into account, it has been estimated that mitochondria dissipate about 80% of the energy derived from oxidized substrates as heat [4]. This observation suggests that, under certain conditions, the released heat could potentially increase the local temperature in the vicinity of mitochondria, with some reports proposing values as high as 10–15 °C above the surrounding environment [5]. Most estimations assume that mitochondria are membrane-formed spheres, with heat production occurring across the surface [6, 7]. However, in situ, heat is produced across the cristae membranes arranged in parallel, which may help to transiently retain heat within the mitochondrial matrix [8].
Several experimental approaches have been developed to assess mitochondrial heat production and temperature changes in intact cells [9]. These include mitochondria-targeted fluorescent proteins such as tsGFP1-mito, gTEMP, emGFP-Mito [10–12], fluorescent polymers [13], and low-molecular-weight sensors like rhodamine, Mito-RTP [14–17], rhosamine-MTY [5], or BODIPY-based reporters [18]. Application of these probes has revealed temperature heterogeneity within a cell. However, a number of biochemical factors such as pH, calcium concentration, membrane potential, and redox state can influence probe fluorescence, potentially leading to apparent signal changes that may be misinterpreted as temperature fluctuations [5, 19]. Therefore, the conclusions of Chrétien et al. [5] were challenged by direct measurements of the effects of temperature on isolated intact mouse liver mitochondria. These studies demonstrated that the stability of respiratory complexes and supercomplexes declines sharply at temperatures above 43 °C, indicating that such high temperatures cause structural and functional damage to mitochondrial machinery. At the same time, mitochondrial-respiration-mediated temperature increase contradicts established principles of physics governing energy transfer, including absolute heat release, thermal conductivity, and the geometric constraints of the cellular environment.
In this work, we re-examine the physical limits of mitochondrial temperature using two complementary approaches. First, we refine the classical diffusion-based estimate, incorporating realistic mitochondrial geometry and maximal metabolic power rather than relying on a coarse cell-level approach. This analysis confirms that even under extreme uncoupled respiration, the steady-state surface temperature rise remains extremely small. Second, we address the plausible loophole in diffusion-based arguments: the possibility that nonequilibrium biochemical activity could actively sustain internal temperature gradients. To examine this scenario, we derive a general bound, based on the Second Law of Thermodynamics, which establishes that “the entropy of a closed system tends to increase over time,” on the largest temperature difference that any biochemically powered heat-pump mechanism could maintain across the inner membrane. By combining this thermodynamic constraint with the known geometric and biophysical parameters, we demonstrate that even the most favorable nonequilibrium mechanisms cannot produce more than a fraction of a degree of heating.
RESULTS
II.
Thermal diffusion result: the 105 gap
A.
We begin by refining the standard estimate of the temperature difference between a mitochondrion and its surroundings, based on Fourier’s law of thermal diffusion:
where is the heat flux (power transfer per unit area), is the temperature gradient, and is the thermal conductivity inside the cell. A natural reference value for is that of water, [20], although recent reports suggest that the cytoplasmic conductivity may be significantly lower, [21].
Mitochondria can exist as interconnected tubular networks with a characteristic radius , or as fragmented spheroids connected end-to-end [22]. Applying Fourier’s law at a distance from the mitochondrial surface gives
Here is the local heat flux in the vicinity of the mitochondrial surface. In our estimations we deliberately adopt the upper theoretical limit, assuming that all the energy released by mitochondrial respiration is converted to heat. Under such conditions, uncoupling agents render the inner membrane permeable to protons, collapsing the proton-motive force. While this accelerates respiration, ATP synthesis ceases, and 100% of the energy released from redox reactions is dissipated as heat. Maximum heat production by mitochondria is limited by their maximal respiratory capacity, defined by the respiratory complexes content [8].
where is the total mitochondrial power consumption (using 480 kJ/mol of O_2_ as the available free energy) and is the surface area of the inner mitochondrial membrane.
In both geometries, spherical or tubular, to obtain we consider the heat flux through the surface at distance from the mitochondrion. The areas are for a sphere and for a cylinder of length . Integrating the stationary heat equation over yields the excess temperature at the mitochondrial surface:
where is the radius of the “neighborhood” surrounding the tubular mitochondrion, determined by the proximity criterion. By construction,
is the volume fraction occupied by the mitochondrial network.
To estimate the power dissipation per unit area, we consider cardiomyocytes as the most metabolically intensive cell type [22]. It has been reported that they consume oxygen at rates as high as 1–2 fmol cell^−1^ s^−1^ [23, 24]. Taking an average cell volume of 35 pL [22], we estimate the mitochondrial heat production per cell volume as
The total mitochondrial volume of cardiomyocytes can reach ~ 40% of the cell volume [25, 26]. For tubular mitochondrial geometry, this volume can be related to surface area through
Putting these estimates together,
This value is consistent with alternative estimates [27], which set the typical power per mitochondrion in the range 0.1–0.01 pW. For a single mitochondrion with surface area , this power corresponds to a heat flux . Some species and cell types may reach as high as .
To estimate the upper bound for the temperature difference between the mitochondrial surface and its environment, we substitute
into Eq. (4):
This reproduces the earlier estimates commonly referred to as the “10^5^ gap” [6, 7]. Note that our treatment explicitly focuses on geometry and heat production at the level of individual mitochondria, rather than using a coarse-grained cell-scale estimate. Taken alone, this refinement would predict an even smaller , but this effect is offset by our use of deliberately extreme parameter values, particularly for and .
Thermodynamic bound on cross-membrane temperature gradient
B.
At first sight, the above analysis appears robust and sufficiently general to rule out the idea of “hot” mitochondria. However, a loophole remains: mitochondria operate under strongly nonequilibrium conditions, where biochemical processes could, in principle, sustain internal temperature gradients by harnessing the available power, much like a standard heat pump that uses mechanical work to reverse the direction of heat flow [28]. Indeed, the inner mitochondrial membrane is an intricate nonequilibrium nanomachine, and from a fundamental standpoint it is conceivable that active reactions could function as a heat pump, generating a steady-state temperature difference between the matrix (interior) and the intermembrane space (exterior). For example, a recent model [29] suggests that ATP synthase could function as a microscopic “thermal ratchet,” producing a localized temperature gradient.
Figure 2 illustrates plausible energy flows when the inner mitochondrial membrane acts as a biochemically powered heat pump. The membrane separates the mitochondrial matrix (pale yellow) at temperature from the surrounding medium (grey) at temperature . Metabolic reactions carried out by the three proton-pumping respiratory chain complexes (dark orange) provide a power input (green), of which a fraction is used to drive the heat pump. This pump transfers heat from the exterior to the interior (blue), so that the total heat flux into the matrix is (magenta). This inward flux must be balanced by conductive heat loss across the membrane of thickness , described by
where is the thermal conductivity and . The scheme illustrates the quantities used to derive the thermodynamic upper bound on the temperature difference across the inner membrane.
Below, we present a general, model-independent analysis that sets the upper bound on excess temperature inside mitochondria, based solely on the Second Law of Thermodynamics [28]. Specifically, we consider a scenario in which the inner mitochondrial membrane acts as a biochemically powered heat pump. In analogy to a classical heat engine, which uses a heat flow from a hotter to a colder reservoir to produce work, a heat pump operates in reverse, converting work to drive a heat flow from the colder to the hotter, thereby sustaining a temperature gradient (Fig. 2).
Let be the temperature of the cellular environment, which we assume to be in equilibrium with the mitochondrial intermembrane space, and let be the temperature inside the inner membrane. As before, let denote the total metabolic power supplied per unit mitochondrial surface area. We assume the mitochondrion is not fully uncoupled, but instead may use a fraction of this power to “pump” heat inward. Let be the heat flux per unit area actively transferred from the exterior to the interior. The power is expended to generate this inverse heat flow and returns as heat on the interior side. Thus, the total heat flux entering the matrix is
Applying the Second Law of Thermodynamics to this membrane heat pump gives the entropy production per unit area:
With , inequality (11) implies
A key property of a heat pump is that the transferred heat flux can greatly exceed the mechanical or biochemical power consumption . We now analyze this extreme regime. Any heat pumped inward must be removed by ordinary thermal conduction across the inner membrane:
where is the membrane thermal conductivity and its thickness.
Combining Eqs. (12) and (13) yields an upper bound for the cross-membrane temperature difference:
Most parameters in Eq. (14) are known with reasonable accuracy: , membrane thickness . Based on experimental measurements of heat transport in lipid bilayers [30] and typical thermal conductivities of bulk fats [20], we adopt the lower-bound estimate for the inner membrane. Using upper-bound values for the efficiency and metabolic power per area , we obtain
This result significantly narrows the “10^5^ gap,” but still effectively rules out the possibility of a substantial temperature difference, even when nonequilibrium driving is taken into account. The new bound may appear to conflict with the ratchet model predicting multi-degree temperature differences [29]; however, that model assumes nearly perfect thermal isolation between compartments. Once the finite thermal conductance of the membrane is included, its predictions necessarily fall below the bound set by the Second Law.
CONCLUSIONS
III.
In this communication, we addressed the puzzling “10^5^ gap” between the reported mitochondrial “overheating” and the fundamental theoretical estimates based on thermal diffusion. After refining the diffusion-based analysis by incorporating realistic mitochondrial geometry and maximal metabolic power, we turned to a more challenging possibility: that mitochondria, as strongly nonequilibrium biochemical machines, might sustain internal temperature gradients beyond those allowed by passive heat diffusion [28, 29].
To address this, we derived a general thermodynamic bound on mitochondrial temperature gradients from the Second Law of Thermodynamics. Allowing the inner membrane to operate as an ideal, respiration-driven heat pump, we determined the maximal entropy-compatible inward heat flux. When this flux is balanced against unavoidable conductive losses, it yields a strict upper limit: even in the most favorable nonequilibrium regime, the temperature difference across the inner mitochondrial membrane cannot exceed a small fraction of a degree.
Our thermodynamic bound closes the loophole left by earlier diffusion-based analyses. We conclude that no molecular mechanism, regardless of complexity or efficiency, can generate or maintain multi-degree mitochondrial overheating without violating fundamental thermodynamic constraints. Consequently, large apparent “temperature” gradient signals reported by intracellular thermometer sensors are most likely due to biochemical modulation of their fluorescence rather than genuine thermal gradients.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Engelhardt W. A., On the dual role of respiration, Molecular and Cellular Biochemistry 5, 25 (1974).4372523 10.1007/BF 01874169 · doi ↗ · pubmed ↗
- 2Engelhardt W., Ortho-und pyrophosphat im aeroben und anaeroben stoffwechsel der blutzellen, Biochemische Zeitschrift 227 (1930).
- 3Rolfe D. F. and Brown G. C., Cellular energy utilization and molecular origin of standard metabolic rate in mammals, Physiological Reviews 77, 731 (1997).9234964 10.1152/physrev.1997.77.3.731 · doi ↗ · pubmed ↗
- 4Brown G. C., Bioenergetic myths of energy transduction in eukaryotic cells, Frontiers in Molecular Biosciences 11, 1402910 (2024).38952719 10.3389/fmolb.2024.1402910 PMC 11215017 · doi ↗ · pubmed ↗
- 5Chrétien D. , Mitochondria are physiologically maintained at close to 50°C, P Lo S Biology 16, e 2003992 (2018).29370167 10.1371/journal.pbio.2003992 PMC 5784887 · doi ↗ · pubmed ↗
- 6Baffou G., Rigneault H., Marguet D., and Jullien L., Reply to: “validating subcellular thermal changes revealed by fluorescent thermosensors” and “the 10(5) gap issue between calculation and measurement in single-cell thermometry”, Nature Methods 12, 803 (2015).26317235 10.1038/nmeth.3552 · doi ↗ · pubmed ↗
- 7Baffou G., Rigneault H., Marguet D., and Jullien L., A critique of methods for temperature imaging in single cells, Nature Methods 11, 899 (2014).25166869 10.1038/nmeth.3073 · doi ↗ · pubmed ↗
- 8Nielsen J. , Plasticity in mitochondrial cristae density allows metabolic capacity modulation in human skeletal muscle, Journal of Physiology 595, 2839 (2017).27696420 10.1113/JP 273040 PMC 5407961 · doi ↗ · pubmed ↗