Reappraisal of the phylogenetic relationships of hirsutella-like fungi across Ophiocordycipitaceae and Polycephalomycetaceae (Hypocreales, Sordariomycetes), and the description of seven new species

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
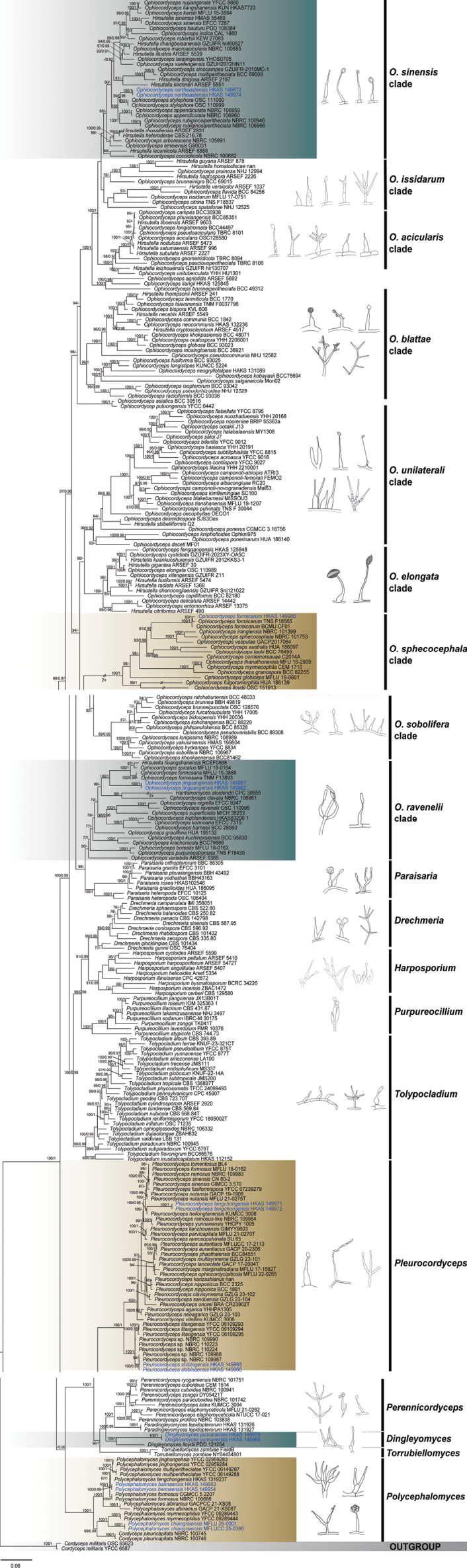 Figure 1
Figure 1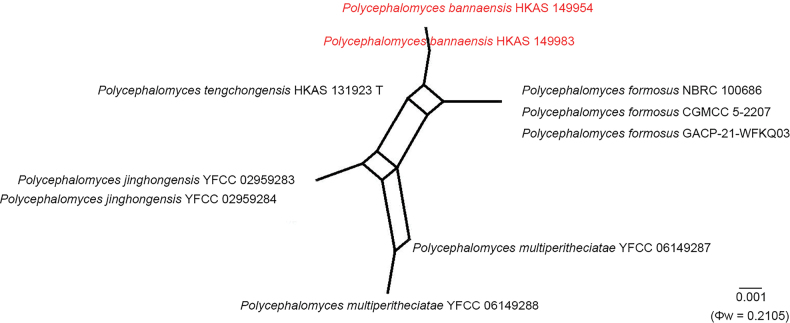 Figure 2
Figure 2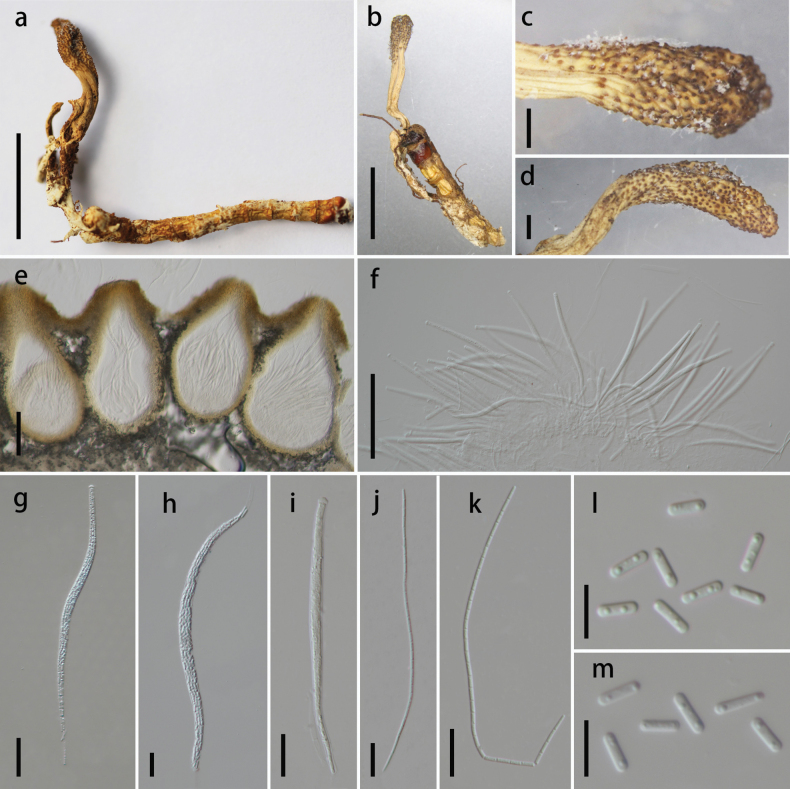 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5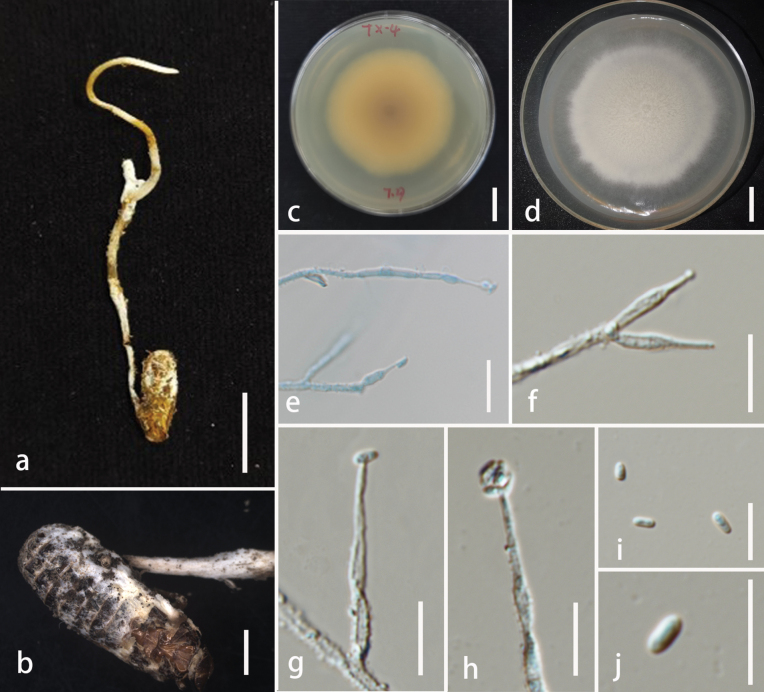 Figure 6
Figure 6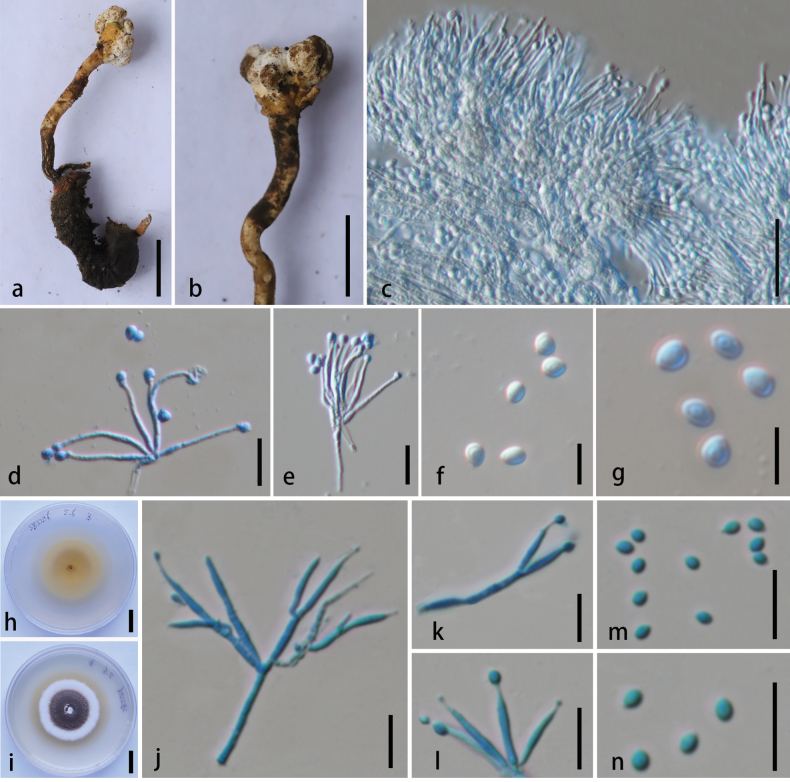 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9| Species | Voucher number | GenBank accession number | Reference | |||||
|---|---|---|---|---|---|---|---|---|
| ITS | nrSSU | nrLSU |
|
|
| |||
|
| NBRC 100745 |
|
|
|
|
|
|
|
|
| NBRC 100746 |
|
|
|
|
|
|
|
|
| PDD 121254 |
|
|
|
|
|
| Johnston et al. (2023) |
|
|
|
|
|
|
|
| ||
|
|
|
|
|
|
|
| ||
|
| CBS 250.82 |
|
|
|
|
|
|
|
|
| IMI 356051 |
|
|
| – | – | – |
|
|
| CBS 596.92 |
|
| – | – | – | – | Gernandt et al. (1999) |
|
| CBS 101434 |
| – | – | – | – | – |
|
|
| OSC 76404 |
|
|
|
|
|
|
|
|
| CBS 142798 |
|
|
|
| – | – |
|
|
| CBS 101432 |
| – | – | – | – | – | Zare et al. (2001) |
|
| CBS 567.95 |
|
|
|
|
|
|
|
|
| CBS 522.80 | – |
|
| – | – | – |
|
|
| CBS 335.80 |
|
|
|
|
|
|
|
|
| CPC 38655 |
| – |
| – | – | – |
|
|
| ARSEF 5407 | – | – |
| – | – | – |
|
|
| BCRC 34226 |
| – | – | – | – | – |
|
|
| CBS 129580 |
| – | – | – | – | – |
|
|
| ARSEF 5599 | – | – |
| – | – | – |
|
|
| ARSEF 5472 | – |
|
| – | – | – |
|
|
| Arsef 5354 | – |
|
| – | – | – |
|
|
| CPC 42872 |
| – |
|
| – |
|
|
|
| ZBAC1472 |
| – | – |
|
|
|
|
|
| ARSEF 5410 | – | – |
| – | – | – |
|
|
| GZUlFR hirl60527 |
| – | – |
| – | – |
|
|
| ARSEF 490 |
| – |
|
| – | – |
|
|
| ARSEF 4517 |
|
|
|
|
| – |
|
|
| ARSEF 5474 | – |
|
|
|
| – |
|
|
| ARSEF 30 | – | – |
|
|
| – |
|
|
| ARSEF 878 |
|
|
|
|
| – |
|
|
| ARSEF 2226 |
| – | – |
|
| – |
|
|
| CBS:216.78 | – | – |
| – | – | – |
|
|
| – | – |
| – | – | – |
| |
|
| RCEF0868 |
| – | – | – | – | – |
|
|
| ARSEF 5539 |
|
|
|
|
| – |
|
|
| ARSEF 5551 |
|
|
|
| – | – |
|
|
| GZUIFR-2012KKS3-1 |
| – |
|
|
| – |
|
|
| ARSEF 8888 |
|
|
|
|
| – |
|
|
| GZUIFR-hir130707 |
| – |
|
|
| – |
|
|
| ARSEF 9603 |
|
|
| – | – | – |
|
|
| ARSEF 5549 |
|
|
|
|
| – |
|
|
| ARSEF 5473 |
|
|
|
|
| – |
|
|
| ARSEF 1369 | – |
|
|
|
| – |
|
|
| ARSEF 2931 |
|
|
|
|
| – |
|
|
| ARSEF 996 |
|
|
|
|
| – |
|
|
| GZUIFR-Snj121022 |
| – |
| – |
| – |
|
|
| HMAS 55469 |
| – | – | – | – | – |
|
|
| Q2 | – |
| – | – | – | – |
|
|
| ARSEF 2197 |
|
|
|
|
| – |
|
|
| ARSEF 2227 |
|
|
|
|
| – |
|
|
| ARSEF 241 |
| – |
|
| – | – |
|
|
| ARSEF 1037 | – |
|
|
|
| – |
|
|
| OSC128580 |
|
|
|
|
| – |
|
|
| YFCC 9016 | – |
|
|
|
|
|
|
|
| ARSEF 5692 |
|
|
|
|
|
|
|
|
| RC20 | – |
| – |
| – | – |
|
|
| NBRC 106960 |
|
|
|
|
|
|
|
|
| NBRC 106959 |
|
|
|
|
|
|
|
|
| NBRC 105891 |
|
|
|
| – |
|
|
|
| BCC 30516 |
| – |
|
|
|
|
|
|
| HUA 186097 |
|
|
|
|
| – |
|
|
| BCC 28560 | – |
| – | – |
|
|
|
|
| YHH 20191 | – |
|
|
|
|
|
|
|
| YHH 20036 | – |
| – |
|
|
|
|
|
| YFCC 9012 | – |
|
|
|
|
|
|
|
| KVL 606 | – |
|
| – | – | – |
|
|
| MISSOU3 | – |
|
|
|
| – |
|
|
|
|
|
|
| – | – |
| |
|
| BBH 49819 | – | – | – |
|
|
|
|
|
| BCC 69015 | – | – |
|
| – |
|
|
|
| BCC 49312 | – | – |
|
| – |
|
|
|
| OSC 128576 | – |
|
|
|
|
|
|
|
|
| – |
|
|
|
|
| |
|
| ATRI3 | – |
|
|
| – | – |
|
|
| FEMO2 | – |
|
|
|
| – |
|
|
| Mal63 | – |
|
| – | – | – |
|
|
| BCC 82180 | – | – | – |
|
|
|
|
|
| TNS | – | – |
|
| – | – |
|
|
| NBRC 106961 |
|
|
|
|
|
|
|
|
| NBRC 100682 |
|
|
|
| – |
|
|
|
| BCC 1842 |
| – |
|
|
|
|
|
|
| YFCC 9027 | – |
|
|
|
|
|
|
|
| C2014A | – |
|
| – |
| – |
|
|
| GZUIFR-2023XY-OA5C | – |
|
| – |
|
|
|
|
| MF01 | – | – |
|
| – | – |
|
|
| ARSEF 14442 | – |
| – |
|
| – |
|
|
| SJS3Des | – |
|
|
|
| – | Saltamachia et al. (2020) |
|
| OSC 110989 | – | – |
|
|
| – |
|
|
|
| – | – | – | – | – |
| |
|
| ARSEF:13375 | – |
| – |
|
| – |
|
|
|
| – |
|
|
| – |
| |
|
| YFCC 8795 | – |
|
|
|
|
|
|
|
| BCC 84256 | – | – |
|
|
| – |
|
|
| BCMU CF01 |
| – | – | – | – | – |
|
|
| TNS | – |
|
|
|
|
|
|
|
|
|
|
|
|
| |||
|
| – |
| – |
|
| – |
| |
|
| TNM | – |
| – |
|
|
|
|
|
| HUA 186139 | – |
|
|
|
|
|
|
|
| YHH 17005 | – |
|
|
|
|
|
|
|
| BCC 93025 |
| – |
|
|
|
|
|
|
| TBRC 8094 | – | – |
|
|
|
|
|
|
|
|
|
|
| – | – |
| |
|
| BCC 93023 |
| – |
|
|
| – |
|
|
| HUA 186132 |
| – |
|
|
| – |
|
|
| BCC 82255 |
| – |
|
|
|
|
|
|
| MY1308 |
| – | – |
| – | – |
|
|
| PDD:108384 |
| – |
| – | – | – | Johnston et al. (2023) |
|
| HKAS83206-1 | – |
| – | – |
|
|
|
|
| YFCC 8834 | – |
|
|
|
|
|
|
|
| CAL 1880 |
|
|
|
|
| – | Shaema et al. (2023) |
|
| NBRC 101399 |
|
|
| – |
| – |
|
|
| BCC 93042 |
| – |
|
| – |
|
|
|
|
| – |
| – | – | – |
| |
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
|
| |
|
| – |
| – |
|
| – |
| |
|
| BCC 48071 |
| – |
|
|
| – |
|
|
| – |
| – |
|
|
|
| |
|
| SC100 | – | – |
|
|
| – |
|
|
| Ophkni975 | – |
|
|
|
|
|
|
|
| – |
|
|
|
|
|
| |
|
| BCC 88229 | – | – | – | – |
| – |
|
|
| EFCC 7315 | – |
| – |
|
|
|
|
|
| – | – |
|
|
|
|
| |
|
| BCC 95830 |
| – |
|
| – |
|
|
|
| YHOS0705 | – |
|
|
|
|
|
|
|
| BCC 76495 |
| – |
|
|
| – |
|
|
|
|
|
|
| – | – |
| |
|
| KUN-HKAS7723 | – | – | – | – |
| – |
|
|
| YHH 2210001 | – |
| – |
|
| – |
|
|
| OSC 151913 | – |
|
|
|
|
|
|
|
| NBRC 108989 |
|
|
|
| – | – |
|
|
|
|
|
|
|
|
|
| |
|
|
| – |
|
| – |
|
| |
|
| NBRC 100685 | – | – |
|
| – |
|
|
|
| BCC 36921 |
| – |
|
|
|
|
|
|
| BCC 69008 | – | – |
|
| – |
|
|
|
| CEM 1710 | – |
|
|
|
| – |
|
|
|
|
|
|
|
|
|
| |
|
| HAKS 131089 |
|
|
|
|
|
|
|
|
| EFCC 9247 |
|
|
|
|
|
|
|
|
| BRIP 55363a | – |
|
|
| – |
|
|
|
|
|
|
|
|
| |||
|
|
|
|
|
|
|
| ||
|
| YFCC 8880 | – |
|
|
|
|
|
|
|
| YHH 20168 | – |
|
|
|
| – |
|
|
| OECO1 | – |
| – | – | – | – |
|
|
| J13 | – |
|
|
|
| – |
|
|
| YHH 2206001 |
|
|
|
|
|
|
|
|
| TBRC 8106 | – | – |
|
| – |
|
|
|
| BCC 85328 | – | – |
|
|
|
|
|
|
|
| – | – |
|
|
|
| |
|
| HUA 186140 | – |
|
|
|
| – |
|
|
| CGMCC 3.18756 |
|
| – |
|
| – |
|
|
| NHJ 12994 | – |
|
|
|
|
|
|
|
| TBRC 8101 | – | – |
|
|
|
|
|
|
| NHJ 12582 | – |
|
|
| – |
|
|
|
| NHJ 12529 | – |
|
|
|
|
|
|
|
| BCC 88308 | – | – | – |
|
| – |
|
|
| YFCC 6442 | – |
|
|
|
|
|
|
|
| TNS F 30044 | – |
| – |
|
| – |
|
|
| TNS | – |
|
|
|
| – |
|
|
| BCC 93036 |
| – |
|
|
|
|
|
|
| BCC 48033 | – | – |
|
|
|
|
|
|
| OSC 110995 | – |
|
|
|
|
|
|
|
| KEW 27083 | – | – |
|
| – | – |
|
|
| NBRC 100946 |
|
|
|
|
|
|
|
|
| NBRC 106966 |
|
|
|
|
|
|
|
|
| Mori02 | – |
|
|
|
|
|
|
|
| J7 | – |
|
|
|
| – |
|
|
| EFCC 7287 |
|
|
|
|
|
|
|
|
| GZUIFR-2010MC-1 |
| – |
|
| – |
|
|
|
| NBRC 106967 |
|
|
|
| – |
|
|
|
| NHJ 12525 | – |
|
|
|
|
|
|
|
| NBRC 101753 |
|
|
|
|
|
|
|
|
|
|
|
|
| – | – |
| |
|
| OSC 111000 |
|
|
|
|
|
|
|
|
| OSC 110999 | – |
|
|
|
|
|
|
|
| YFCC 8815 | – |
|
|
|
|
|
|
|
| MICH 36253 | – |
| – | – |
| – |
|
|
| TNM F0037796 |
|
|
| – | – | – |
|
|
| BCC 1770 |
| – |
|
|
|
|
|
|
| MFU 16-2909 |
| – |
|
|
| – |
|
|
| – |
|
|
| – | – |
| |
|
| YHH HU1301 | – |
|
|
|
| – |
|
|
| ARSEF 5365 | – |
|
|
|
|
|
|
|
| GACP2017064 |
| – |
|
| – |
|
|
|
| GZUIFR Z11 |
|
|
|
|
| – |
|
|
| GZUH2012HN11 |
|
| – |
|
| – |
|
|
| HMAS 199604 | – |
|
| – |
|
|
|
|
|
| – |
| – |
|
|
| |
|
|
| – |
|
|
| – |
| |
|
| HUA 186095 | – |
| – |
|
| – |
|
|
| EFCC 3101 | – |
|
|
|
|
|
|
|
| EFCC 10125 |
|
|
|
|
|
|
|
|
| OSC 106404 | – |
|
|
|
| – |
|
|
| BBC 88305 |
| – |
|
|
| – |
|
|
| BBH 43492 |
| – |
|
|
| – |
|
|
| HKAS102546 |
|
|
|
|
|
|
|
|
|
| – |
|
|
| – |
| |
|
| NBRC 100941 |
|
|
| – |
| – |
|
|
| CEM 1514 | – |
|
|
| – | – |
|
|
|
|
|
|
|
|
|
| |
|
| NTUCC 17-021 |
| – |
|
|
|
|
|
|
| KUMCC 3004 | – | – |
| – | – | – |
|
|
| NBRC 101742 |
|
|
|
|
|
|
|
|
| NBRC 103838 |
|
|
| – |
| – |
|
|
| NBRC 101751 |
|
|
|
|
| – |
|
|
| DY05421 |
| – |
|
| – |
|
|
|
| YHHPA1305 |
|
| – |
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
| GACP 20-2306 |
|
|
|
| – |
|
|
|
| GZLG 23-102 |
| – |
|
| – | – |
|
|
|
|
|
|
| – | – |
| |
|
| YFCC 07239279 |
| – |
|
|
|
|
|
|
| KUMCC 3008 |
|
|
|
|
|
|
|
|
|
|
|
| – | – | – | Nikoh et al. (2000) | |
|
| GACP 17-2004 |
|
|
|
|
|
|
|
|
| GIMYY9603 |
|
|
|
|
| – |
|
|
| YFCC 06109293 |
|
|
|
|
| – |
|
|
| YFCC 06109294 |
|
|
|
|
|
|
|
|
| YFCC 06109295 |
|
|
|
|
| – |
|
|
|
|
|
|
|
|
|
| |
|
| GZLG 23-101 |
|
|
| – | – |
|
|
|
| GZLG 23-103 |
| – |
| – | – | – |
|
|
| BCC 1881 | – |
|
|
| – |
|
|
|
| BCC 2325 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
| GACP 19-1906 |
|
|
|
|
|
|
|
|
| BRA CR23902 |
| – | – | – | – | – |
|
|
|
|
|
|
|
| – |
| |
|
|
|
|
|
|
|
|
| |
|
| BCC 84551 |
| – |
|
|
| – |
|
|
| SU 65 | – | – |
|
|
| – |
|
|
| NBRC 109983 |
| – |
| – | – | – |
|
|
| NBRC 109984 |
|
|
|
|
| – |
|
|
| GZLG 23-104 |
| – |
|
| – |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CN 80-2 |
|
|
|
|
|
|
|
|
| GIMCC 3.570 |
|
|
|
|
| – |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| NBRC 109987 |
| – |
| – | – | – | Unpublished | |
| NBRC 109988 |
| – |
| – | – | – | Unpublished | |
| NBRC 109990 |
| – |
| – | – | – | Unpublished | |
| NBRC 110223 |
| – | – | – | – | – | Unpublished | |
| NBRC 110224 |
| – |
| – | – | – | Unpublished | |
|
| BL4 |
|
|
|
|
|
|
|
|
| KUMCC 3006 |
| – |
|
|
|
|
|
|
| YHCPY 1005 |
| – | – |
|
|
|
|
|
| GACP 21-XS08 |
|
|
|
|
|
|
|
|
| GACPCC 21-XS08 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
|
| |
|
| NBRC 100686 |
|
|
|
|
|
|
|
|
| CGMCC 5.2207 |
|
|
|
|
|
| wang et al. (2020) |
|
| YFCC 02959283 |
|
|
|
|
|
|
|
|
| YFCC 02959284 |
|
|
|
|
|
|
|
|
| YFCC 06149287 |
|
|
|
| – |
|
|
|
| YFCC 06149288 |
|
|
|
|
|
|
|
|
| YFCC 09289443 |
|
|
|
|
|
|
|
|
| YFCC 09289444 |
|
|
|
|
|
|
|
|
|
|
|
| – |
|
|
| |
|
| CBS 744.73 |
|
|
|
|
| – |
|
|
| JX13B01 |
| – |
|
| – | – |
|
|
| FMR 10376 | – | – |
|
|
| – |
|
|
| CBS 431.87 |
| – |
|
|
|
|
|
|
| IOM 325363.1 |
| – |
| – | – | – | Calvillo-Medina et al. (2021) |
|
| IBRC-M 30175 |
| – | – | – | – | – |
|
|
| NHJ_3497 | – |
|
|
|
|
|
|
|
| TK041 |
| – |
|
| – | – |
|
|
| CBS 393.89 |
| – |
| – | – | – |
|
|
| LA100 |
|
|
|
|
| – |
|
|
| ARSEF 2920 |
| – |
|
|
|
|
|
|
| ZBAH632 |
| – | – | – | – | – |
|
|
| MS337 | – |
|
|
|
| – |
|
|
|
| – |
|
| – | – |
| |
|
| CBS 723 70 |
| – | – | – | – | – |
|
|
| KNUF-22-14A |
|
|
| – | – | – |
|
|
| OSC 71235 |
|
|
|
|
|
|
|
|
|
|
|
|
| – |
|
| |
|
| CBS 568.84 | – | – |
| – | – | – |
|
|
| NBRC 106332 |
|
|
| – |
| – |
|
|
| NBRC:100945 |
|
|
|
|
|
|
|
|
| CPC 45907 |
| – |
|
|
| – |
|
|
| TFCC 24099493 |
|
|
|
|
| – |
|
|
| YFCC 875 | – |
|
|
|
|
|
|
|
| YFCC 1805002 | – |
|
|
|
|
|
|
|
| YFCC 879 | – |
|
|
|
|
|
|
|
| JMS200 |
| – |
|
|
| – |
|
|
| KNUF-23-321C |
| – |
|
| – | – |
|
|
| JMS111 |
| – |
|
|
| – |
|
|
| CBS 136897 | – | – |
|
|
| – |
|
|
| CBS:569.84 |
| – |
| – | – | – | Bissett et al. (1983) |
|
| LSB 131 |
|
|
| – | – | – |
|
|
| YFCC 877 | – |
|
|
|
| – |
|
|
| NY04434801 | – |
|
|
|
|
|
|
|
| FieldB | – |
|
|
| – | – |
|
|
| OSC 93623 |
|
|
|
|
|
|
|
|
| YFCC 6587 | – |
|
|
|
|
|
|
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEntomopathogenic Microorganisms in Pest Control · Plant Pathogens and Fungal Diseases · Fungal Biology and Applications
Introduction
Entomopathogenic fungi represent a group specialized in infecting insects. These organisms are distributed across five major fungal phyla, with the highest taxonomic diversity concentrated within Hypocreales (Araújo and Hughes 2016; Thiyagaraja et al. 2025). These fungi are classified into four families: Clavicipitaceae, Cordycipitaceae, Ophiocordycipitaceae, and Polycephalomycetaceae (Sung et al. 2007; Xiao et al. 2023). However, species possessing hirsutella-like asexual morphs are primarily distributed in the families Ophiocordycipitaceae and Polycephalomycetaceae. Ophiocordycipitaceae was proposed by Sung et al. (2007) and originally classified as a subgenus within Cordyceps. Over time, Ophiocordycipitaceae has undergone extensive taxonomic revisions integrating morphological and phylogenetic analyses to resolve its classification and delineate boundaries among constituent genera (Sung et al. 2007; Quandt et al. 2014; Mongkolsamrit et al. 2020; Crous et al. 2020; Araújo et al. 2022; Xiao et al. 2023). Notably, Ophiocordyceps, the type genus of this family, is recognized for its dark, fibrous to wiry stromata and perithecia, with a wide range of hosts (Sung et al. 2007; Ban et al. 2015; Araújo et al. 2018; Xiao et al. 2019). Other genera within Ophiocordycipitaceae also display remarkable morphological diversity and ecological strategies. Drechmeria includes soilborne nematophagous fungi characterized by solitary or verticillate conidiophores (Yu et al. 2018). Harposporium comprises obligate endoparasites of nematodes and rotifers, producing distinctive sickle-shaped conidia (Wang et al. 2007; Yadav et al. 2023). Tolypocladium is notable for its clavicipitoid sexual morphs and production of bioactive secondary metabolites, including cyclosporin derivatives (Yu et al. 2021). Purpureocillium, best represented by P. lilacinum, includes opportunistic entomopathogenic and nematophagous species with characteristic lilac to purple conidiophores (Luangsa-ard et al. 2011). Mongkolsamrit et al. (2019) resurrected the genus Paraisaria by introducing three new species. Recently established genera Hantamomyces and Torrubiellomyces further broaden the morphological and ecological range of the family (Crous et al. 2020; Araújo et al. 2022). Together, these genera illustrate the broad evolutionary diversification and ecological adaptability of Ophiocordycipitaceae. Polycephalomycetaceae, on the other hand, exhibits remarkable ecological adaptability, parasitizing diverse insect and fungal hosts across tropical and subtropical regions. This family was established by Xiao et al. (2023) to accommodate Perennicordyceps, Pleurocordyceps, and Polycephalomyces, which were previously assigned to Ophiocordycipitaceae. Polycephalomycetaceae is supported as a distinct lineage based on combined morphological characters from both sexual and asexual morphs, together with multigene phylogenetic analyses (ITS, nrSSU, nrLSU, tef1-α, rpb1, rpb2). Pleurocordyceps is characterized by stipitate synnemata, acremonium- to hirsutella-like phialides, and dimorphic conidia, representing a well-supported lineage in Polycephalomycetaceae. Polycephalomyces is a morphologically conservative but phylogenetically distinct genus, mainly parasitizing insects and other fungi, and is characterized by synnematous asexual morphs with hirsutella-like phialides. Dingleyomyces, a parasitic genus on the stromata of large Ophiocordyceps species, was introduced by Johnston et al. (2023). The genus was previously placed in Torrubiella as T. lloydii. Wang et al. (2024) established Paradingleyomyces to accommodate the monotypic species Pa. lepidopterorum based on morphological characteristics and multigene phylogenetic analysis.
Hirsutella-like fungi display substantial morphological plasticity, often obscuring generic and species boundaries. As a result, hirsutella-like fungi have played a central role in debates over taxonomic delimitation and evolutionary relationships within these families, warranting focused discussion. Speare (1920) originally defined Hirsutella as synnematous fungi with basally inflated phialides arranged in a hymenial layer and mucous-sheathed conidia. Subsequent studies expanded the genus to include polyphialides, non-inflated phialides, capitate synnemata, non-mucous conidia, didymoconidia, and multiple synanamorphs, reflecting greater morphological diversity (Hodge 1998). Quandt et al. (2014) noted that hirsutella-like fungi are widely distributed within Ophiocordyceps. The resulting morphological ambiguity, ecological overlap, and the need for taxonomic stability prompted the transfer of most hirsutella-like taxa into Ophiocordyceps. Consequently, newly discovered hirsutella-like fungi have predominantly been classified under Ophiocordyceps. Simmons et al. (2015) established a foundational classification system for hirsutella-like anamorphs, delineating six subclades: H. citriformis, H. guyana, H. nodulosa, H. sinensis, H. thompsonii, and a distinct “ant pathogen” subclade. Qu et al. (2018b) later provided comprehensive morphological descriptions for the first five subclades, while Araújo et al. (2018) redefined the “ant pathogen” group as the O. unilateralis clade. Subsequent studies expanded the phylogenetic framework, proposing additional hirsutella-like clades such as O. sobolifera and O. ravenelii (Wang et al. 2018; Sun et al. 2024). For instance, Dai et al. (2024) merged four subclades (H. guyana, H. nodulosa, H. sinensis, and H. thompsonii) into a broader O. sinensis clade, while Xie et al. (2025) proposed two novel clades (O. issidarum and O. acicularis), which have not previously been recognized. These taxonomic inconsistencies highlight the necessity of synthesizing and stabilizing the phylogenetic framework of clades characterized by hirsutella-like anamorphs.
In this study, we collected eight entomopathogenic fungi from Guizhou, Yunnan, and Liaoning Provinces in China, as well as Chiang Rai Province in Thailand. Three of the eight species were recognized as members of Ophiocordyceps, and five species were classified within Polycephalomycetaceae, based on macroscopic and microscopic characteristics. A phylogenetic analysis using ITS, nrSSU, nrLSU, tef1-α, rpb1, and rpb2 was conducted to clarify the taxonomic placements of these species. In addition, given the long-standing taxonomic complexity of hirsutella-like fungi, we further summarized the distribution and morphological diversity of hirsutella-like clades across Ophiocordycipitaceae and Polycephalomycetaceae. This overview provides essential phylogenetic context for interpreting the newly described species and establishes a clearer framework for evaluating convergent phialide morphologies within these families.
Materials and methods
Sample collection, morphological observation, and isolation
Insect specimens infected with entomopathogenic fungi were collected from Guizhou, Liaoning, and Yunnan Provinces in China, as well as from Chiang Rai Province in Thailand. Specimens were obtained from the lower and upper surfaces of living leaves and from leaf litter in both evergreen and deciduous forests with low sunlight exposure. The hosts of some specimens were buried in soil, such as Polycephalomyces bannaensis. Fresh specimens were photographed in the field using a mobile phone camera (Huawei P40), placed in plastic boxes, and transported to the laboratory for further analysis. Fresh fruiting bodies were examined using a stereomicroscope (Leica S9E). Freehand sections of fertile structures were prepared, placed on glass slides, and mounted in either ultrapure water or lactophenol cotton blue solution for microscopic observation using a Leica DM2500 compound microscope equipped with a digital camera and measured using Leica imaging software. The photographic plates were prepared using Adobe Photoshop CC (2018). Fungal isolation was initiated on the day of collection to preclude contamination. A small mass of conidia from synnemata or sclerotia within insect hosts was transferred to potato dextrose agar (PDA) plates using a sterile needle (Wei et al. 2021). Cultures were incubated at 25 °C in the dark for 2 weeks until colonies reached 2–3 cm in diameter. Herbarium specimens were deposited in the Kunming Institute of Botany, Academia Sinica (HKAS), and Mae Fah Luang University (MFLU). Living cultures grown on PDA were deposited in the Kunming Institute of Botany Culture Collection (KUNCC) and the Mae Fah Luang University Culture Collection (MFLUCC). Index Fungorum (IF) numbers were registered following the protocol outlined by Jayasiri et al. (2015) and Index Fungorum (www.indexfungorum.org, accessed on 3 September 2025).
DNA extraction, amplification, and sequencing
Total genomic DNA was extracted from fresh mycelia grown on PDA medium and fungal tissues from dry specimens using a DNA extraction kit (Omega Fungus Genomic DNA Extraction Kit, China), following the manufacturer’s instructions. This study used ITS, nrSSU, nrLSU, tef1-α, rpb1, and rpb2. The ITS region was amplified with primers ITS5 and ITS4, while nrSSU was amplified using NS1 and NS4 (White et al. 1990). The nrLSU region was amplified with the primer pair LR0R and LR5 (Vilgalys and Hester 1990). The tef1-α gene was amplified with primers EF1-983F and EF1-2218R (Rehner and Buckley 2005). The rpb1 region was targeted using primers CRPB1A and RPB1Cr (Castlebury et al. 2004), while rpb2 was amplified with primers fRPB2-5f and fRPB2-7cR (Liu et al. 1999). The PCR amplification conditions for ITS, nrLSU, nrSSU, tef1-α, rpb1, and rpb2 followed a protocol consisting of (1) an initial denaturation at 94 °C for 3 min; (2) 33 cycles of denaturation at 94 °C for 30 s, followed by locus-specific annealing (ITS: 51 °C for 50 s; nrSSU: 47 °C for 1 min 20 s; nrLSU: 50 °C for 30 s; tef1-α: 58 °C for 50 s; rpb1 and rpb2: 51 °C for 40 s) and extension (ITS: 72 °C for 45 s; nrSSU and nrLSU: 72 °C for 1 min 50 s; tef1-α: 72 °C for 1 min; rpb1 and rpb2: 72 °C for 1 min 20 s); and (3) a final extension at 72 °C for 10 min. All PCR products were sequenced at Tsingke Biological Technology in Chongqing, China. The newly generated sequences in this study were deposited in GenBank (www.ncbi.nlm.nih.gov/genbank/) and used for the phylogenetic analysis. Information on all sequences used for the phylogenetic analysis is provided in Table 1.
Phylogenetic analysis
The forward and reverse reads generated in this study were assembled using BioEdit v.7.0.5.3, and the initial identification was performed by BLAST searches in GenBank. Based on the latest literature, updated sequence data were obtained from GenBank. The One-click Fungal Phylogenetic Tool (OFPT) framework (Zeng et al. 2023) was used to construct an initial comprehensive phylogenetic tree for Ophiocordycipitaceae and Polycephalomycetaceae, which was subsequently refined. Single-gene sequence alignments were conducted using the online program MAFFT v.7.110 (https://mafft.cbrc.jp/alignment/server/). TrimAl v1.2 (http://trimal.cgenomics.org) was used to remove uninformative gaps and ambiguous regions (Capella-Gutiérrez et al. 2009), and SequenceMatrix v.1.7.8 was used to concatenate the individual alignments (Vaidya et al. 2011). The final alignment was converted to NEXUS format using AliView v.1.28 (Larsson 2014). Maximum likelihood (ML) and Bayesian inference (BI) algorithms were used to perform phylogenetic analyses of the aligned sequences on the CIPRES Science Gateway portal (www.phylo.org) (Miller et al. 2010). Maximum likelihood analysis was performed using RAxML-HPC BlackBox with rapid bootstrap analysis, followed by 1,000 bootstrap replicates; the GTRGAMMA model was applied to all partitions. Bayesian analysis was used to evaluate posterior probabilities (PP) using MrBayes on XSEDE v.3.2.7a; six simultaneous Markov chains were run for 2 million generations, and trees were sampled every 200 generations (resulting in 10,000 trees). The analysis was terminated when convergence was achieved and the average standard deviation of split frequencies fell below 0.01. Phylogenetic trees were visualized using FigTree v.1.4.2 (Rambaut 2014) and edited using Adobe Illustrator CS6 (Adobe Systems Inc., USA). Maximum likelihood bootstrap values equal to or greater than 70% and PP values equal to or greater than 0.90 were shown above the nodes.
Genealogical concordance phylogenetic species recognition analysis
To assess recombination levels among closely related species, genealogical concordance phylogenetic species recognition (GCPSR) was applied using the pairwise homoplasy index (PHI) test (Bruen et al. 2006). Relationships among closely related taxa were visualized by constructing a phylogenetic network based on a concatenated dataset of five loci (ITS, nrSSU, nrLSU, tef1-α, and rpb1) using the LogDet transformation in SplitsTree v.4 (Huson and Bryant 2006). A Φw value ≤ 0.05 was interpreted as statistically significant evidence of recombination within the dataset. This analysis focused specifically on Polycephalomyces bannaensis due to unresolved phylogenetic relationships. In contrast, other species included in this study did not exhibit such ambiguity and were readily distinguishable using standard phylogenetic methods; therefore, additional recombination analyses were not required.
Results
Phylogenetic analyses
Phylogenetic analyses were conducted using sequence data from six loci (nrLSU, ITS, nrSSU, tef1-α, rpb1, and rpb2), representing 303 taxa from the families Ophiocordycipitaceae and Polycephalomycetaceae. Two strains of Cordyceps militaris (OSC 93623 and YFCC 6587) were used as outgroup taxa. The concatenated alignment consisted of 5,072 characters, including gaps, comprising 844 bp for nrLSU, 570 bp for ITS, 1,025 bp for nrSSU, 921 bp for tef1-α, 688 bp for rpb1, and 1,024 bp for rpb2. The best-scoring maximum likelihood tree, with a log-likelihood value of −128081.371, is presented in Fig. 1.
Phylogram generated from maximum likelihood analysis based on combined nrLSU, ITS, nrSSU, tef1-α, rpb1, and rpb2 sequence data. ML bootstrap values equal to or greater than 70% and PP values equal to or greater than 0.90 are shown above the nodes. Newly generated sequences are indicated in blue.
The phylogenetic analysis included eight genera of Ophiocordycipitaceae (Drechmeria, Hantamomyces, Harposporium, Ophiocordyceps, Paraisaria, Purpureocillium, Tolypocladium, and Torrubiellomyces), five genera of Polycephalomycetaceae (Dingleyomyces, Paradingleyomyces, Perennicordyceps, Pleurocordyceps, and Polycephalomyces), and one incertae sedis taxon (Cordyceps pleuricapitata) within Hypocreales. Ophiocordyceps was represented by nine distinct clades: O. acicularis, O. blattae, O. elongata, O. issidarum, O. ravenelii, O. sinensis, O. sphecocephala, O. sobolifera, and O. unilateralis. Ophiocordyceps northeastensissp. nov. is sister to O. stylophora (99% ML/0.98 PP; Fig. 1) and nests within the O. sinensis clade. Ophiocordyceps jinguangensissp. nov. is sister to a clade containing O. formosana, O. spicatus, and H. huangshanensis, with strong support (100% ML/1.00 PP; Fig. 1), and nests within the O. ravenelii clade. Ophiocordyceps formicarum is sister to a clade including O. irangiensis, O. sphecocephala, and O. vespulae (91% ML/0.99 PP; Fig. 1), grouping within the O. sphecocephala clade. Additionally, five hirsutella-like species (Polycephalomyces bannaensissp. nov., Po. chiangraiensissp. nov., Pleurocordyceps shibingensissp. nov., Pl. tengchongensissp. nov., and Dingleyomyces yunnanensissp. nov.) were confirmed as members of Polycephalomycetaceae. Collectively, hirsutella-like species within Ophiocordycipitaceae and Polycephalomycetaceae comprise 17 distinct clades. In contrast, the O. sphecocephala clade and the Torrubiellomyces clade, although present in the overall phylogram, do not belong to the hirsutella-like lineage as defined in this study. All clades are supported by high bootstrap values and Bayesian posterior probabilities (Fig. 1).
A pairwise homoplasy index (PHI) test was conducted using a five-gene dataset (ITS, nrSSU, nrLSU, tef1-α, and rpb1) to assess recombination levels among clades of Polycephalomyces bannaensissp. nov., Po. tengchongensis, Po. multiperitheciatae, Po. jinghongensis, and Po. formosus. No significant recombination events were detected among these groups (Φw > 0.05), indicating genetic isolation and supporting their recognition as distinct species (Fig. 2).
Split graphs showing results of the pairwise homoplasy index (PHI) test of the new taxa and closely related taxa using LogDet transformation and splits decomposition.
Taxonomy
Ophiocordycipitaceae
Taxon classificationAnimaliaOphiocordycipitaceae
G.H. Sung, J.M. Sung, Hywel-Jones & Spatafora, 2007.
0D3DB5B5-A52A-57D7-8F70-9B9C1A950B89
Ophiocordyceps Petch, 1931.
Notes.
Ophiocordyceps (Ophiocordycipitaceae) is a genus of entomopathogenic fungi encompassing over 350 species (Hyde et al. 2024; Index Fungorum 2025). This genus was originally treated as a subgenus under Cordyceps by Petch (1931) until it was redefined as a distinct genus through multigene phylogenetic analyses by Sung et al. (2007). Species in this genus exhibit diagnostically dark, fibrous to wiry stromata with variable morphologies (clavate, filiform, or branched) and perithecia that are either superficially exposed or embedded within the stromatal matrix. The asci are typically cylindrical and bear prominent apical caps, while the ascospores are generally cylindrical, multiseptate, and may either disarticulate into part-spores or remain intact after discharge (Sung et al. 2007; Ban et al. 2015). Host specificity spans diverse arthropods, parasitizing larvae of Lepidoptera, Coleoptera, Hymenoptera, Hemiptera, Diptera, and arachnids (Kobayasi 1941; Mains 1958; Ban et al. 2015). Quandt et al. (2014) present a concise, thorough, phylogenetically relevant, and taxonomically accurate revision of the family Ophiocordycipitaceae. In this study, we introduce two new species (O. jinguangensis and O. northeastensis) based on phylogenetic and morphological analyses.
Ophiocordyceps
jinguangensis
Taxon classificationAnimaliaOphiocordycipitaceae
X. Zhang, C.J.Y. Li, K.D. Hyde & T.C. Wen sp. nov.
6129B075-B321-57E1-9E06-45220471AFFD
Index Fungorum: IF904199
Etymology.
Reference to the Jinguang Temple Nature Reserve, Yongping County, Dali City, the locality where the type specimen was collected.
Type.
CHINA • Yunnan Province, Dali City, Yongping County, Jinguang Temple Nature Reserve, 25°15'54.11"N, 99°32'13.27"E, alt. 2280 m, 25 July 2024, Cui-Jin-Yi Li, DD247162-1 (holotype: HKAS 149981)
Description.
Parasitic on a larva of Tenebrionoidea (Coleoptera), 37.4 × 2.8–5 mm, yellowish-brown. Sexual morph: Stroma arising from the abdomen and tail of the host, solitary, unbranched, yellow to brown, 10.2–16.5 × 1–2.2 mm. Fertile part up to 3.4–7.1 × 1.2–2.2 mm, cylindrical, yellow to brown, single, rough, with ostiole. Stipe clavate, light brown to brown, the color gradually becoming dark towards the apex, 7.8 × 1.0–1.2 mm. Perithecia 306–496 × 134–223 μm (x̄ = 388.4 × 175.9 µm, n = 15), immersed, ovoid to oblong-ovate. Asci 153.6–273.2 × 5.2–11.8 μm (x̄ = 234.7 × 8.4 µm, n = 20), cylindrical, hyaline, with thickened apex. Apical cap 2.5–5 × 3.5–5.6 μm (x̄ = 3.6 × 4.7 µm, n = 20), hyaline, hemispherical. Ascospores 133.4–187.6 × 1.4–2.4 µm (x̄ = 164.3 × 1.9 µm, n = 15), filiform, hyaline, easily breaking into part-spores. Secondary ascospores 6.7–9.6 × 1.6–2.7 μm (x̄ = 8.1 × 2.2 µm, n = 20), cylindrical, smooth-walled. Asexual morph: undetermined.
Ophiocordyceps jinguangensis (HKAS 149981, holotype). A, B stroma arising from host; C, D fertile part; E perithecia; F immature asci of different lengths; G–I asci; J, K ascospores; L, M secondary ascospores. Scale bars: 1 cm (A); 0.5 cm (B); 0.1 cm (C, D); 100 µm (E, F); 20 µm (G–K); 10 µm (L, M).
Habitat and distribution.
On dead larva of Tenebrionoidea (Coleoptera); currently known from southwestern China.
Additional specimens examined.
CHINA • Yunnan Province, Dali City, Yongping County, Jinguang Temple Nature Reserve, 25°15'54.11"N, 99°32'13.27"E, alt. 2280 m, 25 July 2024, Cui-Jin-Yi Li, DD247162-2 (paratype: HKAS 149982).
Notes.
The newly described species exhibits morphological characteristics closely resembling those of O. formosana (Kobayasi and Shimizu 1980; Li et al. 2016) and O. spicatus (Zha et al. 2021), particularly in sharing features such as yellow to brown, unbranched fertile heads, cylindrical asci with the central portion slightly wider than the apical and basal regions, and filiform ascospores. Phylogenetic analysis places O. jinguangensis as a sister lineage to O. formosana and O. spicatus, with 100% ML/1 PP support (Fig. 1). However, O. jinguangensis is readily distinguished by its stroma lacking the vivid orange to yellow-orange pigmentation characteristic of O. formosana and O. spicatus. It further differs from O. formosana in having shorter asci (153.6–273.2 × 5.2–11.8 µm vs. 366–498 × 8–11 µm) and longer secondary ascospores (6.7–9.6 × 1.6–2.7 µm vs. 2–6 × 1–3 µm). The combined evidence from morphological characteristics and phylogenetic analyses confirms the taxonomic distinction of O. jinguangensis, thereby justifying its designation as a novel species.
Ophiocordyceps
northeastensis
Taxon classificationAnimaliaOphiocordycipitaceae
X. Zhang, K.D. Hyde & T.C. Wen sp. nov.
00B0F3F1-D451-50B2-AF12-DE92CA74819C
Index Fungorum: IF904200
Etymology.
Referring to Lushuihe Village, Fusong County, Baishan City, Jilin Province, China, the locality belongs to Northeast China, where the type specimen was collected.
Type.
CHINA • Jilin Province, Baishan City, Fusong County, Lushuihe Village, 42°50'33.25"N, 127°78'23.05"E, 15 November 2024, Ting-Chi Wen, DB3 (holotype: HKAS 149974)
Description.
Parasitic on larvae of Elateridae (Coleoptera). Sexual morph: undetermined. Asexual morph: Hyphomycetous. Primary synnemata emerging from the junction between head and tail of host, 36.6–64.5 × 0.5–0.9 mm. Secondary synnemata 4.6–25.5 mm in length, 0.7–1.5 mm in width, arising from primary synnemata, solitary, white, becoming brown with age, cylindrical, tapering gradually toward the apex. Phialides 22.3–29.0 × 1.0–4.5 µm (x̄ = 26.3 × 2.4 µm, n = 30), with swollen base and slender neck, hyaline, directly produced on superficial hyphae of secondary branches. Conidia 5.3–9.4 × 2.1–4.9 µm (x̄ = 7.6 × 3.8 µm, n = 30), unicellular, hyaline, oblong-ellipsoid to obovoid.
Ophiocordyceps northeastensis (HKAS 149974, holotype). A habitat of O. northeastensis; B, C synnemata arising from the host; D–F synnemata; G–I phialides; J, K conidia. Scale bars: 1.0 cm (B, C); 0.2 cm (D); 0.1 cm (E, F); 20 µm (G–K).
Habitat and distribution.
On larvae of Elateridae (Coleoptera) from rotten wood; currently known from northeast China.
Additional specimens examined.
CHINA • Jilin Province, Baishan City, Fusong County, Lushuihe Village, 42°50'33.25"N, 127°78'23.05"E, 15 November 2024, Ting-Chi Wen, DB2 (paratype: HKAS 149973).
Notes.
The newly proposed species, O. northeastensis, has only been observed in its asexual morph, while its sister lineage, O. stylophora, has been reported exclusively in its sexual morph, making it impossible to compare their morphological differences. They are proposed as distinct species based on molecular phylogenetic relationships. The new species (DB2203) differs from O. stylophora (OSC 111000) by 35/581 bp (6%, 9 gaps) in ITS, 3/817 bp (0.4%) in nrLSU, 389/1343 bp (28.9%, 388 gaps) in nrSSU, and 33/780 bp (4.2%) in tef1-α.
Polycephalomycetaceae
Taxon classificationAnimaliaPolycephalomycetaceae
Y.P. Xiao, Y.B. Wang, T.C. Wen, H. Yu & K.D. Hyde, 2023.
05C27D27-F091-530F-9C14-011713CA5618
Polycephalomyces
Notes.
Polycephalomyces is a genus established by Kobayasi (1941), with Po. formosus as its type species. Kepler et al. (2013) reclassified several species into Polycephalomyces based on molecular phylogenetic analyses, confirming its placement within Ophiocordycipitaceae under the “One Fungus One Name” principle. This principle unifies the taxonomies of sexual and asexual morphs (Hawksworth et al. 2011; Kepler et al. 2013; Quandt et al. 2014). Matočec et al. (2014) proposed a new genus, Perennicordyceps, for four Polycephalomyces species based on their distinct morphology, including superficial perithecia and hirsutella- or acremonium-like anamorphs. Xiao et al. (2023) transferred Polycephalomyces to Polycephalomycetaceae based on morphological and phylogenetic analyses. Species within this genus exhibit a broad host range, parasitizing insects and other fungi (e.g., Ophiocordyceps spp. and Elaphomyces spp.), highlighting their ecological versatility (Bischoff et al. 2005; Van Vooren and Audibert 2005; Ban et al. 2009; Wang et al. 2012; Matočec et al. 2014; Wang et al. 2015a, 2015b; Xiao et al. 2018; Sun et al. 2019; Wang et al. 2021; Xiao et al. 2023). In this study, we describe two new species of Polycephalomyces, further expanding the known diversity of this ecologically and taxonomically significant genus.
Polycephalomyces
bannaensis
Taxon classificationAnimaliaOphiocordycipitaceae
X. Zhang, N.Y. Liu, K.D. Hyde & T.C. Wen sp. nov.
5D9CA861-AC55-54FD-959C-5687924066F0
Index Fungorum: IF904201
Etymology.
Reference to the Mengzhe Village, Menghai County, Xishuangbanna City, the locality where the type specimen was collected.
Type.
CHINA • Yunnan Province, Dai autonomous prefecture of Xishuangbanna, Menghai County, Mangunxiazhai Town, 21°58'32.33"N, 100°23'5.03"E, alt. 1237 m, 10 August 2024, Nan-Yi Liu, BN24081002-1 (holotype: HKAS 149983).
Description.
Parasitic on adult of Coleoptera. Sexual morph: undetermined. Asexual morph: Hyphomycetous. Synnemata 9–33 mm in length and 0.7–1.3 mm in width, occurring either scattered or clustered on the stipe, branched, white, cylindrical, with an enlarged globose fertile head at the apex. Fertile heads 0.1–0.8 mm in width, globose to subglobose, white, covered with conidial mass. Conidiophores 15–25 µm long (x̄ = 18.8 µm, n = 30), predominantly concentrated within the fertile head, monothetic, occurring either solitarily or in acropleurogenous whorls, and bear 1–4 phialides. Phialides 10–18.9 × 0.9–2.2 µm (x̄ = 13.8 × 1.5 µm, n = 30), narrowly cylindrical, hyaline, smooth-walled, tapering gradually from the middle to the apex. Conidia 2.4–3.3 × 1.5–2.1 µm (x̄ = 2.9 × 1.8 µm, n = 30), unicellular, oblong-ellipsoid to obovoid, hyaline.
Polycephalomyces bannaensis (HKAS 149983, holotype). A habitat of Polycephalomyces bannaensis; B overview of host; C–E conidiophores, phialides, and conidia; F, G conidia. Scale bars: 1 cm (B); 20 µm (C); 10 µm (D, E); 5 µm (F, G).
Habitat and distribution.
On insect of Coleoptera currently known from southwestern China.
Additional specimens examined.
CHINA • Yunnan Province, Dai autonomous prefecture of Xishuangbanna, Menghai County, Mangunxiazhai Town, 21°58'32.33"N, 100°23'5.03"E, alt. 1237 m, 10 August 2024, Xing-Can Peng, BN24081002-2 (paratype: HKAS 149954).
Notes.
Phylogenetically, Polycephalomyces bannaensis is sister to Po. tengchongensis (97% ML/1 PP, Fig. 1). Polycephalomyces bannaensis is distinct in having longer synnemata (9–33 mm long) and oblong-ellipsoid to obovoid conidia, while Po. tengchongensis has shorter synnemata (18.7 mm long) and globose conidia. Additionally, P. bannaensis parasitizes coleopteran insects, whereas Po. tengchongensis infects fungi (e.g., Perennicordyceps cf. elaphomyceticola). The PHI test results (Fig. 2) revealed no significant recombination relationships between Po. bannaensis and its phylogenetically related taxa (Φw = 0.2105). Based on molecular phylogenetic analyses and morphological observations, we propose that these two species are distinct from each other and introduce our collections as a new species.
Polycephalomyces
chiangraiensis
Taxon classificationAnimaliaOphiocordycipitaceae
X. Zhang, K.D. Hyde & T.C. Wen sp. nov.
3C1904F1-45CF-54B5-8022-5D8910129A3C
Index Fungorum: IF904830
Etymology.
Reference to Mueang Chiang Rai District, Chiang Rai Province, the locality where the type specimen was collected.
Type.
THAILAND • Chiang Rai Province, Mueang Chiang Rai District, 20°02'48.30"N, 99°49'31.51"E, alt. 410 m, 28 October 2023, Xian Zhang, TX4-1 (holotype: MFLU 26-0001; ex-type culture: MFLUCC 25-0386).
Description.
Parasitic on Ophiocordyceps sp. (Ophiocordycipitaceae, Hypocreales). Sexual morph: Undetermined. Asexual morph: Hyphomycetous. Colonies on PDA growing slowly, attaining a diameter of 1.4–1.9 cm in 21 days at 25 °C, powdery, vary in color from yellow to white, reverse yellow to brown. Conidiophores normally with 1–2 phialides. Phialides 8.2–19.8 (x̄ = 15.2 µm, n = 10) long, base 1.6–2.2 μm wide, neck 0.6–0.7 μm wide, hyaline, smooth, narrowly subulate with a neck. Conidia 2–3.2 × 0.9–1.5 µm (x̄ = 2.5 × 1.1 µm, n = 30), one-celled, hyaline, ellipsoid.
Polycephalomyces chiangraiensis (MFLU 26-0001, holotype). A overview of Polycephalomyces chiangraiensis; B overview of insect; C, D culture on PDA from obverse and reverse; E–H phialides with conidia; I, J conidia. Scale bars: 10 mm (A); 2 mm (B); 1 cm (C, D); 10 µm (E–J).
Habitat and distribution.
On Ophiocordyceps sp., currently known from northern Thailand.
Notes.
The newly proposed species, Po. chiangraiensis, forms a sister lineage to Po. myrmecophilus with 100% ML/1 PP support (Fig. 1). Morphologically, Po. chiangraiensis differs from Po. myrmecophilus by having a more pronounced difference in width between the base and apex of the phialides and ellipsoid spores. Based on these phylogenetic and morphological distinctions, Po. chiangraiensis is proposed as a novel species within the genus Polycephalomyces.
Pleurocordyceps
Taxon classificationAnimaliaOphiocordycipitaceae
Y.J. Yao, Y. H. Wang, S. Ban, W.J. Wang, Y. Li, K. Wang & P.M. Kirk, 2021.
9E5BB7BD-843D-51EC-B1CF-6A6823916789
Notes.
Pleurocordyceps is a genus of entomopathogenic fungi within the family Polycephalomycetaceae, comprising 23 species (Index Fungorum 2025). This genus was established by Wang et al. (2021) to accommodate species previously classified under Polycephalomyces and Cordyceps, with Pleurocordyceps sinensis (synonym: Paecilomyces sinensis) designated as the type species. Alongside Polycephalomyces and Perennicordyceps, it forms a well-supported monophyletic clade within Hypocreales. Xiao et al. (2023) further clarified its taxonomic placement by formally assigning Pleurocordyceps to the newly established family Polycephalomycetaceae, thereby stabilizing its systematic position within Hypocreales. The sexual morph is characterized by lateral, pulvinate stromata near the host surface, bearing perithecia close to the apex, while the asexual morph produces two distinct conidial types in culture (Wang et al. 2021). Herein, we introduce Pl. shibingensis and Pl. tengchongensis as two new species.
Pleurocordyceps
shibingensis
Taxon classificationAnimaliaOphiocordycipitaceae
X. Zhang, K.D. Hyde & T.C. Wen sp. nov.
A254D637-A2D1-51C5-84C0-C88201D96087
Index Fungorum: IF904202
Etymology.
Reference to Shibing County, Guizhou Province, the locality where the type specimen was collected.
Type.
CHINA • Guizhou Province, Shibing County, 27°09'03.55"N, 108°15'20.08"E, 22 April 2022, Xian Zhang, SB2206 (holotype: HKAS 149965; ex-holotype culture GACP SB2206).
Description.
Parasitic on larva of Scarabaeoidea (Coleoptera), 2.9–4 × 0.5–0.9 cm. Sexual morph: undetermined. Asexual morph: Hyphomycetous. Stroma arising from the head or abdomen of insect, 3–10 × 0.15–1.2 cm, solitary to multiple, flexuous, cylindrical to obclavate, unbranched, yellowish to off-white. Fertile part erects, subglobose, off-white, 1.2 × 0.8 cm. Stipe flexuous, yellowish, 2.3–95 × 0.15–0.25 cm, cylindrical. Conidiophores 28.1–43.7 μm (x̄ = 36.1 µm, n = 20) in length, predominantly concentrated on the fertile head, and bear 1–6 phialides. Phialides 13.3–21.9 × 1.1–1.9 μm (x̄ = 17.1 × 1.4 µm, n = 30), slender, cylindrical, hyaline, smooth-walled, arising from irregularly branched or quasi-verticillate conidiophores. Conidia 2.8–3.6 × 2.2–2.6 µm (x̄ = 3.2 × 2.5 µm, n = 30), one-celled, obovoid, hyaline, smooth-walled, guttulate, and of a single type.
Culture characteristics: Colonies on PDA, attaining a diameter of 23–25 mm within 14 d at 25 °C, dense, leathery, and vary in color from brown to white, thin, in the middle of the white part grew water droplets, reverse yellow with yellowish pigment. Conidiophores 26–35.8 μm long (x̄ = 30.1 µm, n = 20), concentrated, normally with 2 phialides, rarely with 6 phialides. Phialides 11.6–19.7 × 0.8–2 μm (x̄ = 15.2 × 1.4 µm, n = 30), hyaline, smooth-walled, elongated lageniform, tapering gradually from the base to the apex. Conidia 1.5–2.4 × 1–1.7 μm (x̄ = 1.8 × 1.3 µm, n = 30), one-celled, hyaline, ellipse to obovoid.
Pleurocordyceps shibingensis (HKAS 149965, holotype). A overview of host; B synnemata; C, J conidiophores; D, E, K, L phialides; F, G, M, N conidia; H, I culture on PDA from above and below. Scale bars: 1 cm (A, B); 20 µm (C); 10 µm (D, E, J–N); 5 µm (F, G); 1 cm (H, I). (C–G from specimen and J–N from culture.)
Habitat and distribution.
On dead larva of an insect (Scarabaeoidea, Coleoptera); currently known from southwestern China.
Additional specimens examined.
CHINA • Guizhou Province, Shibing County, 27°09'03.55"N, 108°15'20.08"E, 22 April 2022, Xian Zhang, SB2208 (paratype: HKAS 149966).
Notes.
Although both ITS and nrLSU sequences for four previously unpublished Pleurocordyceps strains (NBRC 109987, NBRC 109988, NBRC 109990, and NBRC 110224) are currently available in GenBank, Liu et al. (2024) used only the nrLSU sequences in their phylogenetic analysis, which showed that the strains clustered with Pl. litangensis. We reanalyzed them in this study using both loci. Our results show that Pleurocordyceps sp. NBRC 109987 and NBRC 109988 clustered with Pl. shibingensis in a well-supported clade (100% ML/0.99 PP, Fig. 1), while Pl. litangensis formed an independent branch (100% ML/1 PP, Fig. 1). Morphologically, Pl. shibingensis differs from Pl. litangensis by possessing whorled phialides and obovoid spores. Based on these distinct phylogenetic and morphological characteristics, Pl. shibingensis is proposed as a novel species within Pleurocordyceps.
Pleurocordyceps
tengchongensis
Taxon classificationAnimaliaOphiocordycipitaceae
X. Zhang, K.D. Hyde & T.C. Wen sp. nov.
B060DFFE-21AF-50BC-B6E3-CBB1383CE277
Index Fungorum: IF904203
Etymology.
Reference to Tengchong City, Yunnan Province, the locality where the type specimen was collected.
Type.
CHINA • Yunnan Province, Tengchong City, Houqiao Town, 25°11'30.63"N, 98°15'23.38"E, alt. 2152 m, 18 October 2024, Xian Zhang, HQ5 (holotype: HKAS 149971).
Description.
Parasitic on a larva of Lasiocampidae (Lepidoptera), 37.4 × 4.0–7.6 mm, yellowish-brown. Sexual morph: undetermined. Asexual morph: Synnemata 3.9–5.3 mm long, 0.3–0.6 mm wide, multiple, branched, cylindrical, clavate, capitate, stipitate, with fertile head at the apex, on the stromata of a larva of Lasiocampidae. Conidiophores 20.5–25.5 μm (x̄ = 23.2 µm, n = 15), erect, arising from hyphae, bearing phialides aggregates or clusters, often additional branches occur which are equal in length form stipe structures, 1–4 phialides. Phialides 13.2–16.2 × 0.8–2.1 μm (x̄ = 14.3 × 1.3 µm, n = 30), gathered on the tip of synnema, slender, elongated lageniform, tapering gradually from the base to the apex, hyaline, smooth-walled. Conidia 2.3–3.3 × 1–1.7 µm (x̄ = 2.9 × 1.5 µm, n = 30), one-celled, hyaline, smooth-walled, oval.
Pleurocordyceps tengchongensis (HKAS 149971, holotype). A habitat of Pleurocordyceps tengchongensis; B overview of Pl. tengchongensis; C overview of host; D host with synnemata; E conidiophores; F phialides; G, H conidia. Scale bars: 0.5 cm (C, D); 10 µm (E–H).
Habitat and distribution.
On dead larva of Lasiocampidae (Lepidoptera); currently known from southwestern China.
Additional specimens examined.
CHINA • Yunnan Province, Tengchong City, Houqiao Town, 25°11'30.63"N, 98°15'23.38"E, alt. 2152 m, 18 October 2024, Xian Zhang, HQ6 (paratype: HKAS 149972).
Notes.
The newly proposed species, Pl. tengchongensis, forms a sister lineage to Pl. heilongtanensis with 100% ML/0.98 PP support (Fig. 1). Morphologically, Pl. tengchongensis differs from Pl. heilongtanensis in having hirsutella-like phialides with a rapidly tapering neck (Xiao et al. 2023). Based on these phylogenetic and morphological distinctions, Pl. tengchongensis is proposed as a novel species within the genus Pleurocordyceps.
Dingleyomyces
Taxon classificationAnimaliaPolycephalomycetaceae
P.R. Johnst. & D.C. Park 2023.
979BC71B-8A50-54F6-8830-4EC4FDB78824
Notes.
Dingleyomyces lloydii (synonym: Ophionectria lloydii) is an endemic New Zealand fungicolous fungus that parasitizes the stromata of O. robertsii and O. hauturu. Initially described invalidly as Ophionectria cordyceps, it was later validly published as Ophionectria lloydii (Mains 1958) and subsequently recombined as Torrubiella lloydii (Rossman 1977). However, molecular phylogenetic analyses revealed Torrubiella to be polyphyletic, leading to its taxonomic abandonment (Kepler et al. 2017). Johnston et al. (2023) established a new genus, Dingleyomyces, to accommodate this species based on integrative morphological and phylogenetic evidence. The type species is characterized by small perithecial clusters forming on reduced, non-stipitate stromata. The asexual morph is characterized by conidiophores arising from hyphae, typically bearing two phialides with swollen bases and tapering necks. Conidia are cylindric, straight, and aseptate. In this study, we introduce Dingleyomyces yunnanensis as a new species.
Dingleyomyces
yunnanensis
Taxon classificationAnimaliaPolycephalomycetaceae
X. Zhang, K.D. Hyde & T.C. Wen sp. nov.
B5FE37F0-AF84-5103-84A4-1FD8CE10A283
Index Fungorum: IF904204
Etymology.
Reference to Yunnan Province, the locality where the type specimen was collected.
Type.
CHINA • Yunnan Province, Tengchong City, Houqiao Town, 25°11'30.63"N, 98°15'23.38"E, alt. 2152 m, 18 October 2024, Xian Zhang, HQ4 (holotype: HKAS 149970).
Description.
Parasitic on Ophiocordyceps cf. globiceps (Ophiocordycipitaceae, Hypocreales). Sexual morph: undetermined. Asexual morph: Synnemata 5.7–8.6 mm long, 0.3–0.5 mm wide, cylindrical, clavate, partial enlargement at the apex, stipitate, growing up from the head and tail of insect, multiple, white***. Stipes*** 5.1–7.9 mm long, 0.3–0.5 mm wide, cylindrical, white. Conidiophores concentrated, mono-terverticillate, and having two kinds of phialides. Phialides two types, both types observed on the same synnema. α-phialides 9.2–18.5 × 1.2–1.9 μm (x̄ = 13.4 × 1.6 μm, n = 30), hyaline, smooth, caespitose, palisade-like, crowed, gathered in the top of synnema. β-phialides 23.5–43.6 × 0.7–2.5 μm (x̄ = 32.4 × 1.7 μm, n = 30), hyaline, smooth, solitary, lanceolate, tapering into a long neck. Conidia 3–4.1 × 1.4–2.1 µm (x̄ = 3.6 × 1.7 µm, n = 30), one-celled, hyaline, smooth-walled, fusiform.
Dingleyomyces yunnanensis (HKAS 149970, holotype). A habitat of D. yunnanensis; B overview of D. yunnanensis; C overview of host; D, E synnemata; F–H conidiophores; I α-phialides; J β-phialides; K, L conidia. Scale bars: 0.2 cm (B, C); 0.1 cm (D, E); 100 µm (F); 20 µm (G, H); 10 µm (I–L).
Habitat and distribution.
On Ophiocordyceps cf. globiceps (Ophiocordycipitaceae, Hypocreales); currently known from southwestern China.
Additional specimens examined.
CHINA • Yunnan Province, Tengchong City, Houqiao Town, 25°11'30.63"N, 98°15'23.38"E, alt. 2152 m, 18 October 2024, Xian Zhang, HQ3 (paratype: HKAS 149969).
Notes.
The newly proposed species, D. yunnanensis, is a previously undescribed hirsutella-like species that forms a sister lineage to D. lloydii (the only currently recognized species in Dingleyomyces) with strong statistical support (100% ML/1 PP, Fig. 1). Morphologically, D. yunnanensis differs from D. lloydii in phialide shape and conidial morphology: D. lloydii has pyriform phialides and cylindrical conidia, whereas D. yunnanensis possesses palisade-like to lanceolate phialides and fusiform conidia. Critically, this study provides the first documentation of the asexual morph in Dingleyomyces. Based on these phylogenetic and morphological distinctions, D. yunnanensis is proposed as a novel species within Dingleyomyces.
Discussion
Hirsutella-like morphologies are not only found in Ophiocordyceps but also occur in other genera of Ophiocordycipitaceae, Polycephalomycetaceae, and Clavicipitaceae, complicating the morphological delimitation of Hirsutella. This study re-evaluates clades bearing hirsutella-like anamorphs within Ophiocordycipitaceae and Polycephalomycetaceae by: (1) mapping the morphological diversity of hirsutella-like anamorphs onto phylogenetic trees, and (2) analyzing their host associations and ecological diversification patterns (Fig. 1).
Building upon this phylogenetic framework, we provide a detailed morphological re-examination of 17 clades possessing hirsutella-like anamorphs. The following sections describe key characteristics for each clade, such as phialide morphology, conidial features, and host affiliations. In addition, the Discussion integrates observations from the newly described species and evaluates their significance within their respective clades, including their morphological conformity or deviation, host associations, and phylogenetic placement. These reflections provide insights into patterns of character evolution, clade stability, and the evolutionary relevance of hirsutella-like morphologies, thereby clarifying taxonomic boundaries and evolutionary relationships among these taxa.
Ophiocordyceps
sinensis clade
The O. sinensis clade contains the medically significant Ophiocordyceps sinensis (Sung et al. 2007) and its asexual morph, Hirsutella sinensis (Liu et al. 1989). Phialides in this clade feature cylindrical, slender, or subulate bases that taper gradually or suddenly into distinct necks. Several species, including H. illustris, H. strigosa, H. rhossiliensis, O. sinocampes, and O. nujiangensis, display distinctive warty protrusions on their phialides. Notably, the newly described species O. northeastensis also exhibits conspicuous warty ornamentation on the phialides, further supporting its placement within the O. sinensis clade. Additionally, multiple species produce spores embedded in mucous sheaths, such as H. sinensis, H. illustris, H. strigosa, H. rhossiliensis, O. liangshanensis, and O. multiperitheciata. This clade exhibits remarkably broad host specificity (Simmons et al. 2015), parasitizing diverse organisms ranging from nematodes and mites to insects belonging to Hemiptera, Coleoptera, and Lepidoptera. From an evolutionary perspective, the recurrence of warty phialide ornamentation and mucous-sheathed conidia across multiple lineages within the O. sinensis clade suggests that these characters are evolutionarily conserved and may represent stable diagnostic traits. The broad host spectrum observed in this clade, in contrast, indicates that host shifts and ecological diversification have occurred without substantial modification of key asexual morphological features.
Ophiocordyceps
issidarum clade
The O. issidarum clade, originally named Hirsutella guyana by Simmons et al. (2015), consists of three species (H. guyana, H. haptospora, and H. versicolor). The phialides in this clade are characterized by a cylindrical base that abruptly narrows into a thin neck. Some species, such as H. guyana, O. flavida, and O. brunneinigra, exhibit branched phialides. Xie et al. (2025) later assigned this clade the name O. issidarum, as its sexual forms produce superficial perithecia along with filamentous, multiseptate ascospores that remain intact at maturity (Hyde et al. 2017; Luangsa-ard et al. 2018). Most members of this clade parasitize hosts in the order Hemiptera. However, H. haptospora infects insects in the order Diptera, while H. kirchneri parasitizes insects in the order Trombidiformes.
Ophiocordyceps
acicularis clade
The O. acicularis clade, originally described as Hirsutella nodulosa by Simmons et al. (2015), exhibits monophialidic conidiogenous cells, occasionally polyphialidic, arising laterally or terminally from hyphae. These conidiogenous cells are hyaline, smooth-walled, and taper gradually or abruptly into a slender, short neck. The phialide bases on specimens are comparatively shorter than those in cultured specimens, with the necks exhibiting helical curvature and occasionally bearing verrucose ornamentation (Tasanathai et al. 2020). In contrast, the phialides of O. unituberculata are distinguished by a distinctive apical spherical protrusion (Wang et al. 2018). This clade comprises many cryptic species that parasitize lepidopteran larvae, except for H. leigongshanensis (Tasanathai et al. 2020), which infects coleopteran larvae.
Ophiocordyceps
blattae clade
The O. blattae clade, described as Hirsutella thompsonii by Simmons et al. (2015), is characterized by a more swollen phialide base and a shorter neck compared to other hirsutella-like clades. Its conidia range from spherical to fusiform and exhibit either a warty surface or a mucous sheath. Members of this clade predominantly parasitize cockroaches and termites (Blattodea). While O. communis in this clade exhibits an asexual morph with intermediate characteristics between Hymenostilbe and Hirsutella (Sung et al. 2007), its close relative O. neocommunis was recently reported to show morphological similarity, with the closest genetic distance being classified as a hirsutella-like asexual form (Yang et al. 2025).
Ophiocordyceps
unilateralis clade
The O. unilateralis clade represents one of the most prevalent groups of ant-associated entomopathogenic fungi in tropical forests globally (Evans et al. 2011). This clade comprises three morphologically and ecologically distinct clades: the O. unilateralis core clade, the O. kniphofioides subclade, and the O. oecophyllae subclade (Araújo et al. 2018). The O. kniphofioides subclade is characterized by orange ascomata developing on thorax-emerging stromata with 360° fertile coverage, typically found on hosts in Amazonian moss carpets. In contrast, the O. unilateralis core clade forms dorsally pronotal stromata bearing laterally attached brown to black ascomata, consistent with its “unilateralis” epithet. Ophiocordyceps oecophyllae differs in producing only an asexual morph, with phialides developing directly on the host, particularly from joint regions. Molecular data further confirm that this subclade is well supported (Araújo et al. 2018). Phialides across the O. unilateralis clade exhibit distinct hirsutella-like morphotypes (Evans et al. 2011), characterized by swollen bases that taper either gradually or abruptly into elongated necks, longer than those observed in other clades. While some species display only a single phialide type, others produce polymorphic phialides.
Ophiocordyceps
elongata clade
Species in this clade are associated with diverse lepidopteran and hemipteran insects and are characterized by richly branched stromata, subulate phialides, and conidia enveloped by a mucous sheath (Mongkolsamrit et al. 2024).
Ophiocordyceps
sobolifera clade
This clade includes the O. sobolifera core clade and the O. brunneipunctata complex subclade, comprising pathogens associated with cicada nymphs as well as coleopteran larvae (Mongkolsamrit et al. 2024). Members of the O. sobolifera clade typically produce stromata ranging from reddish-brown to pale yellow, cylindrical ascomata, and immersed perithecia. The asexual forms in this clade feature phialides of variable morphology—some with a spherical base that abruptly narrows into a long filamentous neck (and may branch), while others lack such an elongated neck. The conidia are subglobose to fusoid in shape (Zou et al. 2022; Mongkolsamrit et al. 2024).
Ophiocordyceps
ravenelii clade
This clade comprises taxa that are pathogenic to Coleoptera and is characterized by yellow, orange, or brown stromata, with immersed perithecia located at apical or lateral fertile regions, and filamentous, multiseptate ascospores that disarticulate into cylindrical secondary ascospores upon maturation (Wang et al. 2015c; Mongkolsamrit et al. 2024). The asexual morph is characterized by phialides occurring singly, in opposed pairs, or in verticils, with subtly inflated bases tapering distally into slender necks (Li et al. 2005). Mucous sheaths are commonly absent in species of this clade (Wang et al. 2015c). Although the asexual morph of O. jinguangensis was not observed in the present study, its association with coleopteran hosts and the diagnostic features of its sexual morph are congruent with the morphological circumscription of this clade. Together with its well-supported placement in the phylogenetic analyses, these lines of evidence collectively support the inclusion of O. jinguangensis within this lineage.
Paraisaria clade
The Paraisaria clade has a broad host range (Mongkolsamrit et al. 2019), infecting hosts across Hemiptera, Coleoptera, Diptera, Lepidoptera, Orthoptera, Blattodea, and Hymenoptera. Mongkolsamrit et al. (2019) supported its classification as a valid genus within Ophiocordycipitaceae due to its unique morphological features. Paraisaria is distinctive in producing spherical to subspherical fertile heads. Its asexual stage is linked with hirsutella-like anamorphs. The synnemata comprise verticillately branched conidiophores bearing phialides with swollen bases and thin, tapering necks. Some phialides are borne singly on vegetative hyphae. Conidia are cylindrical to fusiform and aggregated in slimy heads.
Drechmeria clade
The Drechmeria clade is known as endoparasites of nematodes and lepidopteran larvae (David and Jeffrey 1999; Spatafora et al. 2015). However, Drechmeria panacis was isolated as an endophyte from Panax notoginseng (Yu et al. 2018), suggesting broader host-range plasticity and functional diversity within the genus. Morphologically, this clade exhibits hirsutella-like morphology characterized by conidiophores bearing verticillate or solitary phialides. The phialides display a distinct swelling near the base that tapers toward the neck. Conidial morphology is variable, ranging from balanoid, cylindrical, and subglobose to irregular shapes (Zare and Gams 2001).
Harposporium clade
The Harposporium clade predominantly parasitizes invertebrate nematodes, with a minority of isolates derived from coleopteran insects (Kuo et al. 2008; Chen et al. 2025). This fungal group displays two discrete asexual morphologies: (1) spherical to subglobose conidiogenous cells generating arcuate conidia, frequently accompanied by accessory conidia, arthroconidia, and chlamydospores (Evans and Whitehead 2005); and (2) a hirsutella-like morphology characterized by phialides with swollen bases that taper either gradually or abruptly into well-defined necks, bearing elliptical conidia at their apices—as observed in H. peltatum, H. cerberi, and H. anguillulae (Hodge 1997; Evans and Whitehead 2005).
Purpureocillium clade
The Purpureocillium clade exhibits a broad ecological distribution in terrestrial and marine environments, having been isolated from diverse insect hosts, nematodes, crop rhizospheres, soil, and even a corneal ulcer patient (Giné and Sorribas 2017; Liu et al. 2019; Calvillo-Medina et al. 2021; Yang et al. 2024; Chen et al. 2024). This clade displays two distinct asexual morphologies: (1) an acremonium-like anamorph characterized by subglobose, ellipsoidal, or cylindrical conidia aggregated in slimy heads (Luangsa-ard et al. 2011); and (2) a hirsutella-like morph featuring septate conidiophores with warty protuberances, bearing verticillate phialides that produce dry chains of unicellular, subglobose conidia with apiculate bases or limoniform structures (Chen et al. 2024).
Tolypocladium clade
Tolypocladium contains saprotrophic soil dwellers, plant endophytes, and pathogens targeting insects (e.g., mosquito larvae, fireflies, beetles, cicada nymphs, and bat moth larvae), nematodes, and rotifers, as well as parasites of truffle-like fungi (Yu et al. 2021; Dong et al. 2022). This clade exhibits hirsutella-like asexual morphs, with phialides forming singly or in whorls of 2–6. The phialides display swollen bases and narrowly tapering, frequently bent necks. Conidia are produced singly or aggregated in slimy heads (Gams 1971; Wang et al. 2022).
Dingleyomyces clade
The Dingleyomyces clade currently contains two species (D. lloydii and D. yunnanensis). The type species heavily parasitizes Ophiocordyceps hauturu and O. robertsii stromata (Johnston et al. 2023). It represents the first lineage in the family exhibiting a torrubiella-like macromorphology while possessing a hirsutella-like anamorph. The conidiogenous cells taper toward narrow apices and arise from repeatedly branched conidiophores, which often feature swollen cells. The conidia are cylindrical, straight, aseptate, and have rounded ends. The newly described species D. yunnanensis expands the known host range of the clade to a different species of Ophiocordyceps, indicating that host specialization within Dingleyomyces may not be restricted to a single host lineage. Moreover, D. yunnanensis differs from D. lloydii in its asexual morphology, in which the hirsutella-like phialides are consistently solitary rather than arising from branched conidiophores. These differences in host association and phialide arrangement are consistent with its recognition as a distinct species and provide additional morphological characters for delimiting taxa within the Dingleyomyces clade, in agreement with the phylogenetic results.
Perennicordyceps clade
Perennicordyceps comprises insect-parasitic taxa (parasitizing beetle larvae and cicada larvae) and fungicolous species (parasitizing Cordyceps spp., Ophiocordyceps spp., and Elaphomyces spp.). This clade is characterized by acremonium- and hirsutella-like asexual morphs, along with superficial perithecia. Phialides arise from irregularly branched or subverticillate conidiophores. The acremonium-like morph produces conidia (up to 10) enveloped in a mucilaginous sheath, whereas the hirsutella-like morph bears solitary conidia. Conidial morphology varies from globose to ellipsoid or limoniform to fusiform (Matočec et al. 2014; Wang et al. 2020; Xiao et al. 2023).
Pleurocordyceps clade
The Pleurocordyceps clade parasitizes hosts across six insect orders (Coleoptera, Hemiptera, Homoptera, Hymenoptera, Lepidoptera, and Orthoptera) (Xiao et al. 2023, 2024), with additional fungicolous species infecting Ophiocordyceps and Perennicordyceps (Wang et al. 2015b; Xiao et al. 2023, 2024). The Pleurocordyceps clade produces acremonium- or hirsutella-like phialides and two types of conidia. However, sometimes only one type is observed in fresh specimens. These phialides branch verticillately, producing hyaline, smooth-walled conidia. Two types of conidia are usually present in culture: the α-conidia are globose to subglobose or ellipsoidal, forming masses on the fertile head, while the β-conidia are fusiform, arising singly or in chains along the stipe and mycelial surface (Xiao et al. 2023). The two newly described species, Pleurocordyceps shibingensis and Pl. tengchongensis, conform well to the established morphological circumscription and host associations of the Pleurocordyceps clade. Although they do not exhibit obvious morphological novelties or host-range expansions, their well-supported phylogenetic placement, together with concordant morphological characters, provides additional evidence for the stability and diagnosability of this clade. Moreover, the discovery of these species from geographically distinct regions further contributes to documenting the diversity and distribution of Pleurocordyceps, helping to refine species boundaries and the evolutionary framework within the clade. From an evolutionary perspective, the placement of these species supports a pattern of morphological conservatism within Pleurocordyceps, in which species-level diversification is reflected primarily in phylogenetic differentiation rather than conspicuous morphological or ecological shifts.
Polycephalomyces clade
Species of the Polycephalomyces clade parasitize insects from various orders (Orthoptera, Psocoptera, Coleoptera, Hymenoptera, Lepidoptera, and Hemiptera), as well as Ophiocordyceps spp. and myxomycetes (Xiao et al. 2023). Its asexual morph produces synnemata on the host surface or in culture, with branched or unbranched conidiophores that are divergent. Members of the Polycephalomyces clade produce hirsutella-like phialides (1–4 per conidiophore) that arise terminally or intercalarily, typically featuring a narrow cylindrical base that tapers into a long neck—except in Po. ditmarii, where the phialides remain narrowly cylindrical (although further molecular data are needed to confirm its status). Conidia are monomorphic, oblong to cylindrical, hyaline, and smooth-walled (Xiao et al. 2023). The newly described species, Polycephalomyces bannaensis and Po. chiangraiensis, exhibit morphological characters and host associations that are highly consistent with those of previously recognized members of the clade. Although no conspicuous morphological novelties were observed, their distinct and well-supported phylogenetic positions indicate species-level divergence with limited morphological differentiation. This pattern suggests a degree of evolutionary conservatism in key morphological traits within Polycephalomyces, while diversification appears to be more readily captured at the molecular level. The recognition of these species from different geographic regions further contributes to refining the evolutionary framework and species delimitation within the clade.
Conclusion
This study significantly advances our understanding of hypocrealean entomopathogenic fungi by describing seven new hirsutella-like species through multilocus phylogenetic and morphological analyses and by providing the first comprehensive overview of this morphotype across 19 clades in Ophiocordycipitaceae and Polycephalomycetaceae. Our findings clarify the morphological characteristics of hirsutella-like species in these families, documenting both conserved and divergent phialide traits among major phylogenetic lineages. The pronounced morphological diversity revealed across unrelated clades highlights the inherent limitations of relying solely on anamorphic characters for taxonomic inference. Consequently, this study underscores the importance of an integrative taxonomic framework that combines multilocus phylogeny with morphological evidence to resolve persistent taxonomic ambiguities. By mapping phialide morphologies alongside the phylogenetic tree, this work provides a refined context for future studies and emphasizes the need for continued phylogenetic sampling to further elucidate the diversity and ecological roles of these significant fungal groups.
Supplementary Material
XML Treatment for Ophiocordycipitaceae
XML Treatment for Ophiocordyceps jinguangensis
XML Treatment for Ophiocordyceps northeastensis
XML Treatment for Polycephalomycetaceae
XML Treatment for Polycephalomyces bannaensis
XML Treatment for Polycephalomyces chiangraiensis
XML Treatment for Pleurocordyceps
XML Treatment for Pleurocordyceps shibingensis
XML Treatment for Pleurocordyceps tengchongensis
XML Treatment for Dingleyomyces
XML Treatment for Dingleyomyces yunnanensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adachi K, Chiba K (2007) FTY 720 story. Its discovery and the following accelerated development of sphingosine 1-phosphate receptor agonists as immunomodulators based on reverse pharmacology. Perspectives in medicinal chemistry 1: 11–23. 10.1177/1177391 X 0700100002 PMC 275491619812733 · doi ↗ · pubmed ↗
- 2Araújo JP, Hughes DP (2017) Myrmecophilous Ophiocordyceps as a Case Study. The fungal community: its organization and role in the ecosystem, 359 pp. 10.1201/9781315119496-25 · doi ↗
- 3Araújo JPM, Evans HC, Kepler R et al. (2018) Zombie-ant fungi across continents: 15 new species and new combinations within Ophiocordyceps. I. Myrmecophilous hirsutelloid species. Studies in Mycology 90(1): 119–160. 10.1016/j.simyco.2017.12.002PMC 600235629910522 · doi ↗ · pubmed ↗
- 4Araújo JPM, Evans HC, Fernandes IO et al. (2020) Zombie-ant fungi cross continents: II. Myrmecophilous hymenostilboid species and a novel zombie lineage. Mycologia 112(6): 1138–1170. 10.1080/00275514.2020.182209333146584 · doi ↗ · pubmed ↗
- 5Araújo JPM, Moriguchi MG, Uchiyama S et al. (2021) Ophiocordyceps salganeicola, a parasite of social cockroaches in Japan and insights into the evolution of other closely-related Blattodea-associated lineages. IMA Fungus 12: 3. 10.1186/s 43008-020-00053-9PMC 786338133541437 · doi ↗ · pubmed ↗
- 6Araújo JPM, Lebert BM, Vermeulen S et al. (2022) Masters of the manipulator: Two new hypocrealean genera, Niveomyces (Cordycipitaceae) and Torrubiellomyces (Ophiocordycipitaceae), parasitic on the zombie ant fungus Ophiocordyceps camponoti-floridani. Persoonia-Molecular Phylogeny and Evolution of Fungi 49(1): 171–194. 10.3767/persoonia.2022.49.05PMC 1079222838234384 · doi ↗ · pubmed ↗
- 7Ban S, Sakane T, Nakagiri A (2015) Three new species of Ophiocordyceps and overview of anamorph types in the genus and the family Ophiocordycipitaceae. Mycological Progress 14(1): 1017–1030. 10.1007/s 11557-014-1017-8 · doi ↗
- 8Ban S, Sakane T, Toyama K et al. (2009) Teleomorph–anamorph relationships and reclassification of Cordyceps cuboidea and its allied species. Mycoscience 50(4): 261–272. 10.1007/S 10267-008-0480-Y · doi ↗