Gallibacterium anatis Emerging in Indonesia: Isolation and Molecular Characterization From Chickens in a West Java Poultry Farm
Alya Amaliah, Ni Luh Putu Ika Mayasari, Ryan Septa Kurnia, Christian Marco Hadi Nugroho, Muhammad Ade Putra, Agustin Indrawati

TL;DR
This study identifies Gallibacterium anatis in Indonesian chickens, showing it causes respiratory and reproductive issues and has key virulence genes.
Contribution
First molecular evidence of Gallibacterium anatis in Indonesian poultry, including virulence gene detection and phylogenetic analysis.
Findings
14 out of 17 suspected isolates confirmed as G. anatis via molecular methods.
All isolates carried the gyrB virulence gene, with high prevalence of gtxA and flfA.
Sequence similarity of >98.65% with reference G. anatis strains in GenBank confirmed species identity.
Abstract
Gallibacterium anatis (G. anatis) is associated with decreased egg production and respiratory disorders in chickens. The presence of G. anatis in Indonesia has not been reported. This study aimed to identify G. anatis both phenotypically and genotypically in layer, broiler, and breeder chicken from farms in West Java. A total of 23 suspected gallibacteriosis cases were collected from chickens exhibiting respiratory and reproductive disorders between May 2020 and October 2023. Phenotypic characterization was conducted using blood agar culture, Gram staining, and biochemical tests (catalase, oxidase, and TSIA). Genotypic identification of G. anatis was performed using PCR targeting the 16S‐23S rRNA gene, followed by detection of virulence genes (gtxA, flfA, and gyrB). Sequencing and phylogenetic analyses were carried out using BLAST program and MEGA 7. Among the 23 collected samples, 17…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
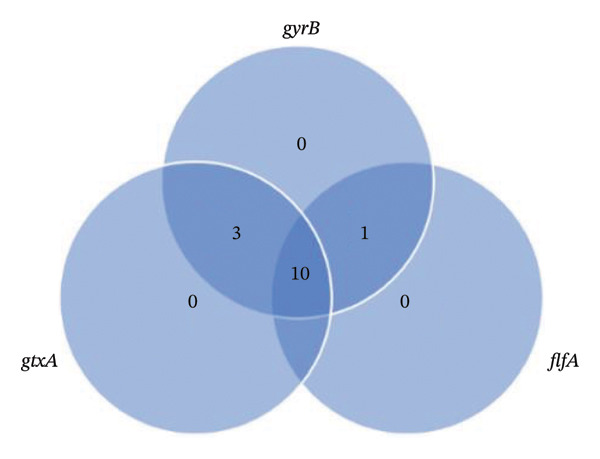 FIGURE 2
FIGURE 2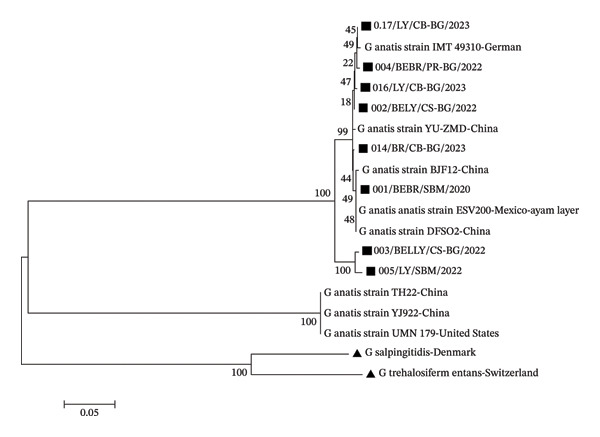 FIGURE 3
FIGURE 3| Target genes | Sequences (5′–3′) | Size (bp) | References |
|---|---|---|---|
|
| F: CAAACCTAATTCAATCGGATG | 1257 | [ |
| R: TGCTTCAATAATTTTCCATTTTC | |||
|
| F: CACCATGGGTGCATTTGCGGATGATCC | 538 | [ |
| R: ATTCGTATGCGATAGTATAGTTC | |||
|
| F: TGTGCGTTTCTGGCCAAGTC | 561 | [ |
| R: CGCTCACCAACTGCAGATTC | |||
| Origin of isolates | Number of samples | Number of phenotypic test | Number of molecular test | |
|---|---|---|---|---|
| Positive | Negative | |||
| Farm A (Breeder‐Layer) | 5 | 5 (100%) | 5 (100%) | 0 (0%) |
| Farm 1 (Layer‐Ciampea) | 3 | 0 (0%) | — | — |
| Farm 2 (Broiler‐Ciseeng) | 3 | 0 (0%) | — | — |
| Farm 3 (Broiler‐Cibinong) | 4 | 4 (100%) | 3 (75%) | 1 (25%) |
| Farm 4 (Layer‐Cibinong) | 4 | 4 (100%) | 3 (75%) | 1 (25%) |
| Farm 5 (Layer‐Cibinong) | 4 | 4 (100%) | 3 (75%) | 1 (25%) |
| Total | 23 | 17 (73.9%) | 14 (82.35%) | 6 (17.64%) |
| Isolates name | Virulence genes | ||
|---|---|---|---|
|
|
|
| |
| 001/BEBR/SBM/2020 | + | + | − |
| 002/BELY/CS–BG/2022 | − | + | + |
| 003/BELY/CS–BG/2022 | + | + | + |
| 004/BEBR/PR–BG/2022 | + | + | + |
| 005/LY/SBM/2022 | + | + | − |
| 012/BR/CB–BG/2023 | + | + | + |
| 014/BR/CB–BG/2023 | + | + | + |
| 015/BR/CB–BG/2023 | + | + | + |
| 016/LY/CB–BG/2023 | + | + | + |
| 017/LY/CB–BG/2023 | + | + | + |
| 018/LY/CB–BG/2023 | + | + | + |
| 020/LY/CB–BG/2023 | + | + | + |
| 021/LY/CB–BG/2023 | + | + | − |
| 022LY/CB–BG/2023 | + | + | + |
| Isolates name | Bacterial references | Access code | Similarity | Query cover |
|---|---|---|---|---|
| 001/BEBR/SBM/2020 |
| 99.38% | 100% | |
| 002/BELY/CS–BG/2022 |
| 99.64% | 100% | |
|
| 99.64% | 100% | ||
| 003/BELY/CS–BG/2022 |
| 99.53% | 100% | |
|
| 99.53% | 100% | ||
|
| 99.53% | 100% | ||
| 004/BEBR/PR–BG/2022 |
| 99.27% | 100% | |
| 005/LY/SBM//2022 |
| 99.73% | 100% | |
|
| 99.73% | 100% | ||
|
| 99.73% | 100% | ||
|
| 99.73% | 100% | ||
|
| CP114281.11 | 99.73% | 100% | |
|
| 99.73% | 100% | ||
| 013/BR/CB–BG/2023 |
| 99.62% | 100% | |
|
| 99.62% | 100% | ||
|
| 99.62% | 100% | ||
|
| 99.62% | 100% | ||
| 016/LY/CB–BG/2023 |
| 99.52% | 100% | |
|
| 99.52% | 100% | ||
|
| 99.52% | 100% | ||
| 017/LY/CB–BG/2023 |
| 99.75% | 100% | |
- —United States Agency for International Development10.13039/100000200
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial infections and disease research · Aquaculture disease management and microbiota · Microbial Metabolism and Applications
1. Introduction
Gallibacterium anatis (G. anatis) is a bacterium that causes gallibacteriosis [1], which plays a role in reducing egg production in laying hens by up to 3%–18%. G. anatis is increasingly recognized as an important pathogen associated with reduced egg production and reproductive tract lesions in laying hens [2]. Infections caused by this bacterium can be primary or secondary [3]. Besides targeting the reproductive organs, this bacterium is also present in various other organs such as the heart, spleen, upper respiratory tract, lower genital tract, and multiple systemic tissues of affected chickens, reflecting its broad tissue tropism [2–5].
Historically, G. anatis was classified as Pasteurella haemolytica; however, in 2003, it was reclassified into a new genus, Gallibacterium [4]. In recent years, reports of G. anatis infections have increased worldwide, indicating its rising clinical importance [2]. A new taxonomy of G. anatis has been identified in several countries [6]. However, there have been no documented reports on the occurrence of G. anatis in Indonesia.
Detection of G. anatis can be performed phenotypically and genotypically. Molecular identification remains essential due to its phenotypic similarity to other respiratory pathogens [2]. Genotypic detection typically uses PCR targeting the 16S and 23S rRNA genes, and identification can be confirmed with MALDI‐TOF; indeed, isolates from reproductive, respiratory, and serosal lesions have been characterized using these methods [4]. Classification using 16S rRNA gene targets is currently widely used in bacterial diagnostics [6]. Genomic analyses have shown considerable diversity among G. anatis isolates, including variation in virulence‐associated genes. Some studies report highly conserved RTX toxin systems as core virulence mechanisms [2, 7], while others identified multiple antibiotic resistance genes varying between organs and flocks [8, 9]. In Morocco, G. anatis was isolated from ovaries, trachea, and cloaca in layer chickens with decreased egg production, indicating its direct impact on productivity [5].
Recent epidemiological studies further underscore the significance of G. anatis. Surveys of poultry flocks in Poland (2022–2023) found a prevalence of 22.5%, with 20% of isolates exhibiting resistance to at least eight antibiotics [10]. All G. anatis biovar haemolytica strains carried the toxin gene gtxA and showed multidrug resistance [9], while additional studies revealed widespread resistance to 20 antimicrobials across respiratory, reproductive, and gastrointestinal isolates [11]. Moreover, G. anatis has been detected in pet birds (∼10.5% prevalence), with isolates showing both high antibiotic resistance and moderate biofilm formation, indicating persistence and treatment challenges [12].
West Java is a province that serves as a center for chicken farming in Indonesia [13], thus serving as a starting point for identifying potentially harmful infectious agents in the poultry industry. Information regarding the presence of G. anatis has not yet been reported in West Java or Indonesia. Therefore, the aims of this study were to isolate and identify the bacterium G. anatis phenotypically and genotypically on chicken farms in West Java, to molecularly characterize the virulence genes of G. anatis, and to analyze the genetic relationship among G. anatis bacteria.
2. Materials and Methods
2.1. Ethical Approval
Ethical approval on this study was obtained from Animal Ethics School of Veterinary Medicine and Biomedical Science, IPB University with number: 121/SKE/X/2023.
2.2. Sample Collection
The research samples consisted of a total of 23 suspected gallibacteriosis cases in West Java from May 2020 to October 2023. The field samples were obtained from several chicken flocks across five different farms that experienced respiratory disorders, accompanied by a 7%–12% reduction in egg production and abnormalities in eggshell formation. All isolates were obtained from tracheal swab, trachea, lung, ovary, and oviduct organs of broilers, layers, and breeder chickens that were suspected of having clinical symptoms of G. anatis infection. Organ samples were placed in sterile plastic, while swab samples were placed in Amies transport medium (Labware), which was then transported using a cold box not exceeding 48 h from sample collection to prevent bacterial growth.
2.3. Isolation and Identification of G. anatis
Samples were obtained from tracheal swabs and reproductive and respiratory organs of chickens exhibiting respiratory disorders or decreased production. Cultures were conducted on blood agar media from archived isolates and field samples, followed by incubation at 37°C for 24–48 h. Colonies were observed macroscopically and bacterial morphology was observed microscopically by Gram staining. Biochemical tests were performed to confirm G. anatis by the catalase test, the oxidase test, and triple sugar iron agar (TSIA) [14].
Isolates exhibiting the phenotypic characteristics of G. anatis were further molecularly confirmed. Confirmation of G. anatis was conducted using the 16S rRNA gene target 1133fgal (5′‐TATTCTTTGTTACCARCGG‐3′) as the forward primer and 23S rRNA 114r (5′‐GGTTTCCCCATTCGG‐3′) as the reverse primer, which yielded a positive result for G. anatis at a band size of 1030 bp [14].
2.4. Detection of Virulence Genes of G. anatis
All samples confirmed as G. anatis by PCR testing were further subjected to the detection of virulence genes of G. anatis bacteria encoding gtxA (cytolytic–hemolytic gene), flfA (flagellar gene), and gyrB (gyrase subunit B). The primer sequences used are listed in Table 1.
2.5. Sequence and Phylogenetic Analyses
Eight isolates of G. anatis were utilized for sequencing analysis using Sanger sequencing with BigDye Terminator V3.1 (Thermo Fisher Scientific GmBH, Dreieich 63303, Germany) [15]. The nucleotide sequence of the G. anatis 16S rRNA gene was analyzed using Basic Local Alignment Search Tool (BLAST) to determine its identity and homology to other isolates in the GenBank database of the National Center for Biotechnology Information (NCBI). Phylogenetic analysis was conducted using the Molecular Evolutionary Genetics Analysis (MEGA) 7 application with the neighbor‐joining method [16]. Phylogenetic trees were constructed by selecting reference isolates with the highest BLAST analysis scores.
3. Results
3.1. Phenotypic and Genotypic Identification of G anatis
A total of 17 (73.9%) of 23 samples showed suspected growth of G. anatis. All of these demonstrated growth as small‐sized, nonmucoid, semitransparent colonies with a distinct hemolytic zone on blood agar (Figure 1). Morphological test results are Gram‐negative and pleomorphic coccobacilli. Biochemical test results were catalase‐positive, oxidase‐positive, and indole‐negative.
FIGURE 1Cultural and microscopic characteristics of Gallibacterium anatis. (a) Growth of G. anatis on blood agar. (b) Gram‐stained G. anatis demonstrating Gram‐negative coccobacilli under 1000× magnification.(a)(b)
Identification of 17 positive G. anatis samples based on phenotypic observation consisted of five sample isolates from a breeder–layer farm, four isolates from broiler farms in Cibinong, and eight isolates from layer farms in Cibinong (Table 2). Molecular confirmation of suspected G. anatis isolates showed that, out of the three layer chicken farms, two farms were confirmed to have G. anatis (Table 2). The farms confirmed to have G. anatis experienced a decrease in egg production ranging from 5% to 20%. G. anatis bacteria were identified in chicken farms in the same area, specifically in the Cibinong region. On the broiler chicken farm, out of the two farms sampled, one farm was confirmed to have G. anatis, with a positivity rate of 75% in chickens experiencing respiratory disturbances ranging from moderate to severe.
3.2. Presence Virulence Genes of G. anatis
Virulence gene analysis was conducted targeting three genes: gtxA, gyrB, and flfA. Among the 14 confirmed G. anatis isolates, a positive result for gtxA was detected in 93.3% (13/14), for gyrB in 100% (14/14), and for flfA in 71.4% (10/14) of the isolates (Table 3). Three isolates were negative for the flfA gene, originating from Sukabumi (broiler, breeder, and layer chicken) and Bogor (layer chicken). Only one isolate (002/BELY/CS–BG/2022) tested negative for gtxA. Analysis of the G. anatis virulence gene combinations showed that each isolate carried at least two virulence genes (28.57%), while the highest prevalence (71.42%) was observed in isolates harboring all three virulence genes (Figure 2).
Distribution patterns of Gallibacterium anatis virulence genes. Each circle is representatives of the virulence gene, such as gtxA, flfA, and gyrB.
3.3. Similarity and Phylogenetic Tree of G. anatis
Similarity analysis using BLAST revealed that the 16S rRNA gene sequence of the isolates were highly similar to G. anatis. Similarity values greater than 98.65% indicate that a bacterial isolate belongs to the same species, while values above 95% can identify the genus [17]. In this study, all nucleotide sequences showed a similarity exceeding 98.65% with G. anatis (Table 4).
The bacterial isolates analyzed here shared similarities with strains from various countries. Specifically, strains YU‐ZMD, TH22, YJ922, DFS02, and BJF12 originated from chickens in China; ESV200 from Mexico; UMN179 from the United States; and IMT49310 from Germany.
Confirmation of the taxonomic proximity of a bacterial isolate by using similarity values can be reinforced by constructing a phylogenetic tree. The phylogenetic tree showed that all isolates clustered within the G. anatis in‐group, forming two main supergroups, while three reference isolates from China and the United States formed a different supergroup. Outgroup species, G. salpingitidis and G. trehalosifermentans formed distinct clusters but remained within the overall Gallibacterium clade.
The phylogenetic tree also indicated that isolates 003/Breeder Layer/Cisarua–Bogor/2022 and 005/Layer/Sukabumi//2022 were more closely related, supported by a bootstrap value of 100% (Figure 3). However, other isolates showed variable relationships with both the tested and reference strains, with bootstrap values below 70%. Bootstrap values reflect the reliability of the phylogenetic tree, where values greater than 70% correspond to a confidence level exceeding 95% [18]. The highest bootstrap values in some relationship suggest that the formed tree could vary if alternative phylogenetic reconstruction methods are applied.
Phylogenetic tree of G. anatis isolates from Indonesia. This phylogenetic tree was constructed using neighbor‐joining with the MEGA 7 application from the 16S‐23S rRNA gene. The branches of the dendrogram show bootstrap values (%) with 1000 replications, and the scale represents one per 1000 nucleotide sequence substitutions of the 16S rRNA gene. () denotes the isolates tested. () denotes outgroup reference isolates.
4. Discussion
In the present study, G. anatis was successfully isolated from poultry farms in West Java, Indonesia. Notably, the presence of G. anatis has also been identified in Europe, Africa, China, India, Japan, North America, and South America [6], underscoring its global distribution. Recent reports further confirm its widespread emergence in commercial poultry systems, particularly in Asia and Europe, where multidrug‐resistant and highly virulent strains have been documented [9]. G. anatis can cause a decrease in egg production in layer and breeder chickens, as well as respiratory disorders in broilers. In this study, G. anatis was identified in layer farms in the Cibinong area. This is consistent with the findings of Yaman and Yapicier [6], who reported G. anatis in Turkish layer farms experiencing decreased egg production. A decrease in production was also observed on Farm 1 in the Ciampea region, but G. anatis was not identified. Such production drops may be linked to other bacterial or viral pathogens that were not identified in this study. Multifactorial drops in egg production have been frequently reported in the field, as coinfections with E. coli, Mycoplasma spp., Avibacterium paragallinarum, or viral pathogens often complicate accurate attribution of production losses [19, 20].
The molecular results for G. anatis in this study indicated that not all isolates showing phenotypic suspicion of G. anatis were successfully molecularly confirmed using the 16S‐23S rRNA gene. One sample from Farms 3, 4, and 5 showed negative results based on molecular analysis. This could be due to the fact that phenotypic testing relies on biochemical pathways and carbon source utilization, which may have similarities with other bacteria, especially within the same family. Bacteria with phenotypic characteristics similar to those of G. anatis include Pseudomonas aeruginosa and Mannheimia haemolytica [21]. These results highlight the importance of combining phenotypic and molecular methods for accurate identification. This challenge aligns with recent findings showing that G. anatis displays substantial phenotypic variability, which can lead to misidentification when relying solely on biochemical assays [22].
The virulence genes of G. anatis have previously been identified. The flfA gene encodes the fimbriae in G. anatis and plays a role in the adhesion process, especially in the oropharyngeal epithelial cells of chickens, which have three fimbrial gene clusters [23, 24]. The presence of flfA can be associated with tissue tropism [25], but the data in this study are not sufficient to establish this correlation. The upper respiratory tract, particularly the trachea, is the common site of G. anatis colonization [2]. However, G. anatis has been detected in internal organs such as the liver, heart, and lungs [14, 15]. These findings suggest that certain strains have the potential for extra‐respiratory dissemination, although the mechanisms underlying this spread remain to be fully elucidated.
The gtxA gene encodes a toxin that can cause inflammation and is responsible for hemolytic activity [26]. G. anatis bacteria are grouped into two biotypes: hemolytic and nonhemolytic. The presence of the gtxA gene can be used to classify isolates as belonging to the hemolytic biotype. This classification can also be observed through phenotypic testing, which results in hemolytic zones on blood agar media. In this study, Isolate 002/BELY/CS‐BG/2022 exhibited a hemolytic zone but tested negative for the gtxA gene. This is likely due to the low concentration of the gtxA gene in the medium during the late log phase of the bacterial growth cycle [27], making it undetectable in molecular testing. Additionally, other isolates that displayed hemolytic zones on blood agar correlated with the presence of the gtxA gene in this test. Similar discrepancies have been reported in recent studies, where gtxA expression was shown to be variable based on the growth phase and environmental conditions, leading to inconsistent molecular detection despite clear phenotypic hemolysis [11].
All isolated samples tested positive for encoding the gyrB, which is associated with virulence. The presence of gyrB in these isolates is significant because the gene encodes the ATPase domain of gyrase, which is essential for bacterial DNA replication [28]. The gyrB gene can be used as a discriminatory marker for identifying G. anatis and differentiating closely related Pasteurellaceae species [22, 29]. The presence of these virulence genes confirms the pathogenic potential of the isolates collected in this study.
Similarity analysis showed that Isolates 016/LY/CB–BG/2023 and 017/LY/CB–BG/2023 originated from the same type of chicken and area; however, they exhibited different reference bacterial strains based on similarity values. Isolate 014/BR/CB–BG/2023, the only broiler chicken tested in this study, showed similar testing results that can also be found in isolates originating from layer chickens or breeders. Interestingly, Isolate 017/LY/CB‐BG/2023, from layer chickens, showed high similarity to isolates from different host species, including cattle. This suggests that the 16S–23S rRNA gene is highly conserved and may not reflect host‐specific adaptation, indicating that sequence similarity does not necessarily correspond to the original host of the isolate. This is consistent with recent genomic studies showing that G. anatis forms host‐independent lineages and that cross‐species similarities are common, especially between poultry and ruminant isolates [9].
Phylogenetic analysis of G. anatis has been conducted extensively to classify this bacterium. Partial use of the 16S‐23S rRNA gene was unable to differentiate isolates based on their origin or epidemiology. Phylogenetic analysis using the 16S‐23S rRNA gene can only group isolates within the same species but not necessarily within the same group with good bootstrap values. In addition to using this gene, the flfA gene has also been utilized for bacterial analysis; however, it also fails to differentiate this gene based on sample origin. Current evidence strongly supports the use of multilocus sequence typing (MLST), core‐genome SNP analysis, or whole‐genome sequencing (WGS) to obtain clearer cluster separation and better infer transmission routes and virulence‐associated lineages [7].
These findings demonstrate that G. anatis is present in West Java poultry farms, with isolates carrying virulence genes that may contribute to decreased egg production and respiratory disorders. Future studies should include larger sample sizes, additional virulence or housekeeping genes, and WGS to better understand the epidemiology, host adaptation mechanisms, and pathogenicity of G. anatis in Indonesian poultry systems.
5. Conclusion
This is the first report to publish findings regarding the presence of G. anatis isolates in Indonesia. A total of 17 isolates were phenotypically identified as G. anatis. Molecular analysis confirmed that 14 of these isolates were G. anatis, and the virulence genes gtxA, gyrB, and flfA were detected, showing diverse patterns. Nucleotide analysis results indicated that the isolates had similarities to G. anatis with various strains. However, the G. anatis strain in this study could not be classified into a specific group in the phylogenetic tree constructed using the 16S‐23S rRNA gene sequence. The presence of G. anatis in chickens experiencing respiratory disorders or decreased production is similar to that of G. anatis in several countries, confirming the existence of this bacterium.
Funding
The publication of this journal was granted by the United States Agency for International Development (USAID) throughout the SEAOHUN One Health Scholarship Program. The contents are the responsibility of the authors and do not necessarily reflect the views of USAID or the United States Government.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Singh B. R. , Singh S. V. , Palanivelu M. , Kumar M. A. , Sinha D. K. , and Kumar O. R. V. , Gallibacterium anatis Outbreaks in Domestic Birds in North India: Antimicrobial and Herbal Drug Sensitivity of Avibacterium and Gallibacterium Isolates, IJPS. (2018) 53, no. 2, 10.5958/0974-8180.2018.00043.0. · doi ↗
- 2Elbestawy A. R. , Ellakany H. F. , Abd El-Hamid H. S. et al., Isolation, Characterization, and Antibiotic Sensitivity Assessment of Gallibacterium anatis Biovar Haemolytica, From Diseased Egyptian Chicken Flocks During the Years 2013 and 2015, Poul Sci. (2018) 97, 1519–1525, 10.3382/ps/pey 007, 2-s 2.0-85047221591.29471426 · doi ↗ · pubmed ↗
- 3Neubauer C. , De Souza-Pilz M. , Bojesen A. M. , Bisgaard M. , and Hess M. , Tissue Distribution of Haemolytic Gallibacterium anatis Isolates in Laying Birds with Reproductive Disorders, Avian Pathology. (2009) 38, 1–7, 10.1080/03079450802577848, 2-s 2.0-60549088915.19089694 · doi ↗ · pubmed ↗
- 4Bojesen A. M. , Christensen H. , Nielsen O. L. , Olsen J. E. , and Bisgaard M. , Detection of Gallibacterium spp. in Chickens by Fluorescent 16S Rrna in Situ Hybridization, Journal of Clinical Microbiology. (2003) 41, no. 11, 5167–5172, 10.1128/JCM.41.11.5167-5172.2003, 2-s 2.0-0242425745.14605154 PMC 262499 · doi ↗ · pubmed ↗
- 5Nassik S. , Tallouzt S. , Karbach N. et al., First Report of Isolation of Gallibacterium anatis From Layer Chickens in Morocco with Decrease in Laying Performance, Avian Diseases. (2019) 63, no. 4, 10.1637/aviandiseases-D-19-00119.31865689 · doi ↗ · pubmed ↗
- 6Yaman S. and Sahan Yapicier O. , Diagnosis of Gallibacterium anatis in Layers: First Report in Turkey, Brazilian Journal of Poultry Science. (2019) 21, no. 3, 10.1590/1806-9061-2019-1019. · doi ↗
- 7Guo F. , Wang D. , Wu H. et al., Genomic Characteristics, Antimicrobial Resistance Profiles and Virulence Factors of Gallibacterium anatis Isolates from Layer Chickens in Northern China, BMC Microbiology. (2025) 25, no. 1, 10.1186/s 12866-025-04440-3.PMC 1258136241184760 · doi ↗ · pubmed ↗
- 8Palmieri N. , Hess C. , and Hess M. , GWAS and Comparative Genomics Reveal Candidate Antibiotic Resistance Genes in the Avian Pathogen Gallibacterium anatis for Six Widespread Antibiotics, Veterinary Microbiology. (2024) 290, 10.1016/j.vetmic.2024.109995.38301451 · doi ↗ · pubmed ↗