Taming the Achilles’ Heel: A Chemical and Structural Design to Address Off-Target Effects in siRNA Therapeutics
Rohith Pavan Parvathaneni, Nithiyanandan Krishnan, Nikolai Hempel, Oommen P. Oommen, Oommen P. Varghese

TL;DR
This paper introduces a new chemical and structural design for siRNA to reduce off-target effects while preserving its effectiveness in RNA interference.
Contribution
The novel 2′-diol modification and extended 3′-overhang design reduce off-target interactions in siRNA therapeutics.
Findings
2′-diol modifications in the seed-region reduced melting temperature and imposed thermodynamic asymmetry.
The US-siRNA design with a 3′-overhang improved antisense strand loading into RISC.
The combined strategy minimizes off-target effects without compromising on-target RNAi activity.
Abstract
Off-target effects represent one of the major bottlenecks for RNA interference (RNAi) technology. To address this issue, we present a novel strategy by combining seed-region chemical modification with an extended 3′-overhang on the sense strand (SS) to mitigate SS-mediated and miRNA-like nontargeted interactions. To modify the seed-region, we developed a novel 2′-diol modification that was selectively installed at different positions within the seed-region of siRNA. For this purpose, we synthesized universal 2′-diacetate phosphoramidites that yielded a free 2′-diol functionality after standard deprotection of oligonucleotides. The 2′-diol moieties with single (positions 3–7) and dual (6 + 7) insertions in the seed-region decreased the melting temperature (T m) by ca. −1 to −4.0 °C, imposing thermodynamic asymmetry. To improve the end-asymmetry of siRNA, we developed a structurally…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
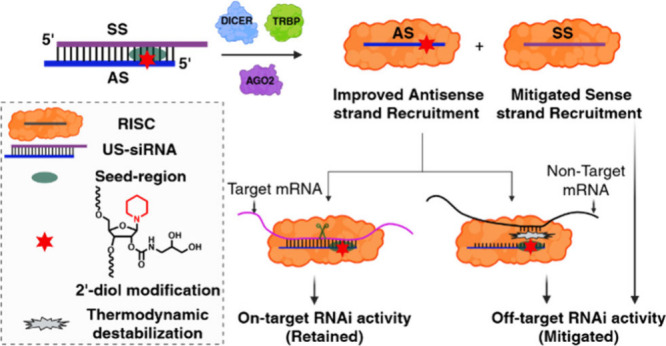 Figure 1
Figure 1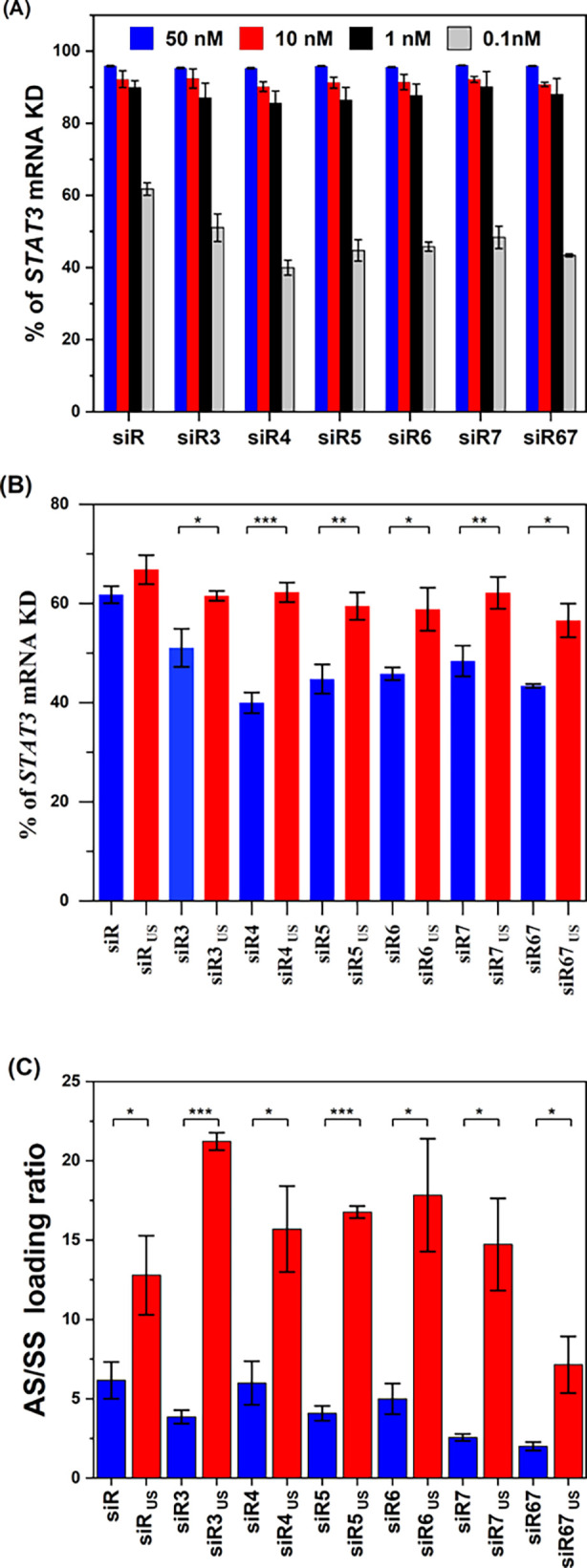 Figure 2
Figure 2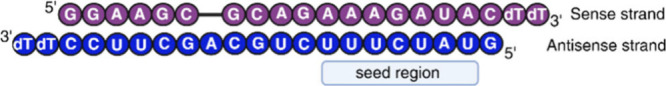 Figure 3
Figure 3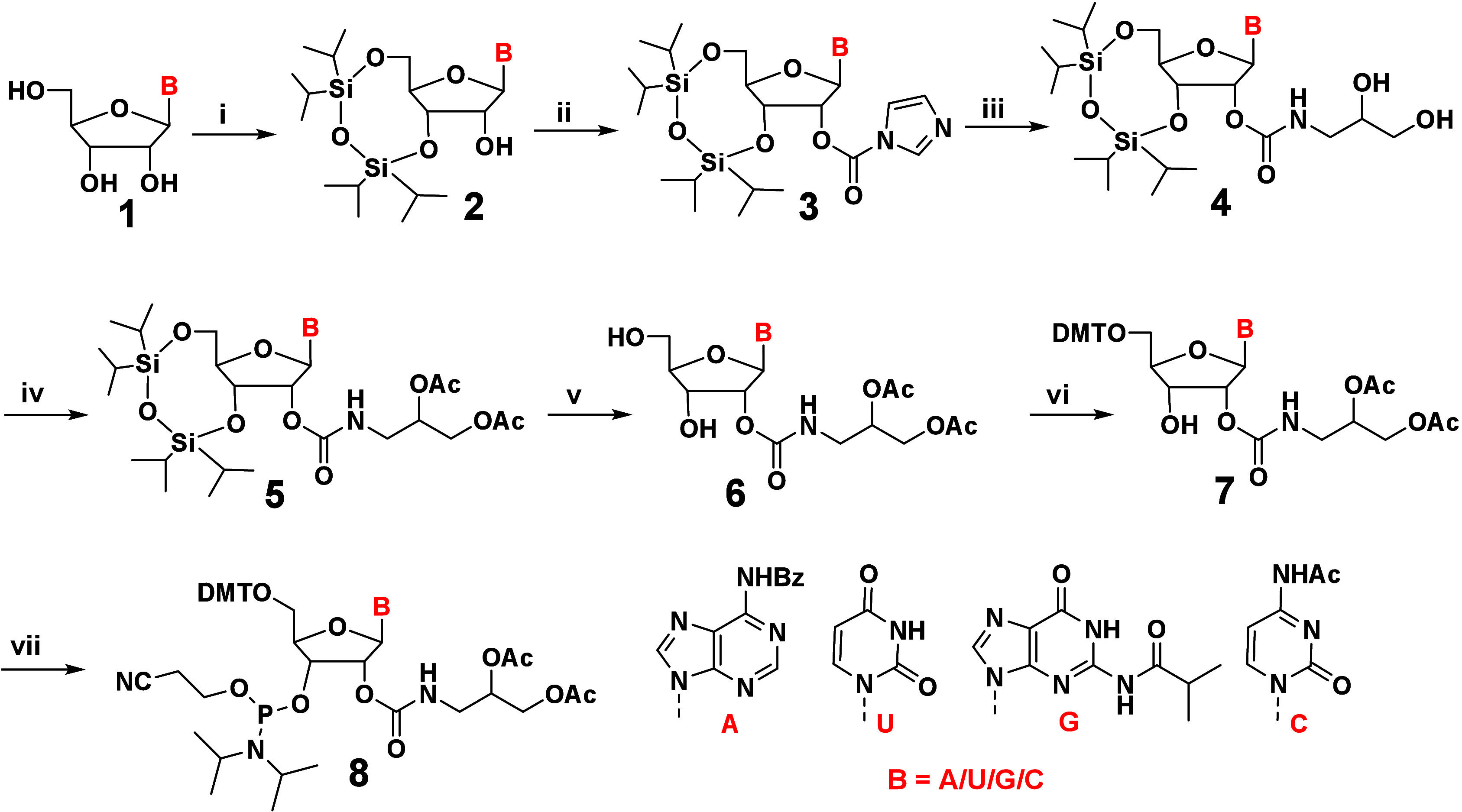 Figure 4
Figure 4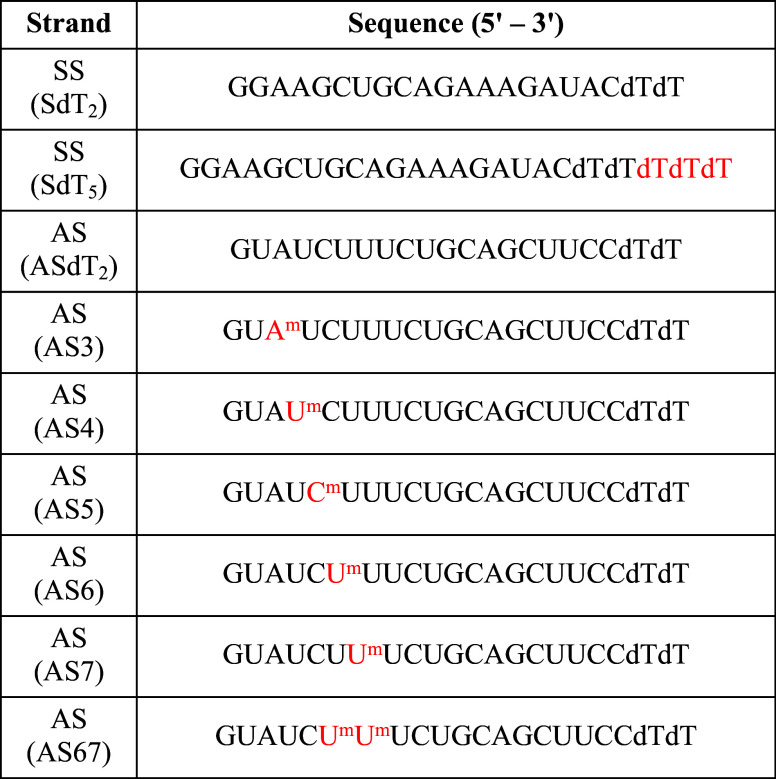 Figure 5
Figure 5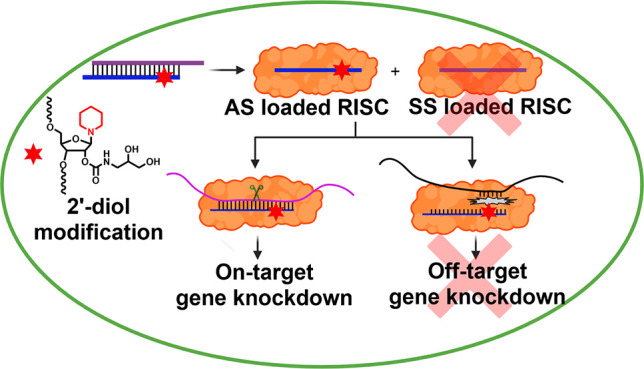 Figure 6
Figure 6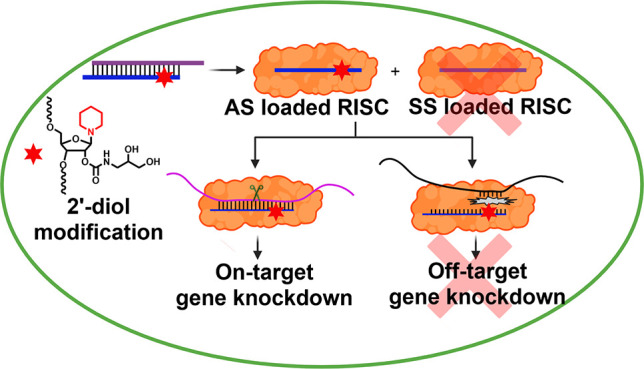 Figure 7
Figure 7- —H2020 Marie Sklodowska-Curie Actions10.13039/100010665
- —Vetenskapsr?det10.13039/501100004359
- —Vetenskapsr?det10.13039/501100004359
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA Interference and Gene Delivery · DNA and Nucleic Acid Chemistry · MicroRNA in disease regulation
RNA interference (RNAi) employing small interfering RNA (siRNA) and microRNA (miRNA) has emerged as a promising technology for the treatment of a variety of diseases.? Currently, there are seven siRNA-based drug formulations (extensively 2′-F and 2′-OMe modified siRNA) that are approved for clinical use to address rare genetic, metabolic, and hematologic disorders.? Although these strategies are primarily designed to address the in vivo stability aspects rather than siRNA-based off-target effects, it should be noted that such modifications impart a sequence-dependent effect on RNAi potency.? The siRNA structure consists of a double stranded sequence composed of sense (passenger) and antisense (guide) strand that undergoes a natural selection process by interacting with proteins such as DICER, TRBP, and Argonaute2 (Ago2), forming a RNA induced silencing complex (RISC) that is responsible for cleaving the target mRNA (mRNA) in a sequence-dependent manner.? This results in post-transcriptional gene silencing with high efficiency and at very low concentrations due to the catalytic activity of the activated RISC.? In the natural gene silencing event, the sense strand (SS) is degraded by Ago2 while the antisense strand (AS) is selectively recruited into RISC, which then binds to a cognate mRNA in a sequence-specific manner.? The selection of the correct strand by the RISC is dependent on differences in thermodynamic asymmetry at the two ends of the sequence. The MID domain of Ago2 selectively binds to the thermodynamically less stable 5′-end of siRNA, thereby regulating this selection process. ?,?
Today, there are advanced siRNA design algorithms ?−? ? that enable the selection of highly specific siRNA with natural thermodynamic asymmetry and minimal homology with other targets, thereby limiting nontargeted knockdown of gene transcripts. Nevertheless, the strand selection is not exclusive and SS is always recruited to some extent.? Another concern of RNAi is miRNA-like off-target effects that require partial complementarity with the seed-region (2–8 nucleotides from the 5′-end) of the AS, which governs the target recognition and gene silencing.?
To improve the strand selection, several approaches have been pursued, including developing asymmetric siRNA, ?,? dicer substrate RNA,? blocking phosphorylation at the 5′-end of the SS,? fork siRNA,? 4′-guanidinium-modified siRNA,? and single-stranded RNA.? We also recently reported that structural modifications of miRNA induce thermodynamic asymmetry to modulate selective strand recruitment (miR or miR*) into RISC by incorporating extended overhangs at the 3′-ends of either strands.? To reduce miRNA-like off-target effects, several approaches were implemented that aimed to lower the melting temperature (T m) within the seed-region.? Seed-region modifications included incorporating deoxy nucleotides,? unlocked nucleic acid (UNA),? glycol nucleic acid (S-GNA),? spacer amidite,? 2′-formamidino,? 2′-deoxy-2′-α-F-2′-β-C-methyl pyrimidine,? alkyl phosphonate,? and amide internucleoside linkages.? These modifications mitigated the off-target effects in a T m-dependent manner. Incorporation of 2′-O-methyl (2′-OMe)? and locked nucleic acids (LNA)? in the seed-region had positional effects with LNA modification showing detrimental impact on the RNAi activity. Thus, we envisaged the design of a modified siRNA that addresses both sense-mediated and miRNA-like off-target effects without compromising the RNAi activity. For this purpose, we developed the 2′-diol chemical modification that was coupled with a structurally modified unsymmetrical siRNA (US-siRNA) design with an extended SS overhang (Figure).
To introduce 2′-diol chemical modifications on oligonucleotides, 2′-diacetate-modified phosphoramidites of adenine (A), uracil (U), guanine (G) and cytosine (C) were synthesized from their respective nucleosides 1 (A, U, G and C) with N ^6^-benzoyl (Bz), N ^2^-isobutyryl (iBu), and N ^4^-acetyl (Ac) protection on the A, G, and C nucleobases, respectively. Briefly, the 3′-5′ hydroxyls were protected by TIPS (1,1,3,3-tetraisopropyldisiloxane) to obtain nucleosides 2. The free 2′-hydroxyl of 2 was activated with carbonyldiimidazole (CDI) to yield compounds 3, which were directly treated with 1-aminoglycerol to obtain 4. Free diols in compounds 4 were protected with an acetyl protecting group to obtain compounds 5. The TIPS group was deprotected using triethylamine trihydrofluoride to generate the 5′- and 3′-dihydroxy compounds 6. The 5′-hydroxyl groups of compounds 6 were exclusively protected using 4,4′-dimethoxytrityl chloride (DMT-Cl) to yield the DMT-protected nucleosides 7, which were subsequently treated with 2-cyanoethyl N,N-diisopropylchlorophosphoramidite (CEP-Cl) to generate phosphoramidites of the respective nucleobases (compounds 8, Scheme). These phosphoramidite building blocks were purified, dried, and directly used to generate 2′-diol-modified oligonucleotides by standard solid-phase oligonucleotide synthesis. The incorporation of diacetate protecting groups on the 2′-diol moiety streamlines the solid-phase oligonucleotide synthesis, as the acetates are quantitatively removed under standard deprotection conditions to regenerate the free 2′-diol functionality in the fully deprotected oligonucleotide.
As a model siRNA sequence, we chose a therapeutic siRNA against signal transducer and activator of transcription 3 (STAT3) mRNA, which is known for its oncogenic role in various cancer models.? The SS of the STAT3 siRNA was structurally modified with three additional deoxy-thymidine (dT) overhangs at the 3′-end to induce structural asymmetry, and the AS strands were chemically modified with a single 2′-diol insertion at positions 3, 4, 5, 6, 7 and dual substitutions at positions 6 and 7 to impart thermodynamic asymmetry and selective destabilization of the seed-region (Table). The various combinations of siRNA duplexes with SS overhang (SdT_2_ and SdT_5_) and AS seed-region (ASdT_2_, AS3, AS4, AS5, AS6, AS7, and AS67) variants were annealed together for subsequent biophysical and biochemical evaluations (Table S1).
To determine the thermal asymmetry, we performed duplex melting studies for various combinations of chemically and structurally modified siRNA duplexes (Table). The incorporation of 2′-diol moieties within the AS seed-region progressively induces thermal destabilization, yielding a larger T m drop as the modification is moved from position 3 toward position 7 (−1.2 to −2.5 °C) and culminating in a 4 °C drop when dual modifications at positions 6 and 7 are introduced. As anticipated, structurally modified US-siRNA with canonical AS (siR_US_) lowered the T m by 0.6 °C, suggesting thermodynamic asymmetry as a result of extended overhang (Table).? While the ΔT m of −0.6 °C is modest for US-siRNA alone, the convergence of chemical and structural perturbations act synergistically, where the flexible extended overhang amplifies the thermodynamic destabilization introduced by the 2′-diols, thereby exhibiting a greater net T m drop of −2.3 to −5.1 °C. Notably, the US-siRNA exhibits a pronounced T m drop (up to −1 °C) with respect to their canonical sense variants (structurally unmodified SS), making them particularly interesting candidates for further evaluation of RNAi activity and relative RISC loading.
To quantify the effect of chemical and structural modification on RNAi activity, we determined the STAT3 expression levels by quantitative PCR analysis in a human osteosarcoma cell line (MG63). We first quantified the effect of seed-region modification on the RNAi activity. For this purpose, we evaluated STAT3 knockdown levels of different 2′-diol-modified siRNA sequences siR3, siR4, siR5, siR6, siR7, and siR67, having modifications at the 3, 4, 5, 6, 7, and 6/7 positions, respectively (Table S1). Interestingly, these modifications retained full gene silencing potency across three concentrations (50, 10, and 1 nM) identical to canonical siRNA (siR). However, at 0.1 nM all 2′-diol variants exhibited a position-dependent reduction in knockdown relative to siR (FigureA). Interestingly, this position-dependent reduction in knockdown was circumvented when structurally unsymmetrical design was incorporated within these sequences (siR_US_, siR3_US_, siR4_US_, siR5_US_, siR6_US_, siR7_US_, and siR67_US_). This indicates that although the seed-region destabilization can trim the RNAi potency at subnanomolar dose, the structural modification on the SS overhang restores the functional performance and is well tolerated by RNAi machinery (FigureB and Figure S32).
To probe the mechanism, we employed the stem-loop RT-qPCR assay to determine if our chemical and structural design has any influence in relative abundance of AS or SS post-transfection.? The stem-loop experiment with both chemically and structurally modified siRNA revealed that the incorporation of the 3′-overhang itself resulted in an increase of in AS loading from 6.2 to 12.8 for siR and siR_US_. This suggests that our unsymmetrical design itself has an effect on selective AS loading. However, the insertion of 2′-diol in the seed region resulted in a modest reduction of relative AS/SS ratios to ∼4, ∼6, ∼4, ∼5, ∼2.5, and ∼2 for siR3, siR4, siR5, siR6, siR7, and siR67, respectively, despite their lower T m values. This apparent discrepancy suggests that the overall reduction in T m does not necessarily create the end-focused thermodynamic asymmetry, which is essential for the recognition by the MID-domain for selective strand selection. Interestingly, structurally modified US-siRNA sequences with 2′-diol insertions further augmented the AS/SS RISC loading ratios significantly with respect to their canonical sense variants by reaching ∼21 for siR3_US_, ∼15.5 for siR4_US_, ∼16.5 for siR5_US_, ∼17.5 for siR6_US_, ∼14.5 for siR7_US_, and ∼7 for siR67_US_ (FigureC). Although the relative abundance of a single strand in the cell lysate does not directly demonstrate the RISC loading of that strand, given the low enzymatic stability of unmodified single-stranded RNA, the observed increase implies stabilization of a specific strand by an intracellular mechanism, presumably RISC, imparting catalytic activity. The observed improvement of relative AS recruitment with US-siRNA reveals the preservation of the thermodynamic end-asymmetry that is augmented with 2′-diol substitutions, producing a synergistic enrichment of guide strand recruitment with improved RNAi activity. Such enrichment of guide strand recruitment can potentially minimize SS-mediated off-target effects, analogous to previous observation with UNA.?
Furthermore, to assess the capacity of 2′-diol modifications to discriminate against near-complementary off-targets, we introduced a single nucleotide bulge by omitting the uridine nucleoside at position 7 of the SS and annealed each chemically modified AS variant (Figure). Although the bulge lies distal to the diol insertions, all 2′-diol modified duplexes exhibited increased the T m drop relative to the canonical siRNA, with dual substitutions producing the greatest effect with an exceptional T m drop of −10.3 °C. This indicates that the 2′-diol modification amplifies the destabilizing effect even for sequences that has remote bulged regions (Table). Introducing a bulge together with an extended SS overhang better models partial miRNA-like pairing and amplified the mismatch discrimination, leading to even stronger thermal penalties for imperfect pairing (Table, T m drops up to −13.7 °C). Such pronounced destabilization is expected to minimize undesired binding with nontargeted mRNA transcripts, thereby maintaining on-target knockdown and thermodynamically discriminating the off-target effects. While T m measurements alone cannot predict the RISC-mediated targeting outcomes, these findings suggest that such chemical and structural modifications enhance the discrimination against imperfectly paired interactions, representing an intriguing avenue for future studies on off-target modulation.
In summary, we developed a rational siRNA design by combining chemical modifications in the seed region with a structural modification at the 3′-end of the SS. This strategy not only enhanced thermodynamic asymmetry but also improved the biophysical and biochemical properties of the siRNA. The 2′-diol substitutions thermodynamically destabilized the seed region and increased discrimination against imperfect duplex pairing, but when applied alone they lowered the relative RISC loading of the AS and reduced RNAi on-target activity. Remarkably, introducing an extended 3′-overhang on the SS restored end-focused thermodynamic asymmetry, thereby boosting AS loading while maintaining on-target activity. Together, these complementary modifications represent a promising design framework for future studies to evaluate their applicability across diverse siRNA sequences, enabling safer and more efficient gene silencing. Future work will focus on systematically exploring the sequence dependence of these modifications, elucidating their RISC-mediated off-target effects, and assessing their impact through transcriptome-wide assays. Such chemically and structurally engineered siRNAs could be readily integrated into current clinical modalities to achieve sustained, target-specific silencing with reduced side effects.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kanasty R.Dorkin J. R.Vegas A.Anderson D.Delivery Materials for si RNA Therapeutics Nat. Mater.2013121196797710.1038/nmat 376524150415 · doi ↗ · pubmed ↗
- 2Xiao B.Wang S.Pan Y.Zhi W.Gu C.Guo T.Zhai J.Li C.Chen Y. Q.Wang R.Development, Opportunities, and Challenges of si RNA Nucleic Acid Drugs Mol. Ther Nucleic Acids 202536110243710.1016/j.omtn.2024.10243739897581 PMC 11787428 · doi ↗ · pubmed ↗
- 3Foster D. J.Brown C. R.Shaikh S.Trapp C.Schlegel M. K.Qian K.Sehgal A.Rajeev K. G.Jadhav V.Manoharan M.Kuchimanchi S.Maier M. A.Milstein S.Advanced Si RNA Designs Further Improve In Vivo Performance of Gal N Ac-Si RNA Conjugates Mol. Ther.201826370871710.1016/j.ymthe.2017.12.02129456020 PMC 5910670 · doi ↗ · pubmed ↗
- 4Zamore P. D.Tuschl T.Sharp P. A.Bartel D. P.RN Ai: Double-Stranded RNA Directs the ATP-Dependent Cleavage of MRNA at 21 to 23 Nucleotide Intervals Cell 20001011253310.1016/S 0092-8674(00)80620-010778853 · doi ↗ · pubmed ↗
- 5Dykxhoorn D. M.Lieberman J.The Silent Revolution: RNA Interference as Basic Biology, Research Tool, and Therapeutic Annu. Rev. Med.20055640142310.1146/annurev.med.56.082103.10460615660519 · doi ↗ · pubmed ↗
- 6Matranga C.Tomari Y.Shin C.Bartel D. P.Zamore P. D.Passenger-Strand Cleavage Facilitates Assembly of Si RNA into Ago 2-Containing RN Ai Enzyme Complexes Cell 2005123460762010.1016/j.cell.2005.08.04416271386 · doi ↗ · pubmed ↗
- 7Khvorova A.Reynolds A.Jayasena S. D.Functional si RN As and Mi RN As Exhibit Strand Bias Cell 2003115220921610.1016/S 0092-8674(03)00801-814567918 · doi ↗ · pubmed ↗
- 8Schwarz D. S.Hutvágner G.Du T.Xu Z.Aronin N.Zamore P. D.Asymmetry in the Assembly of the RN Ai Enzyme Complex Cell 2003115219920810.1016/S 0092-8674(03)00759-114567917 · doi ↗ · pubmed ↗