P4HA2 Participates in Pathogenesis of Refractive Error by Regulating Collagen Posttranslational Modification and Extracellular Matrix Balance
Yanling Liu, Shanshan Dong, Furong Huang, Ruotong Li, Xi He, Yizheng Jiang, Qi Tian, Haofeng Xiong, Liangyun Jin, Qiongsi Wang, Lu Xia, Jingjing Chen, Huidan Wu, Hui Guo, Xiangtian Zhou, Kun Xia, Zhengmao Hu

TL;DR
This study shows that the P4HA2 gene plays a key role in causing refractive errors like myopia by affecting collagen and extracellular matrix balance in the eye.
Contribution
The study reveals a novel mechanism by which P4HA2 influences myopia through collagen hydroxylation and ECM imbalance.
Findings
P4HA2 knockout mice showed impaired visual acuity and disrupted collagen fibril arrangement in the sclera and cornea.
Reduced collagen hydroxylation in P4HA2 knockout models led to decreased thermal stability and ECM imbalance.
Elevated fibronectin and reduced Collagen I levels were observed in P4HA2 knockout models.
Abstract
Myopia represents a refractive anomaly characterized by impaired vision resulting from a misfocused image in front of the fovea. Although numerous genes linked to high myopia (HM) have been identified, the exact etiology and pathogenesis mechanisms of HM remain predominantly obscure. In a prior investigation, a mutation in the P4HA2 gene was identified in association with HM. To illuminate the potential mechanisms of action of P4HA2 in HM, we established a P4HA2‐knockout mouse line (P4ha2 −/−) and a P4HA2‐knockout HEK293 cell line for this study. P4ha2 −/− mice exhibited compromised visual acuity and altered light transmission pathways as evidenced by multiple biometric assessments. Furthermore, we observed a time‐dependent disruption in the arrangement of collagen fibrils in the sclera and cornea of the P4ha2 −/− mice, attributed to diminished thermal stability due to decreased…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
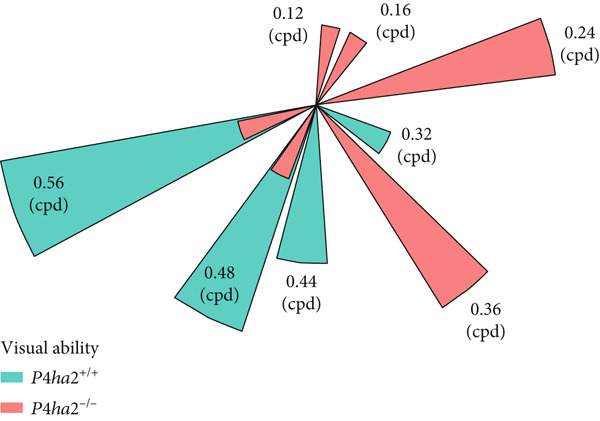 Figure 1
Figure 1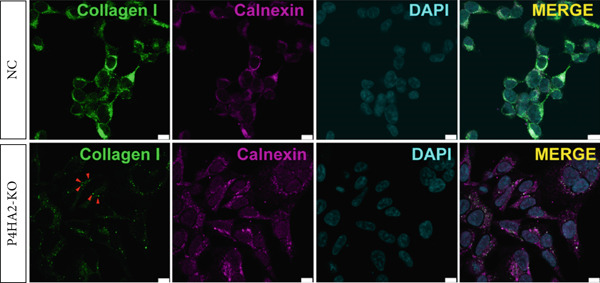 Figure 2
Figure 2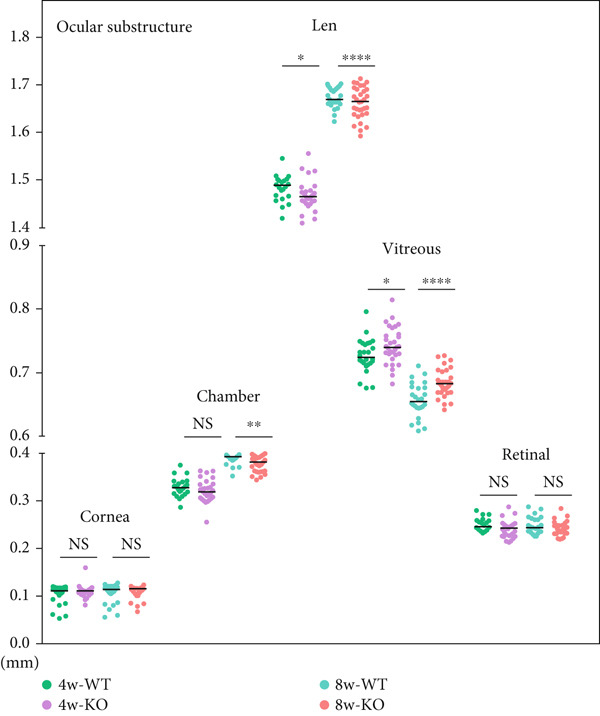 Figure 3
Figure 3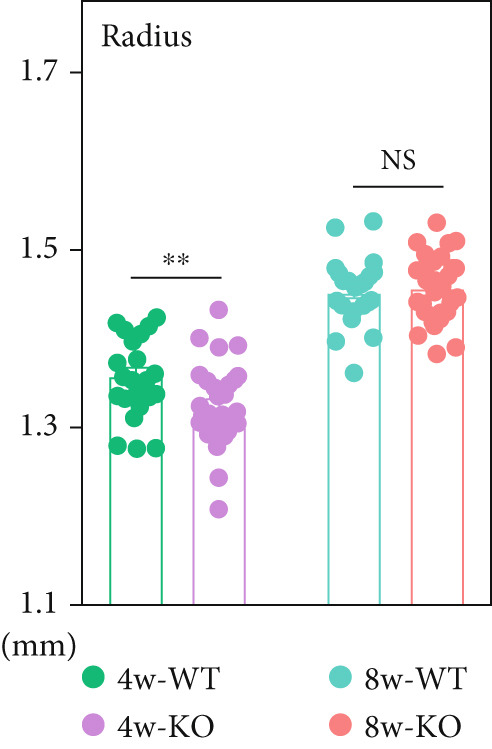 Figure 4
Figure 4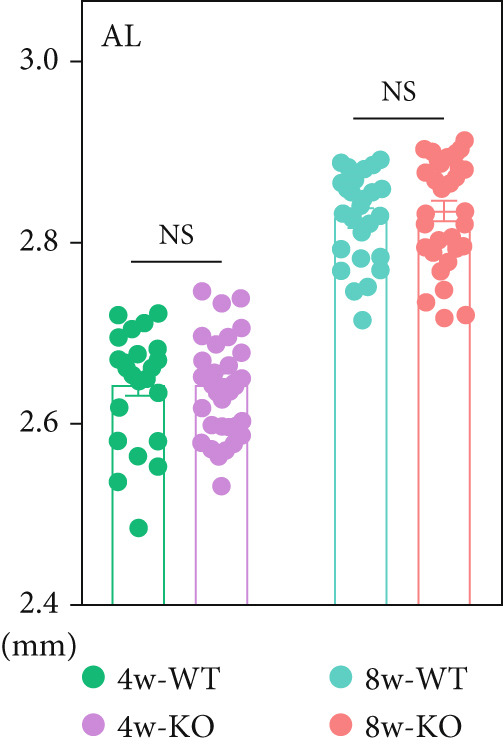 Figure 5
Figure 5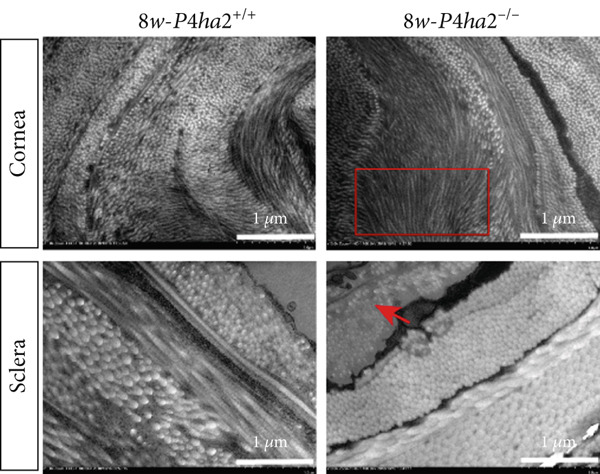 Figure 6
Figure 6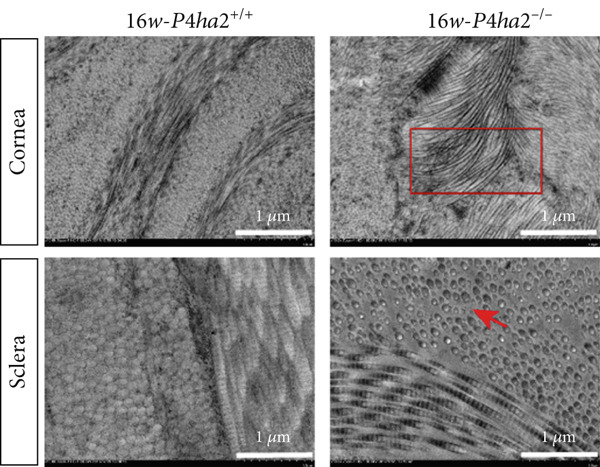 Figure 7
Figure 7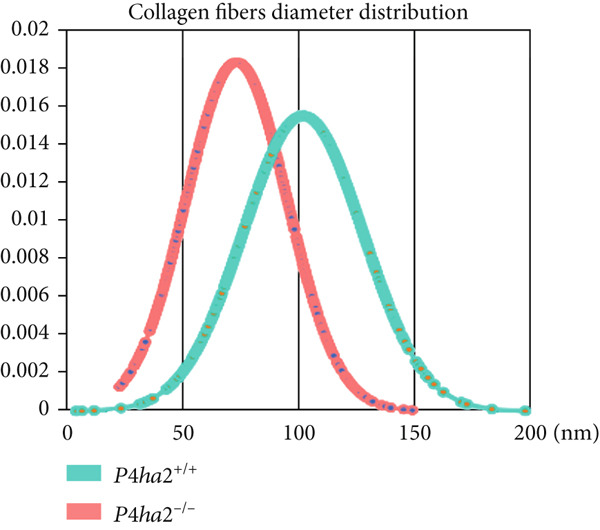 Figure 8
Figure 8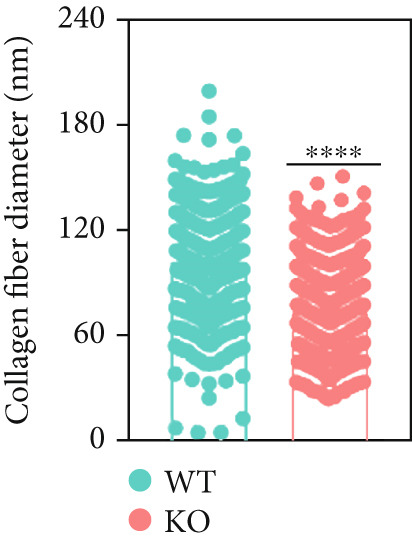 Figure 9
Figure 9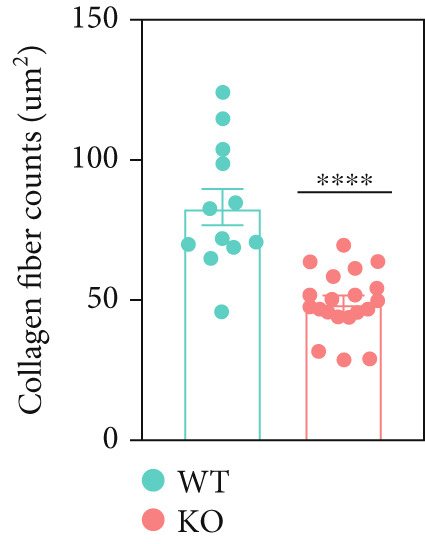 Figure 10
Figure 10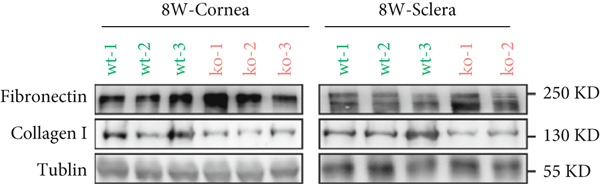 Figure 11
Figure 11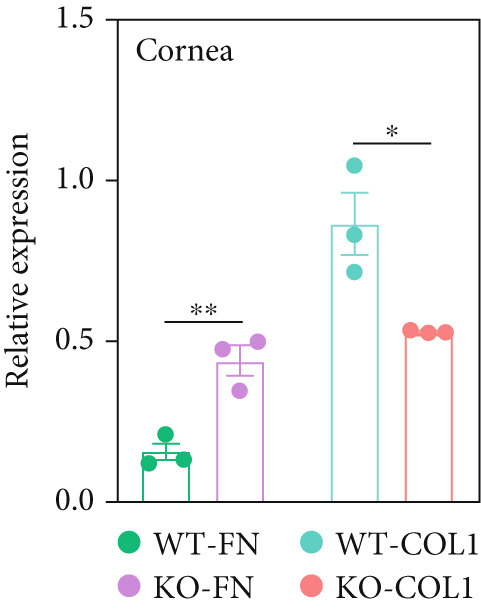 Figure 12
Figure 12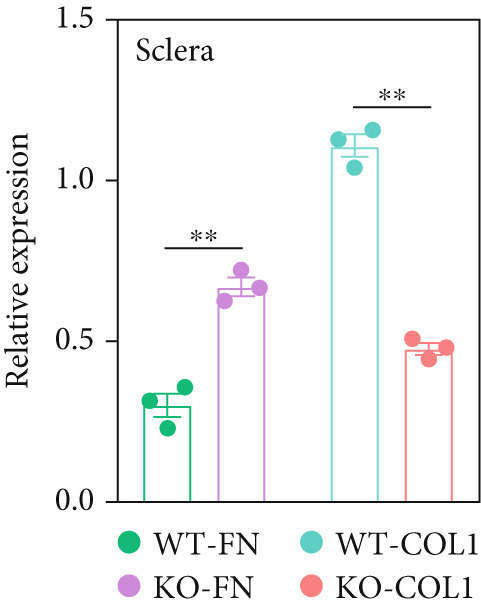 Figure 13
Figure 13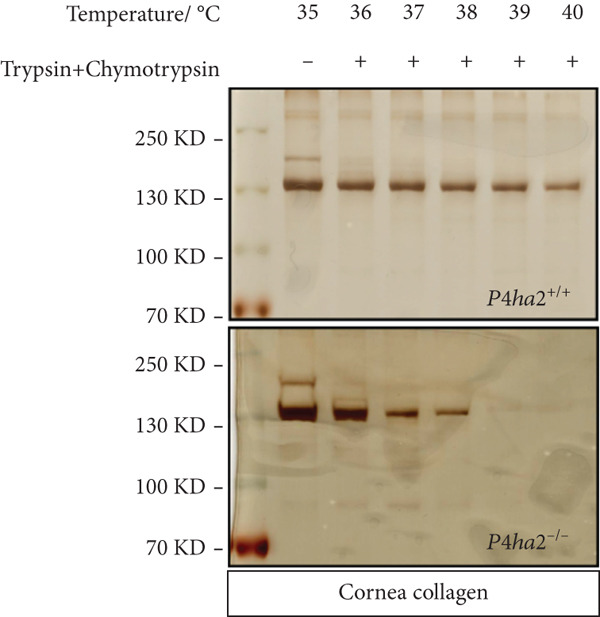 Figure 14
Figure 14 Figure 15
Figure 15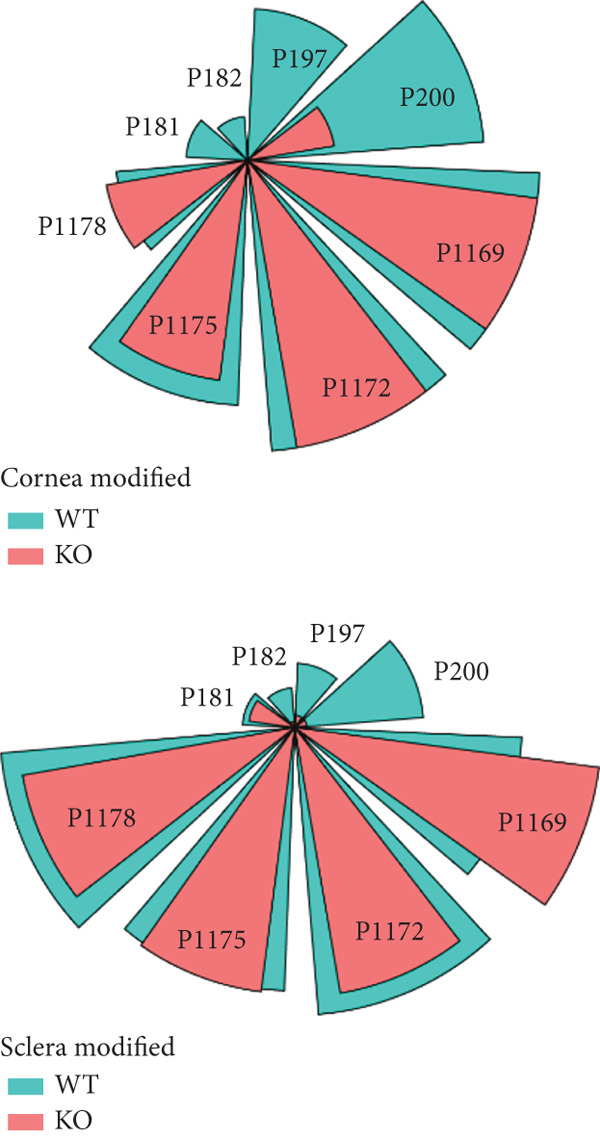 Figure 16
Figure 16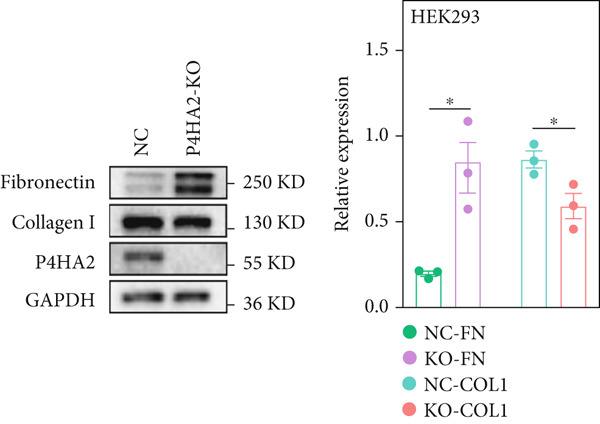 Figure 17
Figure 17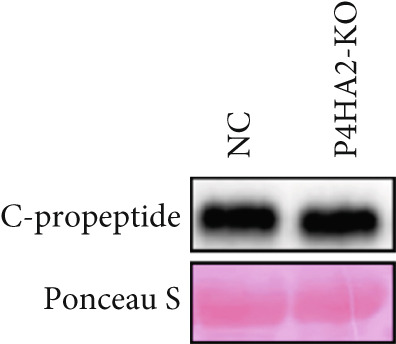 Figure 18
Figure 18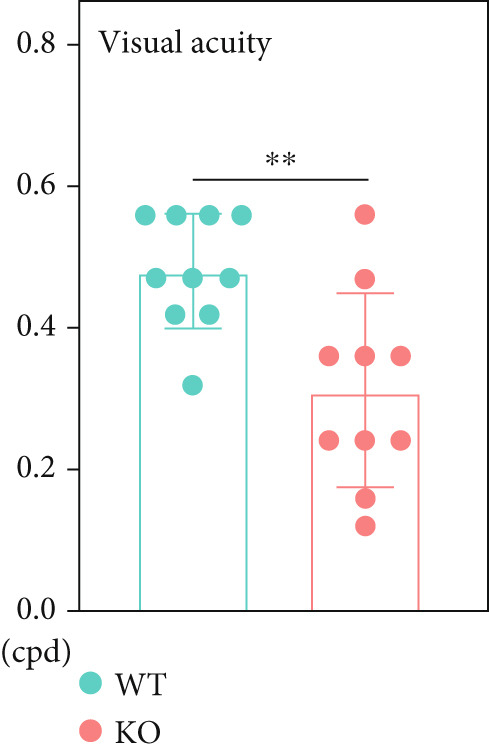 Figure 19
Figure 19- —National Natural Science Foundation of China10.13039/501100001809
- —National Key Research and Development Program of China10.13039/501100012166
- —Natural Science Foundation of Hunan Province10.13039/501100004735
- —Guangzhou Basic and Applied Basic Research Scheme
- —George Mason University10.13039/100006369
- —Department of Education of Zhejiang Province10.13039/501100008867
- —Hangzhou Medical College
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCorneal surgery and disorders · Ocular and Laser Science Research · Connective Tissue Growth Factor Research
1. Introduction
Myopia represents a prevalent ocular condition with a growing global incidence. Predictions suggest that myopia′s prevalence will escalate to 49.8%, with 20% of cases classified as high myopia (HM) [1]. This classification is based on a recent World Health Organization report, which defines myopia as having a diopter of less than −0.5 in either eye, whereas HM is typically defined as having less than −6.0 diopters in most scholarly investigations. HM is distinguished by an elongated axial length (AL), a thinner sclera, and a progressively compromised visual acuity. Among the myriad complications associated with HM, retinal detachment and macular degeneration are the most prevalent. Notably, HM stands as the foremost cause of irreversible blindness in East Asia [2].
The etiology of myopia is complex, stemming from a combination of environmental and genetic factors, although early research has primarily emphasized environmental influences. Generally, prolonged periods of reading in an improper posture have been identified as a significant contributor to myopia onset in most individuals [3]. Furthermore, the risk of myopia appears to heighten when visual impairment manifests during crucial stages of visual development. Recent investigations have underscored a familial predisposition to myopia, particularly among patients with HM. Twin studies have indicated that HM heritability ranges from 50% to 90% [4, 5]. Moreover, advancements in genetic analysis, such as linkage analysis and next‐generation sequencing, have pinpointed several genes associated with HM, including BSG [6], P4HA2 [7, 8], LEPREL1 [9, 10], ZNF644 [11, 12], SLC39A5 [13], VIPR2 [14] and ARR3 [15].
Research into the pathogenesis of HM has predominantly focused on extracellular matrix (ECM) remodeling and degeneration, particularly within the sclera, given the observed thinner sclera and elongated AL in both HM patients and animal models induced with concave lenses [16, 17]. Previous studies have identified mutations in collagen posttranslational hydroxylation modification genes, including P4HA2 [7, 8] and LEPREL1 [9, 10], within the HM pedigree as well as in sporadic cases. Additionally, various signaling pathways involved in ECM regulation have been implicated in HM pathogenesis. For instance, reduced expression of SMAD1, a key regulator of the TGFβ signaling pathway, has been noted in HM patients with SCL39A5 mutations [13, 18]. Furthermore, the HIF pathway has been implicated in HM development through its influence on Collagen I expression, a crucial ECM component [19]. Although ECM abnormalities are widely recognized as the underlying cause of HM, it′s noteworthy that thinner sclera have only been observed in specific mouse models, such as P3h2 ^−/−^ mice [20]. Conversely, in mice with modified HM‐associated genes like Bsg ^+/G901A^ [6] and Zfp644 ^−/−^ [21] mice, ECM abnormalities were not reported.
In our prior investigation, we identified mutations in the P4HA2 gene within the HM pedigree, as well as in sporadic cases, indicating a robust correlation between P4HA2 and the onset of HM. In this current study, we established P4ha2 ^−/−^ mice and P4HA2 knockout cell lines to examine ocular phenotypes and ECM remodeling. These models were utilized to provide additional insights into the pathogenesis of HM and refractive error.
2. Materials and Methods
2.1. Animals
All animal experiments were conducted in strict compliance with the Animal Care and Use Committee guidelines of Central South University and received ethical approval for research involving animals from the same institution.
Knockout C57BL/6 mice for the P4HA2 gene were generated by CRISPR‐Cas9 technology, specifically targeting a 438 bp segment around Exon 5 (Figure S8). Expression of P4HA2 in eye ball had also been detected (Figure S9). The animals were housed in cages under controlled conditions, including a temperature of 25°C, a 12:12 light–dark cycle, and access to food and water available.
2.2. Water Visual Test
The water visual test was performed according to established protocols, using primary thresholds to assess the visual abilities of mice [22]. Briefly, the assessment was carried out in a trapezoid water tank equipped with two monitors displaying various grey gratings, with the highest grating set at 0.56 cycles per degree (cpd). The testing comprised three phases: pretraining, training, and testing. During each phase, water temperature was maintained between 22°C and 24°C, whereas room temperature was kept at 24°C–25°C. Mice that achieved an 80% correction rate during training (16/20 trials) progressed to the testing phase, where a 70% correction was required to advance to higher gratings. Grating limitations were recorded when correction rates fell below 70%.
2.3. Biometric Measurements
For the mice recruited, we tracked the developments of eyes by measuring biometric indexes in 4‐week age and 8‐week age. Noninvasive and noncontacting measurements were prior. Refractive parameters were measured in a darkened room with an eccentric infrared photorefractor (EIR). For each mouse, both eyes were tested for three times to obtain a mean value. Then the mice were anesthetized by intraperitoneal injecting a mixture of ketamine (12%) and xylazine hydrochloride (0.53%). The axial components of the eye parameters, including anterior chamber depth (ACD), lens thickness (LT), vitreous chamber depth (VCD), and AL, cornea thickness, retina thickness and cornea radius, were measured using a custom‐made optical coherence tomography (OCT) [22]. Normal OCT was conducted to observe the retina structure using Phoenix MICRON. After 24 h dark adaption and 10 min dilation with atropine (1%), electroretinogram (ERG) was conducted using RETI‐animal—Roland Consult.
2.4. Transmission Electron Microscope
Eye balls were deprived from randomly chosen eyes of mice and immersed into ice 2.5% glutaraldehyde solution. For each group, three mice were sacrificed. Dissection was conducted under stereoscope. Cornea, sclera, and retina tissues were obtained and incubated in ice 2.5% glutaraldehyde solution for 4 h and then washed with ice PB buffer for five times: 2% osmic acid was added into each tissue and then washed with ice ddH_2_O for five times. Diverse concentration of ethanol (50%, 70%, 80%, 90%, 100%) were used to replace the water and SPI‐Pon 812 embedding set were used to embed tissues as described before [23]. Leica EM UC7 ultramicrotome was used to obtain 70 nm slices and classical staining with 2% uranyl acetate and lead citrate were performed [24]. All the TEM images were captured using HITACHI 7700 TEM.
2.5. Thermal Stability Test
Collagens were isolated from 8‐week mice using 1% pepsin (in 3% acetic acid) digestion and NaCl precipitation. Collagen solution was prepared and neutralized with NaOH solution. The concentration of collagen solution was identified using BCA protein assay kit and a total of 70 μg was prepared for each sample. Thermal stability test was performed as described before by using trysin–chymotrypsin double digestion [25]. Digested samples were separated using SDS‐PAGE gels. Sliver stain kits were used to display the results.
2.6. LC‐MS/MS Spectrum
Collagens were isolated from mice cornea and sclera, and precipitated with 4 M NaCl solution as described above. LC‐MS/MS spectrum procedures were conducted as reported [26]. Fully digested trypsin/chymotrypsin collagen solutions were vacuumed and rehydrated. Electrospray MS was performed on the tryptic peptides using an LCQ Deca XP ion trap mass spectrometer equipped with in‐line liquid chromatography (LC) by a ChromXP C8 capillary column (3 μm, 120 Å) eluted at 4.0 μL/min. Liquid phase was conducted by nano LC 415 (SCIEX, Concord) with C18 reversed‐phase chromatography columns (75 μm × 15 cm C18‐3 μm 120 Å, Chrom XP Eksigent). Mass spectrum was performed in Triple TOF 6600 system (SCIEX, Concord, Ontario, Canada) equipped with Nanospray III ion source (SCIEX, United Stateas). Spray voltage is 2.4 kV, air curtain pressure is 35PSI, atomization pressure is 12PSI, and heating temperature is 150°C. The scan time of single‐level TOF‐MS single map is 250 ms and up to 40 second‐level maps with charge 2^+^ to 4^+^ and single‐second count greater than 260 cps can be collected under each IDA cycle.
Protein sequences used for MS analysis were obtained from the Uniprot genome database. PEAKS Studio 8.5 (Version 8.5, Bioinformatics Solutions Inc. Waterloo, Canada) was used to analysis the results. The mass error of primary mass spectrometry is 20 ppm, the mass error of secondary mass spectrometry is 0.5 Da, and the false positive rate of peptides is controlled to 5% FDR.
2.7. P4HA2 Kncokout HEK293 Cell Line Generation
P4HA2‐kncokout HEK293 cell line was generated by lentivirus packed CRISPR Cas9 system with LentiCRISPR v2 plasmid inserting with 5 ^′^‐CACCGTAGCCGCTTCACCAGTTTGT‐3 ^′^ and 5 ^′^‐AAACACAAACTGGTGAAGCGGCTAC‐3 ^′^. One μg/mL puromycin was used to eliminate all the untransfected cells. A total of 20 transfected cells were cultured in a 10 cm dish and cultured for 7 days. Subclones were picked and then cultured for another 7 days to maintain enough cells. By PCR (F: 5 ^′^‐ CTGGCCTGGTTACTTCATGC ‐3 ^′^, R: 5 ^′^‐ CCACTGGGCAACTGGATTAAG ‐3 ^′^) and immunoblotting, P4HA2‐kncokout HEK293 cell line was identified.
2.8. Immunoblotting and Immunofluorescence
For immunoblotting, the protein aliquots including cornea, sclera, and cell lysates, were resolved in NP40 lysis buffer, were separated by gel electrophoresis in 10% polyacrylamide mini gels and were transferred onto a polyvinylidene difluoride membrane. Western blot analysis of the protein levels was performed using antibodies including Collagen I (ab260043, Abcam), Fibronectin (ab2413, Abcam), β‐actin (ab8226, Abcam), and GAPDH (ab8245, Abcam). After incubation with antibodies overnight in 4°C, secondary antibodies, including goat anti‐rabbit (115‐035‐146, Jackson ImmunoResearch) and goat anti‐mouse IgG (111‐035‐144, Jackson ImmunoResearch) were applied. West Femto Maximum Sensitivity Substrate (34094, Thermofisher) were used to visualize the results. Biorad ChemiDoc system was used for imaging. Image J was used to export grey density analysis.
For immunofluorescence, samples were fixed with 4% PFA and incubated with 5% BSA (0.1% PBST) for 1 h in room temperature, followed by incubation with certain antibodies, including Collagen I (ab34710, Abcam) and Collagen II (ab34712, Abcam), for 16 h. Secondary antibodies, including anti‐rabbit 488 (A‐11070, Thermofisher) and anti‐mouse cy3 (A‐21427, Thermofisher).
The corneal samples, Western blot analyses were performed using three biological replicates per group (3 vs. 3). For the scleral samples, three WT and two KO biological replicates were included. For samples in Figure 4, For Figure 4, we used data from three biological replicates.
2.9. Statistical Analysis
Data analysis was conducted using GraphPad statistical software [27], employing one‐way ANOVA and unpaired t‐tests to determine statistical significance (p < 0.05).
3. Results
3.1. P4ha2
−/− Mice Displayed Impaired Visual Acuity
In this study, preliminary thresholds were used to assess visual acuity. Eight‐week‐old male mice were selected for the test. Following the pretraining phase, all mice in the testing phase demonstrated correction rates exceeding 80% (16 out of 20 trials) during training. During the testing phase, both P4ha2 ^+/+^ and P4ha2 ^−/−^ mice groups (n = 10 each) were evaluated. The median visual acuity for each group was 0.47 and 0.3 cpd, respectively (Figure 1a). The average visual acuity of P4ha2 ^+/+^ mice was significantly higher than that of P4ha2 ^−/−^ mice (p < 0.01) (Figure 1b).
Figure 1 P4ha2 ^−/−^ mice displayed compromised visual acuity and altered eye structure. (a) Visual acuity of P4ha2 ^+/+^ (wild type, WT) mice and P4ha2 ^−/−^ (knockout, KO,) mice (N = 10); (b) Average visual acuity of the WT and KO mice; (c) corneal thickness, anterior chamber depth, lens thickness, vitreous depth, and retina thickness (H) of 4‐ and 8‐week‐old mouse eyes (N_WT_ = 14, N_KO_ = 16); corneal radius (d) and axial length (e) of 4‐ and 8‐week‐old mouse eyes, measured using optical coherence tomography (N_WT_ = 14, N_KO_ = 16); ∗p < 0.05; ∗∗p < 0.01, ∗∗∗p < 0.001, by unpaired t‐test.(a)(b)(c)(d)(e)
3.2. P4ha2
−/− Mice Displayed Abnormal Eye Structure
Using custom‐made OCT, we meticulously tracked the dynamic evolution of major ocular components in both P4ha2 ^+/+^ and P4ha2 ^−/−^ mice (n = 13 and 16, respectively) across various ages (4 and 8 weeks). By the 4‐week mark, P4ha2 ^−/−^ mice exhibited significantly reduced corneal radius (p < 0.01) and increased vitreous depth (p < 0.05). Additionally, a decrease in LT (p < 0.05) and ACD (p = 0.1033) was noted in P4ha2 ^−/−^ mice. Although P4ha2 ^−/−^ mice tended to have thicker corneas than P4ha2 ^+/+^ mice, this disparity did not reach statistical significance (p = 0.1248) (Figure 1c and Figure 1d). However, the difference in AL between the two groups remained statistically insignificant (p = 0.7943) (Figure 1e).
By the age of 8 weeks, P4ha2 ^−/−^ mice displayed deeper ACD (p < 0.01) and longer VCD compared with P4ha2 ^+/+^ mice (p < 0.0001). Other parameters, such as AL and corneal thickness, maintained consistency with those observed in 4‐week‐old mice, albeit without statistical significance (Figures 1c, 1d, and 1e). Additionally, we observed a lower central corneal refraction in 8‐week‐old P4ha2 ^−/−^ mice, as determined by EIR (Figure S1a).
To assess potential impairments in retinal structure and function among P4ha2 ^−/−^ mice, we conducted an ERG test on 8‐week‐old mice. Photopic and scotopic A and B waves, along with scotopic oscillatory potentials, did not exhibit significant differences between the two groups (Figures S1b, S1c, and S1d). Furthermore, examination of various retinal layers in 8‐week‐old P4ha2 ^−/−^ mice revealed normal structure (Figure S2). Thus, both the retinal structure and function of P4ha2 ^−/−^ mice appeared to be relatively intact.
3.3. P4ha2
−/− Mice Displayed Progressively Reduced Collagen Amount in Sclera and Cornea
At the age of 8 weeks, one eye from both P4ha2 ^+/+^ and P4ha2 ^−/−^ mice was randomly selected for examination of corneal and scleral ultrastructure (N = 3). Comparing with the P4ha2 ^+/+^ counterparts, P4ha2 ^−/−^ mice exhibited disorganized collagen fibrils in the cornea and thinner collagen fibrils in the outer layer of the sclera (Figure 2a). To elucidate time‐dependent degenerative processes, one eye from 16‐week‐old P4ha2 ^+/+^ and P4ha2 ^−/−^ mice (N = 3) was randomly chosen for ultrastructural analysis of the cornea and sclera. At the end of 16 weeks, there were no discernible changes in the 16‐week‐old P4ha2 ^+/+^ mice. However, strikingly abnormal collagen fibrils were evident in the 16‐week‐old P4ha2 ^−/−^ mice. Notable disarrangement was observed in the corneal structure of the 16‐week‐old P4ha2 ^−/−^ mice, accompanied by disturbed and thinner collagen fibrils in the sclera (Figure 2b). The number (Figure 2c,d) and diameter (Figure 2e) of fibrils were measure within randomly chosen 1 μm^2^ area were conducted and notably decreased in the sclera of 16‐week‐old P4ha2 ^−/−^ mice was observed. Although a lack of OCT data at 16 weeks prevents definitive age‐matched morphological correlation, the ultrastructural findings still support the possibility of refractive phenotype development.
Figure 2. Transmission electron microscopy results and extracellular matrix (ECM) component imbalance in prolyl 4‐hydroxylase II–null mice. (a–b) The cornea and sclera ultrastructure of (a) 8‐ and (b) 16‐weeks‐old mice, the red rectangle indicates misarrangement of collagen fibrils in the cornea, and the red arrows indicates sparse fibrils in sclera, the scale bar indicates 10 μm; (c) the distribution of the diameter of collagen fibrils in 16‐weeks‐old WT and KO sclera (fibrils N_WT_ = 1003, fibrils N_KO_ = 991); (d) the average diameter of collagen fibrils in WT groups was higher than in KO groups (fibrils N_WT_ = 1003, fibrils N_KO_ = 991); (e) the density of collagen fibrils was higher in WT groups; (f) fibronectin and Collagen I expression in the cornea and sclera tissues of 8‐week‐old P4ha2 ^−/−^ mice mice; and (g) and (h) represent the gray density of each lane; ∗p < 0.05; ∗∗p < 0.01, ∗∗∗p < 0.001, by unpaired t‐test.(a)(b)(c)(d)(e)(f)(g)(h)
Further assessment involved examining the expression levels of Collagen I and fibronectin in corneal and scleral tissues from 8‐week‐old mice. We observed decreased Collagen I and increased fibronectin levels in both corneal (Figure 2f,g) and scleral (Figure 2f,h) tissues, indicating an imbalance in the ECM.
3.4. Collagen Thermal Stability Was Reduced Due to the Lack of Proline Hydroxylation in Non‐(GPP)n Position P181, P182, P197, and P200
As proline hydroxylation is responsible for collagen thermostability, we performed a thermal stability test on collagens (mainly Collagen I) extracted from mouse cornea using trypsin–chymotrypsin digestion at various temperatures. Compared with those in the control, which maintained thermal stability even at 38°C, corneal collagens from *P4ha2^−/−^
- mice degenerated at 37°C (Figure 3a). This result indicates a reduction in hydroxylation; thus, we performed LC‐MS/MS to confirm proline hydroxylation in collagens. As the conserved (GPP)n domain is essential for collagen stability, we analyzed the 4‐hydroxyprolin (4Hyp) condition of the Collagen I α1 chain, which was also abundant in all tissues. The results showed that there was no change in *P4ha2^−/−^
- mice (Figure 3b and Figure S3). However, the other 4Hyp of the Collagen I α1 chain in P181, P182, P197, and P200 were reduced in the cornea and sclera (Figure 3b,c and Figures S4a, S4b, S4c, and S4d). We also observed multiple reductions of 4Hyp in other types of collagens (Table S1–S3). In summary, the 4Hyp position, which is crucial for collagen stability, remained unchanged, but other positions that were less bone‐related important showed reductions, which were consistent with the lack of osteogenesis imperfecta in patients with P4HA2 mutations.
Figure 3. Thermal stability of and 4Hyp in collagen. (a) The thermal stability of Collagen I was reduced in P4ha2 ^−/−^ corneal collagen isolated from 8‐week‐old mice; (b) (GPP)n and EPG in Collagen I; and (c) the 4Hyp in highly conserved (GPP)n remained unchanged in the Collagen I α1 chain of the cornea and sclera at positions P1169, P1172, P1175, and P1178. However, in Collagen I α1 chain positions P181, P182, P197, and P200, 4Hyp displayed a reduction in P4ha2 ^−/−^ mice.(a)(b)(c)
3.5. ECM‐Associated Proteins Showed Imbalance and Accumulation in Prolyl 4‐Hydroxylase II–Null Cells
To investigate how the lack of P4HA2 induces thinner collagen fibrils, as observed using TEM, we generated a P4HA2‐knockout HEK293 cell line (Figure S5). First, we examined the expression of Collagen I and fibronectin in the HEK29 knockout cell lines. The results showed decreased Collagen I and increased fibronectin levels (Figure 4a). The content of C‐propeptide of Collagen I in the culture supernatant remained unchanged (Figure 4b), which indicated that the decrease in Collagen I level in the *P4HA2-*knockout HEK293 cells was mainly caused by degeneration induced by the lack of 4Hyp. Furthermore, using immunofluorescence, we observed that Collagen I accumulated in the endoplasmic reticulum of cells (Figure 4c). Collagen II also showed similar accumulation in the knockout cells (Figure S6). However, these accumulations were not colocalized with KLHL12, a component of the collagen transporter Coat Protein II complex (Figure S7), which indicated that this might not be a transportation‐related problem.
Figure 4. Extracellular matrix (ECM) alteration in prolyl 4‐hydroxylase II–null HEK293 cells. (a) Fibronectin and Collagen I expression in the prolyl 4‐hydroxylase II–null HEK293 cell line (KO); (b) c‐propeptide of Collagen I remained unchanged; and (c) abnormal Collagen I (green) accumulation (red arraow) in the endoplasmic reticulum (red) in P4HA2 HEK293 KO cells. The scale bar indicates 75 μm, ∗p < 0.05; ∗∗p < 0.01, ∗∗∗p < 0.001, by unpaired t‐test.(a)(b)(c)
4. Discussion
This study involved the generation of P4ha2 ^−/−^ mice characterized by impaired visual acuity, mirroring the phenotypic traits observed in patients with HM. Previous studies had showed no apparent phenotypic abnormalities in P4ha2 ^−/−^ mice [25]. Extensive optical assessments demonstrated that P4ha2 ^−/−^ mice exhibited atypical eye structures while maintaining normal retinal structure and function. Furthermore, ultrastructural analysis unveiled thinner and disrupted collagen fibrils within the sclera of P4ha2 ^−/−^ mice.
Approximately 22% of amino acids found in collagens consist of proline or hydroxylated proline, a composition that mitigates the entropy cost associated with collagen folding [28]. Collagens feature (ProHypGly)n and Gly‐Xaa‐Yaa repeats, with the Y position typically undergoing hydroxylation catalyzed by 4‐proline‐hydroxylase [29]. When the Y position is lysine, it can also be hydroxylated by lysyl hydroxylases by Lysyl hydroxylases such as PLODs [30]. This enzymatic process, particularly targeting Hyp at the Y position, enhances the thermal stability of collagens [31, 32]. Notably, the hydroxylation of (GPP)n repeats exhibits evolutionary conservation across species [33], exerting a more significant impact compared to other hydroxyprolines like P181 [28].
In our investigation, we observed no alterations in 4Hyp within (GPP)n repeats in the Collagen I α1 chain, the predominant collagen chain in mice. However, the loss of hydroxyproline seemed uneven in all collagen. We identified almost complete loss of 4Hyp in P197 and P200, which were all EPG sequences which coincident with recent observation [34]. This finding also aligns with the observed normal skeletal and muscle phenotypes in P4ha2 ^−/−^ mice [25, 35]. We also assumed that other 4Hyp change in Collagen I α2 chain, Collagen III α1 chain, Collagen V, and so on. might also contribute to thermal instability. The resultant instability of collagen fibrils at physiological temperature could lead to progressive degeneration over time, potentially contributing to the worsening vision impairment observed in HM patients.
Recent studies have highlighted the significant involvement of collagens in the pathogenesis of myopia [36]. In our research, we observed a reduction in Collagen I levels and an increase in fibronectin levels within corneal and scleral tissues, as well as in P4HA2‐knockout cell lines. This finding corresponded with the sparse collagen fibrils distribution observed in TEM results. Such an imbalance in Collagen I and fibronectin content could profoundly impact the refractive properties of the cornea. Notably, the refractive index of the human cornea, which ranges from 1.401 to 1.373 [37], is primarily influenced by Collagen I, having a refractive index of 1.43 [38]. The observed decrease in Collagen I and concurrent increase in fibronectin levels would undoubtedly alter the refractive properties of the cornea, thereby affecting the path of light transmission. Additionally, changes such as a shorter ACD and longer vitreous depth would further contribute to alterations in the light path.
It seemed that longer AL was not shown in every gene editing mouse model and every HM patient. Several HM‐associated gene‐modified mouse models have also been reported. In Vipr2 ^−/−^ mice [14], AL showed no statistically increased, however, alterations in refractive elements were identified, which was similar to what we observed in P4ha2 ^−/−^ mice. Other animal models, including Bsg ^+/G901A6^, Zfp644 ^−/−21^, and Zfp644 ^S673G/S673G21^ mice as well as P4ha2 ^−/−^ mice, displayed normal retinal function, but altered refractive elements. Longer AL was only reported in Zfp644 ^−/^ and Zfp644 ^S673G/S673G^ mice. In Bsg ^+/G901A^ mice, researches only observed increasing ΔAL. In P3ha2 ^−/−^ mice [20], researchers reported thinner sclera but no clue of AL.
As what we observed in patients with Cohen syndrome [39] show HM phenotypes without longer AL, disturbed and misarranged collagen fibrils and decreased Collagen I in the P4ha2 ^−/−^ mouse cornea resulted in refractive index changes in the cornea, which also leading to changes in the light path. In summary, although the AL of P4ha2 ^−/−^ mice remained unchanged, the refractive error observed in P4ha2 ^−/−^ mice could largely be attributed to abnormal refractive systems and deviated light paths.
In conclusion, P4HA2 contributes to the pathogenesis of refractive error by changing the light pathways by altering the ratio of refractive elements and ECM components. Furthermore, the progressive aggravation of refractive error might also be explained by time‐dependent collagen reduction caused by the lack of P4HA2. Our study presents that P4HA2 participates in pathogenesis of HM by regulating collagen posttranslational modification and ECM balance.
NomenclatureHMHigh myopia P4ha2 ^−/−^
P4HA2‐knockout mouse lineALAxial lengthECMExtracellular matrixTEMTransmission electron microscopyACDAnterior chamber depthLTLens thicknessVCDVitreous chamber depthOCTOptical coherence tomographyERGElectroretinographyEIREccentric infrared retinoscopy
Conflicts of Interest
The author declares no conflicts of interest.
Author Contributions
Yanling Liu and Shanshan Dong contributed equally to this paper. Shanshan Dong is co‐first author.
Funding
This work was supported by the National Natural Science Foundation of China (Grant Number 82271908 to Zhengmao Hu and Grant Number 82130043 to Kun Xia), the National Key R&D Program of China (Grant Number 2021YFA0805202 to Zhengmao Hu), Natural Science Foundation of Hunan Province (Grant Number 2025JJ80300 to Zhengmao Hu), Guangzhou Basic and Applied Basic Research Scheme (2023A04J1175 to Shanshan Dong), Plan on enhancing scientific research in GMU (02‐410‐2302311XM to Shanshan Dong), the general research projects of Zhejiang Provincial Department of Education (Y202456738 to Yanling Liu), and the Basic Research Program Youth Research Fund of Hangzhou Medical College (KYQN2024003 to Yanling Liu).
Supporting Information
Additional supporting information can be found online in the Supporting Information section. Supporting Information files including the following figures and tables:
Supporting information
Supporting Information 1 Figure S1. The results of EIR and ERG test showed no difference among P4ha2^+/+^ (WT) mice and P4ha2^−/−^ (KO) mice. Figure S2. Normal retina structure were observed in P4ha2^+/+^ (WT) mice and P4ha2^−/−^ (KO) mice. Figure S3. Hydroxylation Modified Secondary Mass Spectrogram of Collagen I α1 in position P1169, P1172, P1175, P1178. Figure S4. (a–d) Hydroxylation modified secondary mass spectrogram of Collagen I α1 in position (a) P181, (b) P182, (c) P197, and (d) P200. Figure S5. Identification of P4HA2 KO HEK293 cell line. Figure S6. Abnormal Collagen I (green) accumulated in ER (red) of P4HA2 HEK293 KO cells. Figure S7. Collagen I (green) were not colocalizing with KLHL12. Figure S8. P4ha2 gene knockout mice construction strategies. Figure S9. Expression of P4ha2 in mice eyes.
Supporting Information 2 Table S1. Four Hyp reduction in mice sclera. Table S2. Four Hyp reduction in mice cornea. Table S3. Four Hyp reduction in mice skin.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Holden B. A. , Fricke T. R. , Wilson D. A. , Jong M. , Naidoo K. S. , Sankaridurg P. , Wong T. Y. , Naduvilath T. J. , and Resnikoff S. , Global Prevalence of Myopia and High Myopia and Temporal Trends From 2000 Through 2050, Ophthalmology. (2016) 123, no. 5, 1036–1042, 10.1016/j.ophtha.2016.01.006, 2-s 2.0-84963943910, 26875007.26875007 · doi ↗ · pubmed ↗
- 2Morgan I. G. , Ohno-Matsui K. , and Saw S. M. , Myopia, Lancet. (2012) 379, 1739–1748, 10.1016/S 0140-6736(12)60272-4, 2-s 2.0-84860511273.22559900 · doi ↗ · pubmed ↗
- 3Dirani M. , Tong L. , Gazzard G. , Zhang X. , Chia A. , Young T. L. , Rose K. A. , Mitchell P. , and Saw S. M. , Outdoor Activity and Myopia in Singapore Teenage Children, British Journal of Ophthalmology. (2009) 93, no. 8, 997–1000, 10.1136/bjo.2008.150979, 2-s 2.0-68049102939, 19211608.19211608 · doi ↗ · pubmed ↗
- 4Peet J. A. , Cotch M. F. , Wojciechowski R. , Bailey-Wilson J. E. , and Stambolian D. , Heritability and Familial Aggregation of Refractive Error in the Old Order Amish, Investigative Ophthalmology & Visual Science. (2007) 48, no. 9, 4002–4006, 10.1167/iovs.06-1388, 2-s 2.0-35148831622, 17724179.17724179 PMC 1995233 · doi ↗ · pubmed ↗
- 5Wojciechowski R. , Congdon N. , Bowie H. , Munoz B. , Gilbert D. , and West S. K. , Heritability of Refractive Error and Familial Aggregation of Myopia in an Elderly American Population, Investigative Ophthalmology & Visual Science. (2005) 46, no. 5, 1588–1592, 10.1167/iovs.04-0740, 2-s 2.0-20444491538.15851555 PMC 3092734 · doi ↗ · pubmed ↗
- 6Jin Z. B. , Wu J. , Huang X. F. , Feng C. Y. , Cai X. B. , Mao J. Y. , Xiang L. , Wu K. C. , Xiao X. , Kloss B. A. , Li Z. , Liu Z. , Huang S. , Shen M. , Cheng F. F. , Cheng X. W. , Zheng Z. L. , Chen X. , Zhuang W. , Zhang Q. , Young T. L. , Xie T. , Lu F. , and Qu J. , Trio-Based Exome Sequencing Arrests de Novo Mutations in Early-Onset High Myopia, Proceedings of the National Academy of Sciences of the United States of America. (2017) 114, no. 16 · doi ↗ · pubmed ↗
- 7Guo H. , Tong P. , Liu Y. , Xia L. , Wang T. , Tian Q. , Li Y. , Hu Y. , Zheng Y. , Jin X. , Li Y. , Xiong W. , Tang B. , Feng Y. , Li J. , Pan Q. , Hu Z. , and Xia K. , Mutations of P 4HA 2 Encoding Prolyl 4-Hydroxylase 2 Are Associated With Nonsyndromic High Myopia, Genetics in Medicine. (2015) 17, no. 4, 300–306, 10.1038/gim.2015.28, 2-s 2.0-84926482761, 25741866.25741866 · doi ↗ · pubmed ↗
- 8Napolitano F. , Di Iorio V. , Testa F. , Tirozzi A. , Reccia M. G. , Lombardi L. , Farina O. , Simonelli F. , Gianfrancesco F. , Di Iorio G. , Melone M. A. B. , Esposito T. , and Sampaolo S. , Autosomal-Dominant Myopia Associated to a Novel P 4HA 2 Missense Variant and Defective Collagen Hydroxylation, Clinical Genetics. (2018) 93, no. 5, 982–991, 10.1111/cge.13217, 2-s 2.0-85043282188, 29364500.29364500 · doi ↗ · pubmed ↗