Rats exposed to a low resource environment in early life display sex differences in blood pressure, autonomic activity, and brain and kidney pro-inflammatory markers during adulthood
Jonna Smith, Savanna Smith, Kylie Jones, Angie Castillo, Jessica L. Bolton, Ahfiya Howard, Luis Colon-Perez, Faith Femi-Ogunyemi, Allison Burkes, Mark Cunningham

TL;DR
Rats exposed to a low-resource environment early in life show sex-specific differences in blood pressure and inflammation in adulthood, offering insights into how childhood poverty may lead to hypertension in humans.
Contribution
This study reveals sex-specific physiological and inflammatory mechanisms linking early life resource deprivation to adult hypertension in a rodent model.
Findings
LBN males had elevated blood pressure and increased sympathetic nerve activity compared to controls.
LBN females showed no changes in blood pressure but had increased parasympathetic nerve activity.
Males exposed to LBN had reduced brain and kidney pro-inflammatory cytokines, possibly as a compensatory mechanism.
Abstract
Poverty, a low resource state, is a common adverse childhood experience (ACE) and early life stress (ELS). People who experienced childhood poverty are at greater risk for developing hypertension during adulthood, with sex differences. To determine the possible mechanisms of these sex differences, we investigated the alterations in blood pressure (BP), autonomic activity, and inflammation in the brain and kidneys of rats exposed to an impoverished environment during the early life, by using the limited bedding and nesting (LBN) rodent model. The LBN model mimics childhood poverty by creating a low resource environment on postnatal days 2–9. After weaning, offspring were separated by sex and LBN exposure and were evaluated at 16–18 weeks of age (Adulthood). LBN males displayed an increase in BP compared to the control (CON), whereas LBN females showed no changes. Sympathetic nerve…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
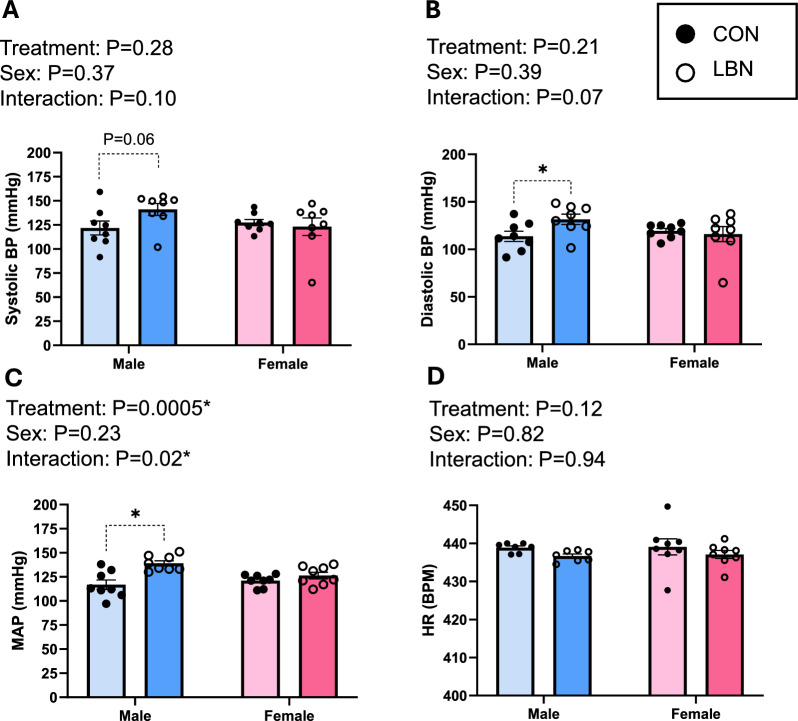 Figure 1
Figure 1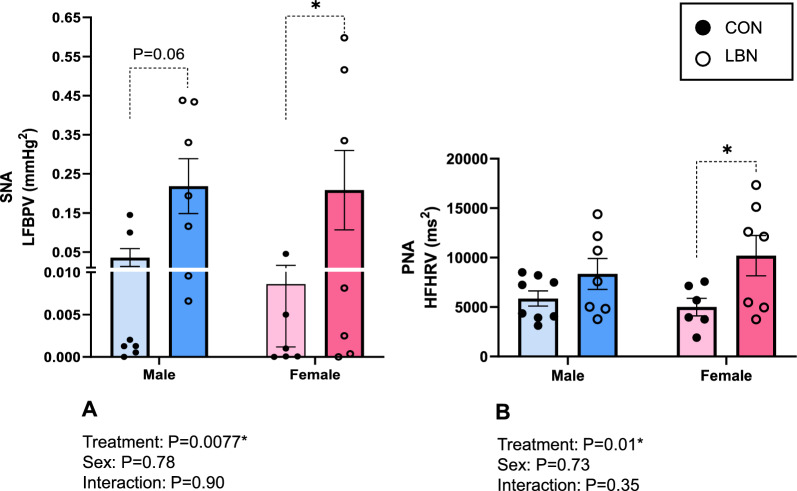 Figure 2
Figure 2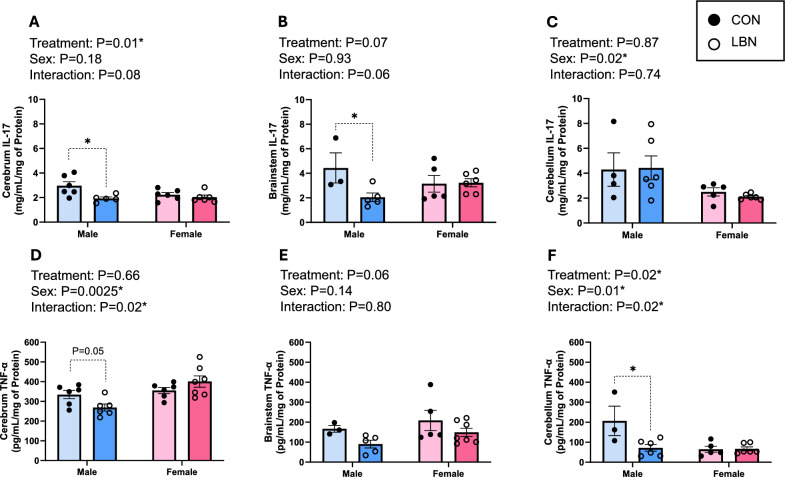 Figure 3
Figure 3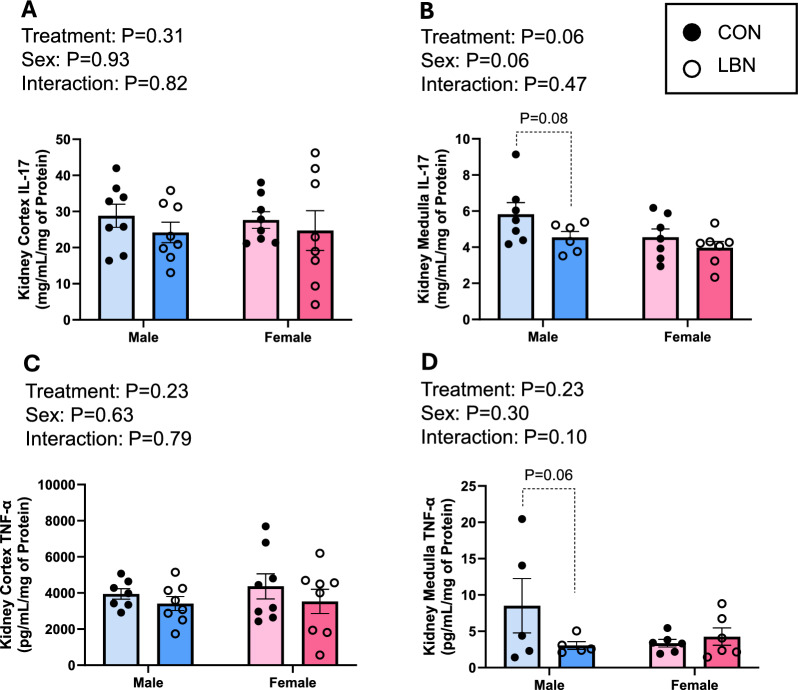 Figure 4
Figure 4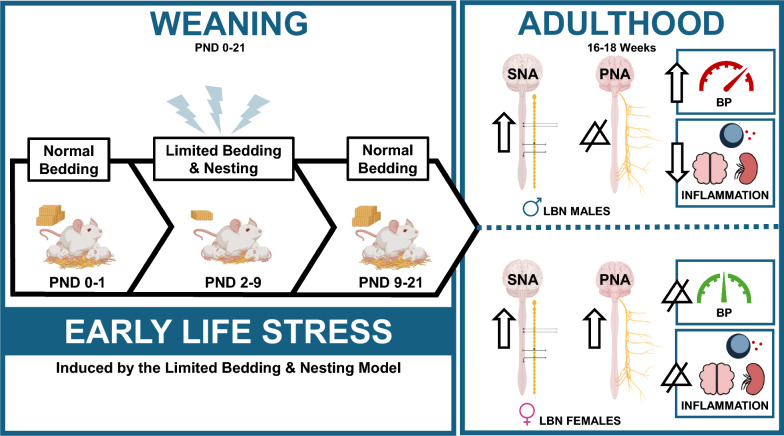 Figure 5
Figure 5- —https://doi.org/10.13039/100014261National Institute for Health Care Management Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsStress Responses and Cortisol · Sodium Intake and Health · Neuroscience of respiration and sleep
Background
Poverty impacts ~ 333 million children worldwide and increases the risk of developing hypertension, cardiovascular, and cerebrovascular diseases later in life [1]. Poverty is defined as resource deprivation that prevents individuals from maintaining a minimum standard of living [2, 3]. Often, the definition of poverty is limited to its characterization of one’s economic status; an individual making an income less than $30,000 a year in the United States (US) is considered below the poverty line [4]. However, poverty is more than just a lack of income. Poverty restricts access to vital resources, such as food, water, shelter, transportation, education, jobs, safety, and healthcare [5].
Poverty is a major public health concern that disproportionally affects children and is considered an adverse childhood experience (ACE). ACEs are traumatic events that occur before the age of 18 years, which can include neglect, abuse, household dysfunction, and importantly poverty [6–9]. According to the Centers for Disease Control (CDC), approximately 64% of the US population has experienced at least one type of ACE, while 17% reported experiencing four or more ACEs. ACEs can negatively impact a person’s mental and physical health throughout life [10–12].
Epidemiological studies of ACEs and animal models of early life stress (ELS) report an increase in cardiovascular and cerebrovascular diseases during adulthood [9, 11, 13–18]. Studies not only show increased health risks during adulthood, but also sex differences in blood pressure (BP) [19–21]. Multiple studies show that males, especially Black males, have a greater risk of developing hypertension, being diagnosed with hypertension at an earlier time point in adulthood, and display a larger increase in BP compared to females that have experienced ACEs [20, 22–24]. However, these findings are controversial. In more recent studies, adult women display a greater increase in hypertension and other cardiovascular diseases (CVDs) when exposed to ACEs, such as physical and sexual abuse [11, 24–29]. In a new study, by Spikes et al*.* 2025, it was revealed that Black women reported more ACEs and had a stronger positive correlation between childhood trauma and impaired arterial compliance, thus making them more susceptible to CVDs later in life [11, 24]. Therefore, based on these later studies, it can be inferred that maybe women, exposed to ACEs, are at a greater risk of CVDs during adulthood [11, 24–29]. However, conversely, some studies suggest no sex differences, while others show that post-menopausal women have equal or greater rates of hypertension compared to males who experienced ACEs [30]. Thus, the notion as to which sex has greater negative health outcomes after being exposed to ACEs is undetermined. These discrepancies in the literature may be attributed to differences in demographic populations, the type and intensity of different ACE exposures, self-reporting in males vs. females, and which cardiovascular output was measured, such as hypertension, stroke, myocardial infarction, and/or cardiac death. From a physiological perspective, these sex differences may also be explained by changes in hormones, neuroendocrine activation, immunological alterations, and autonomic activity [13, 24, 31–33]. Despite the plethora of studies linking ACEs to CVDs during adulthood, the exact mechanisms that predispose this vulnerable population of people to hypertension are unknown and the focus of this study.
Several factors are known to alter BP in humans and rodents exposed to ELS. In this study, we chose to focus on alterations in autonomic function and inflammation. Autonomic dysfunction can result in increased sympathetic nerve activity (SNA), in turn, which will increase systemic and local inflammation to facilitate the development of hypertension [17, 26, 34, 35]. Circulating and local tissue inflammation can also play a role in elevating BP independent of increased SNA [17, 34, 36, 37]. Specifically, an increase in brain and kidney pro-inflammatory cytokines, such as interleukin-17 (IL-17) and tumor necrosis factor-alpha (TNF-α), are shown to increase BP in rodents with or without ELS, with sex differences [37–40]. IL-17 and TNF-α are known to regulate BP, via an increase in systemic and/or tissue inflammation. Elevated renal IL-17 and TNF-α concentrations can alter sodium transporters, increase the production of renin–angiotensin–aldosterone system components, change renal hemodynamics, and cause renal dysfunction to increase BP [36, 41]. These same cytokines in the brain can contribute to cerebrovascular dysfunction and increased blood–brain-barrier (BBB) permeability. An increase in BBB permeability permits the influx of these cytokines into the brain, leading to further damage, inflammation, and autonomic activation, all of which will increase BP [42]. Additionally, IL-17 and TNF-α can elicit the production of other cytokines, thus generating a pro-inflammatory cytokine storm [43, 44]. Studies from our lab and others have implicated that these cytokines participate in the pathology and pathogenesis of hypertension, ischemic strokes, preeclampsia, hypertension in postpartum preeclamptic women, and importantly, in human and animals exposed to ELS/ACEs [25, 38, 45–48]. Examining the modifications in autonomic activation and renal/brain inflammation may provide clues into the sex differences associated with CVD risk in people exposed to ACEs.
To study ELS, in the context of a low resource environment, we utilized the limited bedding and nesting (LBN) rodent model, to assess the BP, autonomic activity, and alterations in inflammation within different regions of the brain and kidney. The LBN model mimics childhood poverty by reducing bedding and nesting material during weaning [45, 49–51]. This model creates a chronically stressed environment, characterized by unpredictable maternal care [45, 49–52]. These fragmented behaviors observed in dams are also observed in people living in impoverished environments [53]. In rodents, the radical changes in maternal care, such as alterations in duration and frequency of grooming, nest building, and rough handling, can mirror neglect and abuse experienced by children during poverty [45, 53]. This ELS model has demonstrated that LBN rodents have alterations in brain structures (such as the hippocampus, amygdala, and prefrontal cortex), hypothalamic–pituitary–adrenal (HPA) axis, cognitive decline, and depression [17, 49, 51]. Despite these novel findings, no studies have investigated sex differences and effect of LBN exposure on BP, autonomic activity, and inflammation in rodents during adulthood. These studies are necessary because ACEs are common in adults, and the physiological mechanisms connecting ACEs and increased risk of CVDs and cerebrovascular dysfunction are unknown [17, 54].
The objective of this study is to characterize changes in BP, autonomic activity, and inflammation in the brain and kidney of male and female rats that experienced a low resource (LBN) environment during weaning. We hypothesize that LBN male rats will display elevated BP, changes in autonomic activity, and inflammation in the brain and kidney, whereas LBN females will display no changes in these measurements. The results from this study will provide insights into the sexually dimorphic mechanisms that link childhood poverty to the development of hypertension during adulthood.
Methods
Experimental procedure
Timed-pregnant Sprague–Dawley rats (ENVIGO; Indianapolis, IN) were received on gestational day (GD) 11–12 and were divided into two groups: Control (CON; n = 6) and LBN (n = 7) dams. The rats were housed in a 12-h light/dark cycle with controlled temperature and humidity. Food and water were provided ad libitum throughout the entire experimental protocol. All animals and procedures used in this study were approved by the University of North Texas Health Fort Worth Institutional Animal Care and Use Committee (IACUC) and in accordance with the National Institute of Health (NIH) Guide for the Care and Use of Laboratory Animals.
At GD 20–22, both CON and LBN dams gave birth naturally. From postnatal day (PND) 1–21, the CON dams weaned their pups in a normal environment with the typical amounts of bedding and nesting material. From PND 2–9, LBN dams and pups were relocated to a modified LBN environment, with 75–80% less bedding material, to induce ELS [45, 51, 52]. The LBN environment included a slightly elevated mesh floor on the bottom of a clean cage to allow for droppings to fall through and to provide another layer of stress. During PND 2–6, dam behaviors were recorded, analyzed, and evaluated to calculate entropy, which is a measurement of fragmented behaviors for the dams [49, 55]. The dams were monitored for frequency, total duration, and the mean duration of self-grooming, pup grooming, nursing, nest building, eating and drinking, and transportation of pups in the cage. The measurements were then used to generate an entropy score for our dams, using a computational algorithm [49, 55]. The entropy score utilizes the conditional probability of each behavior, in sequence with other behavior types, to determine if the dams’ behavior was fragmented and unpredictable. Many other research groups have used the entropy score to validate the efficacy of the LBN model in generating ELS [45, 49, 52]. The wire mesh was removed, and normal quantities of bedding and nesting materials were provided to the LBN group on PND 10, which continued until PND 21. After the weaning period, all pups were separated by sex and treatment, then aged to 16–18 weeks of age, which is equivalent to young adulthood in humans [56].
To measure the mean arterial pressure (MAP), heart rate (HR), sympathetic nerve activity (SNA), and parasympathetic nerve activity (PNA), carotid catheterization surgery was performed at 16–18 weeks of age. After the BP, HR, SNA, and PNA were recorded, we humanely euthanized the animals to collect blood and organs. The brain and kidney samples were snap-frozen and stored in a −80 °C freezer. Later, the brains were sectioned into cerebrum, brainstem, and cerebellum, whereas the kidneys were separated into medulla and cortex for experimentation and analysis. The organ sections were then homogenized and centrifuged at 10,000G for 20 min to obtain supernatants for future use in colorimetric enzyme-linked immunosorbent assays (ELISAs), namely IL-17 and TNF-α.
Carotid catheterization surgery
At 16–18 weeks of age, we performed carotid catheterization surgeries on all offspring, as described previously by us and others [45, 47, 48]. Briefly, to perform the carotid catheterization surgery, we made an incision into the ventral neck under 2–4% nasal isoflurane, isolated the carotid artery, then implanted a small catheter. We secured the catheter to the artery with surgical thread and weaved it through the dorsal neck. While under anesthesia, core temperature was maintained via a heating pad. This surgery lasted ~ 15 to 20 min per rat. Post-surgery, the rat was placed in a warm, clean cage with a 2 mg tablet of bacon-flavored Rimadyl, to monitor for safe recovery. BP and autonomic activity were recorded the next day.
BP, SNA, and PNA recordings
Systolic BP, diastolic BP, MAP, heart rate (HR), low frequency BP variability (LFBPV) for SNA, and high frequency HR variability (HFHRV) for PNA were recorded via a PowerLab 16/35 AD Instruments apparatus (ADInstruments, Australia) as performed and described previously [57]. To measure SNA, we examined the variability in MAP tracings at a low frequency output (0.2–0.8 Hz) using power spectral analysis calculations [58, 59]. To obtain PNA, we also used the power spectral analysis to analyze HR variability at the high frequency output (0.75–2.0 Hz) [58]. The power spectral analysis values were calculated using the Fast Fourier Transform (FFT) algorithm (2,048 values, 50% overlapping segments) [58].
Inflammation assays
To determine IL-17 and TNF-α concentrations in the brain and kidney, we performed the following ELISAs: IL-17 (DY8410) and TNF-α (DY510-05; R&D Systems, Minneapolis, MN) according to the manufacturer’s instructions and previous studies [47]. Before loading the samples into the 96-well plates, we diluted the kidney cortices to 1:10, while the kidney medullas and brain sections were pipetted without dilution. Both assays were read at 450 nm using the BioTek Epoch 2 microplate reader (Agilent Technologies, Santa Clara, CA) alongside Generation 5 software (Santa Clara, CA).
Statistics
To analyze dam behavior and entropy score, we utilized the Student’s t-test. Body weight, organ weight, BP, inflammation, and autonomic activity were analyzed by a two-way ANOVA with a Fisher’s least significant difference (LSD) post-hoc, using GraphPad Prism 10 (v. 10.0.3; Santa Clara, CA) and Excel (Microsoft). These data were reported as mean ± standard error of the mean (SEM), where statistical significance was defined as *P < 0.05.
Results
LBN Induced behavioral changes and increased entropy score in dams
Dams exposed to the LBN model during PND 2–9 exhibited several behavioral modifications in frequency of events, total duration of events, and average time of each event during exposure to the LBN (Table 1). We observed increased frequency of licking and grooming of pups (10.2 ± 1.16 vs. 5.5 ± 1.07A.U.; P = 0.03), self-grooming (7.74 ± 0.68 vs. 2.75 ± 0.64A.U.; P = 0.0009), nest building (3.86 ± 0.84 vs. 1.9 ± 0.33A.U.; P = 0.13), eating (4.23 ± 0.83 vs. 1.65 ± 0.60A.U.; P = 0.06), and off-nest (11.23 ± 1.14 vs. 5.5 ± 0.95 A.U.; P = 0.008). Furthermore, the total duration of licking and grooming the pups (441.65 ± 52.94 vs. 283.80 ± 40.89 s; P = 0.07), self-grooming (258.83 ± 22.61 vs. 98.69 ± 30.34 s; P = 0.002), and eating (373.80 ± 80.49 vs. 137.73 ± 32.42 s; P = 0.06) also increased. However, there were no significant changes in total duration of nest building (144.11 ± 42.81 vs. 114.37 ± 29.20 s; ns) and off-nest time (571.33 ± 150.59 vs. 405.15 ± 178.83 s; ns). There was a significant decrease in total duration of nursing time (1,803.63 ± 150.85 vs. 2554.70 ± 166.52 s; P = 0.01) with LBN dams, but no change in nursing event frequency (9.89 ± 1.57 vs. 7.65 ± 0.82A.U.; ns). Overall, no significant changes in the average time of each behavior were observed during this period (Table 1). To validate the exposure of chronic stress on the dams and pups, an entropy score was calculated. The LBN dams showed increased entropy (1.16 ± 0.05 vs. 0.83 ± 0.02A.U.; P = 0.001) compared to the CON. Other groups have also demonstrated a higher entropy score in LBN dams, reflecting a chronically stressed environment with fragmented maternal behaviors (Table 1) [45, 51, 52].Table 1. Behavioral Changes of Postpartum Dams (PND 2–6) Exposed to LBNMeasuresNumber of eventsTotal duration (s)Mean duration of each event (s)Behaviors****CON (Mean ± SEM)****LBN (Mean ± SEM)P-ValueCON (Mean ± SEM)****LBN (Mean ± SEM)P-ValueCON (Mean ± SEM)****LBN (Mean ± SEM)P-ValueLicking & Grooming (LG)5.5 ± 1.0710.2 ± 1.160.03283.80 ± 40.89441.65 ± 52.920.0764.15 ± 14.6347.76 ± 2.500.17Self-Grooming (SG)2.75 ± 0.647.74 ± 0.680.000998.69 ± 30.34258.83 ± 22.610.00222.23 ± 2.7132.15 ± 3.780.1Nursing (N)7.65 ± 0.829.89 ± 1.570.342554.70 ± 166.521803.63 ± 150.850.01485.69 ± 80.81317.28 ± 70.710.17Nest-Building (NB)1.9 ± 0.333.86 ± 0.840.13114.37 ± 29.20144.11 ± 42.810.6431.43 ± 6.9229.65 ± 6.070.86Eating (E)1.65 ± 0.604.23 ± 0.830.06137.73 ± 32.42373.80 ± 80.490.0653.74 ± 11.9567.39 ± 11.120.45Carrying Pups (C)0.3 ± 0.240.69 ± 0.160.194.91 ± 4.546.33 ± 1.870.742.64 ± 2.283.84 ± 1.070.6Off-Nest (O) = O + M5.5 ± 0.9511.23 ± 1.14ss0.008405.15 ± 178.83571.33 ± 150.590.51128.31 ± 48.9297.53 ± 34.140.61CON (Mean ± SEM)****LBN (Mean ± SEM)P-ValueEntropy0.83 ± 0.021.16 ± 0.050.001All data for the behavior observations were analyzed with two-way ANOVA and the Fisher’s LSD test for post-hoc test for comparisons. Entropy analysis was compared by using student t-test. Comparisons with P < 0.05 are bolded. Comparisons with 0.050 < P < 0.100 are underlined. Both significant and marginal comparisons are posted
Body weight and organ weight analysis
Body weight, brain weight, total kidney weight, and heart weight of adult offspring showed no differences between treatment in male or female LBN vs. CON rats (Table 2). However, there was a sex effect, with males having a larger body weight and smaller brain-to-body weight ratio than females in both LBN and CON rats (Table 2).Table 2. Measurement of Body Weight, and Organ WeightsCON MLBN MCON FLBN FTreatment effect(P-Value)Sex effect(P-Value)Interaction(P-Value)(AVG ± SEM)(AVG ± SEM)(AVG ± SEM)(AVG ± SEM)Body Weight (g)446.13 ± 13.28444.43 ± 12.01256.00 ± 6.71259.25 ± 5.92 > 0.9999 < 0.00010.7369Brain Weight (g/kg of BW)4.45 ± 0.124.44 ± 0.077.18 ± 0.176.90 ± 0.180.2933 < 0.00010.3902Total Kidney Weight (g/kg of BW)5.64 ± 0.095.73 ± 0.145.81 ± 0.145.66 ± 0.190.87850.76960.3981Heart Weight (g/kg of BW)3.00 ± 0.093.21 ± 0.133.24 ± 0.193.25 ± 0.230.64080.21000.6785All measurements at 16–18 weeks (adulthood) were analyzed with two-way ANOVA and the Fisher’s LSD test for post-hoc test for comparisons. Comparisons with P < 0.05 are bolded
LBN & sex: effects on BP, HR, and autonomic activity
Systolic BP, diastolic BP, MAP, HR, and autonomic activity (SNA and PNA) were measured in adult CON and LBN rats (Figs. 1A-D and 2A-B). Both systolic (Fig. 1A) and diastolic BP (Fig. 1B) did not show an overall LBN exposure or sex effect. However, sex and LBN status did show a tendency for an interaction (Systolic BP: P = 0.10; Diastolic BP: P = 0.07) effect (Figs. 1A-B). When checking for multiple comparisons, LBN males had elevated systolic (141 ± 6 vs. 122 ± 7 mmHg, P = 0.06) (Fig. 1A) and diastolic (132 ± 5 vs. 114 ± 5 mmHg, P = 0.03) (Fig. 1B) BP compared to CON males, whereas females no differences. Moreover, MAP showed both a treatment and interaction effect, in which LBN exposure had an effect in males but no effect in females (Fig. 1C). In fact, LBN males exhibited a 16% (~ 23 mmHg) increase in MAP (139 ± 3 vs. 117 ± 5 mmHg; P = 0.0001) compared to the CON males (Fig. 1C). There were no changes in MAP between LBN and CON females. There were no differences in HR among the groups (Fig. 1D).Fig. 1A) Systolic BP, B) diastolic BP, C) MAP, and D) HR were measured in LBN males (n = 7–8), CON males (n = 7–8), LBN females (n = 8), and CON females (n = 8). All bar graphs display individual data points with each group’s mean ± SEM, with significance determined at P < 0.05. Groups were statistically analyzed with two-way ANOVA and the Fisher’s LSD test for post-hoc analysis. Individual data points are represented by closed circles for CON offspring and open circles for LBN offspring. Male bar graphs are blue and female bar graphs are pink
To determine autonomic activity, we measured LFBPV (SNA) and HFHRV (PNA). There was a significant increase in SNA with LBN exposure in both males and females (P = 0.0077) (Fig. 2A). LBN males had a sixfold increase in SNA (0.218 ± 0.07 vs. 0.04 ± 0.02mmHg^2^, P = 0.06) and LBN females displayed a 23-fold increase in SNA (0.209 ± 0.10 vs. 0.009 ± 0.01mmHg^2^, P = 0.05) compared to their respective CON groups (Fig. 2A). Conversely, PNA was only increased in LBN females vs. CON females (10,199.71 ± 2,047.22 vs. 5,004.83 ± 892.29ms^2^; P = 0.01) (Fig. 2B). No changes were observed in PNA between LBN males and CON males (ns).Fig. 2A) Low Frequency BP Variability (LFBPV; SNA) and B) High Frequency Heart Rate Variability (HFHRV; PNA) were measured in LBN males (n = 7), CON males (n = 7–8), LBN females (n = 7), and CON females (n = 6). All bar graphs display individual data points with each group’s mean ± SEM, with significance determined at P < 0.05. Groups were compared with two-way ANOVA and the Fisher’s LSD test for post-hoc analysis. Individual data points are represented by closed circles for CON offspring and open circles for LBN offspring. Male bar graphs are blue and female bar graphs are pink
LBN & sex: effects on brain inflammation
IL-17 and TNF-α were examined in different segments of the brain: cerebrum, brainstem, and cerebellum outlined in figs. 3A-F. There was a trend in the interaction effect (sex X LBN), observed in the cerebrum (P = 0.08) and brainstem (P = 0.06) for IL-17. There were no significant changes in cerebral IL-17 in females (Fig. 3A). Although in figs. 3A-B, LBN males exhibited a decrease in IL-17 concentrations in the cerebrum (1.93 ± 0.13 vs. 2.96 ± 0.34 mg/mL/mg of Protein, P = 0.01) and brainstem (2.04 ± 0.35 vs. 4.43 ± 4.43 ± 1.23 mg/mL/mg of Protein; P = 0.02). There was no difference in cerebellar IL-17 between LBN vs. CON rats within each sex, but there was an overall sex difference (P = 0.02) with females having less IL-17 compared to males (Fig. 3C).Fig. 3A) Cerebrum, B) brainstem, and C) cerebellum IL-17 concentrations were measured in LBN males (n = 5–6), CON males (n = 3–6), LBN females (n = 5–6), and CON females (n = 6). D) Cerebrum, E) brainstem, and F) cerebellum TNF-α concentrations were measured in LBN males (n = 5–6), CON males (n = 3–6), LBN females (n = 6–7), and CON females (n = 5–6). All bar graphs display individual data points with each group’s mean ± SEM, with significance determined at P < 0.05. Groups were compared with two-way ANOVA and the Fisher’s LSD test for post-hoc analysis. Individual data points are represented by closed circles for CON offspring and open circles for LBN offspring. Male bar graphs are blue and female bar graphs are pink
Cerebral TNF-α concentration was decreased for LBN vs. CON males (268.81 ± 17.99 vs. 334.28 ± 20.91 pg/mL/mg of Protein; P = 0.05), with no differences in LBN vs. CON females (Fig. 3D). No differences were observed with brainstem TNF-α (Fig. 3E). However, cerebellum TNF-α showed a fourfold decrease between LBN vs. CON males (71.90 ± 16.43 vs. 206.79 ± 73.65 pg/mL/mg of Protein; P = 0.003), with no changes presented in females (Fig. 3F).
LBN & sex: effects on kidney inflammation
Concentrations of IL-17 and TNF-α were also investigated within the kidney cortex (Figs. 4A & C) and medulla (Figs. 4B & D). IL-17 within the kidney cortex revealed no effects with LBN exposure and/or sex (Fig. 4A). However, IL-17 in the kidney medulla tended to decrease in LBN vs. CON males (4.55 ± 0.33 vs. 5.83 ± 0.64 mg/mL/mg of Protein; P = 0.08) (Fig. 4B). No differences exist between sex and LBN exposure vs. CON females for kidney medulla IL-17.Fig. 4A) Kidney cortex and B) kidney medulla IL-17 concentrations were measured in LBN males (n = 6–8), CON males (n = 7–8), LBN females (n = 7–8), and CON females (n = 7–8). C) Kidney cortex and D) medulla TNF- α concentrations were also measured in LBN males (n = 5–8), CON males (n = 5–7), LBN females (n = 6–8), and CON females (n = 6–8). All bar graphs display individual data points with each group’s mean ± SEM, with significance determined at P < 0.05. Groups were compared with two-way ANOVA and the Fisher’s LSD test for post-hoc analysis. Individual data points are represented by closed circles for CON offspring and open circles for LBN offspring. Male bar graphs are blue and female bar graphs are pink
In the kidney cortex, there was no differences in LBN exposure and/or sex between groups for TNF-α (Fig. 4C). Although, kidney medulla TNF-α tended to decrease in LBN vs. CON males (3.04 ± 0.53 vs. 8.52 ± 3.74 pg/mL/mg of Protein; P = 0.06) (Fig. 4D).
Discussion
LBN exposure induces sex differences in BP, autonomic activity, and inflammation in the brain and kidney (Fig. 5). Specifically, LBN males exhibit an increase in BP, increased SNA, and decreased pro-inflammatory markers in the cerebrum, brainstem, and cerebellum. LBN females depicted no differences in BP, an increase in both SNA and PNA, and no alterations in brain or kidney inflammation. Results from our study suggests that the elevation in BP for LBN males may be due to an increase in SNA. However, LBN females may be protected against BP elevation due to a simultaneous increase in PNA, despite increased SNA. The sex differences in inflammation may be a compensatory response to BP, in which LBN males show a reduction pro-inflammatory cytokines to mitigate brain damage from an increase in BP. Meanwhile, LBN females do not show changes in inflammation, since BP is not different between groups. It is important to note that these sexually dimorphic differences in BP, autonomic activity, and brain/kidney inflammation may also be explained by differences in sex hormones, which was not examined in this study [11, 19–21, 25, 29].Fig. 5**)** A Graphic design summarizing the major findings of this study
Children exposed to ACEs, including poverty, have an increased risk of poor health outcomes such as a heightened risk of hypertension, inflammation, and stroke as adults [9, 11, 13–18, 20, 22, 23, 25, 27–30, 60]. Several clinical studies show that ACEs are linked to sex differences in the presentation of hypertension [8, 11, 14, 20, 24, 25, 27–30, 32, 61–63]. For example, in Huang et al*.’s* study, they found that persons experiencing ACEs increased the risk of hypertension later in life, indirectly, through cardiometabolic dysregulations (such as hyperlipidemia and hyperglycemia), systemic inflammation, and obesity [61]. In a similar study, conducted in the United Kingdom, Deschenes et al*.* found that British civil service employees, who reported ACEs had a higher risk for developing heart coronary disease, partially mediated through cardiometabolic dysregulation and hypertension [32]. Additionally, Su et al*.’s* 2015 longitudinal study demonstrated a strong, positive relationship between the number of ACES and BP; participants who were exposed to more ACEs exhibited a larger increase in BP during early adulthood compared to those who did not experience any ACEs [8].
The association between elevated BP and ACEs is not only observed in clinical studies, but also in animal models of ELS, using maternal separation and/or early weaning models [64–66]. Franco et al*.’s* 2013 study revealed that 25-week-old adult Wistar male rats exposed to early weaning were hypertensive with extensive oxidative stress [64]. Reho and Fisher 2015 investigated changes in vascular function and BP in a maternal separation rodent model. In these studies, they found that maternal separation caused an increase in arterial contractility at PND 21, but no changes in BP. However, at PND 35, the relationship was reversed, in which BP was increased while arterial contractility was unchanged [65]. Genest et al*.* 2004 showed that neonatal maternal separation led to a stark 20% increase in MAP in males, but no changes in females at 8–10 weeks of age [66]. Note that these results are like our findings, in which LBN males, during adulthood, had significantly increased MAP, while females remained unchanged. Together, these data show that ELS, whether it be via maternal separation or our model of LBN, is linked to hypertension.
The LBN model reduces resources to induce chronic stress in dams and pups during weaning. Typically, the LBN model is used to evaluate outcomes of depression and cognitive dysfunction along with changes in brain morphology [49, 52, 67, 68]. However, our study utilized this model to investigate the mechanisms that link living in a low resource environment during childhood to changes in autonomic activity, BP regulation, and organ inflammation. To validate the LBN model, we recorded and analyzed the dams’ behavior during LBN exposure. We found changes in licking and grooming, self-grooming, nursing, and time off-nest, which correlated with changes that others have observed with this model [49, 69]. Moreover, we calculated an entropy score, which measures maternal behavior fragmentation, and showed that our score was higher than the CONs, consistent with findings from others who use this model have found [49] (Table 1).
We observed sex differences in adult BP, with LBN males showing an increase, and LBN females displaying no changes. Many potential mechanisms could facilitate these differences in BP between males and females, which include alterations in autonomic activity, inflammation, oxidative stress, vascular dysfunction, HPA axis dysfunction, endothelin-1, and decreased NO bioavailability. However, in this study, we chose to focus on autonomic activity and inflammation in the kidney and brain using the LBN model of ELS, due to their crucial role in BP regulation.
Changes in autonomic activity, both sympathetic and parasympathetic, are known to influence BP [70, 71]. Whereas increased SNA and/or decreased PNA will elevate BP [72]. In our study, LBN males had an increase in SNA, while LBN females showed an increase in both SNA and PNA. An increase in SNA is demonstrated in both human and animal studies when the subjects are exposed to an ACE or ELS [17, 33, 66, 73]. For example, a study investigating the association between childhood trauma and catecholamine response to psychological stressors found that 3-methoxy-4-hydroxy-phenylglycol (MHPG), a metabolite of norepinephrine, was elevated in participants who experienced ACEs [74–76]. An increase in norepinephrine concentrations in most studies suggests an increase in SNA, which is what we observed in our LBN rodents. A study by Renard et al*.* 2005 similarly found sex differences in SNA in mice exposed to maternal separation. In this study, maternally deprived females exhibited slightly higher basal plasma norepinephrine levels compared to CON females. However, plasma norepinephrine was unchanged in maternally deprived males compared to the CON [76]. Another study by González-Pardo et al*.* in 2020 further suggested sex differences in SNA, showing that norepinephrine turnover in maternally separated male mice was decreased compared to the CON, while maternally separated female mice was increased compared to CON [73]. Note that an increase in norepinephrine turnover also suggests an increase in SNA. Although these previous studies suggest some sex differences in autonomic activity with ELS exposure, our study showed that both LBN males (hypertensive) and LBN females (normotensive) had increased SNA, with females having a greater increase in SNA, which appears to be supported by clinical data. However, despite the increase in SNA in our study, LBN females also demonstrated an increase in PNA. PNA can influence BP and often works in antagonism to offset an increase in SNA [75, 76]. Based on our observations, we predict that perhaps LBN male rats had increased BP due to an increase in SNA, whereas LBN female rats were protected against BP elevation due to an increase in PNA to balance out the increase SNA.
Human and animal subjects exposed to ACEs and/or ELS are associated with an increase in systemic and targeted organ inflammation throughout life [13, 73, 77]. An increase in inflammation is often characterized by an increase in pro-inflammatory cytokines, decrease in anti-inflammatory cytokines, and change in both types of cytokines to favor an inflammatory state [78]. The pro-inflammatory cytokines that have been augmented in human and animal models are TNF-α, IL-17, IL-6, and C-reactive protein [77, 79]. However, these changes in pro-inflammatory cytokines are controversial and can differ depending on the species, age, and ELS events, intensity, and duration.
Two important pro-inflammatory cytokines that we investigated in the brains and kidneys are IL-17 and TNF-α, which both directly and indirectly influence BP regulation and can cause hypertension [38, 80–83]. In this study, we hypothesized that both IL-17 and TNF-α would be increased in the brain and kidneys of hypertensive LBN males and not changed in normotensive LBN females. While our hypothesis was accurate with LBN females, the LBN males displayed the opposite of what we predicted. LBN males showed a reduction in pro-inflammatory cytokines (IL-17 and TNF-α) despite having elevated BP and increased SNA. Therefore, the decrease in brain and kidney pro-inflammatory cytokines may be an initial compensatory mechanism acting in response to elevated BP and/or SNA. This compensatory relationship observed in this study is not uncommon. Others have observed these same changes in pro-inflammatory markers (e.g., TNF-α and IL-1β) due to increased SNA or norepinephrine concentrations [84, 85]. Thus, the changes in organ inflammation in LBN rodents may be an attempt to lower and/or maintain normal BP and to prevent tissue damage.
Our study suggests that the reduction of brain/kidney inflammation may serve as a compensatory mechanism, due to an increase in SNA and BP in LBN males. The literature supports these findings; and suggests that the relationship between BP and inflammation as well as SNA and inflammation can be complex. Whereas, most studies show a positive correlation between BP and inflammation, especially in hypertension, where there is typically increased inflammation [86]. However, some studies have demonstrated a U-shaped relationship with BP and inflammation, where extended states of untreated and chronic hypertension are associated with decreases in inflammation [87].
The relationship between SNA and inflammation is also nuanced, as the site of inflammation, the pathological state, the concentration of neurotransmitters, and the type of receptors on immune cells need to be considered [35, 88]. Moreover, increases in SNA causes the release of norepinephrine which binds to beta 2 adrenergic receptors on B cells, macrophages, mast cells, and NK cells to reduce inflammation [88]. Thus, the ratio of α to β adrenergic receptors on immune cells can determine the inflammatory response to norepinephrine. In fact, α adrenergic receptors are preferred to be activated at low physiological concentrations of norepinephrine, while β adrenergic receptors are favored at high physiological concentrations of norepinephrine. When β adrenergic receptors are targeted, macrophages can shift from a neutral or M1 phenotype to the M2 phenotype, which will increase anti-inflammatory cytokines such as IL-10, IL-4, and IL-13. Furthermore, when β adrenergic receptors are activated on immune cells, there will be a decrease in the production of pro-inflammatory cytokines, IL-6 and TNF-α [88–93]. Note that in our study, TNF-α decreased in the brain and kidneys of LBN males, paired with increased SNA. Therefore, we predict that both BP elevation and increased SNA caused a counteractant decrease in brain and kidney inflammation in LBN males. However, because LBN females displayed an increase in PNA to neutralize the simultaneous increase in SNA, these rats did not experience a rise in BP nor changes in inflammation. Future experiments are warranted to defend this hypothesis.
Limitations and future directions
It is important to note that this study is observational and not causal. Although the findings in this study are novel, we cannot reveal the exact timing, order of sequence of physiological events, and/or how each physiological change influences the other changes, i.e., how changes in SNA or inflammation can alter BP. To address these issues, longitudinal studies (before puberty, after puberty, and early adulthood) and gain of function and/or inhibition studies with the autonomic nervous system and/or inflammation need to be performed. Other pro-inflammatory (IL-6, IL-1β and C-reactive protein) and anti-inflammatory (IL-4 and IL-10) cytokines should be explored in specific brain regions (such as the rostral ventrolateral medulla, paraventricular nucleus, and sections of the hippocampus), known to change BP. Additionally, it would be helpful to confirm these trends in inflammatory alterations via western blot, immunohistochemistry, and flow cytometry in brain and kidney, which help identify the specific location and cells within the organs that demonstrate these changes.
Although in this study we measured SNA and PNA using LFBPV and HFHRV, which is widely used, direct measurements of autonomic activity via microneurography are necessary to validate our findings [58, 59, 94]. The limitations of using LFBPV and HFHRV are dependent on external factors that may influence the rodents’ measurements of BP and HR. These factors consist of, and are not limited to, the use and type of anesthesia, respiratory rate, temperature, CO_2_ and oxygen content, along with the stress levels of the rodents during recordings. In this study, we attempted to regulate these variables by making sure that all measurements were taken in optimal conditions in live rats, with no anesthesia and reduced stress.
In addition, this study did not measure the changes in sex hormones in response to BP, autonomic activity, and brain and kidney inflammation in LBN rats. However, observing sex hormones, such as estrogen, progesterone, and testosterone, and other androgens, in both males and females, may shed light on the sex differences observed in our study.
Sex hormones are known to modulate BP, oxidative stress, autonomic activity, and inflammation [31, 95–97]. Several studies show that estrogen and/or estrogen receptor activation, in premenopausal women, reduces the production of reactive oxygen species, lowers inflammation, increases antioxidant capacity, and enhances NO production [31, 34, 98, 99]. Estrogen administered to male rats with ischemic reperfusion injury displayed a reduction in infarct size and cardiac injury, suggesting that estrogen has cardioprotective effects in males too [99]. These studies indicate that estrogen is protective against the development of CVDs in both males and females.
Estrogen can also affect HPA axis activity, depending on the estrogen concentration as well as the type and location of estrogen receptor activation. Studies show that elevated concentrations of estrogen reduce HPA activity via estrogen receptor beta (ER-β) activation [100]. Conversely, lower concentrations of estrogen and/or activation of estrogen receptor alpha (ER-α), in the paraventricular nucleus, enhances HPA activity by impairing the HPA negative feedback loop to increase corticotropin releasing hormone secretion, thereby elevating cortisol levels [100, 101]. Therefore, the ratio of ER-α to ER-β receptors may be important to determining the role of estrogen to activate or inhibit HPA axis activity [102]. Likewise, the abundance and activation of specific estrogen receptors may play a crucial role in cardioprotective effects of both sexes.
Estrogen also assists in increasing parasympathetic tone and decreasing sympathetic tone in females, which is attenuated after menopause [34, 96, 103]. Estrogen has similar effects in males [104, 105]. Studies have shown that both male and female rats given estrogen will decrease sympathetic tone and increase parasympathetic tone [104, 105]. On the other hand, testosterone increases SNA, via a reduction in NO bioavailability and increases in inflammation in both males and females [31, 106–108]. Although removal of endogenous testosterone in males, will increase the risk of CVDs, such as hypertension [109–111]. This same trend is also observed in aging men, with decreasing testosterone production. In females, removing testosterone appears to have no significant effects on BP and the risk of CVDs [112]. Furthermore, both estrogen and testosterone can activate or downregulate the amount of α or β adrenergic receptors to promote vasoconstriction (via ER-α activation) or NO mediated vasodilation (via ER-β activation) [113, 114]. Thus, the activation of specific hormones and/or hormone receptors may help to explain sex differences observed in this study. Future studies will address the role of sex hormones in the pathogenesis of hypertension in the context of ELS in adult offspring.
Perspectives and significance
In summary, our data display sex differences, in which adult LBN males have elevated BP, possibly due to increased SNA. On the other hand, our data suggest that LBN females may be protected from increased BP due to a simultaneous increase in PNS and SNA. Furthermore, we detected no changes in pro-inflammatory cytokines in the brain and kidneys of LBN females but found a decrease in LBN males. The reduction in pro-inflammatory cytokines, IL-17 and TNF-α, in LBN males may serve as a compensatory mechanism to lower BP and/or prevent tissue damage. Identifying the sexually dimorphic alterations in the pathology of adults exposed to an ELS may help to derive novel treatments for patients who have experienced ACEs.
Societal implications and conclusions
Understanding the mechanism of hypertension in animals that experience ELS will help scientists and physicians better understand the pathology and pathogenesis of hypertension in individuals who have experienced poverty as an ACE. Poverty is a pervasive and prevalent issue that continues to impact the health and well-being of many individuals and families worldwide. Unfortunately, with the lag in wage growth and increase in income inequality, along with a rise in inflation, food costs, high unemployment, and lack of affordable housing, it is predicted that childhood poverty rates will increase [115]. To change the negative trajectory of poor health outcomes in people that have experienced ACEs, we advocate that 1) scientists conduct more experiments to determine the impact of resource deprivation (i.e., poverty) on the development of hypertension with sex differences, and 2) the mechanisms that link ELS/ACEs to hypertension development with sex differences.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Spikes TA, Thorpe RJ Jr., Michopoulos V, et al. Effect of early life trauma exposure on vascular dysfunction in Black men and women. J Am Heart Assoc. 2025;14(5):e 036498. 10.1161/JAHA.124.036498.10.1161/JAHA.124.036498 PMC 1213268639996445 · doi ↗ · pubmed ↗
- 2Vegetabile Brian G. S-OSA, Davis Elysia Poggi, Baram Tallie Z., Stern Hal S. Estimating the entropy rate of finite Markov chains with application to behavior studies. Research. J Educ Behav Stat. 2019 10.3102/1076998618822540.
- 3Janicki-Deverts D, Cohen S, Matthews KA, Jacobs DR Jr. Sex differences in the association of childhood socioeconomic status with adult blood pressure change: the CARDIA study. Psychosom Med. 2012;74(7):728–35. 10.1097/PSY.0b 013e 31825 e 32e 8.10.1097/PSY.0b 013e 31825 e 32e 8PMC 343423022822232 · doi ↗ · pubmed ↗
- 4Rochira V, Carani C. Estrogens, Male Reproduction and Beyond. In: Feingold KR, Ahmed SF, Anawalt B, et al, eds. Endotext. 2000.25905171 · pubmed ↗