Taxonomy and phylogeny of Pseudochaetosphaeronema associated with rubber trees from Yunnan Province, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
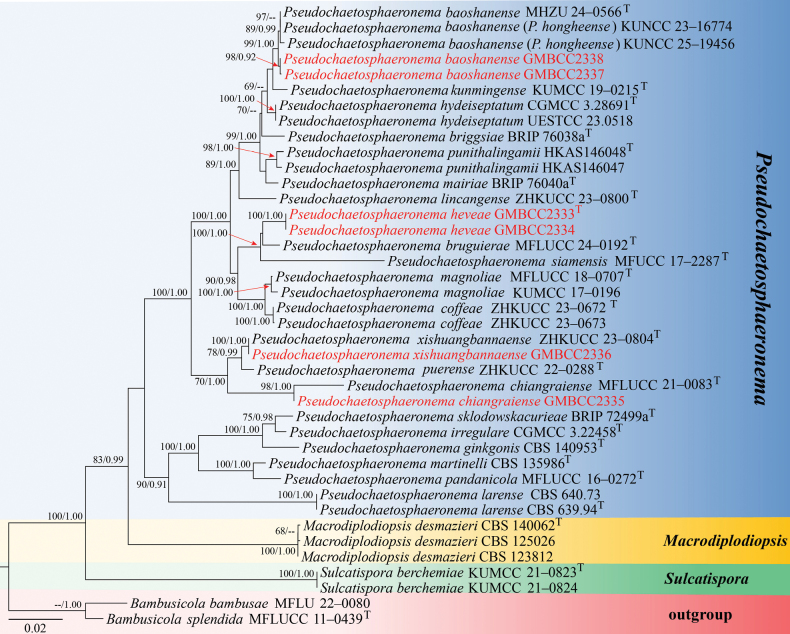 Figure 1
Figure 1 Figure 2
Figure 2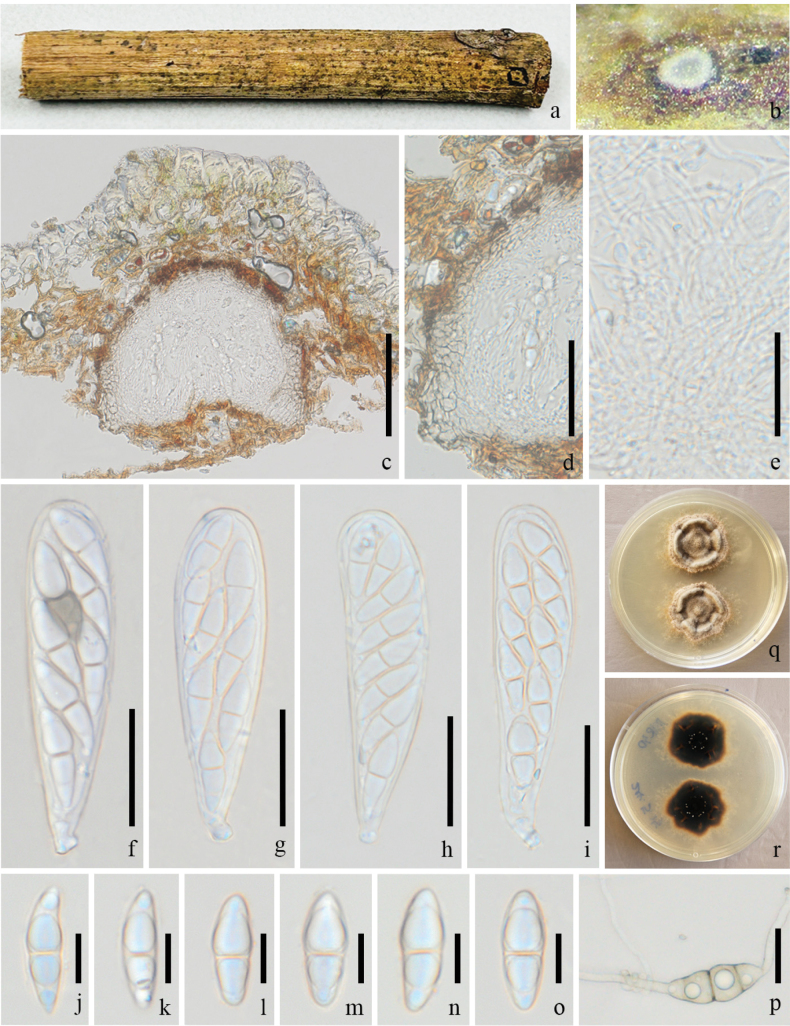 Figure 3
Figure 3 Figure 4
Figure 4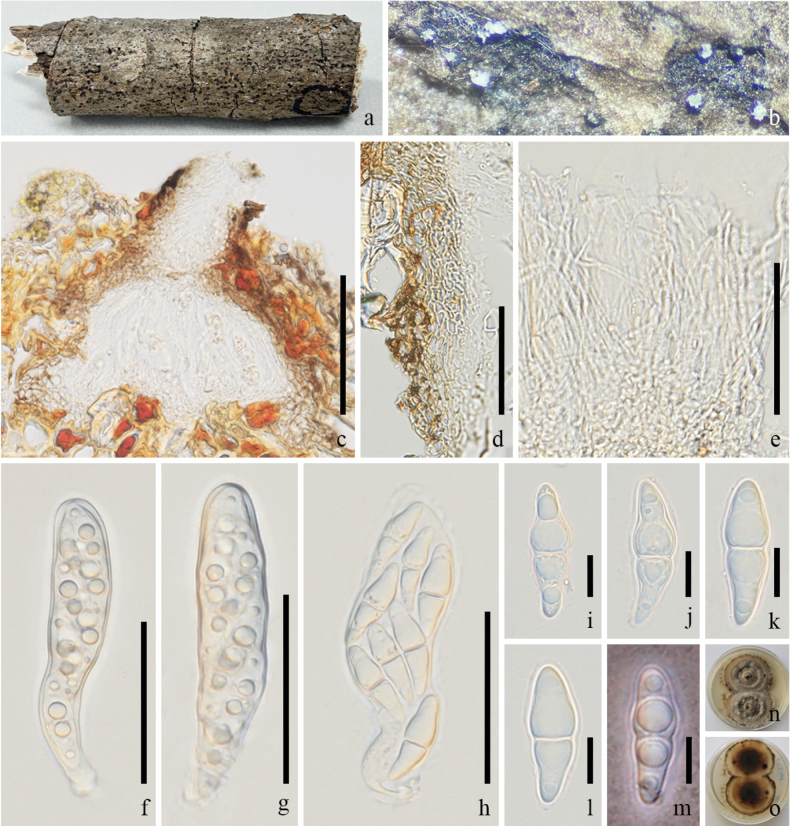 Figure 5
Figure 5| Species | Strain numbers | GenBank Accession Numbers | |||
|
|
|
| |||
|
| MFLU 22–0080 |
|
|
|
|
|
| MFLUCC 11–0439 T |
|
|
|
|
|
| CBS 123812 |
|
| N/A | N/A |
|
| CBS 125026 |
|
| N/A | N/A |
|
| CBS 140062 T |
|
| N/A | N/A |
|
| MHZU 24–0566 T |
|
|
|
|
|
| KUNCC 23–16774 |
|
|
|
|
|
| KUNCC 25–19456 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| BRIP 76038a T |
|
| N/A |
|
|
| MFLUCC 24–0515 T |
|
|
|
|
|
| MFLUCC 21–0083 T |
|
| N/A |
|
|
|
|
|
|
|
|
|
| ZHKUCC 23–0672 T |
|
|
|
|
|
| ZHKUCC 23–0673 |
|
|
|
|
|
| CBS 140953 T |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CGMCC 3.28691 T |
|
|
|
|
|
| UESTCC 23.0518 |
|
|
|
|
|
| CGMCC 3.22458 T |
|
|
|
|
|
| KUMCC 19–0215 T |
|
|
|
|
|
| CBS 639.94 T |
|
|
|
|
|
| CBS 640.73 |
|
|
|
|
|
| ZHKUCC 23–0800 T |
|
|
|
|
|
| KUMCC 17–0196 |
|
|
|
|
|
| MFLUCC 18–0707 T |
|
|
|
|
|
| BRIP 76040a T |
|
| N/A | N/A |
|
| CBS 135986 T |
|
|
|
|
|
| MFLUCC 16–0272 T |
|
|
| N/A |
|
| ZHKUCC 22–0288 T |
|
|
|
|
|
| HKAS146048 T |
|
|
|
|
|
| HKAS146047 |
|
|
|
|
|
| MFUCC 17–2287 T |
|
|
|
|
|
| BRIP 72499a T |
|
| N/A | N/A |
|
| ZHKUCC 23–0804 T |
|
|
|
|
|
| ZHKUCC 23–0805 |
|
|
|
|
|
|
|
|
|
|
|
|
| KUMCC 21–0823 T |
|
|
|
|
|
| KUMCC 21–0824 |
|
|
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Entomopathogenic Microorganisms in Pest Control
Introduction
Hevea brasiliensis (Euphorbiaceae) is a tropical tree extensively cultivated in Southeast Asia as the primary source of natural rubber (Nair 2021). Rubber is one of Yunnan’s major cash crops, particularly in Xishuangbanna (China), where plantations have expanded rapidly over recent decades and now cover a significant portion of the landscape, driving local income (Ling et al. 2022). Research on fungi associated with Hevea brasiliensis has primarily focused on pathogenic fungi, with an emphasis on identifying and controlling leaf, stem, and root diseases (Lin et al. 2021). Endophytic fungi have also received attention, particularly regarding their potential roles in plant growth, stress tolerance, and biocontrol (Xu et al. 2019). More recently, attention has expanded to saprobic fungi, which decompose dead plant material (Senwanna et al. 2021; Nizamani et al. 2023). Numerous taxa have been described from this host, underscoring its role as a reservoir of fungal diversity (Theodoro and Batista 2014; Senwanna et al. 2021; Xu et al. 2022, 2023, 2024, 2025; Manawasinghe et al. 2024; Liu et al. 2025).
The family Macrodiplodiopsidaceae, with Macrodiplodiopsis as its type genus, was established by Crous et al. (2015) within the suborder Massarineae (Pleosporales, Dothideomycetes). Subsequent studies expanded its generic composition to include Camarographium, Macrodiplodiopsis, Pseudochaetosphaeronema, and Pseudomonodictys in this family (Ariyawansa et al. 2015; Tanaka et al. 2015). However, later phylogenetic analyses and taxonomic revisions demonstrated that some of these genera are placed in other families (Wanasinghe et al. 2017; Phookamsak et al. 2022). As a result, the current circumscription of Macrodiplodiopsidaceae is more restricted, comprising only Macrodiplodiopsis and Pseudochaetosphaeronema (Wijayawardene et al. 2022; Hyde et al. 2024). Members of the family are mainly saprobes on decaying plants, although some species have been reported as opportunistic human pathogens (e.g. Pseudochaetosphaeronema larense, P. martinelli) (Ahmed et al. 2014, 2015; Boonmee et al. 2021; De Silva et al. 2022). Macrodiplodiopsidaceae is characterized by dark-brown, obovoid, asymmetric, euseptate ascospores in the sexual morph and by globose to subglobose conidiomata with ellipsoid to clavate conidia in the asexual morph (Crous et al. 2015).
Pseudochaetosphaeronema, one of the genera in Macrodiplodiopsidaceae, was established by Punithalingam (1979) with P. larense as the type species and currently comprises 20 species in Species Fungorum (2026). Species of Pseudochaetosphaeronema exhibit a widespread but predominantly tropical and subtropical distribution, with the highest diversity reported from Asia, particularly in China (Yunnan Province), and Thailand (Tibpromma et al. 2018; Jayasiri et al. 2019; Hyde et al. 2020; Boonmee et al. 2021; De Silva et al. 2022; Manawasinghe et al. 2024; Xu et al. 2024; Apurillo et al. 2025; Du et al. 2025; Lu et al. 2025; Wanasinghe et al. 2025); additional records have been reported from Australia, Venezuela, and Martinique (Punithalingam 1979; Ahmed et al. 2015; Tan and Shivas 2024). The genus has a broad host range, occurring primarily as a saprobe on diverse woody plants, including Bruguiera cylindrica, Cercis chinensis, Coffea sp., Hevea brasiliensis, Magnolia garrettii, Olea europaea, and Tamarindus sp. (Boonmee et al. 2021; De Silva et al. 2022; Li et al. 2023; Manawasinghe et al. 2024; Xu et al. 2024; Apurillo et al. 2025; Du et al. 2025; Lu et al. 2025). The members of Pseudochaetosphaeronema have been isolated as endophytes from Geijera salicifolia and Ginkgo biloba (Zhang et al. 2016; Tan and Shivas 2024) and as an opportunistic pathogen reported from human feet (Punithalingam 1979; Ahmed et al. 2014, 2015). Morphologically, the asexual morph is characterized by black, obpyriform pycnidia with a long neck, hyaline conidiophores, monoblastic or monophialidic conidiogenous cells, and subspherical to ellipsoidal conidia (Apurillo et al. 2025; Lu et al. 2025), while the sexual morph produces black, globose to subglobose ascomata with bitunicate asci and hyaline, fusiform ascospores (Boonmee et al. 2021; Li et al. 2023; Xu et al. 2024; Lu et al. 2025).
In this study, four Pseudochaetosphaeronema species were collected and isolated from rubber plants. Based on morphology and multi-gene phylogenetic analyses, we introduce a new species, report two new host records, and document an additional collection of Pseudochaetosphaeronema from H. brasiliensis in Yunnan Province, China. In previous studies, three species of Pseudochaetosphaeronema (P. lincangense, P. puerense, and P. xishuangbannaense) were discovered on rubber trees. This study increases the number of Pseudochaetosphaeronema species associated with H. brasiliensis to six. In addition, based on our phylogenetic analyses, P. hongheense (KUNCC 23–16774 and KUNCC 25–19456) is grouped together with P. baoshanense strains; therefore, we synonymized it under P. baoshanense based on morphology and multi-locus phylogenetic analyses.
Materials and methods
Sampling, examination, and isolation
Dead decaying branches of Hevea brasiliensis with ascomycetous fungal fruiting bodies were collected from Yunnan Province in China. Ecological data were recorded in accordance with Rathnayaka et al. (2024). Samples were transported to the mycology laboratory of Qujing Normal University in sealed plastic bags for further processing. Macro-morphological characteristics of fungal fruiting bodies were examined using a stereomicroscope (Leica S8AP0, Tokyo, Japan), while micro-morphological features were observed and photographed with a compound microscope (Olympus BX53, Tokyo, Japan). Fungi were isolated using single-spore isolation, as described by Senanayake et al. (2020). All morphological features were measured using Tarosoft (R) Image Framework version 0.9.7. and photographic plates were edited and combined in Adobe Photoshop CC 2017. Herbarium specimens were deposited at the Guizhou Medical University (GMB-W), China. Living cultures are deposited in the Guizhou Medical University Culture Collection (GMBCC). Facesoffungi (FoF) numbers and Index Fungorum Registration Identifiers (IF) were obtained as per Jayasiri et al. (2015) and Index Fungorum (2026), respectively. The data is also deposited in the Greater Mekong Subregion database (Chaiwan et al. 2021).
DNA extraction, PCR amplification, and sequencing
Fungal genomic DNA was extracted from one or two-month-old fresh fungal mycelium growing on potato dextrose agar (PDA) using a DNA Extraction Kit-BSC14S1 (BioFlux, Hangzhou, P.R. China) following the manufacturer’s instructions. The DNA product was kept at 4 °C for DNA amplification and at −20 °C for long-term preservation. DNA was amplified by polymerase chain reaction (PCR) for four genes: internal transcribed spacer region (ITS), partial 28S large subunit nuclear ribosomal DNA (LSU), partial 18S small subunit rDNA (SSU), and partial translation elongation factor 1-alpha (tef1-α). The partial ITS was amplified with ITS5 and ITS4 (White et al. 1990); LSU with LR0R and LR5 (Vilgalys and Hester 1990); SSU with NS1 and NS4 (White et al. 1990); and tef1-α with EF1-983F and EF1-2218R (Rehner and Buckley 2005). PCR was carried out in 25 μL reaction mixtures containing 12.5 μL 2× Master Mix (EasyTaq™ DNA Polymerase, dNTPs, and buffer; Beijing TransGen Biotech, Beijing, China), 8.5 μL ddH_2_O, 2 μL template DNA, and 1 μL of each primer (10 pM). The thermal cycling program followed Tibpromma et al. (2018) and Tian et al. (2024): initial denaturation at 94 °C for 3 min; 35 cycles of 94 °C for 30 s, annealing at 56 °C for 50 s, and extension at 72 °C for 1 min; and a final extension at 72 °C for 10 min. PCR products were purified and sequenced at Sangon Biotech Co., Kunming, China.
Phylogenetic analyses
Newly generated sequences were subjected to BLAST searches against NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi) to identify closely related taxa. Reference sequences from relevant publications (Du et al. 2025; Lu et al. 2025; Wanasinghe et al. 2025) and BLAST results were downloaded from GenBank and combined with the new sequences (Table 1). The relevant sequence data and newly generated DNA sequences were combined, and automatic alignment and editing were completed using OFPT (Zeng et al. 2023). Afterward, all datasets were concatenated into FASTA files; the alignments were visually checked and manually improved where necessary using the AliView program (Larsson 2014). Phylogenetic analyses were referenced in Dissanayake et al. (2020) and conducted using maximum likelihood (ML) and Bayesian posterior probability (BYPP) algorithms on the CIPRES Science Gateway portal (https://www.phylo.org/). ML analyses were performed with RAxML-HPC v.8, using the GTRGAMMA substitution model and 1,000 rapid bootstrap replicates. Bayesian analyses were conducted in MrBayes v.3.2.7a, with substitution models selected using MrModeltest v.2.2 (Nylander 2004); the best model for ITS, LSU, SSU, and tef1-α was GTR+I+G. Six simultaneous Markov chains were run for one million generations, automatically stopped when the topological convergence diagnostic reached 0.01, with a 25% burn-in. Trees were sampled every 100^th^ generation. The resulting trees were visualized using FigTree v.1.4.0 (Rambaut 2012) and edited in Microsoft PowerPoint 2021 and Adobe Photoshop CC 2017. The final alignments and trees were deposited in TreeBASE, under submission ID 32404 (Fig. 1) (http://www.treebase.org/).
Phylogram generated from maximum likelihood analysis based on combined LSU, ITS, SSU, and tef1-α sequences of 40 strains. Bootstrap values (ML ≥ 60%) and Bayesian posterior probabilities (BPP ≥ 0.90) are shown at the nodes. The tree is rooted with Bambusicola bambusae (MFLU 22–0080) and B. splendida (MFLUCC 11–0439). New isolates are indicated in red; ex-type strains are marked with “T.” Bootstrap values (ML ≤ 60%) and Bayesian posterior probabilities (BYPP ≤ 0.90) are only shown at the nodes for new isolates of this study.
Results
Phylogenetic analyses
The phylogenetic tree topologies obtained from RAxML and BI analyses were essentially similar. The RAxML analysis of the combined dataset yielded the best-scoring tree (Fig. 1), which was constructed from 3,167 base pairs (799 bp for LSU, 535 bp for ITS, 1,014 bp for SSU, and 819 bp for tef1-α). The final ML optimization likelihood value was -11667.153902. The matrix contained 650 distinct alignment patterns, with 11.78% of the characters being undetermined or gaps. The GTR+I+G model was applied for the combined ITS, LSU, SSU, and tef1-α dataset. The estimated base was: A = 0.239072, C = 0.251367, G = 0.268721, T = 0.240840; substitution rates AC = 1.455396, AG = 3.219017, AT = 1.80661, CG = 1.384182, CT = 8.948669, GT = 1.000000; and gamma distribution shape parameter α = 0.128109. The final RAxML tree is shown in Fig. 1.
In this phylogenetic tree, the results are similar to those reported by Du et al. (2025) and Wanasinghe et al. (2025). In this study, our six isolates clustered within Pseudochaetosphaeronema. Two strains of our isolates (GMBCC2337 and GMBCC2338) were grouped with P. baoshanense (MHZU 24–0566, holotype) and P. hongheense (KUNCC 23–16774: ex-type, KUNCC 25–19456) with 99% ML/1.00 BYPP statistical support. Another two of our strains (GMBCC2333, GMBCC2334) formed an independent lineage within Pseudochaetosphaeronema and were well distinct from the closely related species (P. bruguierae, MFLU 24–0192, holotype). The remaining two strains clustered with members of Pseudochaetosphaeronema. Strain GMBCC2335 clustered with the ex-type of P. chiangraiense (MFLUCC 21–0083) with 98% ML and 1.00 BYPP support, and GMBCC2336 clustered with the ex-type strain of P. xishuangbannaense (ZHKUCC 23–0804) with 100% ML and 1.00 BYPP support, which supports that they are the same species (P. chiangraiense and P. xishuangbannaense, respectively).
Taxonomy
Pseudochaetosphaeronema
baoshanense
Taxon classificationFungiPleosporalesMacrodiplodiopsidaceae
L. Lu, K.D. Hyde & Tibpromma Fungal Diversity 25: 1–275 (2025)
281EAF88-03D3-5D1B-9C73-90AF400ABA2E
Index Fungorum: IF903641
Facesoffungi Number: FoF17573
Description.
Saprobic on a decaying branch of Hevea brasiliensis. Sexual morph: Undetermined. Asexual morph: Coelomycetous. Conidiomata 140–170 × 170–200 μm (x̄ = 153 × 184 μm, n = 10), solitary, globose to subglobose or pyriform, dark brown, immersed or erumpent, with black spots on the host surface, unilocular. Conidiomatal wall 50–60 μm wide, composed of two layers, outer layers brown, inner layers pale brown to hyaline, arranged in a textura angularis to textura prismatica, darker at the outer layer, fusing and indistinguishable from the host tissues. Conidiophores arising from the inner layers lining the conidioma, or at the base, often reduced to conidiogenous cells. Conidiogenous cells 6–10 × 2–3 μm (x̄ = 7.23 × 2.14 μm, n = 15), monophialidic, cylindrical, integrated, hyaline or pale brown, smooth-walled. Conidia 11–15 × 5–6 μm (x̄ = 12.4 × 5.6 μm, n = 40), oblong to ellipsoidal, or fusoid, with rounded ends, sometimes truncate and narrow at the base, yellowish to pale brown, smooth, thick-walled, 1–3-septate, some slightly constricted at the septum.
Pseudochaetosphaeronema baoshanense (GMB-W1228, New host record). a. Host branch piece; b. Conidiomata on host (arrows show conidiomata); c, d. Longitudinal section of conidiomata; e. Conidiomatal wall; f, g. Conidiogenous cells with conidia; h–k. Conidia; l. Germinated conidium; m, n. Colonies on PDA (surface and reverse). Scale bars: 100 μm (c, d); 50 μm (e); 20 μm (f, g, l); 5 μm (h–k).
Culture characteristics.
Conidia germinated on PDA within 12 hours. Colonies circular, raised, brown, with entire margin; reverse dark brown at the center, fading to pale brown at the edge.
Material examined.
China, • Yunnan Province, Dehong Dai and Jingpo Autonomous Prefecture, Mang City, on a decaying branch of Hevea brasiliensis, 18 April 2024, Rui-Fang Xu, DHR23 (GMB-W1227), living culture GMBCC2337; China, Yunnan Province, Lincang City, 28 July 2022, LCR24 (GMB-W1228), living culture GMBCC2338.
Known distributions.
China, Yunnan Province (Lu et al. 2025; Wanasinghe et al. 2025; this study).
Known hosts.
Coffea sp. (Lu et al. 2025), dead twigs of an unknown deciduous plant (Wanasinghe et al. 2025), Hevea brasiliensis (this study).
Notes.
Phylogenetic analyses show that strains GMBCC2337 and GMBCC2338 clustered with Pseudochaetosphaeronema baoshanense (MHZU 24–0566, holotype) and P. hongheense (KUNCC 23–16774; KUNCC 25–19456) with 99% ML and 1.00 BI support (Fig. 1). Morphologically, the species isolated from Hevea brasiliensis is similar to the species described by Lu et al. (2025) from Coffea sp., in having cylindrical conidiogenous cells and yellowish to brown, oblong to ellipsoidal or fusoid conidia. However, the conidiomata wall of our samples (GMB-W1227, GMB-W1228) is thicker than the holotype of P. baoshanense (MHZU 24–0566) (50–60 μm vs. 20–40 μm), and conidia of our samples are sometimes slightly constricted at the septa, whereas in the holotype they are non-constricted. While the ITS, LSU, and SSU regions were identical between the new isolates (GMBCC2337 and GMBCC2337) and P. baoshanense (MHZU 24–0566), only the tef1-α gene region showed four base pairs differences. Based on the combined molecular evidence, our collections are identified as P. baoshanense. According to our knowledge, this is the first report of P. baoshanense on Hevea brasiliensis.
Pseudochaetosphaeronema hongheense was introduced by Wanasinghe et al. (2025), and this fungus was isolated from Honghe, Yunnan Province. In the present study, P. hongheense (KUNCC 23–16774, KUNCC 25–19456) clusters with P. baoshanense (MHZU 24–0566, holotype) with 97% ML support, but Bayesian support is low. Pairwise nucleotide comparisons showed no differences in ITS, LSU, and tef1-α sequences, with only a base-pair gap in SSU. Morphologically, P. hongheense (HKAS146045, holotype) and P. baoshanense (MHZU 24–0566, holotype) are similar in conidiomatal structure, conidiogenous cells, and brown, septate conidia. However, P. baoshanense differs from P. hongheense in thicker conidiomatal wall (20–40 μm vs. 5–15 μm), conidia that are oblong to ellipsoidal or fusoid and not constricted at the septa, and in producing 1–3-septate conidia that are only slightly thick-walled at maturity (Lu et al. 2025). Pseudochaetosphaeronema hongheense is characterized by distinctly constricted, consistently 3-septate, thick-walled conidia (Wanasinghe et al. 2025). Based on the molecular and morphological evidence, P. hongheense is considered a synonym of P. baoshanense.
Pseudochaetosphaeronema
chiangraiense
Taxon classificationFungiPleosporalesMacrodiplodiopsidaceae
Wijesinghe, Boonmee & K.D. Hyde, Fungal Diversity 111: 75 (2021)
A12C9031-F077-5A8A-A05C-D85ABDCB7425
Index Fungorum: IF558549
Facesoffungi Number: FoF09950
Description.
Saprobic on a decaying branch of Hevea brasiliensis. Sexual morph: Ascomata 170–280 × 200–280 μm (x̄ = 221 × 223 μm, n = 10), solitary, scattered, immersed, uniloculate, globose to subglobose. Peridium 30–50 μm wide, composed of several brown to pale brown cells of textura angularis. Hamathecium 1–2 μm wide, numerous, filiform, unbranched, pseudoparaphyses. Asci 80–123 × 23–29 μm (x̄ = 99 × 25 μm, n = 15), 8-spored, bitunicate, fissitunicate, cylindrical to clavate, with a short pedicel, apex rounded, with ocular chamber. Ascospores 20–32 × 7–13 μm (x̄ = 26 × 10 μm, n = 30), overlapping, 2–3 seriate, fusiform, with obtuse ends, 1-septate at the center, constricted at the septa, hyaline, guttulate, thick and smooth-walled. Asexual morph: Undetermined.
Pseudochaetosphaeronema chiangraiense (GMB-W1225, New host record). a. Host branch piece; b. Ascoma on the host; c. Longitudinal section of ascoma; d. Partial peridium; e. Pseudoparaphyses; f–i. Asci; j–o. Ascospores; p. Germinated ascospore; q, r. Colonies on PDA (surface and reverse). Scale bars: 100 μm (c); 30 μm (d–k); 20 μm (l–p).
Culture characteristics.
Ascospores germinated on PDA within 24 hours, and ascospores turned brown after germination. Colonies irregular, wrinkled, brown, crateriform with undulate margin; reverse dark brown.
Material examined.
China, • Yunnan Province, Dehong Dai and Jingpo Autonomous Prefecture, Mang City, on a decaying branch of Hevea brasiliensis, 18 April 2024, Rui-Fang Xu, DHR30 (GMB-W1225), living culture GMBCC2335.
Known distributions.
Thailand (Boonmee et al. 2021) and Yunnan Province, China (Li et al. 2023; this study).
Known hosts.
Tamarindus sp. (Boonmee et al. 2021), Olea europaea (Li et al. 2023), and Hevea brasiliensis (this study).
Notes.
Pseudochaetosphaeronema chiangraiense was collected from Tamarindus sp. in Chiang Rai, Thailand (Boonmee et al. 2021), and was also later recorded on Olea europaea in Sichuan Province, China (Li et al. 2023). In this study, our isolate (GMBCC2335) was grouped with P. chiangraiense (MFLUCC 21–0083, ex-type). The morphology of our collection (GMB-W1225) is similar to the holotype of P. chiangraiense (Boonmee et al. 2021), but differs only in size: holotype with a thinner peridium when compared to our collection (13–17 μm vs. 30–50 μm), and a bit big in pseudoparaphyses (2–4.5 μm vs. 1–2 μm); larger asci (50–110 × 15–30 μm vs. 80–123 × 23–29 μm), and longer ascospores (20–45 μm vs. 20–32 μm). However, the new isolate was genetically identical to P. chiangraiense in the ITS, LSU, and SSU regions but differed by three base pairs in the tef1-α gene. Therefore, we identify our strain as P. chiangraiense based on phylogenetic and morphological evidence. According to our knowledge, this is the first record of P. chiangraiense on H. brasiliensis in China.
Pseudochaetosphaeronema
heveae
Taxon classificationFungiPleosporalesMacrodiplodiopsidaceae
R.F. Xu, K.D. Hyde & Tibpromma sp. nov.
D18B6048-16B5-545F-8D03-71D0B8F3FABD
Index Fungorum: IF904508
Facesoffungi Number: FoF18921
Etymology.
Refers to the host genus Hevea.
Holotype.
GMB-W1222
Description.
Saprobic on a decaying branch of Hevea brasiliensis. Sexual morph: Undetermined. Asexual morph: Coelomycetous. Conidiomata 125–170 × 140–225 μm (x̄ = 148 × 190 μm, n = 10), solitary to aggregated, globose to subglobose or pyriform, immersed to semi-immersed, unilocular, dark brown to black, appearing as black dots on the host surface. Conidiomatal wall 20–70 μm wide, brown to dark brown, textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 3–8 × 2–6 μm (x̄ = 6 × 4.4 μm, n = 10), monophialidic, cylindrical or ampulliform, hyaline, smooth-walled, with guttules. Conidia 8–14 × 3–4 μm (x̄ = 11.25 × 3.23 μm, n = 50), fusiform or fusoid, slightly curved, 1–2-septate, constricted at septum, cell swollen sometimes, rounded at both ends, hyaline, smooth, thin-walled, without a mucilaginous sheath.
Pseudochaetosphaeronema heveae (GMB-W1222, Holotype). a. Host branch piece; b. Conidiomata on host; c. Longitudinal section of conidioma; d. Conidiomatal wall; e–h. Conidiogenous cells with conidia; i–m. Conidia; n. Germinated conidium; o, p. Colonies on PDA (surface and reverse). Scale bars: 100 μm (c), 50 μm (d), 30 μm (e), 10 μm (f–n).
Culture characteristics.
Conidia germinating on PDA within 12 hours. Colonies irregular, umbonate, white to cream, with a central brown droplet, undulate margin; reverse dark brown with pale brown edge, producing brown pigment on PDA media.
Material examined.
China, • Yunnan Province, Dehong Dai and Jingpo Autonomous Prefecture, Mang City, on a decaying branch of Hevea brasiliensis, 18 April 2024, Rui-Fang Xu, DHR27 (GMB-W1222, holotype), ex-type living culture GMBCC2333 = GMBCC2334.
Notes.
In the phylogenetic analyses, GMBCC2333 and GMBCC2334 formed a distinct lineage, sister to P. siamensis (MFLUCC 17–2287, ex-type) and P. bruguierae (MFLUCC 24–0515, ex-type) (Fig. 1). Sequence comparisons showed that GMBCC2333 differs from P. siamensis by 13/510 bp (ITS, 2.55%), 3/810 bp (LSU, 0.20%), 2/1,005 bp (SSU, 0.19%), and 99/645 bp (tef1-α, 15.35%); differences with P. bruguierae included 8/497 bp (ITS, 1.61%), 2/810 bp (LSU, 0.25%), and 28/893 bp (tef1-α, 3.14%). Morphologically, P. heveae (GMB-W1222, holotype) can be distinguished from P. coffeae (MHZU 23–0081, holotype) by smaller conidiomata (125–170 × 140–225 μm vs. 150–190 × 180–240 μm) and conidiogenous cells (3–8 × 2–6 μm vs. 6–9 × 3–6 μm), as well as shorter conidia (8–14 × 3–4 μm, 1–2-septate vs. 12–16 × 3–5 μm, 1–3-septate) (Lu et al. 2025). It also differs from P. bruguierae (MFLU 24–0192, holotype), which has larger conidiomata (230–400 × 300–370 μm), narrower conidiogenous cells (7–10 × 1–2.7 μm), and smaller conidia (5–10 × 2–3 μm), 2–3-septate with guttules (Apurillo et al. 2025). Based on molecular and morphological evidence, we introduce our new collection as a new species, P. heveae.
Pseudochaetosphaeronema
xishuangbannaense
Taxon classificationFungiPleosporalesMacrodiplodiopsidaceae
R.F. Xu & Tibpromma, MycoKeys 103: 71–95 (2024) [as ‘ xishuangbannaensis’]
685CFB43-F334-588A-B627-227D7D617B8C
Index Fungorum: IF901422
Facesoffungi Number: FoF15198
Description.
Saprobic on a decaying branch of Hevea brasiliensis. Sexual morph: Ascomata 110–330 × 212–335 μm, (x̄ = 218 × 263 μm, n = 10), white spot on the surface, sometimes inconspicuous on host surface, solitary, scattered, immersed, globose to subglobose or pyiform, or ellipsoid, uni-loculate, brown, with ostiole. Ostiole 70–155 × 60–85 μm, (x̄ = 105 × 70 μm, n = 5), central, brown, papillate. Peridium 30–60 μm wide, composed of several layers, brown to pale brown to hyaline cells of textura intricata. Hamathecium comprises 2–3 μm wide, numerous, filiform, unbranched, hyaline, cellular pseudoparaphyses. Asci 80–110 × 16–30 μm (x̄ = 95 × 21 μm, n = 15), 8-spored, bitunicate, obovoid, short, distinct pedicel, apex rounded with an ocular chamber. Ascospores 26–36 × 10–12 μm (x̄ = 30 × 11 μm, n = 20), hyaline, fusiform, with pointed ends, 1-septate, occasionally 3-septate, constricted at the middle septum, wider upper cell, constricted at the septa, guttulate, thick-walled, with a thin mucilaginous sheath. Asexual morph: Undetermined.
Pseudochaetosphaeronema xishuangbannaense (GMB-W1226). a. Host branch piece; b. Ascomata on the host; c. Longitudinal section of ascoma; d. Partial peridium; e. Pseudoparaphyses; f–h. Asci; i–l. Ascospores; m. Ascospore with India ink; n, o. Colonies on PDA (surface and reverse). Scale bars: 100 μm (c); 50 μm (d–h); 10 μm (i–m).
Culture characteristics.
Ascospores germinate on PDA within 24 hours. Colonies circular, umbonate, entire margin; brown from above, dark brown at center and edge, with brown ring on reverse.
Material examined.
China, • Yunnan Province, Dehong Dai and Jingpo Autonomous Prefecture, Mang City, on a decaying branch of Hevea brasiliensis, 18 April 2024, Rui-Fang Xu, DHR42 (GMB-W1226), living culture GMBCC2336.
Known distributions.
China, Yunnan Province, Xishuangbanna Dai Autonomous Prefecture (Xu et al. 2024), China, Yunnan Province, Dehong Dai and Jingpo Autonomous Prefecture (this study).
Known hosts.
Hevea brasiliensis (Xu et al. 2024, this study)
Notes.
In the phylogenetic analyses, strain GMBCC2336 formed a clade with strains of P. xishuangbannaense (ZHKUCC 23–0804 and ZHKUCC 23–0805) with 100% ML/1.00 BYPP statistical supports (Fig. 1). Compared with the holotype of P. xishuangbannaense (ZHKU 23–0107) with our sample (GMB-W1226) has smaller ascomata (110–330 × 212–335 μm vs. 270–410 × 370–480 μm), a thinner peridium (30–60 μm vs. 40–90 μm), smaller asci (80–110 × 16–30 μm vs. 130–180 × 25–35 μm), and smaller, 1–3-septate ascospores (26–36 × 10–12 μm vs. 30–50 × 10–20 μm, 3–5-septate) (Xu et al. 2024). Despite these size differences, the overall morphology and phylogeny align with those of P. xishuangbannaense. The nucleotide comparisons show differences of 3 bp (570 bp) and 1 bp (954 bp) of ITS and SSU, respectively, between the new isolate (GMBCC2336) and the ex-type of P. xishuangbannaense (ZHKUCC 23–0804). Therefore, strain GMBCC2336 is identified as P. xishuangbannaense and represents a newly collected specimen from Dehong, about 415 km from the type locality in Xishuangbanna.
Discussion
This study provides new insights into the diversity of Pseudochaetosphaeronema associated with Hevea brasiliensis in Yunnan Province, China. We introduce P. heveae as a novel species, establish new host records for P. baoshanense and P. chiangraiense, and provide an additional collection of P. xishuangbannaense. The presence of multiple Pseudochaetosphaeronema taxa on a single host highlights the rich diversity of saprobes in tropical and subtropical ecosystems. The rubber tree hosts a wide variety of fungi due to the warm, humid climate of tropical and subtropical regions where it grows, which further promotes fungal development and diversity (Talley et al. 2002; Senwanna et al. 2021). The number of Pseudochaetosphaeronema species from H. brasiliensis now increases to six with the addition of P. lincangense and P. puerense by Manawasinghe et al. (2024) and Xu et al. (2024).
Moreover, based on multi-locus phylogenetic analyses, pairwise nucleotide comparisons, and morphology comparisons, P. hongheense is synonymized with P. baoshanense. We observed slight morphological differences among collections of P. baoshanense, even though they are from Yunnan Province; these differences may be due to their local environment (Baoshan (type locality), Honghe (Wanasinghe et al. 2025), Lincang, and Dehong (this study), host plant (coffee plant, dead twigs of an unknown deciduous tree, and rubber tree), or geographical origin.
The recognition of Pseudochaetosphaeronema heveae as a distinct new species is strongly supported by a combination of morphological and phylogenetic evidence. Although phylogenetic analyses placed P. heveae in a separate lineage from P. siamensis, the statistical support was relatively low. This limited resolution is likely due to insufficient information obtained with only ITS, LSU, and SSU markers within the genus, as observed in previous studies; by contrast, the present study, which included tef1-α, provided greater discriminatory power. Nonetheless, the consistent tree topologies recovered from both ML and BI analyses, coupled with stable morphological characters, lend confidence to the delimitation of P. heveae. This case highlights the importance of integrating multiple loci, especially protein-coding, in fungal taxonomy. The low phylogenetic support for P. heveae suggests that additional loci, such as RPB2 and TUB2, may be needed to resolve intrageneric relationships among species. Our study was conducted extensively across Yunnan Province, and the high diversity of Pseudochaetosphaeronema species observed suggests that sampling of Hevea brasiliensis across Southeast Asia and South America may reveal more species (Boonmee et al. 2021; De Silva et al. 2022; Li et al. 2023; Xu et al. 2024; Lu et al. 2025). Future research integrating morphology, multigene phylogenetics, and ecological data will be crucial for clarifying species boundaries, host specificity, and functional roles of this genus in tropical ecosystems.
The discovery of Pseudochaetosphaeronema baoshanense and P. chiangraiense on Hevea brasiliensis represents new host records, reflecting the ecological adaptability of Pseudochaetosphaeronema. Pseudochaetosphaeronema baoshanense was originally described from Coffea sp. in Yunnan Province, China, while P. chiangraiense was first reported from Tamarindus sp. in Thailand and subsequently on Olea europaea in Sichuan Province, China (Boonmee et al. 2021; Li et al. 2023; Lu et al. 2025). Based on the distribution of their host plants across tropical and subtropical regions, P. baoshanense and P. chiangraiense appear well adapted to warm, humid environments. Their occurrence in these climatic conditions suggests that temperature plays an important role in shaping their ecological distribution and host ranges. Our study extends their host range to Hevea, suggesting that these fungi may be saprobes capable of colonizing a diverse range of woody substrates. Whether these species exhibit host preference or are generalist colonizers remains uncertain, but their repeated detection across unrelated hosts highlights the need for broader ecological surveys.
Our additional collection of Pseudochaetosphaeronema xishuangbannaense confirms its occurrences on Hevea brasiliensis beyond the original locality in Xishuangbanna, Yunnan (Xu et al. 2024). The isolate exhibited species-level consistency with the type material. Although some differences were observed in ascomata, peridium, asci, and ascospore size, such intraspecific variation is not uncommon among coelomycetous fungi; environmental or host-related factors may influence it. Further collections of P. xishuangbannaense from Hevea are needed to confirm its saprobic preferences in local rubber plantations. The species could potentially play a crucial role in wood decomposition.
Rubber tree plantations are managed forests where a single tree species is grown (Wang and Zhang 2025). However, some rubber plantations in China (in the provinces of Hainan and Yunnan) incorporate coffee, tea, and pineapple as intercrops (Qi et al. 2024). Among microorganisms, fungi are among the most important because they help break down material, recycle nutrients, and decompose dead material. This keeps the plantation ecosystem healthy. In this study, we found several Pseudochaetosphaeronema species on dead rubber tree wood in a saprobic life mode. However, they might switch their lifestyle. First, they live as endophytes within a healthy tree, and then become active decomposers when the wood dies. Furthermore, these wood-decaying fungi likely interact with other microbes.
In conclusion, this study documents the discovery of a new species, two new host records, and a new collection of Pseudochaetosphaeronema from H. brasiliensis in China. These findings not only expand the known diversity and host range of the genus but also underscore the ecological significance of saprobic fungi in rubber plantations. By integrating molecular and morphological data, we contribute to the taxonomic framework for Pseudochaetosphaeronema and lay a foundation for future studies on its biodiversity, ecology, and potential interactions with economically important hosts. Beyond documenting taxonomic novelties, this study emphasizes the broader ecological relevance of saprobic fungi associated with H. brasiliensis. A deeper understanding of these rubber-associated fungal communities will contribute to a more comprehensive assessment of plantation ecosystem health and provide a foundation for future studies on microbial interactions, functional diversity, and potential applications in biological control and biodegradation.
Supplementary Material
XML Treatment for Pseudochaetosphaeronema baoshanense
XML Treatment for Pseudochaetosphaeronema chiangraiense
XML Treatment for Pseudochaetosphaeronema heveae
XML Treatment for Pseudochaetosphaeronema xishuangbannaense
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahmed SA, van de Sande WWJ, Stevens DA, Fahal A, van Diepeningen AD, Menken SBJ, de Hoog GS (2014) Revision of agents of black-grain eumycetoma in the order Pleosporales. Persoonia 33: 141–154. 10.3767/003158514 X 684744 PMC 431293025737597 · doi ↗ · pubmed ↗
- 2Ahmed SA, Desbois N, Quist D, Miossec C, Atoche C, Bonifaz A, Hoog S (2015) Phaeohyphomycosis caused by a novel species: Pseudochaetosphaeronema martinelli. Journal of Clinical Microbiology 53. 10.1128/JCM.01456-15PMC 454091726135866 · doi ↗ · pubmed ↗
- 3Apurillo CCS, Phukhamsakda C, Hyde KD, Thiyagaraja V, Jones EBG (2025) New fungal genus, three novel species, and one new record from mangroves, with reclassification of Melanconiella (Melanconiellaceae) species. Myco Keys 116: 25–52. 10.3897/mycokeys.116.137351 PMC 1199761140235465 · doi ↗ · pubmed ↗
- 4Ariyawansa H, Hyde K, Jayasiri S, Buyck B, Kandawatte T, Dai D-Q, Dai Y, Daranagama D, Jayawardena R, Lücking R, Ghobad-Nejhad M, Niskanen T, Thambugala K, Voigt K, Zhao R-L, Li G-J, Doilom M, Boonmee S, Yang Z, Chen X-H (2015) Fungal Diversity Notes 111–252 – Taxonomic and phylogenetic contributions to fungal taxa. Fungal Diversity 75: 27–274. 10.1007/s 13225-015-0346-5 · doi ↗
- 5Boonmee S, Wanasinghe DN, Calabon MS, Huanraluek N, Chandrasiri SKU, Jones GEB, Rossi W, Leonardi M, Singh SK, Rana S, Singh PN, Maurya DK, Lagashetti AC, Choudhary D, Dai Y-C, Zhao C-L, Mu Y-H, Yuan H-S, He S-H, Phookamsak R, Jiang H-B, Martín MP, Dueñas M, Telleria MT, Kałucka IL, Jagodziński AM, Liimatainen K, Pereira DS, Phillips AJL, Suwannarach N, Kumla J, Khuna S, Lumyong S, Potter TB, Shivas RG, Sparks AH, Vaghefi N, Abdel-Wahab MA, Abdel-Aziz FA, Li G-J, Lin W-F, Singh U, Bhatt RP, Lee · doi ↗ · pubmed ↗
- 6Chaiwan N, Gomdola D, Wang S, Monkai J, Tibpromma S, Doilom M, Wanasinghe DN, Mortimer PE, Lumyong S, Hyde KD (2021) https://gmsmicrofungi.org: an online database providing updated information of microfungi in the Greater Mekong Subregion. Mycosphere: Journal of Fungal Biology 12: 1513–1526. 10.5943/mycosphere/12/1/19 · doi ↗
- 7Crous PW, Carris LM, Giraldo A, Groenewald JZ, Hawksworth DL, Hemández-Restrepo M, Jaklitsch WM, Lebrun MH, Schumacher RK, Stielow JB, van der Linde EJ, Vilcāne J, Voglmayr H, Wood AR (2015) The Genera of Fungi – fixing the application of the type species of generic names – G 2: Allantophomopsis, Latorua, Macrodiplodiopsis, Macrohilum, Milospium, Protostegia, Pyricularia, Robillarda, Rotula, Septoriella, Torula, and Wojnowicia. IMA Fungus 6: 163–198. 10.5598/imafungus.2015.06.01.11PMC 450008226203 · doi ↗ · pubmed ↗
- 8De Silva N, Hyde K, Lumyong S, Phillips A, Bhat D, Maharachchikumbura S, Thambugala K, Tennakoon D, Suwannarach N, Karunarathna S (2022) Morphology, phylogeny, host association and geography of fungi associated with plants of Annonaceae, Apocynaceae and Magnoliaceae. Mycosphere: Journal of Fungal Biology 13: 955–1076. 10.5943/mycosphere/13/1/12 · doi ↗