Differential effects of focused ultrasound neuromodulation in Parkinson’s disease mice versus healthy mice
Leqi Yang, Kevin Xu, Dingyue Zhang, Andrew Stark, Yimei Yue, Alexxai Kravitz, Yaoheng Yang, Hong Chen

TL;DR
Focused ultrasound neuromodulation improves motor function in Parkinson’s disease mice but not in healthy mice, showing its effects depend on disease state.
Contribution
Demonstrates disease-state-dependent effects of FUS neuromodulation in a Parkinson’s disease model.
Findings
FUS inhibited GPe calcium activity in PD mice, lasting ~3 minutes post-stimulation.
Motor improvements in PD mice lasted at least 50 minutes after FUS.
No significant effects were observed in healthy mice, and no tissue damage was detected.
Abstract
Objective. Focused ultrasound (FUS) neuromodulation holds strong potential for treating neurological disorders, but most preclinical studies have been performed in healthy animal models. How disease states influence the FUS neuromodulation effects remains poorly understood, limiting clinical translation. Approach. We used Parkinson’s disease (PD) as a model to compare the calcium and behavioral responses to FUS neuromodulation in healthy and diseased mice. The PD mouse model was the unilateral dopamine depletion model, induced by injecting 6-hydroxydopamine into the left middle forebrain bundle. FUS was targeted at the left external globus pallidus (GPe) in freely moving mice using a wearable device. Calcium activity in the GPe was monitored via fiber photometry, and motor behavior was assessed using video tracking. Main results. In unilateral PD mice, FUS significantly inhibited GPe…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
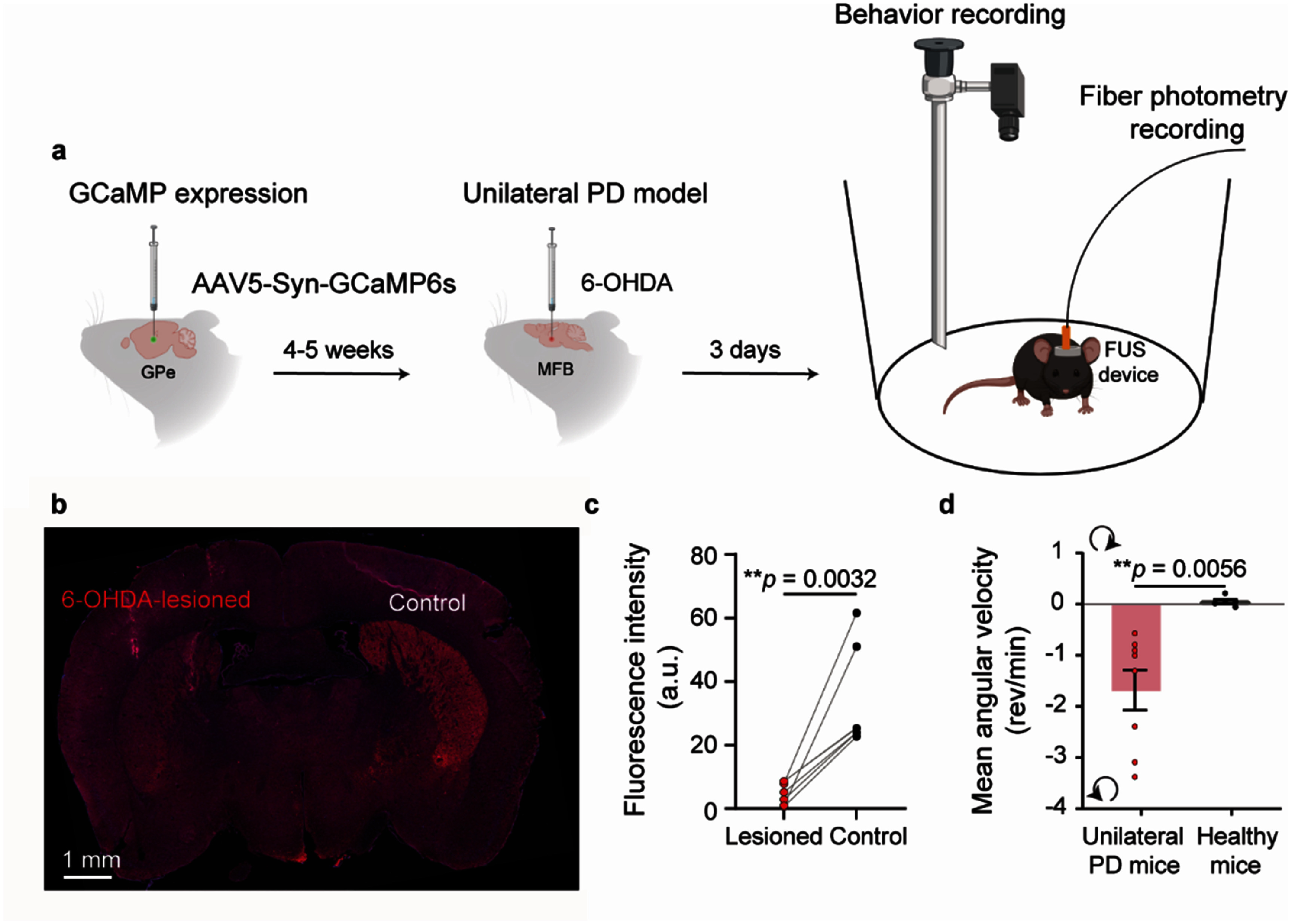 Figure 1
Figure 1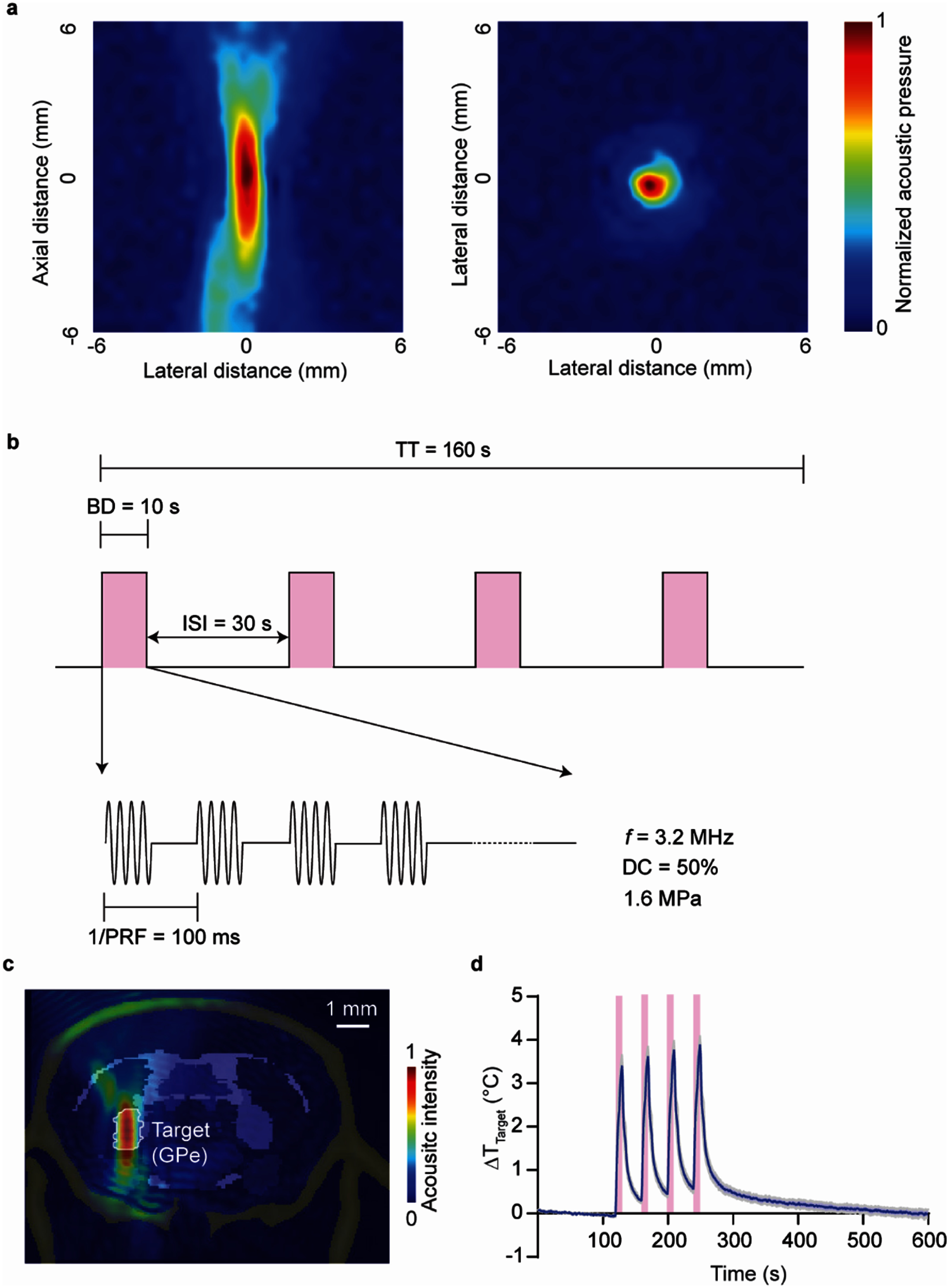 Figure 2
Figure 2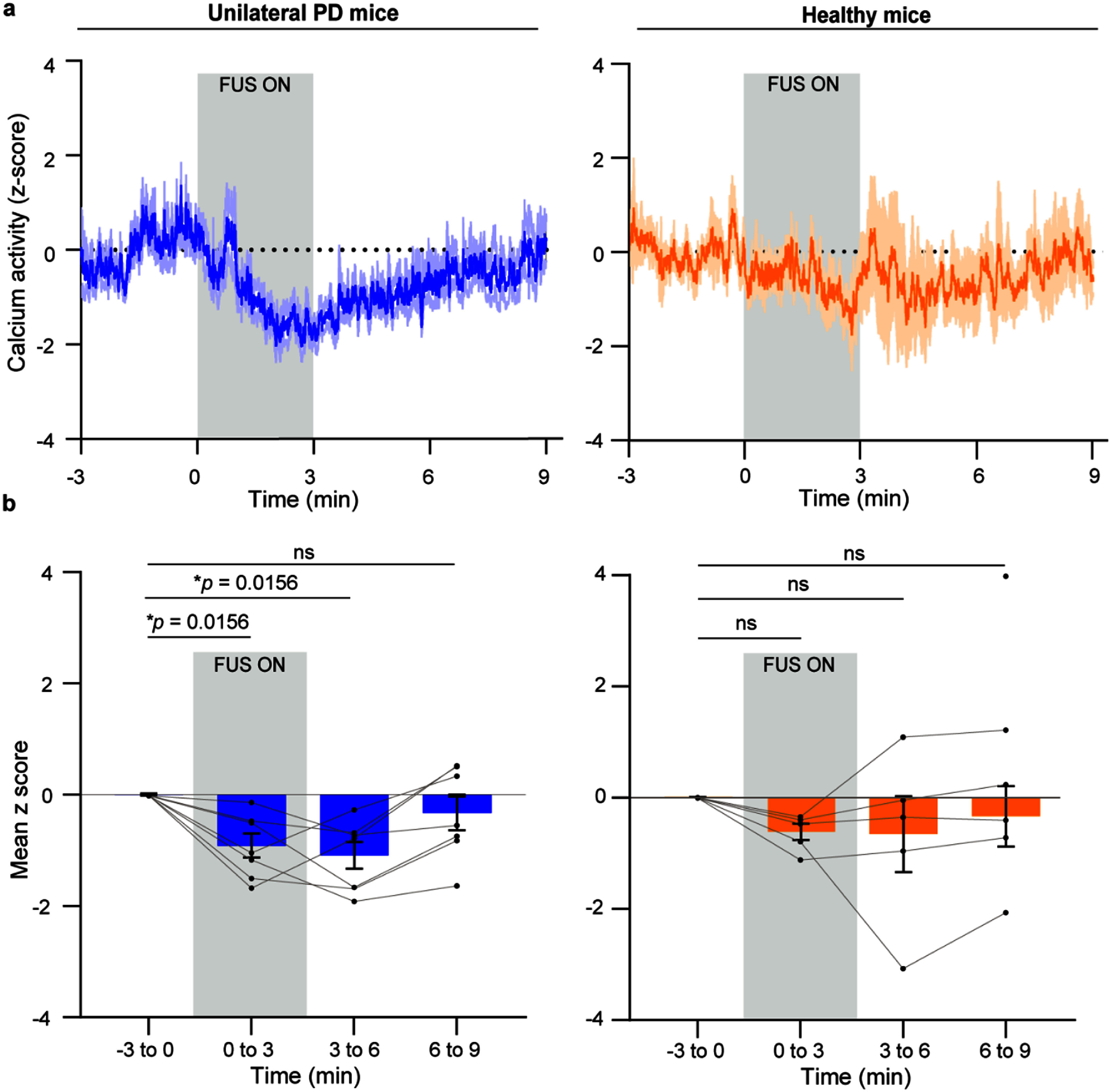 Figure 3
Figure 3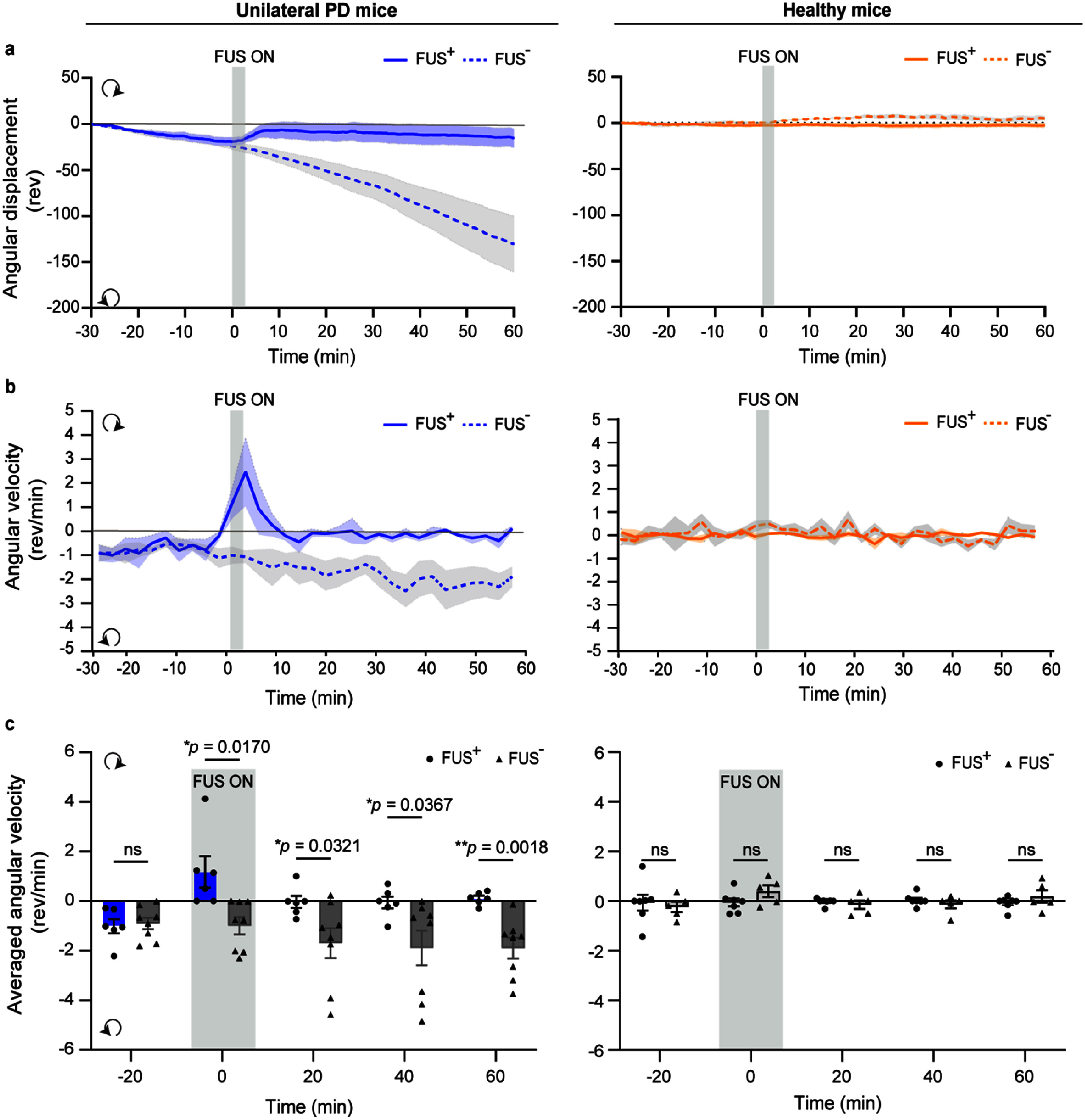 Figure 4
Figure 4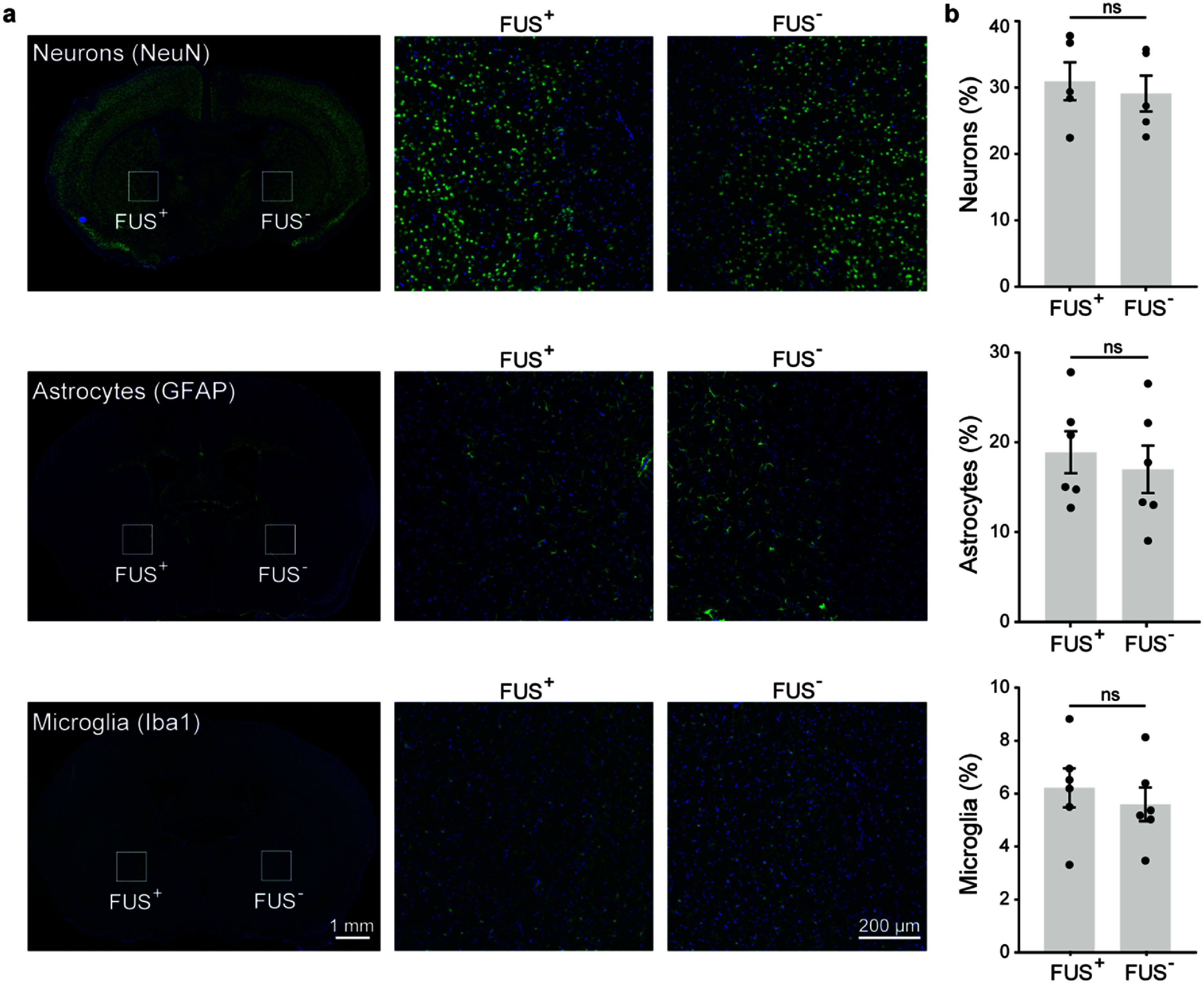 Figure 5
Figure 5| Time (min) | Unilateral PD mice mean | Unilateral PD mice 95% CI | Healthy mice mean | Healthy mice 95% CI |
|---|---|---|---|---|
| −3–0 | 0 | [0, 0] | 0 | [0, 0] |
| 0–3 | −0.92 | [−1.44, −0.38] | –0.62 | [−1.03, −0.22] |
| 3–6 | −1.09 | [−1.68, −0.50] | –0.66 | [−2.57, 1.24] |
| 6–9 | −0.33 | [−1.09, −0.43] | –0.35 | [–1.85, 1.16] |
| Time (min) | Unilateral PD mice/FUS+ | Unilateral PD mice/FUS– | Healthy mice/FUS+ | Healthy mice/FUS– |
|---|---|---|---|---|
| –20 | −1.01 [−1.75, −0.27] | –0.90 [–1.46, −0.34] | −0.06 [−0.84, 0.71] | −0.17 [−0.53, 0.20] |
| 0 | 1.17 [−0.45, 2.79] | –0.99 [–1.82, −0.16] | −0.05 [−0.44, 0.34] | 0.28 [−0.15, 0.72] |
| 20 | −0.04 [−0.65, 0.58] | –1.69 [–3.12, −0.25] | −0.02 [−0.15, 0.10] | −0.10 [−0.41, 0.20] |
| 40 | −0.06 [−0.67, 0.55] | –1.88 [–3.53, −0.23] | 0.04 [−0.18, 0.26] | −0.09 [−0.38, 0.20] |
| 60 | 0.11 [−0.16, 0.37] | –1.89 [–2.89, −0.90] | −0.07 [−0.30, 0.16] | 0.14 [−0.30, 0.57] |
- —National Institute of Neurological Disorders and Stroke10.13039/100000065
- —National Institute of Diabetes and Digestive and Kidney Diseases10.13039/100000062
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsUltrasound and Hyperthermia Applications · Vagus Nerve Stimulation Research · Ultrasound and Cavitation Phenomena
Introduction
Focused ultrasound (FUS) neuromodulation has emerged as a promising noninvasive approach for treating neurological disorders such as Parkinson’s disease (PD) [1], Alzheimer’s disease (AD) [2], and essential tremor [3]. Compared with deep brain stimulation (DBS), FUS offers the distinct advantage of being noninvasive. Unlike other noninvasive neuromodulation techniques, such as transcranial magnetic stimulation (TMS) and transcranial direct current stimulation (tDCS), FUS uniquely combines deep brain penetration with high spatial precision. By adjusting the acoustic parameters, FUS can both excite and inhibit neuronal activity across species, including rodents [4, 5], non-human primates [6], and humans [7]. Despite its potential, FUS neuromodulation outcomes are variable and can be influenced by multiple factors beyond ultrasound parameters. One key factor is the brain state at the time of stimulation [8].
The nervous system is in a constant state of flux, transitioning through various brain states. These different brain states are regulated by varying levels of activity in interconnected brain regions and by the modulation of connections between these regions [9]. When an external neurostimulation is applied, its effects can vary depending on the brain’s current state—a phenomenon known as state-dependent neuromodulation. This concept frames the brain as a dynamic system in which responses to external stimuli are critically shaped by ongoing neural activity [9].
Neurological diseases significantly alter both local neuronal activity and large-scale brain network connectivity [10, 11], potentially changing how the brain responds to neuromodulation. Prior studies had shown that the effects of TMS and tDCS differed between healthy individuals and patients with neurological disorders [12, 13]. For example, Mir et al showed that TMS applied to the dorsal premotor cortex facilitated motor-evoked potentials (MEPs) in healthy subjects but had no such effect in PD patients off medication [14]. Inghilleri et al found that TMS increased MEPs in healthy controls but decreased MEPs in patients with AD [15]. Hsu et al reported tDCS had a small effect size in healthy adults but a large effect size in adults with AD [16]. These findings highlight the importance of accounting for disease state when applying neuromodulation to reduce variability and improve efficacy. However, the impact of disease states on FUS neuromodulation remains largely unexplored. Addressing this gap is critical for the clinical translation of FUS-based therapies and elucidating the mechanisms through which disease states influence stimulation outcomes.
PD is one of the most prevalent neurodegenerative disorders and the most extensively studied condition in preclinical FUS neuromodulation research [17]. It serves as a model for investigating disease-state-dependent effects of FUS neuromodulation. PD is characterized by progressive degeneration of dopaminergic neurons in the substantia nigra pars compacta, leading to widespread dysfunction in the basal ganglia circuitry (BGC) [18]. In the healthy brain, dopamine maintains a balance between the direct and indirect pathways within the BGC [19]. In PD, dopamine depletion disrupts this balance, resulting in altered activity throughout the BGC [20]. These disease-related circuit changes may fundamentally reshape how the brain responds to FUS neuromodulation. However, this effect has not been fully understood or characterized.
In PD, the globus pallidus (GP) becomes pathologically hyperactive [21]. DBS targeting the GP has been used to suppress this activity and alleviate motor dysfunction in patients [22]. Notably, recent studies show that inhibiting specific external GP (GPe) neuronal subpopulations using optogenetics or DBS can produce long-lasting motor recovery in dopamine-depleted mouse models [23, 24], which makes the GPe an attractive node for PD treatment.
In this study, we investigated whether PD alters calcium and rotational behavior responses to FUS neuromodulation. Specifically, we chose the GPe as our target region to evaluate whether ultrasound could induce sustained motor improvement. Using a unilateral PD mouse model generated by 6-hydroxydopamine (6-OHDA) injection into the left hemisphere, we delivered FUS to the left GPe via a wearable device. We recorded the calcium activity using fiber photometry and assessed motor behavior via video tracing. In unilateral PD mice, FUS inhibited GPe calcium activity during the stimulation and for approximately 3 min after stimulation, accompanied by improved motor function as evidenced by reduced ipsilateral circling lasting for at least 50 min. In contrast, healthy mice exhibited no calcium change and also showed no rotational behavior changes. These findings demonstrate that disease state significantly modulates the effects of FUS neuromodulation and underscore the importance of evaluating neuromodulation strategies in pathological contexts.
Methods
Animals
2.1.
Adult C57BL/6NCrl mice (6–10 weeks old; Charles River Laboratories) were randomly assigned to experimental groups. All mice were housed in the Washington University School of Medicine animal facility under controlled environmental conditions (23°C–26°C, 35%–65% humidity) with a 12 h light/dark cycle and ad libitum access to standard chow. All animal procedures were approved by the Institutional Animal Care and Use Committee at Washington University in St. Louis and conducted in accordance with NIH guidelines for animal research (protocol no. 24–0222).
A total of 69 mice were included across six experimental studies. (1) TH staining (n = 6; figure 1(c)). (2) Baseline differences between unilateral PD mice and healthy controls (n = 13, figure 1(d)). (3) Temperature elevation in the target region during FUS stimulation (n = 5; figure 2(d)). (4) Fiber photometry recordings in unilateral PD mice (n = 7) and healthy mice (n = 6) (figure 3). (5) Behavioral assessments including unilateral PD mice with FUS treatment (n = 6), unilateral PD mice without FUS treatment (n = 8), healthy mice with FUS treatment (n = 7), and healthy mice without FUS treatment (n = 5) (figure 4). Three mice with unilateral 6-OHDA injections were excluded due to the absence of a clear rotational bias. (6) Staining for safety evaluation (n = 6; figure 5), and one mouse in the NeuN staining group was excluded due to poor staining quality.
Experimental setup and unilateral PD model validation. (a) Mice were injected with AAV5-Syn-GCaMP6s into the left GPe for fiber photometry recording. After a 4–5 week expression period of GCaMP6s, 6-OHDA was injected into the MFB in the left hemisphere to generate a unilateral PD mouse model. On the same day, baseplates were implanted on the mice’s heads to enable future unilateral FUS stimulation targeting the left GPe. 3 d after the 6-OHDA lesion, a wearable FUS device was mounted onto the baseplates. Behavioral activity was recorded using a camera to monitor changes before, during, and after FUS stimulation. The calcium activity was recorded with fiber photometry. (Images created with BioRender.com) (b) A representative TH (red) and DAPI (blue) staining image of mice with unilateral 6-OHDA injection (left hemisphere). (c) Quantification of TH staining (n = 6). (d) Quantification of baseline mean angular velocity in unilateral PD (n = 8) and healthy mice (n = 5). Positive and negative values indicate contralateral and ipsilateral rotations, respectively.
Transducer acoustic intensity profile and ultrasound parameters. (a) Acoustic pressure fields in the axial focal plane and lateral focal plane were measured by a hydrophone in a water tank. (b) Schematic of ultrasound parameters in the study. FUS was delivered with a center frequency of 3.2 MHz, an acoustic pressure of 1.6 MPa after skull attenuation, a pulse repetition frequency (PRF) of 10 Hz, and a duty cycle (DC) of 50%. The burst duration (BD) was 10 s with an inter-stimulus interval (ISI) of 30 s, for a total of four stimulations with a total time (TT) of 160 s. (c) Simulation of ultrasound pressure fields with skull. (d) Temperature rise in the target area measured by an implanted fiber-optic thermometer (n = 5).
FUS induced different calcium responses in unilateral PD mice (n = 7) and healthy mice (n = 6). (a) Calcium activity (ΔF/F, z-score) of neurons before, during, and after FUS stimulation in unilateral PD mice (left) and healthy mice (right). (b) Quantification of the mean z-score of calcium activity across four time periods in unilateral PD mice (left) and healthy mice (right). Each dot represents one mouse. The box plots show mean ± SEM. The statistical analysis was performed via Friedman test followed by Wilcoxon signed-rank tests. The vertical bar indicates the duration of FUS stimulation.
FUS evoked different rotational behavior changes in unilateral PD mice and healthy mice. (a) Angular displacement over time in unilateral PD mice (left) and healthy mice (right), comparing FUS-stimulated (FUS+) and non-stimulated (FUS–) groups. (b) Angular velocity over time in unilateral PD and healthy mice across FUS+ and FUS—groups. Positive values indicate contralateral (clockwise) rotation, while negative values indicate ipsilateral (counterclockwise) rotation. (c) Quantification of mean angular velocity over five time periods. Sample sizes: unilateral PD mice/FUS+ (n = 6), unilateral PD mice/FUS—(n = 8), healthy mice/FUS+ (n = 7), healthy mice/FUS—(n = 5). The box plots show mean ± SEM. The statistical analysis was performed via mixed-model followed by Tukey’s post hoc test. The vertical bar indicates the duration of FUS stimulation.
FUS stimulation does not induce cellular damage in the targeted brain region (n = 6). (a) Representative immunofluorescence images showing staining of neurons (NeuN), astrocytes (GFAP), and microglia (Iba1). Blue color represents DAPI and green color represents specific cell types. (b) Quantification of the ratio of positively stained cells to total DAPI-stained cells in the FUS-stimulated and contralateral control regions. Data are presented as mean ± SEM. Statistical comparisons were made using paired two-tailed t-tests.
Unilateral 6-OHDA injection
2.2.
To induce a unilateral PD model, mice were anesthetized with 1.5%–2.0% isoflurane in oxygen and secured in a stereotaxic frame (Kopf instruments). A small hole was drilled through the skull above the medial forebrain bundle (MFB) in the left hemisphere (coordinates: AP = −0.5 mm, ML = −1.1 mm relative to bregma). A Hamilton syringe was used to inject 1 µl of 6-hydroxydopamine (6-OHDA, 5 µg µl^−1^ in 0.9% NaCl; Sigma-Aldrich) into the MFB at a depth of 5 mm from the skull surface at a rate of 0.3 µl min^−1^. The needle was left in place for 3 min post-injection before being slowly withdrawn. The injection site was then sealed with bone wax. To confirm the successful establishment of the unilateral PD mouse model, inclusion was based on behavioral verification of an effective 6-OHDA lesion. After recovery from surgery, locomotor behavior was assessed to evaluate lesion efficacy. Only animals that exhibited clear ipsilateral circling behavior as shown in figure 1(d) were included for subsequent experiments related to unilateral PD mice.
Calibration of ultrasound transducers and simulation
2.3.
The ultrasound transducer was made using a lead zirconate titanate ceramic resonator (DL-43, DeL Piezo Specialties). Calibration of the ultrasound transducer was performed in degassed water without and with ex vivo mouse skulls. To assess skull attenuation, freshly extracted mouse skulls were positioned in front of the transducer, and the acoustic pressure fields were measured using a calibrated hydrophone (HGL-0200, Onda; 200 µm active element). The transmission efficiency of the ultrasound was calculated as the ratio of the maximum peak negative pressures measured with and without the skull in place. Using this approach, the average transmission efficiency was approximately 50.9%.
The detailed procedure of ultrasound simulation was described previously [25]. Briefly, the k-space pseudospectral method-based solver, k-Wave [26] was used for simulation. To accurately represent the acoustic interaction with the skull, we derived the skull’s acoustic properties from computed tomography (CT) scans of a mouse head using a μCT scanner (Rigaku). The density and sound speed of the skull and brain tissue were converted from the Hounsfield units of the CT images using the function ‘hounsfield2density’ in the k-Wave toolbox. From the CT, the skull thickness is ∼0.3–0.4 mm.
Brain temperature measurement
2.4.
Under 1.7% isoflurane anesthesia, mice were secured in a stereotaxic frame, and a small burr hole was drilled above the left GPe (AP = −0.3 mm, ML = −2.2 mm, DV = −3.6 mm). A baseplate was affixed to the skull and aligned to the GPe as previously described. A wearable transducer with a 1 mm-diameter central opening was then attached to the baseplate. A fiber-optic thermometer sampling at 60 Hz was inserted through the opening and slowly lowered into the GPe. After allowing approximately 5 min for the signal to stabilize, 2 min of baseline brain temperature were recorded. FUS was subsequently delivered using the parameters shown in figure 2(b). Post-stimulation recording continued until the brain temperature returned to baseline. Temperature traces were then normalized to the mean value of the 2 minute baseline period.
Ultrasound stimulation and behavioral testing
2.5.
After the 6-OHDA injection, a baseplate for FUS coupling was affixed to the skull. A dot was marked on the skull to indicate the location of the left GPe (AP = −0.3 mm, ML = −2.2 mm, DV = −3.6 mm relative to bregma). The baseplate (centered on the dot) was secured using dental adhesive (Metabond), with its center aligned to the dot marker. Buprenorphine SR (1.0 mg kg^−1^, s.c.) was administered for postoperative analgesia. Mice were allowed to recover for at least 3 days before further experimentation. For healthy control mice, baseplates were affixed using the same procedure.
To minimize stress, mice were habituated to the behavior test setup and exposed to the sounds of the function generator and power amplifier on the day prior to stimulation. On the test day, mice were briefly anesthetized (1.5%–2.0% isoflurane). The wearable ultrasound transducer was encased in a custom 3D-printed housing and filled with degassed ultrasound gel (Aquasonic) to ensure optimal acoustic coupling. The transducer was securely mounted onto the baseplate on the mouse head using adhesive, providing a tight and stable connection throughout the experiment. For studies involving fiber photometry recording, the wearable transducer was custom-modified by drilling a 2 mm-diameter central hole to accommodate the optical fiber. This modification allowed the mating sleeve to pass through the center of the transducer and connect the stainless-steel ferrule to the implanted cannula, enabling simultaneous optical recording and ultrasound stimulation at the same target site. After placement in a circular open-field arena, mice were allowed to fully recover from anesthesia for one hour. Mouse behavior was recorded using a camera (Logitech C920X, 30 fps) before, during, and after FUS stimulation.
Ultrasound parameters were: frequency = 3.2 MHz; peak negative pressure = 1.6 MPa (after skull attenuation); duty cycle = 50%; pulse repetition frequency = 10 Hz; stimulus duration = 10 s; inter-stimulus interval = 30 s; number of stimulations = 4. Pulses were smoothed to avoid auditory artifacts [27]. A custom MATLAB script and Arduino Uno controlled the stimulation onset. A red LED synchronized to pulse onset provided visual cues. Videos were processed in Bonsai (v2.7) to extract position and angular orientation. Angular displacement and angular velocity were quantified using custom MATLAB code. Time 0 was defined as the onset of the first FUS pulse. Average angular velocity was calculated at five different time points: 20 min before FUS, during FUS, 20 min post-FUS, 40 min post-FUS, 60 min post-FUS.
Virus injection and fiber photometry recording
2.6.
To evaluate calcium activity in the GPe during FUS stimulation, we used fiber photometry to record GCaMP6s signals. Under 1.7% isoflurane anesthesia, mice were secured in a stereotaxic frame, and a small burr hole was drilled on the skull above the left GPe (AP = −0.3 mm, ML = −2.2 mm, DV = −3.6 mm). A glass capillary was slowly lowered to DV = −3.6 mm from the skull surface, and AAV5-Syn-GCaMP6s (0.345 µl; 7.0 × 10^12^ vg/mL; Addgene) was infused at 23 nl s^−1^ into the left GPe utilizing a microinjector (Nanoject II, Drummond Scientific Company). The capillary was left in place for 10 min before being slowly withdrawn. The hole was then sealed with bone wax. After four weeks to allow for viral expression, an optical fiber cannula (MFC_200/245-0.37_6mm_ZF1.25_FLT, Doric Lenses) was implanted ∼200 µm above the injection site through the baseplate opening, ensuring alignment of the fiber tip and ultrasound focus at the GPe.
Excitation light (470 nm, 4% power) was delivered via the fiber, and GCaMP6s emission was collected using a fiber photometry system (FP3002, Neurophotometrics) synchronized with FUS stimulation. Signals were acquired using Bonsai (v2.7). Data were analyzed using a custom MATLAB script as described previously [4]. Z score values were computed using the MATLAB ‘zscore’ function. Mean z-scores were calculated for four time windows: 160 s pre-FUS, 160 s during FUS, 160 s immediately post-FUS, and 320 s delayed post-FUS.
Histological analysis
2.7.
Approximately 6 h after FUS stimulation, mice were perfused with PBS, and brains were fixed overnight in 4% paraformaldehyde, then cryoprotected in 30% sucrose. Tyrosine hydroxylase (TH) staining was performed to confirm dopaminergic neuron depletion in unilateral PD models. Fluorescence intensity of TH staining was quantified using ImageJ. Striatal intensity in both the lesioned and control hemispheres was normalized to the cortical intensity. Immunostaining was performed using primary antibodies against NeuN (Abcam, cat. no. 104 225; 1:1000), GFAP (Abcam, cat. no. 207 165; 1:1000), and Iba1 (Abcam, cat. no. 178 846; 1:1000), as well as DAPI mounting medium (Vector). Sections were imaged using a multichannel Keyence BZ-X800 microscope (×20 objective) and analyzed with BZ-X800 analyzer software. Cell counts were obtained using QuPath (University of Edinburgh). The percentage of NeuN-, GFAP-, or Iba1-positive cells relative to the total number of DAPI-stained nuclei was calculated per animal.
Statistical analysis
2.8.
Statistical tests were conducted using GraphPad (Prism). Normality was assessed using the Quantile–Quantile (Q–Q) plot. Data was analyzed using Friedman tests followed by Wilcoxon signed-rank post hoc test, mixed-model followed by Tukey’s post hoc test or paired two-tailed t-tests. Results were considered statistically significant at p < 0.05. Data is reported as mean ± standard error of the mean (SEM).
Results
Differential calcium responses in unilateral PD mice vs healthy mice
3.1.
We first injected 6-OHDA into the MFB in the left hemisphere to establish a unilateral PD mouse model. TH immunostaining revealed a marked loss of TH-positive fibers in the striatum of the lesioned hemisphere (left) compared to the contralateral control hemisphere (right) (figures 1(b) and (c)). At the behavioral level, unilaterally lesioned PD mice exhibited ipsilateral circling behavior in the open field test, whereas healthy mice showed no rotational preference (figure 1(d)).
To assess the impact of FUS on GPe neurons in unilateral PD and healthy conditions, we performed fiber photometry recordings of calcium signals in both unilateral PD and healthy mice (figure 1(a)). FUS was delivered to the left GPe according to the stimulation protocol shown in figure 2(b). In unilateral PD mice, we observed a consistent decrease in ΔF/F (z-score) of GPe calcium activity during FUS stimulation compared with that before stimulation. Notably, this calcium suppression continued after FUS ended and gradually returned to baseline levels about three minutes post-stimulation (figure 3(a), left). After normality test (figure S6), a Friedman test revealed a significant effect of time on the calcium signal in unilateral PD mice, p = 0.0056. Post hoc Wilcoxon signed-rank tests indicated significant decreases in calcium at 0–3 min (p = 0.0156) and 3–6 min (p = 0.0156) compared with the −3–0 min baseline (figure 3(b), left). The same analysis in healthy mice detected no significant effect of time on the calcium signal, p = 0.0666 (figures 3(a) and (b), right, table 1). Together, these findings demonstrate that FUS induces a prolonged or ‘offline’ inhibitory effect on GPe calcium activity in unilateral PD mice, but not in healthy mice. This highlights a disease-state–dependent difference in calcium response to identical FUS parameters.
Differential behavioral responses in unilateral PD mice vs healthy mice
3.2.
To test whether FUS could alleviate the unilateral PD-related motor deficit as shown in figure 1(d), we applied unilateral stimulation to the left GPe using the same parameters as described earlier. FUS significantly reduced circling behavior in unilateral PD mice, as evidenced by a decrease in total accumulated angular displacement at the end of the recording period (–15.06 ± 10.31 revolutions with FUS vs −130.30 ± 30.35 revolutions without FUS, figure 4(a), left). In contrast, FUS had no significant effect on angular displacement in healthy mice (–2.39 ± 2.68 revolutions with FUS vs −5.12 ± 2.41 revolutions without FUS, figure 4(a), right). This effect was further confirmed by analyzing angular velocity over time across groups (figures 4(b) and (c)). In unilateral PD mice, unilateral FUS stimulation of the GPe reversed the 6-OHDA-induced ipsilateral rotation bias to contralateral rotation during the stimulation (1.17 ± 0.63 rev min^−1^ with FUS vs −0.99 ± 0.35 rev min^−1^ without FUS), indicating a shift toward balanced movement (figure 4(b), left). After the stimulation, we observed sustained motor recovery, with no apparent rotational preference in unilateral PD mice for at least 50 min (figure 4(c)). In healthy mice, FUS did not alter angular velocity (–0.05 ± 0.16 rev min^−1^ with FUS vs 0.40 ± 0.23 rev min^−1^ without FUS, figures 4(b) and (c)). A mixed model revealed a significant effect of the FUS state (stimulated vs non-stimulated), F(1,12) = 12.81, p = 0.0038, a close to significant effect of the Time, F(2.371, 28.45) = 2.732, p = 0.0739, and a significant interaction between these two factors on rotational behavior, F(4, 48) = 2.825, p = 0.0349. Post hoc Tukey tests revealed significant differences between the FUS-stimulated and non-stimulated conditions at the 0, 20, 40, and 60 min time-points (figure 4(c)). An identical analysis on the healthy mice did not detect a significant effect of time, FUS state, or an interaction between these two factors on the rotational behavior (table 2). These results demonstrate that the same FUS neuromodulation parameters can alleviate unilateral PD-related circling behavior while failing to alter rotational behavior in healthy mice, suggesting that FUS-induced behavioral effects are brain state-dependent (PD vs healthy).
FUS neuromodulation was safe at the cellular level
3.3.
To assess the safety of our FUS parameters, we calculated the mechanical index (MI) and the cumulative equivalent minutes at 43 °C (CEM43). The MI was 0.89 and CEM43 = 0.0218 at the target area based on the temperature measurement shown in figure 1(d). Both values fall well within the safety thresholds defined by the ITRUSST guidelines [28], indicating minimal risk of mechanical or thermal damage under our stimulation conditions. We also assessed the safety of FUS stimulation by performing immunohistochemical staining for neurons (NeuN), astrocytes (GFAP), and microglia (Iba1) in healthy mice. No significant differences were observed in the number of these cell types between the FUS-stimulated GPe in the left hemisphere and the non-stimulated contralateral GPe in the right hemisphere (figure 5), suggesting that the FUS parameters used in this study were well-tolerated at the cellular level.
Discussion
This study compared the effects of FUS neuromodulation on calcium activity and behavior in PD versus healthy mice. In unilateral PD mice, FUS inhibited calcium activity during stimulation, with suppression persisting for approximately 3 min after stimulation ended. In contrast, in healthy mice, FUS did not induce significant calcium activity change. Behaviorally, FUS alleviated the characteristic ipsilateral circling behavior in unilateral PD mice, which lasted for at least 50 min after FUS stopped. However. The same FUS stimulation had no noticeable effect on healthy controls. This study highlighted that the disease state significantly modulates the effects of FUS neuromodulation, underscoring the importance of evaluating neuromodulation strategies in pathological contexts to optimize therapeutic efficacy.
The concept of state-dependent neuromodulation emerged more than two decades ago in studies using TMS and tDCS. Silvanto and Pascual-Leone showed that the efficacy of TMS depends on the baseline cortical excitability of the targeted brain region [8], whereas Antal et al demonstrated that tDCS effects are influenced by the subject’s cognitive state [29]. Recent studies have extended the concept of state dependency to FUS neuromodulation, demonstrating that its effects vary with behavioral and neural states. For example, Wang et al reported that FUS-evoked spiking activity in the cortex differs between anesthetized, awake, and running states [30]. Nguyen et al found that baseline brain rhythms modulate FUS outcomes [31]. Yang et al used functional magnetic resonance imaging to show that FUS either suppressed or activated neural responses depending on whether it was paired with sensory input or delivered at rest [32]. Prieto et al demonstrated in vitro that FUS could either inhibit or enhance firing in hippocampal neurons depending on spike frequency [33]. However, nearly all previous research has focused on state dependency in healthy systems. Our study adds to this body of work by demonstrating that disease state also significantly influences the outcomes of FUS neuromodulation.
Several studies have reported the therapeutic potential of FUS in PD animal models. For instance, in 6-OHDA-lesioned rats, Sung et al found that FUS improved locomotor function, potentially by exerting neuroprotective effects and reducing 6-OHDA-induced neurotoxicity [34]. In a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine induced PD mouse model, Zhou et al showed that FUS protected dopaminergic neurons by modulating apoptotic pathways, including the downregulation of Bax and cleaved caspase-3, and upregulation of Bcl-2 [35]. Wang et al demonstrated that FUS reduced beta-band activity (13–30 Hz) in the basal ganglia, which is typically elevated in PD [36], and hypothesized that this effect arose from direct modulation of the BGC. Previous studies showed that inhibiting specific GPe neuronal subpopulations using optogenetics or DBS can produce long-lasting motor recovery in dopamine-depleted mouse models [23, 24]. Prior studies have shown that FUS can generate mild focal heating that preferentially produces inhibitory effects [37, 38]. Guided by this evidence, we selected FUS parameters designed to induce mild heating at the focal zone with the goal of inhibiting GPe neuronal activity while maintaining established safety margins (figures 2(c) and (d)). Our findings demonstrate that FUS directly inhibits the GPe calcium activity and improves motor behavior in a 6-OHDA-induced unilateral PD mouse model. Notably, the behavioral improvement as reflected by increased angular velocity persisted for at least 50 min after stimulation, well beyond the FUS application period. Comparable offline effects have been observed with optogenetic stimulation [23] and DBS [24] targeting the GPe. However, unlike these approaches, FUS does not require gene modification or invasive implantation, making it a promising candidate for clinical translation. Importantly, this sustained benefit was not accompanied by detectable tissue damage, as no significant changes were observed in the number of neurons, astrocytes, or microglia within the FUS-stimulated GPe (figure 5). One potential mechanism underlying this prolonged effect is that FUS may dissociate the firing rates of PV neurons from those of Lhx6 neurons in the GPe, promoting long-lasting behavioral rescue [23]. While the exact mechanism of this sustained effect remains to be elucidated, these findings highlight the therapeutic potential of FUS neuromodulation for PD. It is also important to note that different combinations of acoustic parameters can shift the response from inhibition to excitation, or produce minimal effects, and this is also dependent on the intrinsic properties and ongoing state of the brain. Although our parameter set was chosen to induce inhibition, alternative settings could elicit excitatory responses that may either benefit or worsen Parkinsonian symptoms. These parameter-dependent effects warrant systematic investigation in future studies.
This study has several limitations. The 6-OHDA mouse model, while widely used, does not fully recapitulate the progressive and multifactorial nature of human PD. Future work should examine whether FUS induces differential effects at different stages of disease progression or in other PD models. Fiber photometry offers a valuable approach for monitoring population-level calcium dynamics in freely behaving animals. However, it remains an indirect measure of neuronal activity. Complementary electrophysiological techniques in future studies will help clarify how disease states influence ultrasound neuromodulation outcomes at the level of neuronal spiking. In addition, although our findings establish a clear difference between PD and healthy mice, the generalizability of these results across species, including humans, remains uncertain. Furthermore, whether FUS elicits disease-dependent effects in other neurological disorders beyond PD remains to be explored. Nonetheless, our findings provide important insight into how disease states modulate the efficacy of FUS neuromodulation and highlight the need to account for these differences in both research and clinical applications.
In conclusion, this study demonstrates that FUS neuromodulation exerts distinct effects on calcium activity and motor behavior depending on the brain state. In unilateral PD mice, FUS inhibited GPe calcium activity both during and after stimulation and alleviated unilateral PD-related circling behavior. In contrast, the same FUS parameters did not induce calcium activity change in healthy mice and also did not affect behavior. These results support the concept that FUS neuromodulation may be affected by the disease state, emphasizing the need to consider the brain state when designing and applying FUS-based therapies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lee K S Clennell B Steward T G Gialeli A Cordero-Llana O Whitcomb D J 2022 Focused ultrasound stimulation as a neuromodulatory tool for Parkinson’s disease: a scoping review Brain Sci.1228910.3390/brainsci 1202028935204052 PMC 8869888 · doi ↗ · pubmed ↗
- 2Beisteiner R et al 2020 Transcranial pulse stimulation with ultrasound in Alzheimer’s disease—a new navigated focal brain therapy Adv. Sci.7190258310.1002/advs.201902583 PMC 700162632042569 · doi ↗ · pubmed ↗
- 3Riis T S Losser A J Kassavetis P Moretti P Kubanek J 2024 Noninvasive modulation of essential tremor with focused ultrasonic waves J. Neural Eng.2101603310.1088/1741-2552/ad 27ef 38335553 · doi ↗ · pubmed ↗
- 4Yang Y et al 2023 Induction of a torpor-like hypothermic and hypometabolic state in rodents by ultrasound Nat. Metab.5789803789–80310.1038/s 42255-023-00804-z 37231250 PMC 10229429 · doi ↗ · pubmed ↗
- 5Tufail Y Matyushov A Baldwin N Tauchmann M L Georges J Yoshihiro A Tillery S I H Tyler W J 2010 Transcranial pulsed ultrasound stimulates intact brain circuits Neuron 6668194681–9410.1016/j.neuron.2010.05.00820547127 · doi ↗ · pubmed ↗
- 6Verhagen L et al 2019 Offline impact of transcranial focused ultrasound on cortical activation in primates Elife 8e 4054110.7554/e Life.4054130747105 PMC 6372282 · doi ↗ · pubmed ↗
- 7Sarica C et al 2022 Human studies of transcranial ultrasound neuromodulation: a systematic review of effectiveness and safety Brain Stimul.1573746737–4610.1016/j.brs.2022.05.00235533835 · doi ↗ · pubmed ↗
- 8Silvanto J Pascual-Leone A 2008 State-dependency of transcranial magnetic stimulation Brain Topography 211101–1010.1007/s 10548-008-0067-018791818 PMC 3049188 · doi ↗ · pubmed ↗